1. Introduction
Hospital-acquired infections (HAIs), especially those from Gram-negative organisms that tend to become multidrug resistant (MDR), pose serious threats to healthcare systems [
1].
Acinetobacter baumannii is a non-spore forming, Gram-negative rod that is ubiquitous in the environment, and an opportunistic human pathogen.
A. baumannii infections commonly occur in severely ill patients with extended hospital stays. These bacteria readily acquire antibiotic resistance, resulting in difficult to treat infections which may require higher doses and longer treatment durations of more toxic antibiotics [
2,
3,
4,
5].
A. baumannii is one of the five pathogens labeled as an urgent threat by the Centers for Disease Control and Prevention (CDC), due to its ability to become resistant to multiple standard-of-care antibiotics [
6]. In fact, roughly 50% of
A. baumannii isolates are MDR [
7]. The mechanisms of resistance found in this bacterium include the presence of beta lactamases, mutation of target sites, changes in membrane permeability, and overproduction of efflux pumps [
8]. First-line treatments for
A. baumannii infections typically include cefepime, ampicillin-sulbactam, or meropenem [
7]. However, with the emergence of carbapenem-resistant pathogens, newer agents such as cefiderocol, a novel beta-lactam antibiotic that uses a siderophore moiety to bind to bacterial iron transporters, have been used [
9,
10]. Cefiderocol was shown to be as effective as imipenem-cilastatin for urinary tract infections in a clinical trial [
11], but was associated with higher mortality rates compared to other standard treatments in another study [
10]. Given this, treatment options for MDR
A. baumannii remain limited.
Most infections (greater than 90%) are nosocomial rather than community acquired [
12]. Pathogens are classified as urgent, serious, or concerning threats according to the CDC and are monitored through the Multi-site Gram-negative Surveillance Initiative [
13]. Carbapenem-resistant
Acinetobacter baumannii (CRAB) and carbapenem-resistant
Enterobacterales (CRE) are considered urgent threats, while other highly resistant organisms, such as vancomycin-resistant
Enterococci (VRE) and multidrug-resistant
Pseudomonas aeruginosa, are considered serious threats. Compared to other multidrug-resistant organisms, the number of CRAB infections tends to be lower, but still substantial. In 2017, there were 8500 hospital infections from CRAB that resulted in over 700 deaths, compared to 13,100 hospital infections with 1100 deaths from CRE in the United States. In the same year, there were 54,500 hospital infections with 5400 deaths from MDR
P. aeruginosa and 32,600 hospital infections with 2700 deaths from VRE [
14].
Globally, there were approximately 1,000,000
A. baumannii HAIs in 2019, about half of which were resistant to carbapenems [
15]. The presence of multidrug resistance varies by country. A 2019 metanalysis evaluated the prevalence of
A. baumannii hospital and ventilator associated pneumonia (HAP and VAP) and found that Central America, Cuba, Mexico, and Latin America/Caribbean had a lower rate of infections, but 100% of those infections were MDR [
16], compared to the United States that had nearly triple the amount of HAPs and VAPs with an MDR prevalence of 69% [
16]. During that same year, the prevalence of HAP and VAP infections from MDR
A. baumannii was 91% in Greece [
16], and a 4-year cohort study conducted in Greece showed that the all-cause hospital mortality rate among 91 patients with a pandrug-resistant (PDR)
A. baumannii infection was as high as 68% [
17]. Another study conducted on 92 Tunisian ICU patients showed a mortality rate up to 60% among those infected with
A. baumannii ventilator associated pneumonia [
18]. Furthermore, an analysis of 1423 U.S. patients with
A. baumannii infections from 2009 to 2013 revealed that 80% were MDR [
19]. Patients infected with MDR strains typically have more severe illness and are associated with higher frequencies of inappropriate antibiotic therapy, further perpetuating the rise of MDR isolates [
19].
MDR
A. baumannii infections are difficult to treat, which makes controlling hospital outbreaks extremely important, yet it remains a difficult task. This is largely due to
A. baumannii’s ability to survive in many different environments and under adverse conditions [
20,
21,
22,
23,
24]. They have evolved different survival mechanisms, such as resistance to desiccation [
12], which allows for survival for extended periods of time on surfaces [
4,
12]. In fact, one study revealed the presence of
A. baumannii on the bedrails of a previously infected patient up to 9 days after discharge [
12,
25]. This emphasizes the importance of proper decontamination of hospital rooms in preventing the spread of MDR bacteria. Germicides and microbicides are synonymous terms defined by the CDC as agents that destroy microorganisms [
26]. Microbicides are routinely used in the hospital environment for disinfection and sterilization to limit the spread of nosocomial infections.
