
Transcriptional Regulator DasR Represses Daptomycin Production through Both Direct and Cascade Mechanisms in Streptomyces roseosporus



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
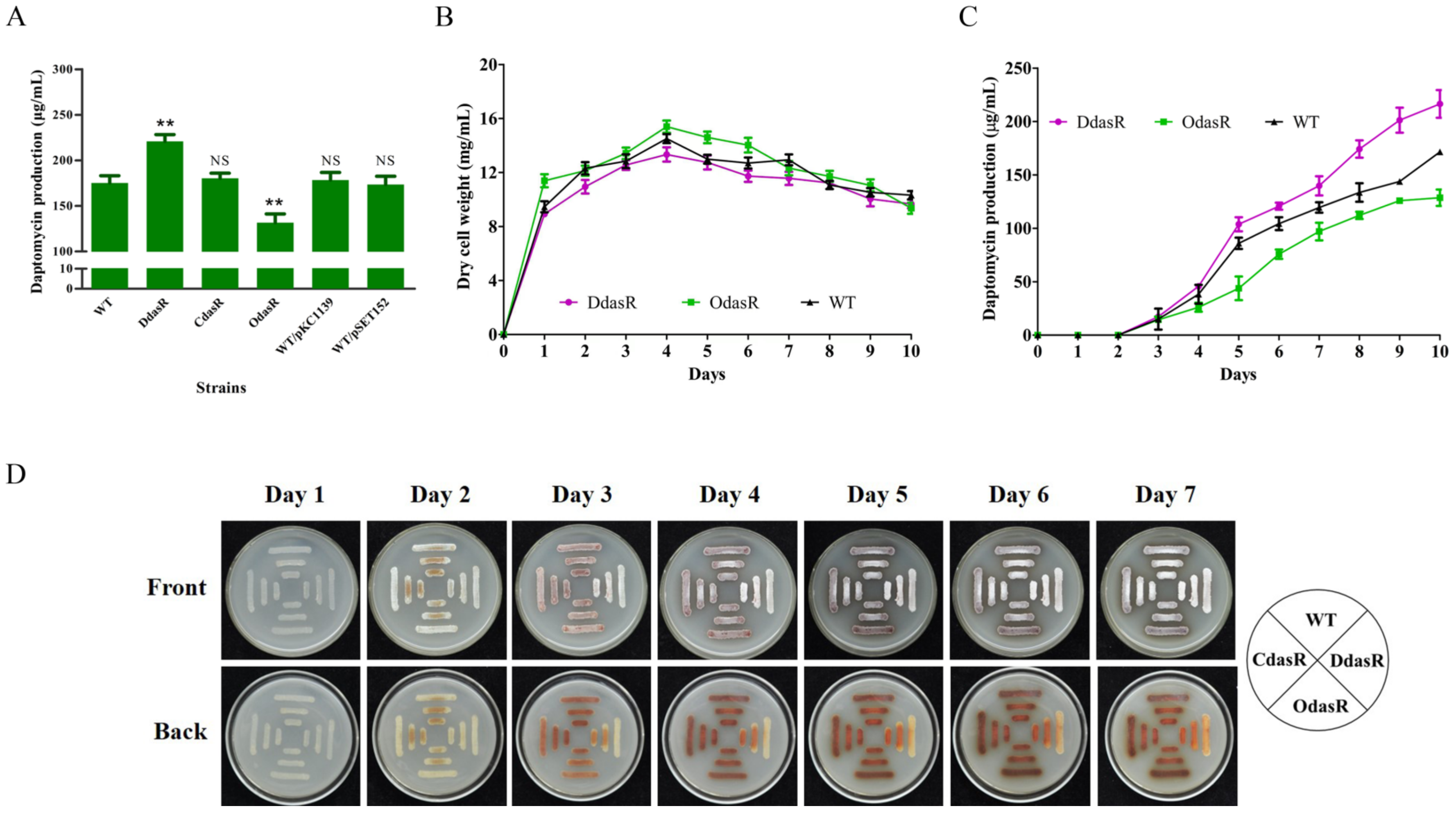
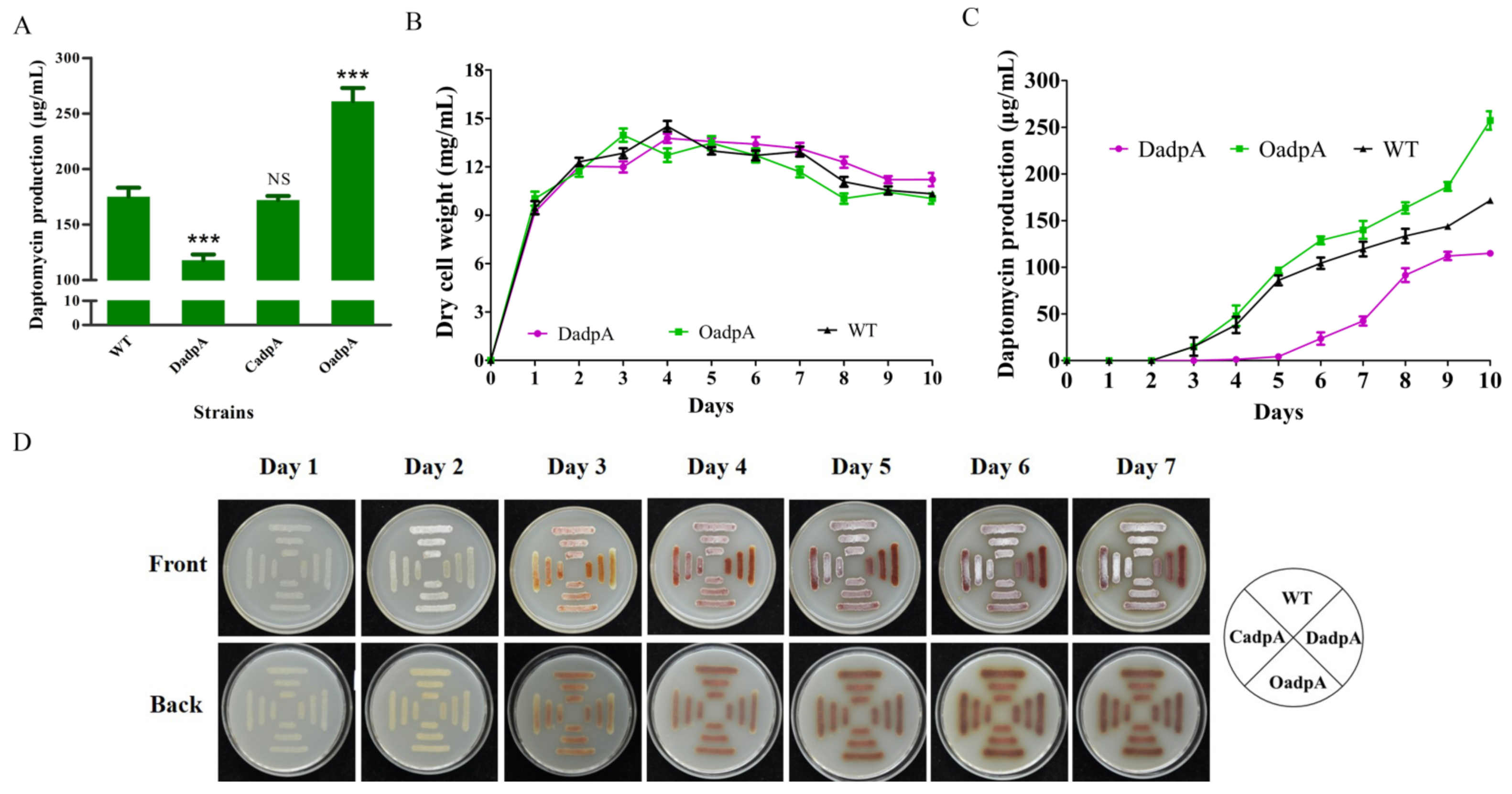
2.1. DasR Inhibits Daptomycin Production and Morphological Development, but Promotes Pigment Production
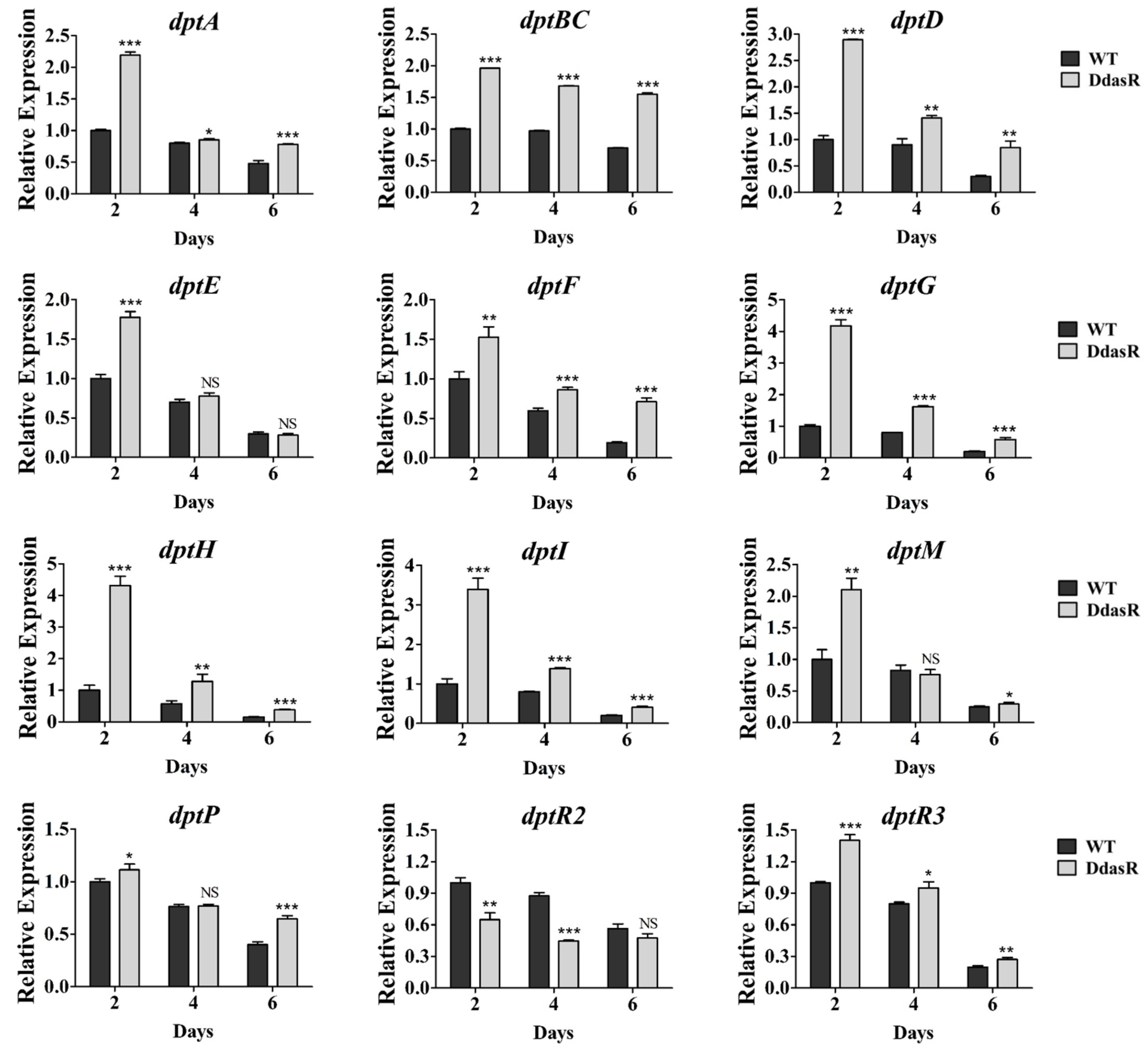
2.2. DasR Regulates the Transcription of dpt Genes
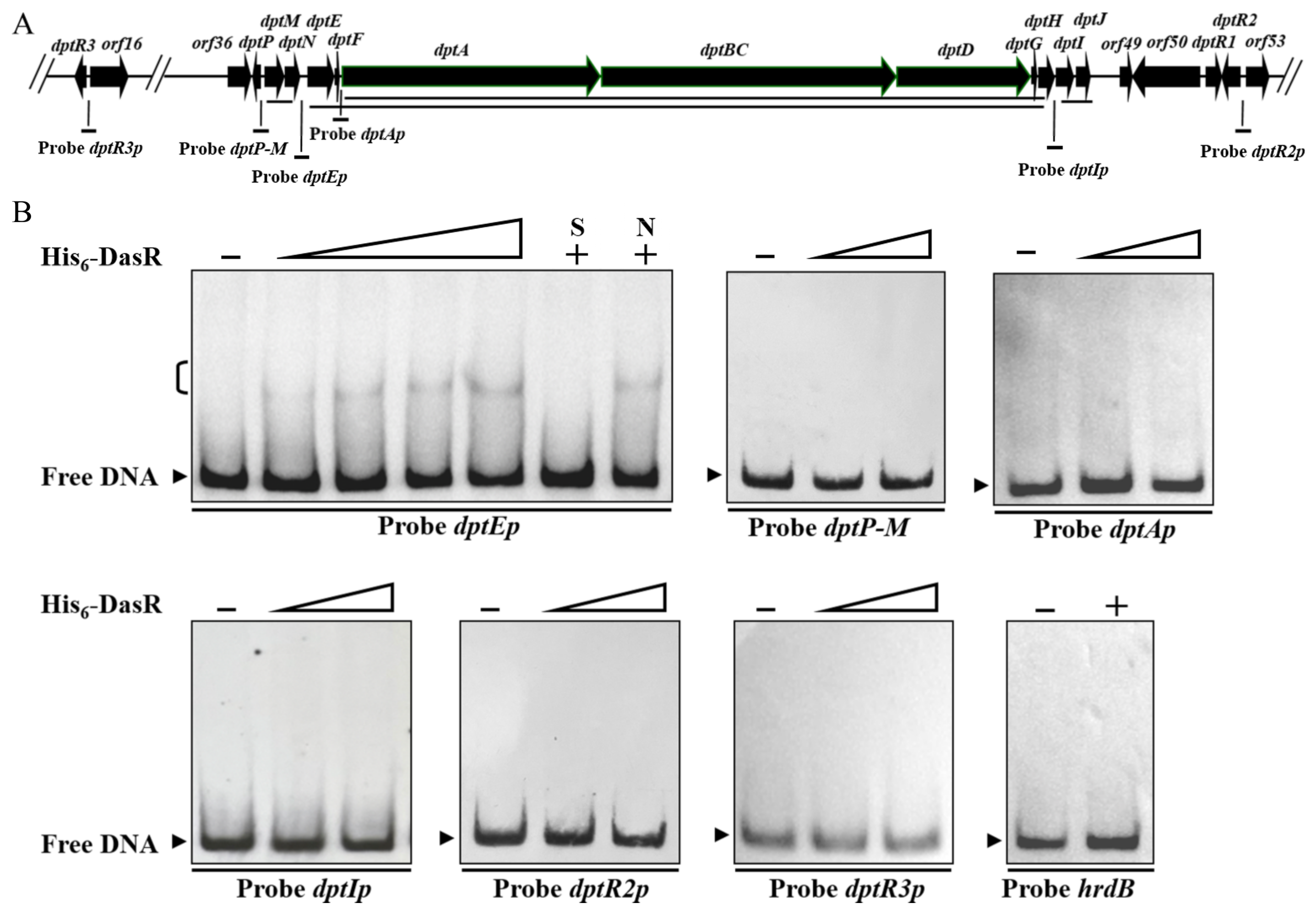
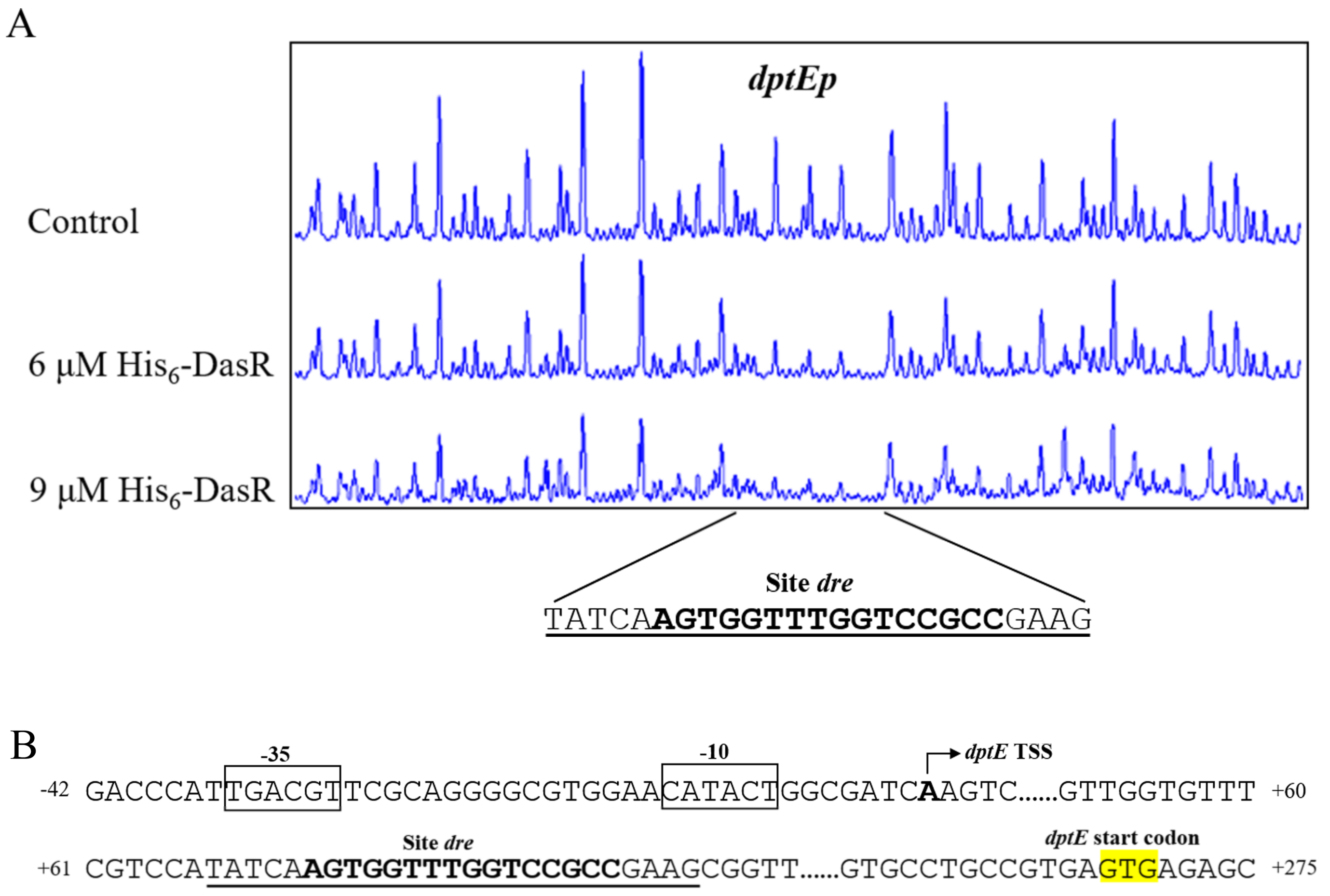
2.3. DasR Binds Specifically to the dptE-Promoter Region
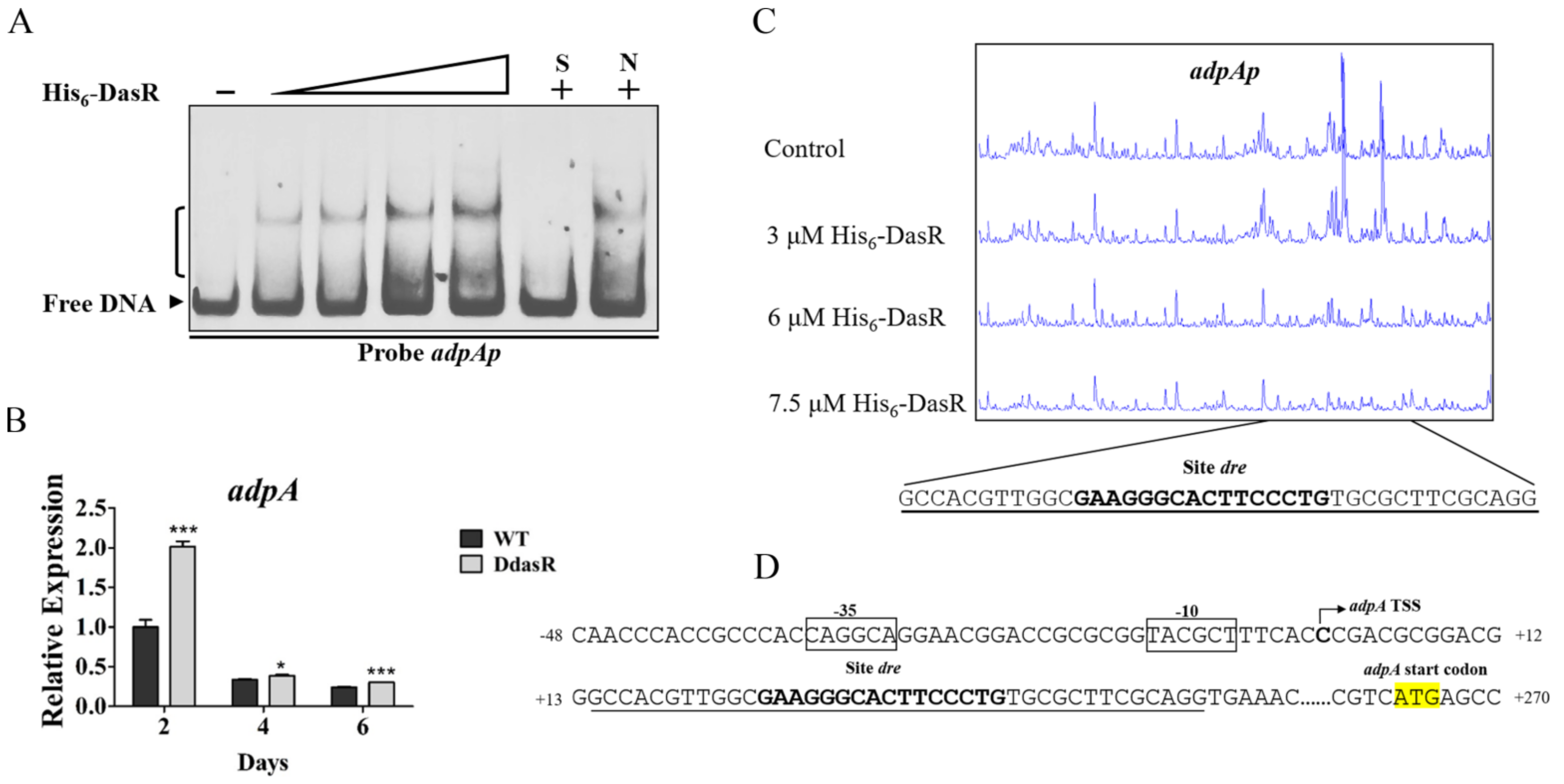
2.4. DasR Directly Represses adpA Involved in Daptomycin Production and Morphological Development
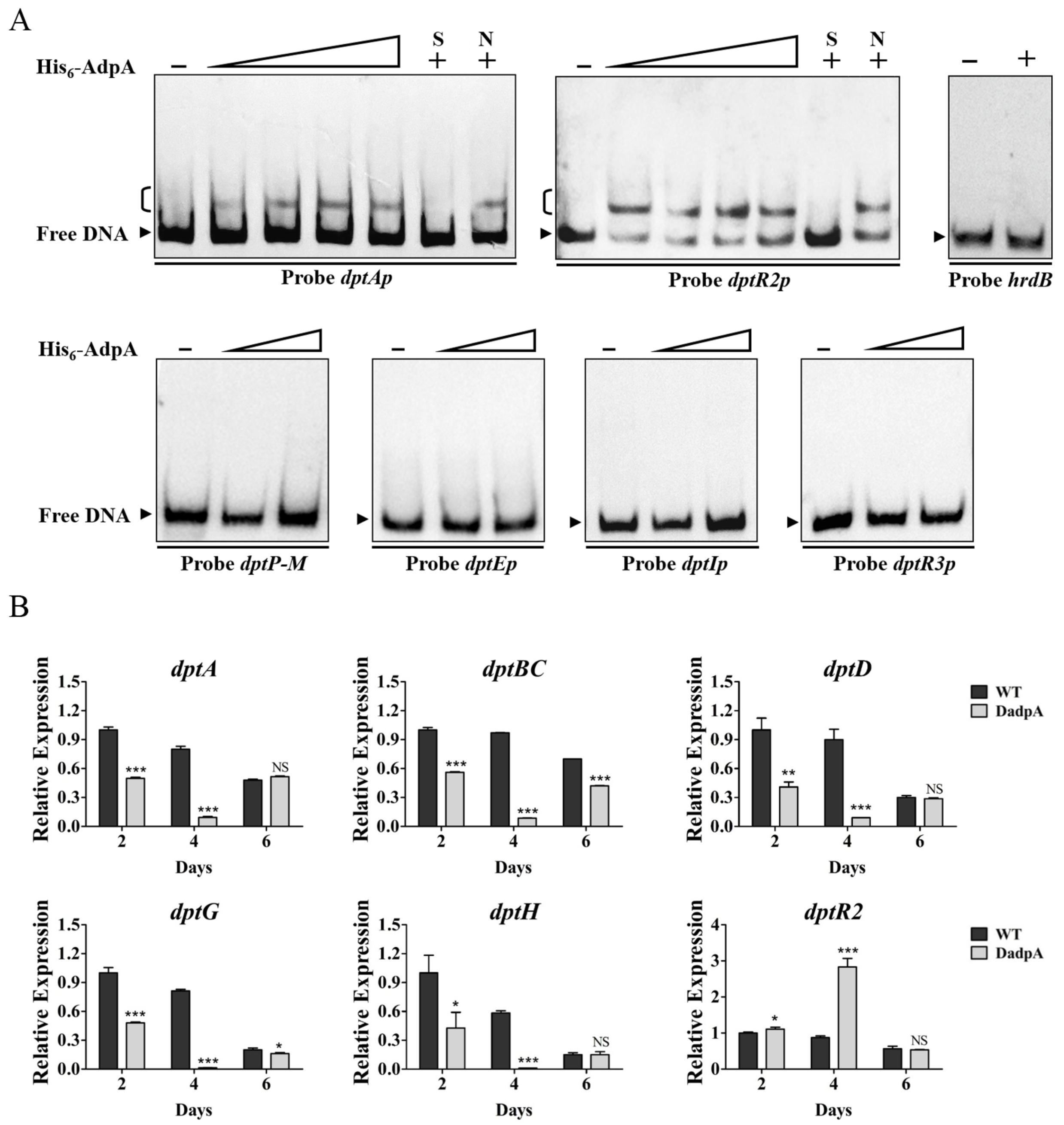
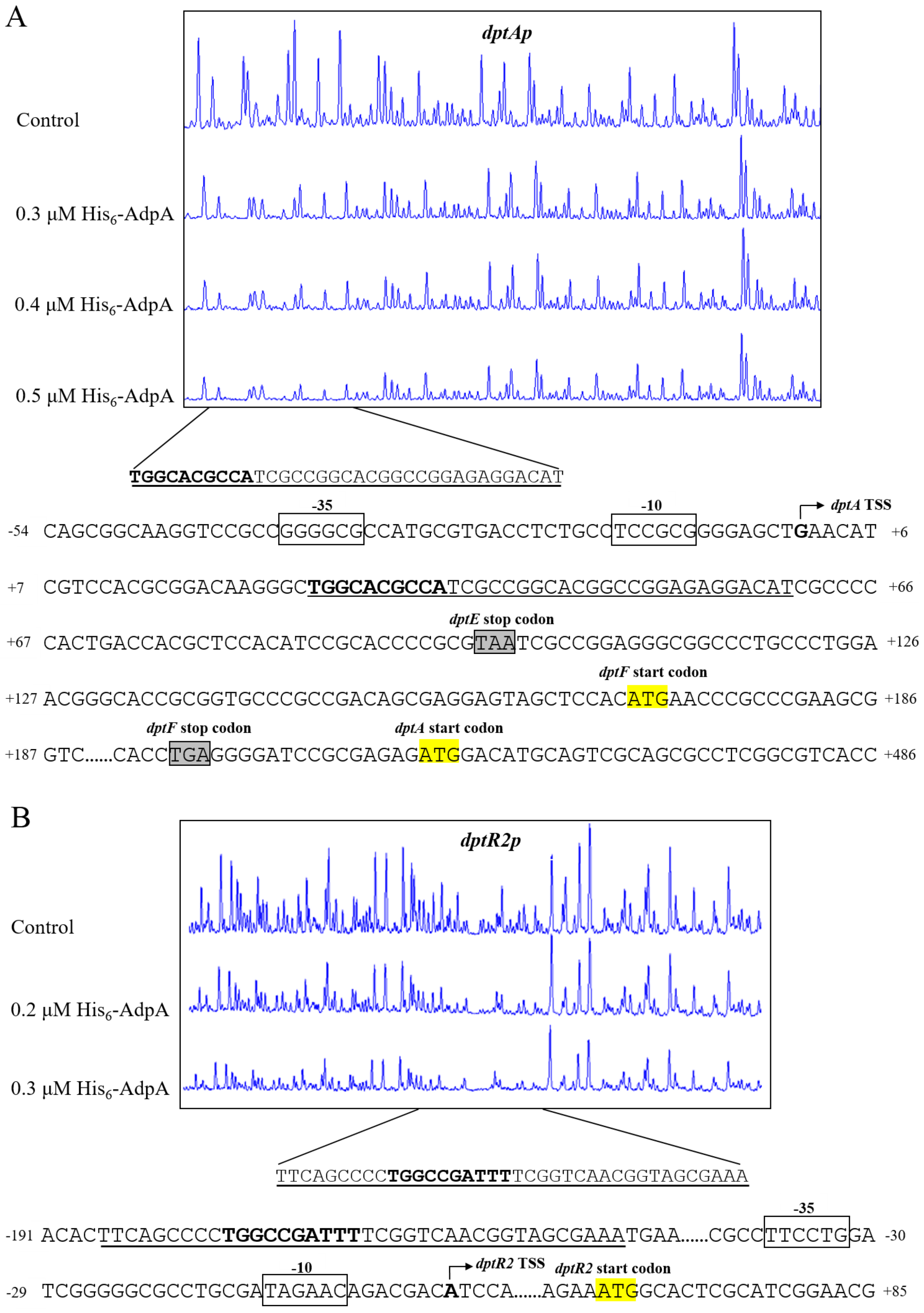
2.5. AdpA Directly Regulates Expression of dptA Operon and dtpR2
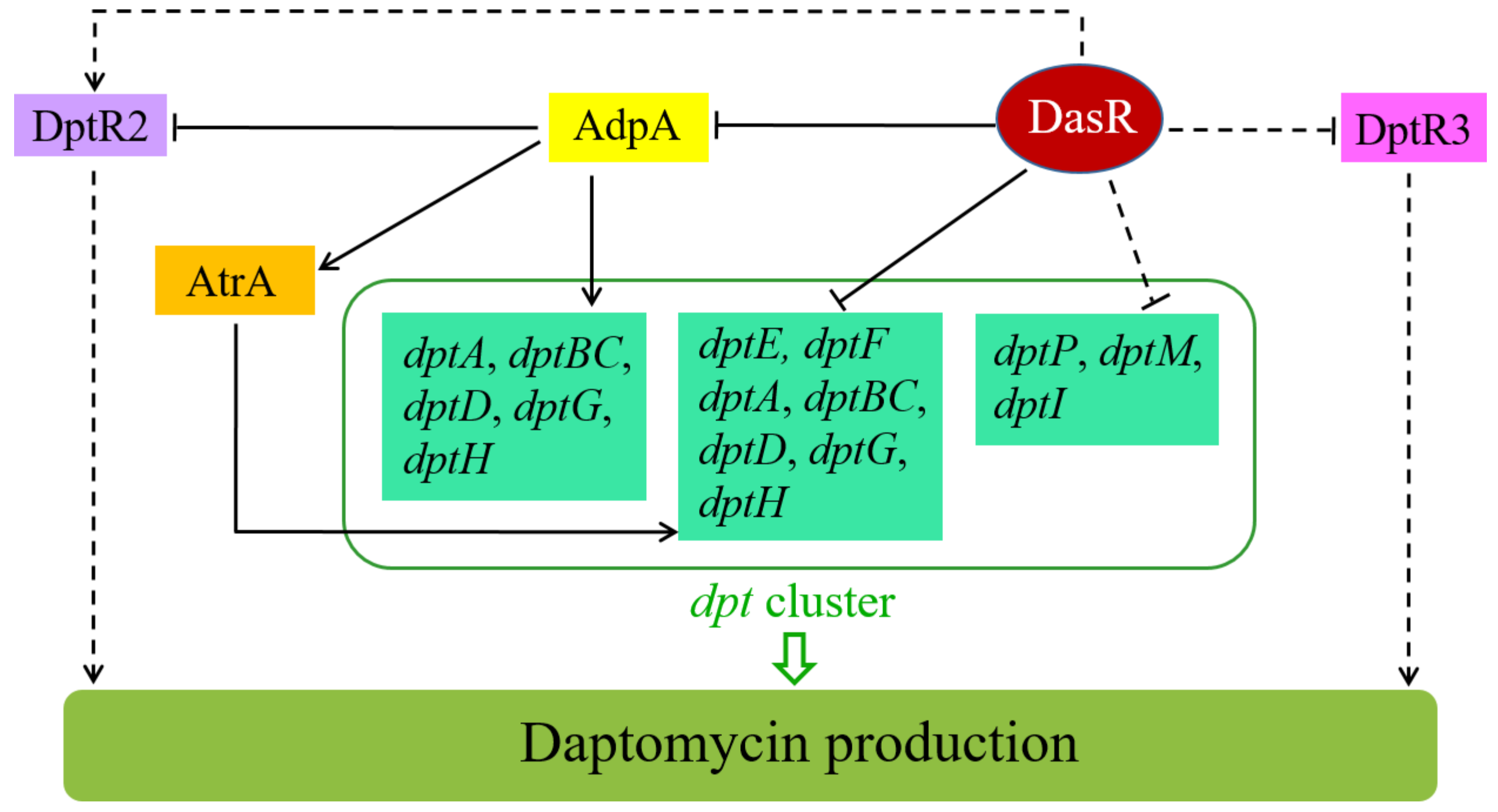
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, Primer Pair, and Growth Conditions
4.2. Construction of S. roseosporus Mutant Strains
4.3. Production and Analysis of Daptomycin
4.4. Reverse Transcription and Quantitative Real-Time PCR (RT-qPCR) Analysis
4.5. Overexpression and Purification of His6-DasR and His6-AdpA
4.6. EMSAs
4.7. DNase I Footprinting Assay
4.8. 5′ Rapid Amplification of cDNA Ends (5′ RACE)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Urem, M.; Swiatek-Polatynska, M.A.; Rigali, S.; van Wezel, G.P. Intertwining nutrient-sensory networks and the control of antibiotic production in Streptomyces. Mol. Microbiol. 2016, 102, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Van Wezel, G.P.; McDowall, K.J. The regulation of the secondary metabolism of Streptomyces: New links and experimental advances. Nat. Prod. Rep. 2011, 28, 1311–1333. [Google Scholar] [CrossRef] [PubMed]