A. baumannii has an innate ability to become resistant to microbicides, further contributing to its ability to survive in the harshest conditions [
27]. For example, in 283
A. baumannii ICU isolates, the minimum inhibitory concentrations (MICs) increased after treatment with benzalkonium chloride (BAC) and chlorhexidine gluconate (CHG). In addition, the minimum bactericidal concentrations (MBCs) for drug-resistant isolates were also generally higher [
27]. Biofilm formation enhances microbicide resistance.
A. baumannii biofilms are more resistant than planktonic bacteria to BAC and CHG on ceramic surfaces [
28]. Compounding the problem further,
Acinetobacter strains that form biofilms are frequently multidrug resistant [
29]. As biofilms are also inherently more drug-tolerant, some newer technologies are being explored as alternative therapeutics, including silver nano-particles [
30]. In a promising study, nanoparticles were shown to reduce biofilm formation and downregulate the expression of biofilm producing genes among
A. baumannii wound isolates [
31]. However, evaluation of safety and efficacy is still underway [
30].
Acinetobacter biofilms remain challenging to eradicate from surfaces and such infections are still difficult to treat.
The mechanism of action for each antiseptic agent varies. The mechanism of action for ethanol, CHG, and quaternary ammonium compounds (QACs) is the degradation of cell membranes [
32], resulting in cell lysis and further leading to the degradation of cellular proteins and enzymes [
33]. Bleach is a potent oxidizer and disrupts cellular activities, such as protein synthesis. The skin antiseptic povidone rapidly degrades proteins [
32]. Resistance may emerge when an agent is used at a sub-lethal concentration, or for an inappropriate amount of time [
34]. The overproduction of certain efflux pumps is a mechanism of microbicide resistance employed by bacterial pathogens [
33]. The QAC-specific efflux pumps are members of the small multidrug-resistance (SMR) family [
35], and many of the mechanisms that lead to antibiotic resistance, coincide with the mechanisms of microbicide resistance [
5,
34,
36]. For example, the
qacE efflux pump gene is commonly found on integrons with the
sul1 gene, which confers resistance to sulfonamides [
35]. Furthermore, the presence of
qacEΔ1 and
qacE has been correlated with the presence of certain beta-lactamase genes (
blaVIM and
blaNDM-1) and has additionally been associated with tolerance to CHG [
5]. CepA is an efflux pump that increases the MICs of
Klebsiella pneumoniae after exposure to CHG [
32,
37]. While the
qacA gene is frequently seen among Gram-positive pathogens [
38], a recent study of 44
A. baumannii isolates in Saudi Arabia revealed that 16.7% of strains contained the
qacA gene [
39].
The role that microbicide-resistance genes play in hospital outbreaks remains controversial. One study reported that the
qacE and
qacEΔ1 genes were identified among 100 MDR
A. baumannii isolates from an ICU, however no evidence of microbicide tolerance was identified [
40]. In agreement, a different study found no differences in bacterial survival among 20 MDR strains and 20 sporadic pan-susceptible strains after exposure to antiseptics [
41]. Furthermore, there was no significant correlation between the presence of
cepA,
qacE, and
qacEΔ1 genes and an increased MIC among 44 MDR
A. baumannii,
Pseudomonas aeruginosa, and
K. pneumoniae strains [
39]. However, another study found that
A. baumannii strains that were resistant to carbapenems, fluoroquinolones, aminoglycosides, and tetracyclines were associated with microbicide tolerance [
42].
Hospitals routinely use a variety of disinfecting agents, such as QACs, bleach, and ethanol. These are usually in the form of hospital disinfectant wipes and are used to clean surfaces and medical equipment. Antiseptics such as 10% povidone and 2% CHG are also commonly used for hand hygiene. For antiseptics to be effective against bacteria they must be used at specific contact or dwell times and meet certain Food and Drug Administration (FDA) standards. In general, this requires the germicide to kill 99.9% of bacteria on a hard, non-porous surface after a maximum time of 5 min [
43]. Different disinfectant wipes require different dwell times. Sani-cloth
® wipes containing bleach are considered bactericidal, viricidal, and fungicidal when a dwell time of 4 min is used. However, the efficacy against certain pathogens varies based on dwell time [
44]. At 1 min, it is effective against
A. baumannii isolates that are resistant to cefazolin, ampicillin, gentamicin, piperacillin, trimethoprim-sulfamethoxazole, and intermediately resistant to cefotaxime and ceftriaxone, but not necessarily carbapenem-resistant
A. baumannii [
44]. Furthermore, while it is effective against
A. baumannii after 1 min, it is not effective against all pathogens until after 4 min of disinfection [
44]. Ethanol typically has a dwell time of 30 s to 1 min [
45]. QAC wipes have different dwell times depending on the components. 0.61% dodecyl dimethyl ammonium chloride in 27% ethanol and 25% isopropanol (0.61% DDAC) is used mainly in the ICU and has a dwell time of 1 min [
46], whereas the combination QAC wipe of 0.25% alkyl dimethyl ethyl benzyl ammonium chloride, 0.25% alkyl dimethyl benzyl ammonium chloride in 55% isopropanol (0.50% BAC) has a dwell time of 2 min [
47].
Most of the research performed so far on microbicide tolerance or resistance among A. baumannii isolates are performed under optimal settings with microbicides used at the proper dwell times. Therefore, a possible reason for hospital outbreaks of multidrug-resistant organisms could be due to improper cleaning techniques. Our goal is to explore the interaction of MDR bacteria with microbicides as potential contributors to hospital outbreaks. We hypothesized that in addition to accumulating drug-resistance determinants, MDR A. baumannii also accumulates mutations that allow for greater antiseptic tolerance when compared to pan-susceptible (PS) strains. We found no significant differences between MDR and PS strain survival among planktonic bacteria. As we did not find significant differences between MDR and PS strains under planktonic conditions, we tested the efficacy of antiseptic agents against bacterial biofilms. Our results demonstrated increased bacterial survival, but no differences between MDR and PS strains. Finally, we determined which strains contained the known microbicide-resistance genes and showed that significantly more microbicide-resistant bacteria survived compared to microbicide-sensitive bacteria after specific treatments. Interestingly, we found that the presence of microbicide-resistance genes was more likely in PS strains. This suggests that PS A. baumannii may be underestimated in the propagation of hospital outbreaks and highlights the need for increased hospital precautions for dealing with the spread of both MDR and PS organisms.
3. Discussion
Hospital-acquired infections can be detrimental to a patient’s health, and outbreaks can be influenced by a multitude of external factors. From 2012–2018, the number of CRAB infections was downtrending [
53]. The hospital rates of carbapenem resistance were approximately 20–35% based on antibiograms, but resistance can vary year to year [
54]. Recently, the COVID-19 pandemic resulted in increased CRAB hospital outbreaks [
55,
56,
57], which may be attributable to empiric antibiotic overuse [
56]. Given the current crisis of antibiotic resistance, preventing and controlling outbreaks has become more important than ever. Besides practicing antimicrobial stewardship, proper decontamination—a term defined by the Occupational Safety and Health Administration (OSHA) to describe the process by which pathogenic organisms are destroyed either physically or chemically—should be employed. This process can be through cleaning, sanitizing, disinfecting, or sterilizing. Cleaning refers to the removal of pathogens using a detergent or surfactant. Sanitizing refers to reducing the number of pathogens within 30 s. Disinfecting refers to the destruction of pathogens, except for sporulating organisms. Sterilizing is the process by which all pathogens are effectively killed, including those that undergo sporulation [
26]. The germicidal formulations used in this study mimic hospital disinfectant wipes used by Cooper University Hospital and are approved by the Environmental Protection Agency (EPA) to be effective against specific MDR