- Zuttion, F.; Colom, A.; Matile, S.; Farago, D.; Pompeo, F.; Kokavecz, J.; Galinier, A.; Sturgis, J.; Casuso, I. High-speed atomic force microscopy highlights new molecular mechanism of daptomycin action. Nat. Commun. 2020, 11, 6312. [Google Scholar] [CrossRef]
- Gonzalez-Ruiz, A.; Seaton, R.A.; Hamed, K. Daptomycin: An evidence-based review of its role in the treatment of Gram-positive infections. Infect. Drug Resist. 2016, 9, 47–58. [Google Scholar]
- Debono, M.; Abbott, B.J.; Molloy, R.M.; Fukuda, D.S.; Hunt, A.H.; Daupert, V.M.; Counter, F.T.; Ott, J.L.; Carrell, C.B.; Howard, L.C.; et al. Enzymatic and chemical modifications of lipopeptide antibiotic A21978C: The synthesis and evaluation of daptomycin (LY146032). J. Antibiot. 1988, 41, 1093–1105. [Google Scholar] [CrossRef]
- Robbel, L.; Marahiel, M.A. Daptomycin, a bacterial lipopeptide synthesized by a nonribosomal machinery. J. Biol. Chem. 2010, 285, 27501–27508. [Google Scholar] [CrossRef]
- Miao, V.; Coeffet-Legal, M.F.; Brian, P.; Brost, R.; Penn, J.; Whiting, A.; Martin, S.; Ford, R.; Parr, I.; Bouchard, M.; et al. Daptomycin biosynthesis in Streptomyces roseosporus: Cloning and analysis of the gene cluster and revision of peptide stereochemistry. Microbiology 2005, 151, 1507–1523. [Google Scholar] [CrossRef]
- Wittmann, M.; Linne, U.; Pohlmann, V.; Marahiel, M.A. Role of DptE and DptF in the lipidation reaction of daptomycin. FEBS J. 2008, 275, 5343–5354. [Google Scholar] [CrossRef]