A. baumannii strains after one a minute exposure of bleach and DDAC, but two minutes for BAC.
The goals of this study were to understand the implications of antibiotic and microbicide-resistance among
A. baumannii isolates and to assess their potential contribution to hospital outbreaks. We selected disinfecting wipes and skin antiseptics that are routinely used in Cooper University Hospital. We showed that all wipe formulations were effective disinfectants against free-living, planktonic
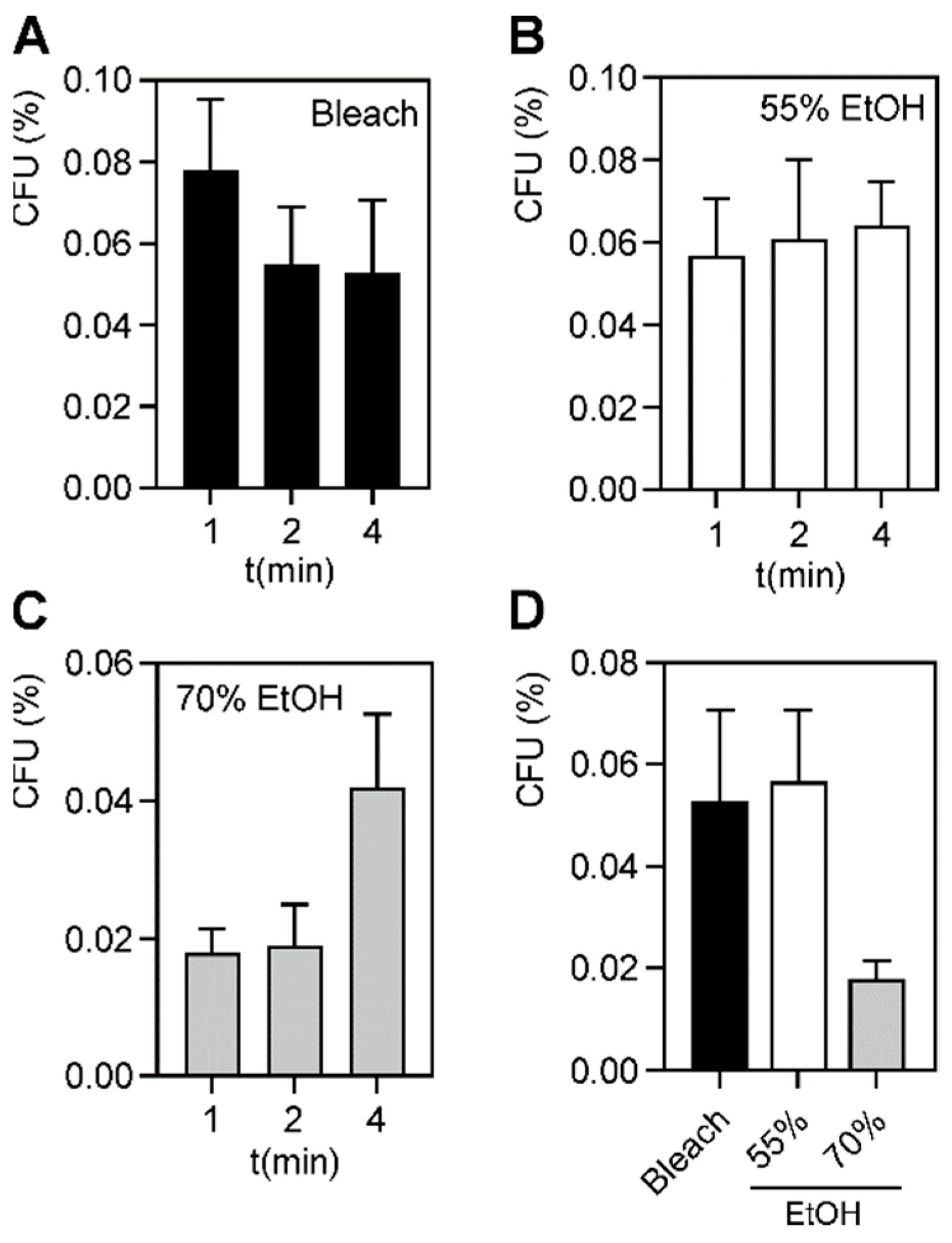
A. baumannii strains at 1, 2, and 4 min. Since we tested five different hospital wipes and two skin antiseptics, we compared the efficacy of these different microbicides. We found that povidone, CHG, and the QACs killed 100% of planktonic bacteria. However, following exposure to 0.63% bleach, 55% ethanol, and 70% ethanol at their proper dwell times, a small subpopulation (<0.1%) of bacteria repeatedly survived. For disinfectants to properly destroy pathogens, they must be used at the proper dwell time. The standard dwell times for DDAC against
A. baumannii is 1 min and 2 min for BAC [
46,
47]. Bleach is a disinfectant at 1 min, but sterilizes at 4 min, and common hospital practice recommends using a contact time of 4 min [
44]. 50% ethanol is generally germicidal after 60 s and 70% ethanol is germicidal after 30 s [
45]. Even at an extended dwell time of 4 min, we found that a small subpopulation remained (
Figure 1 and
Table S2). This suggests that in any population of
A. baumannii, microbicide-tolerant persisters may exist, even if the strains were not exposed to these agents prior. Bacterial persistence is a commonly observed phenomenon during drug exposure and is believed to be a factor in recurring bacterial infections [
58,
59]. Recently, bacterial persisters were found in
Escherichia coli in response to QACs [
60]. It is possible that these persisters contribute to the spread of
A. baumannii infections in hospital settings, as these bacteria survive well on hard surfaces [
4,
12,
20,
21,
22,
23,
24,
25]. These results were obtained following proper dwell times, so this may become even more significant if improper disinfecting techniques are used.