- Yuan, P.; Zhou, R.; Chen, X.; Luo, S.; Wang, F.; Mao, X.; Li, Y. DepR1, a TetR family transcriptional regulator, positively regulates daptomycin production in an industrial producer, Streptomyces roseosporus SW0702. Appl. Environ. Microbiol. 2016, 82, 1898–1905. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, Q.; Zhuang, S.; Chen, Z.; Wen, Y.; Li, J. A MarR family transcriptional regulator, DptR3, activates daptomycin biosynthesis and morphological differentiation in Streptomyces roseosporus. Appl. Environ. Microbiol. 2015, 81, 3753–3765. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Hui, M.; Li, R.; Zhang, S. Pleiotropic regulation of daptomycin synthesis by DptR1, a LuxR family transcriptional regulator. World J. Microbiol. Biotechnol. 2020, 36, 173. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ren, N.; Luo, S.; Chen, X.; Mao, X.; Li, Y. DptR2, a DeoR type auto-regulator, is required for daptomycin production in Streptomyces roseosporus. Gene 2014, 544, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Luo, S.; Zhou, R.; Wang, F.; Yu, P.; Sun, N.; Chen, X.; Tang, Y.; Li, Y. Transcriptional regulation of the daptomycin gene cluster in Streptomyces roseosporus by an autoregulator, AtrA. J. Biol. Chem. 2015, 290, 7992–8001. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Luo, S.; Li, Y. Negative regulation of daptomycin production by DepR2, an ArsR-family transcriptional factor. J. Ind. Microbiol. Biotechnol. 2017, 44, 1653–1658. [Google Scholar] [CrossRef]
- Luo, S.; Chen, X.; Mao, X.; Li, Y. Transposon-based identification of a negative regulator for the antibiotic hyper-production in Streptomyces. Appl. Microbiol. Biotechnol. 2018, 102, 6581–6592. [Google Scholar] [CrossRef]
- Zheng, Y.; Sun, C.; Fu, Y.; Chen, X.; Li, Y.; Mao, X. Dual regulation between the two-component system PhoRP and AdpA regulates antibiotic production in Streptomyces. J. Ind. Microbiol. Biotechnol. 2019, 46, 725–737. [Google Scholar] [CrossRef]
- Huang, X.; Ma, T.; Tian, J.; Shen, L.; Zuo, H.; Hu, C.; Liao, G. wblA, a pleiotropic regulatory gene modulating morphogenesis and daptomycin production in Streptomyces roseosporus. J. Appl. Microbiol. 2017, 123, 669–677. [Google Scholar] [CrossRef]
- Wu, J.; Chen, D.; Wu, J.; Chu, X.; Yang, Y.; Fang, L.; Zhang, W. Comparative transcriptome analysis demonstrates the positive effect of the cyclic AMP receptor protein Crp on daptomycin biosynthesis in Streptomyces roseosporus. Front. Bioeng. Biotechnol. 2021, 9, 618029. [Google Scholar] [CrossRef]
- Yan, H.; Lu, X.; Sun, D.; Zhuang, S.; Chen, Q.; Chen, Z.; Li, J.; Wen, Y. BldD, a master developmental repressor, activates antibiotic production in two Streptomyces species. Mol. Microbiol. 2020, 113, 123–142. [Google Scholar] [CrossRef]
- Colson, S.; Stephan, J.; Hertrich, T.; Saito, A.; van Wezel, G.P.; Titgemeyer, F.; Rigali, S. Conserved cis-acting elements upstream of genes composing the chitinolytic system of streptomycetes are DasR-responsive elements. J. Mol. Microbiol. Biotechnol. 2007, 12, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Rigali, S.; Nothaft, H.; Noens, E.E.; Schlicht, M.; Colson, S.; Muller, M.; Joris, B.; Koerten, H.K.; Hopwood, D.A.; Titgemeyer, F.; et al. The sugar phosphotransferase system of Streptomyces coelicolor is regulated by the GntR-family regulator DasR and links N-acetylglucosamine metabolism to the control of development. Mol. Microbiol. 2006, 61, 1237–1251. [Google Scholar] [CrossRef]