Antibiotic resistance is selected for when a subpopulation of bacteria survives exposure to a drug, likely by upregulating efflux pumps or obtaining additional resistance genes through plasmid exchange [
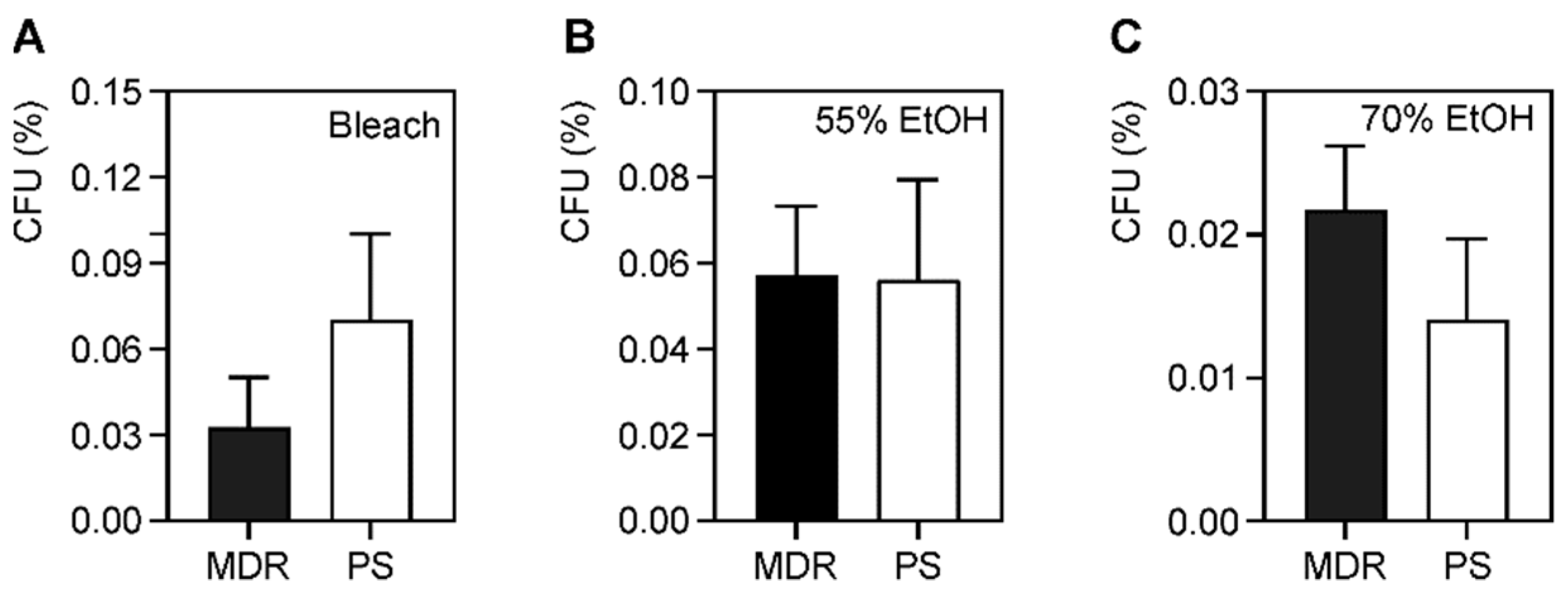
61]. A similar phenomenon likely exists for microbicide tolerance or resistance. We hypothesized that MDR strains either more easily acquire additional determinants that confer microbicide resistance or contain drug-resistance determinants that confer cross-protection against microbicides. We expected that the MDR strains would be more resistant to some microbicides than PS strains. While we did not find any differences in the percentage of survival between MDR and PS strains during the time-kill assays with bleach (
Figure 2), the MDR strains had a higher average MBC. With a higher MBC, this made the MBC/MIC ratio >4, indicating that bleach is not bactericidal against these strains, but is bacteriostatic (
Table 2). This was also true for the microbicide-resistant strains. The MDR strains also had an increased MBC to ethanol (
Table 3), whereas the microbicide-resistant strains had an increased MBC in isopropanol (
Table 4), but both these agents were still bactericidal. The increase in MBC indicates an evolutionary trend and may imply that upon further exposure to the microbicide, it is more likely that they will develop increased tolerance or resistance [
41]. These findings highlight the importance of using antiseptics well above the MIC or MBC to ensure proper disinfection [
62]. It is also important to note that we only evaluated ten strains; future studies should evaluate more strains to better understand the relationship between antibiotic and microbicidal-resistance genes.