- Rigali, S.; Titgemeyer, F.; Barends, S.; Mulder, S.; Thomae, A.W.; Hopwood, D.A.; van Wezel, G.P. Feast or famine: The global regulator DasR links nutrient stress to antibiotic production by Streptomyces. EMBO Rep. 2008, 9, 670–675. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lin, C.; Li, X.; Tang, Z.; Qiao, J.; Zhao, G. DasR positively controls monensin production at two-level regulation in Streptomyces cinnamonensis. J. Ind. Microbiol. Biotechnol. 2016, 43, 1681–1692. [Google Scholar] [CrossRef]
- Liao, C.; Xu, Y.; Rigali, S.; Ye, B. DasR is a pleiotropic regulator required for antibiotic production, pigment biosynthesis, and morphological development in Saccharopolyspora erythraea. Appl. Microbiol. Biotechnol. 2015, 99, 10215–10224. [Google Scholar] [CrossRef]
- Ng, I.S.; Ye, C.; Zhang, Z.; Lu, Y.; Jing, K. Daptomycin antibiotic production processes in fed-batch fermentation by Streptomyc-es roseosporus NRRL11379 with precursor effect and medium optimization. Bioprocess Biosyst. Eng. 2014, 37, 415–423. [Google Scholar] [CrossRef]
- Coeffet-Le Gal, M.F.; Thurston, L.; Rich, P.; Miao, V.; Baltz, R.H. Complementation of daptomycin dptA and dptD deletion mutations in trans and production of hybrid lipopeptide antibiotics. Microbiology 2006, 152, 2993–3001. [Google Scholar] [PubMed]