The relationship between microbicidal and drug-resistance is equivocal. Even though some studies identified the presence of microbicidal-resistance genes in MDR pathogens [
5,
34,
36,
39], evidence of phenotypic resistance has not been demonstrated in vitro [
39,
40]. On the other hand, some research has been able to demonstrate such a relationship [
42]. It is likely that microbicide tolerance, rather than antibiotic resistance is the significant driver determining bacterial survival in adverse environments [
5,
32,
35,
37,
42]. We screened our collection for several known microbicidal-resistance genes from the SMR family that included the common efflux pump genes
qacA,
qacE, and
qacEΔ1, and
cepA. The
qac resistance genes confer resistance to quaternary ammonium compounds, such as BAC and DDAC [
35]. They also can confer resistance to specific antimicrobials, such as
qacE with sulfonamides [
35] and
cepA with CHG [
5,
32,
37]. Our data does not demonstrate a relationship between the two types of resistance, as microbicidal-resistance gene distribution in our strain collection was not associated with antimicrobial resistance (
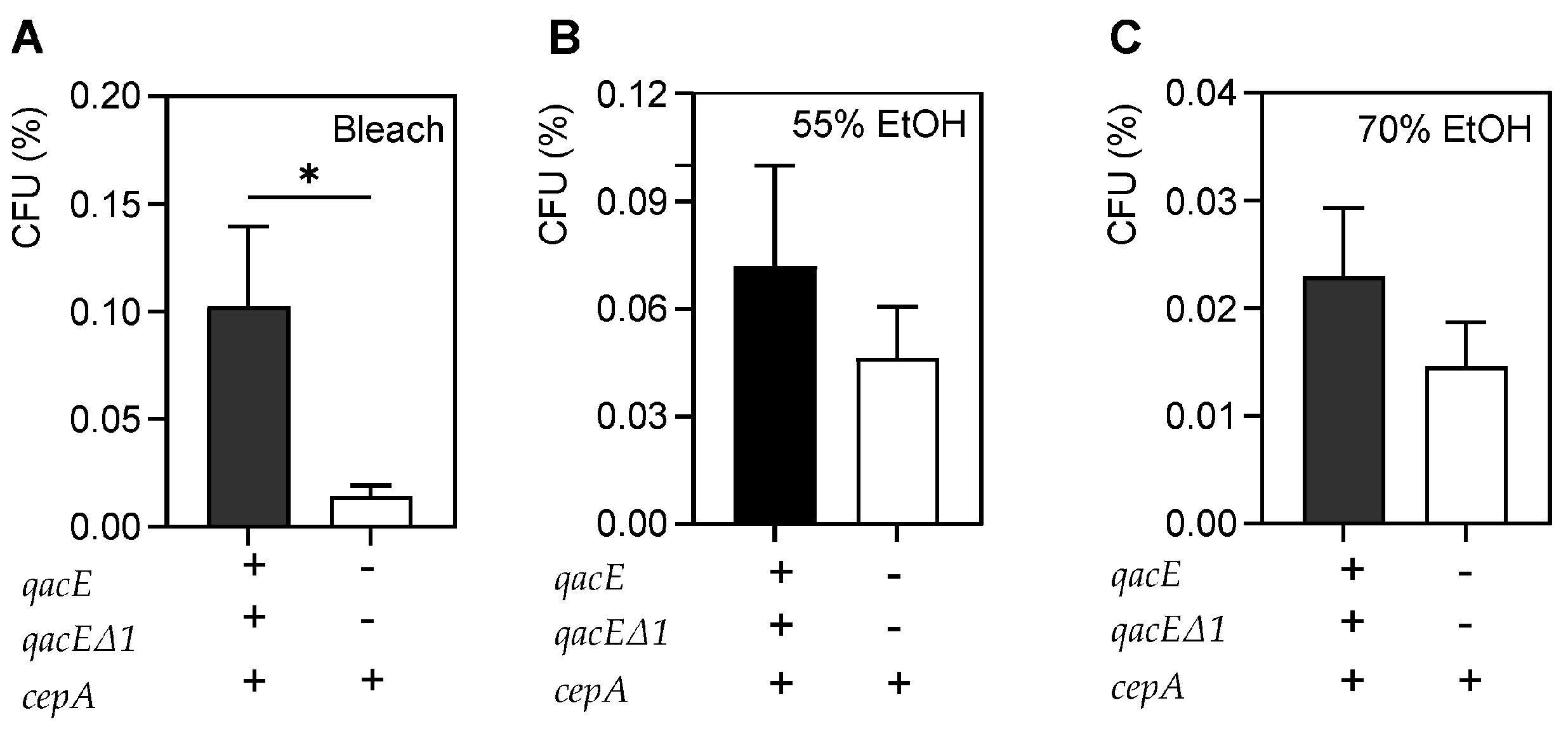
Table 1). Only one MDR strain in our collection had the three microbicide-resistance genes whereas and all others were PS. It is possible that MDR strains continue to acquire additional microbicidal-resistance genes during the evolution of a prolonged hospital outbreak. Of course, it is also possible that there are additional, uncharacterized resistance mechanisms in our collection. In addition, we do not know which efflux systems are expressed in these strains, if any. While there was no planktonic bacterial survival after QAC, povidone or CHG exposure among microbicide-resistant strains, there was a significant increase in bacterial survival after bleach treatment compared to microbicide-susceptible strains (
Figure 3). This suggests that some of these efflux pumps may be expressed and that their expression confers more tolerance to bleach exposure.
As we did not observe differences among free-living MDR and PS bacteria, we thought that differences might emerge following biofilm growth. The bacterial biofilm encases the cells an extracellular matrix that protects cells in vivo from immune defenses and antibiotics, and protects them from environmental insults such as surface disinfectants [
63,
64]. In the natural environment, the ability to form biofilms is critical for
A. baumannii survival [
65]. We assessed microbicide efficacy against
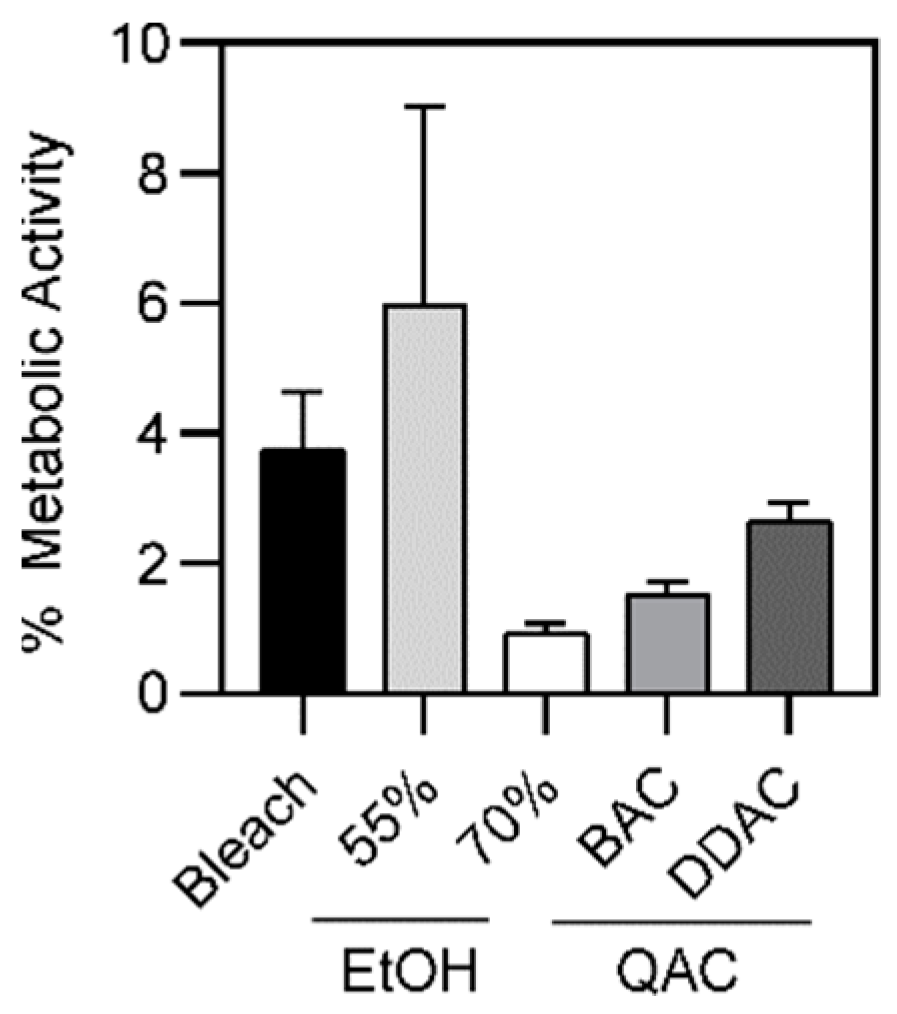
A. baumannii following biofilm formation. We found that larger percentages of the population survived, when compared to free-living bacteria. On average, 3.7%, 5.6%, 0.9%, 1.5%, and 2.6% of bacteria survived after exposure to 4 min of bleach, 1 min of 55% ethanol, 1 min of 70% ethanol, 2 min of 0.5% BAC, and 1 min of 0.61% DDAC, respectively (
Table S3). Again, increasing the exposure time did not result in an increase in bacterial killing (
Table S4). Rather, we observed that longer exposure to 70% ethanol and 0.61% DDAC resulted in statistically significant more bacterial survival compared to the 1-min time point. The small observed increase in survival is likely not biologically relevant. In addition, no differences were observed between MDR and PS strains (
Figure 6), however differences were seen between microbicide-resistant and sensitive strains (
Figure 7), suggesting that drug resistance is a separate phenomenon from microbicide resistance. This data suggests that the surviving subpopulation is tolerant or resistant to these microbicidal agents. Similar to a prior study which highlighted the connection between biofilm formation and antiseptic resistance [
28], our findings of bacterial survival of >0.1% demonstrate that biofilm formation is an innate mechanism of microbicide tolerance. Together, our results suggest the need for a more extensive decontamination process when biofilms might be involved. For instance, a previous study demonstrated that it took up to 10 min to destroy
A. baumannii biofilms [