- Higo, A.; Hara, H.; Horinouchi, S.; Ohnishi, Y. Genome-wide distribution of AdpA, a global regulator for secondary metabolism and morphological differentiation in Streptomyces, revealed the extent and complexity of the AdpA regulatory network. DNA Res. 2012, 19, 259–273. [Google Scholar] [CrossRef]
- Zhu, J.; Sun, D.; Liu, W.; Chen, Z.; Li, J.; Wen, Y. AvaR2, a pseudo gamma-butyrolactone receptor homologue from Streptomyces avermitilis, is a pleiotropic repressor of avermectin and avenolide biosynthesis and cell growth. Mol. Microbiol. 2016, 102, 562–578. [Google Scholar] [CrossRef]
- Ohnishi, Y.; Yamazaki, H.; Kato, J.Y.; Tomono, A.; Horinouchi, S. AdpA, a central transcriptional regulator in the A-factor regulatory cascade that leads to morphological development and secondary metabolism in Streptomyces griseus. Biosci. Biotechnol. Biochem. 2005, 69, 431–439. [Google Scholar] [CrossRef]
- Wolanski, M.; Donczew, R.; Kois-Ostrowska, A.; Masiewicz, P.; Jakimowicz, D.; Zakrzewska-Czerwinska, J. The level of AdpA directly affects expression of developmental genes in Streptomyces coelicolor. J. Bacteriol. 2011, 193, 6358–6365. [Google Scholar] [CrossRef] [PubMed]
- Den Hengst, C.D.; Tran, N.T.; Bibb, M.J.; Chandra, G.; Leskiw, B.K.; Buttner, M.J. Genes essential for morphological development and antibiotic production in Streptomyces coelicolor are targets of BldD during vegetative growth. Mol. Microbiol. 2010, 78, 361–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ji, J.; Li, X.; Wang, J.; Li, S.; Pan, G.; Fan, K.; Yang, K. Angucyclines as signals modulate the behaviors of Streptomyces coelicolor. Proc. Natl. Acad. Sci. USA 2014, 111, 5688–5693. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Liu, G.; Yang, H.; Tian, Y.; Tan, H. The pleiotropic regulator AdpA-L directly controls the pathway-specific activator of nikkomycin biosynthesis in Streptomyces ansochromogenes. Mol. Microbiol. 2009, 72, 710–723. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, J.; Zhuo, J.; Li, Y.; Tian, Y.; Tan, H. Activation and mechanism of a cryptic oviedomycin gene cluster via the disruption of a global regulatory gene, adpA, in Streptomyces ansochromogenes. J. Biol. Chem. 2017, 292, 19708–19720. [Google Scholar] [CrossRef]
- Bu, X.; Weng, J.; He, B.; Xu, M.; Xu, J. A novel AdpA homologue negatively regulates morphological differentiation in Streptomyces xiamenensis 318. Appl. Environ. Microbiol. 2019, 85, e03107–e03118. [Google Scholar] [CrossRef]
- Tschowri, N.; Schumacher, M.A.; Schlimpert, S.; Chinnam, N.B.; Findlay, K.C.; Brennan, R.G.; Buttner, M.J. Tetrameric c-di-GMP mediates effective transcription factor dimerization to control Streptomyces development. Cell 2014, 158, 1136–1147. [Google Scholar] [CrossRef]
- Tenconi, E.; Urem, M.; Swiatek-Połatynska, M.A.; Titgemeyer, F.; Muller, Y.A.; van Wezel, G.P.; Rigali, S. Multiple allosteric effectors control the affinity of DasR for its target sites. Biochem. Biophys. Res. Commun. 2015, 464, 324–329. [Google Scholar] [CrossRef]
- Bierman, M.; Logan, R.; O’Brien, K.; Seno, E.T.; Rao, R.N.; Schoner, B.E. Plasmid cloning vectors for the conjugal transfer of DNA from Escherichia coli to Streptomyces spp. Gene 1992, 116, 43–49. [Google Scholar] [CrossRef]