66]. Furthermore, if there is extensive grime or other visible soil, additional wipes and pre-cleaning with applied friction is recommended for disinfection [
44,
46,
47]. Our findings, along with prior studies suggest that bacterial biofilms may require pre-cleaning prior to disinfection. As it is difficult to know when bacterial biofilms are present, the safest option is to implement a pre-cleaning step with use of friction as part of standard hospital operating procedures.
In sum, a more holistic approach that extends beyond surface decontamination may be needed to combat HAI’s. Surfaces in hospitals and health care facilities serve as reservoirs for pathogens, and hands are usually the vector transferring those microorganisms to patients. Thus, hand hygiene is important in minimizing transfer of pathogens. Cleaning and disinfecting the environment where the patient resides is fundamental to reduce the microorganism burden in patient’s environment. Most, if not all housekeeping surfaces need to be cleaned with soapy water or a detergent disinfectant, according to product specifications, where an emphasis is placed on scheduling and the technique used to clean and disinfect [
67,
68]. In addition, to focus on the environmental surfaces, there has been heavy reliance on development of strategies to prevent HAIs, often referred to as “bundles” of infection prevention. A method called the “bundle” approach consists of ongoing hospital staff education with regards to maintaining strict hand hygiene, isolation precautions, environmental cleaning, and surveillance of high-risk areas. In 2009, a hospital in Spain assessed the long-term impact of the bundle approach from 1994 to 2003. Results showed that the rates of colonization and infection significantly decreased from 0.82 cases per 100 admissions in 1994, to 0.46 in 1996–1997, to 0.21 in 1998–2003 [
69]. Thus, while proper decontamination is important, a multi-disciplinary approach is essential when attempting to combat this problematic pathogen.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}