- Li, L.; Guo, J.; Wen, Y.; Chen, Z.; Song, Y.; Li, J. Overexpression of ribosome recycling factor causes increased production of avermectin in Streptomyces avermitilis strains. J. Ind. Microbiol. Biotechnol. 2010, 37, 673–679. [Google Scholar] [CrossRef]
- Luo, S.; Sun, D.; Zhu, J.; Chen, Z.; Wen, Y.; Li, J. An extracytoplasmic function sigma factor, σ(25), differentially regulates avermectin and oligomycin biosynthesis in Streptomyces avermitilis. Appl. Microbiol. Biotechnol. 2014, 98, 7097–7112. [Google Scholar] [CrossRef] [PubMed]
- Zianni, M.; Tessanne, K.; Merighi, M.; Laguna, R.; Tabita, F.R. Identification of the DNA bases of a DNase I footprint by the use of dye primer sequencing on an automated capillary DNA analysis instrument. J. Biomol. Tech. 2006, 17, 103–113. [Google Scholar] [PubMed]
- Macneil, D.J.; Klapko, L.M. Transformation of Streptomyces avermitilis by plasmid DNA. J. Ind. Microbiol. 1987, 2, 209–218. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Q.; Zhu, J.; Li, X.; Wen, Y. Transcriptional Regulator DasR Represses Daptomycin Production through Both Direct and Cascade Mechanisms in Streptomyces roseosporus. Antibiotics 2022, 11, 1065. https://doi.org/10.3390/antibiotics11081065
Chen Q, Zhu J, Li X, Wen Y. Transcriptional Regulator DasR Represses Daptomycin Production through Both Direct and Cascade Mechanisms in Streptomyces roseosporus. Antibiotics. 2022; 11(8):1065. https://doi.org/10.3390/antibiotics11081065
Chicago/Turabian StyleChen, Qiong, Jianya Zhu, Xingwang Li, and Ying Wen. 2022. "Transcriptional Regulator DasR Represses Daptomycin Production through Both Direct and Cascade Mechanisms in Streptomyces roseosporus" Antibiotics 11, no. 8: 1065. https://doi.org/10.3390/antibiotics11081065