
Rational Use of Danofloxacin for Treatment of Mycoplasma gallisepticum in Chickens Based on the Clinical Breakpoint and Lung Microbiota Shift

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Isolation and Cultivation of MG
2.3. Animals
2.4. Determination of ECV
2.4.1. Antimicrobial Susceptibility Determination
2.4.2. Formulation of ECV
2.5. Establishment of COPD
2.5.1. Selection of Pathogenic Strain M19
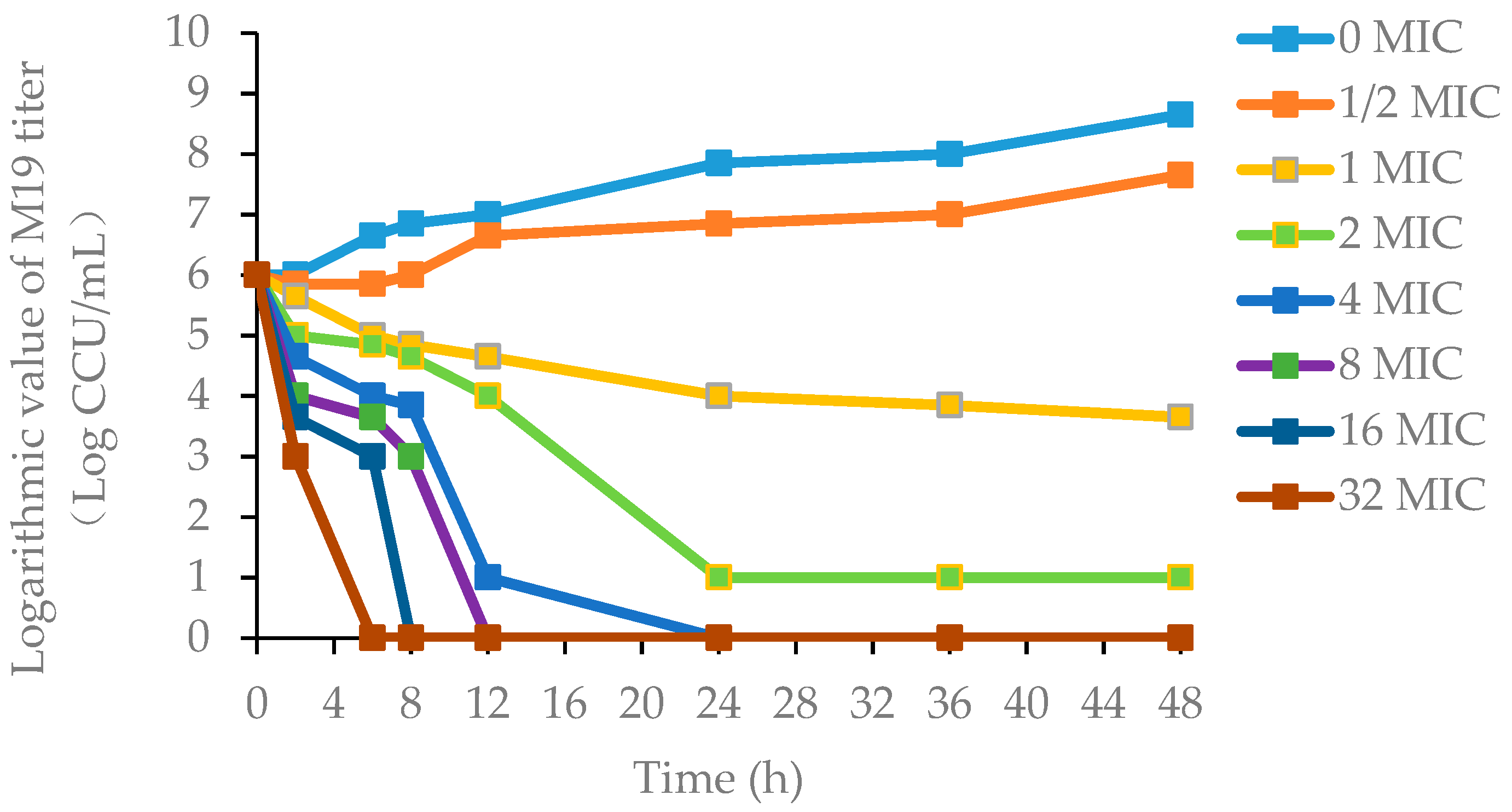
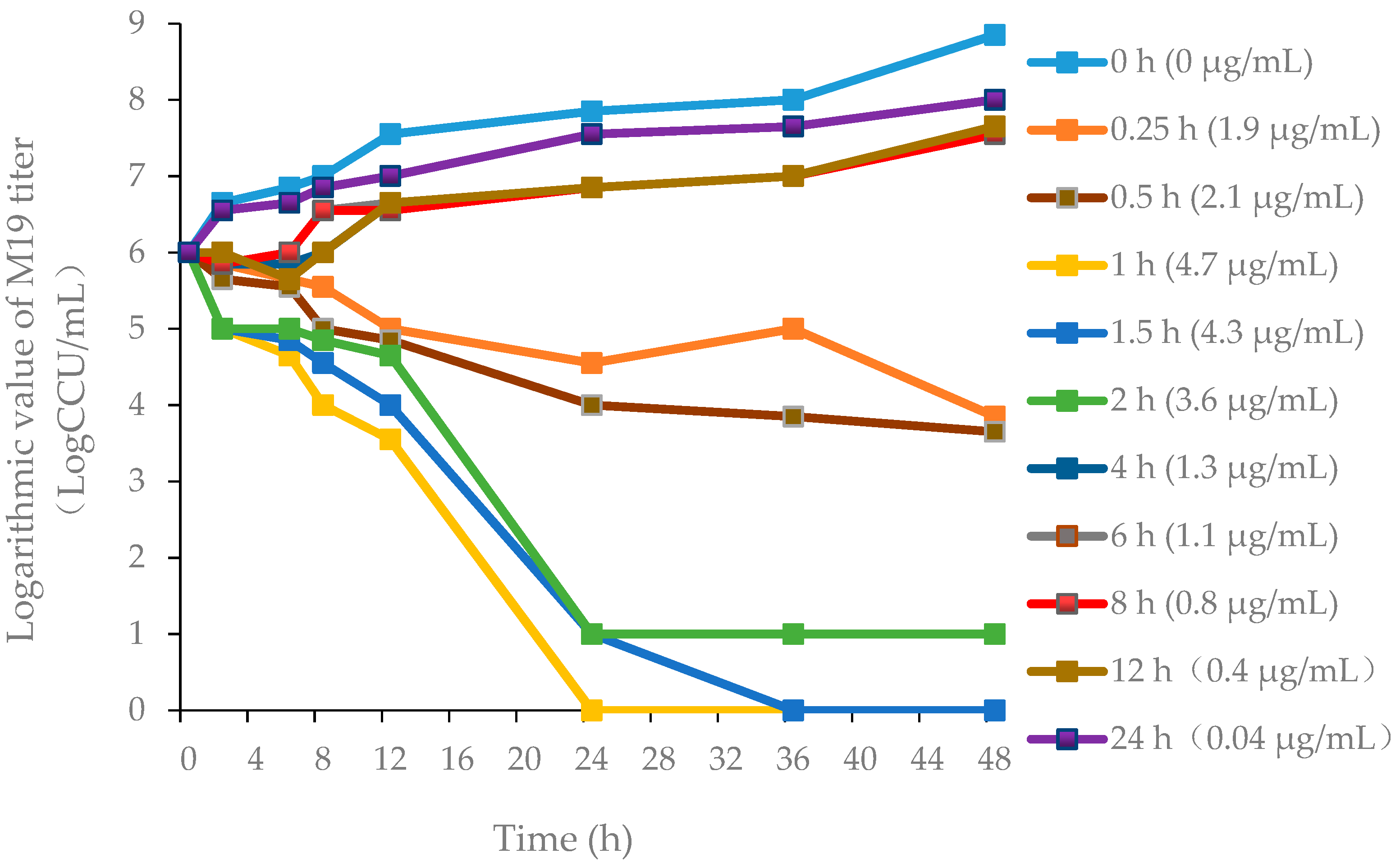
2.5.2. PD Study of M19 In Vitro and Ex Vivo
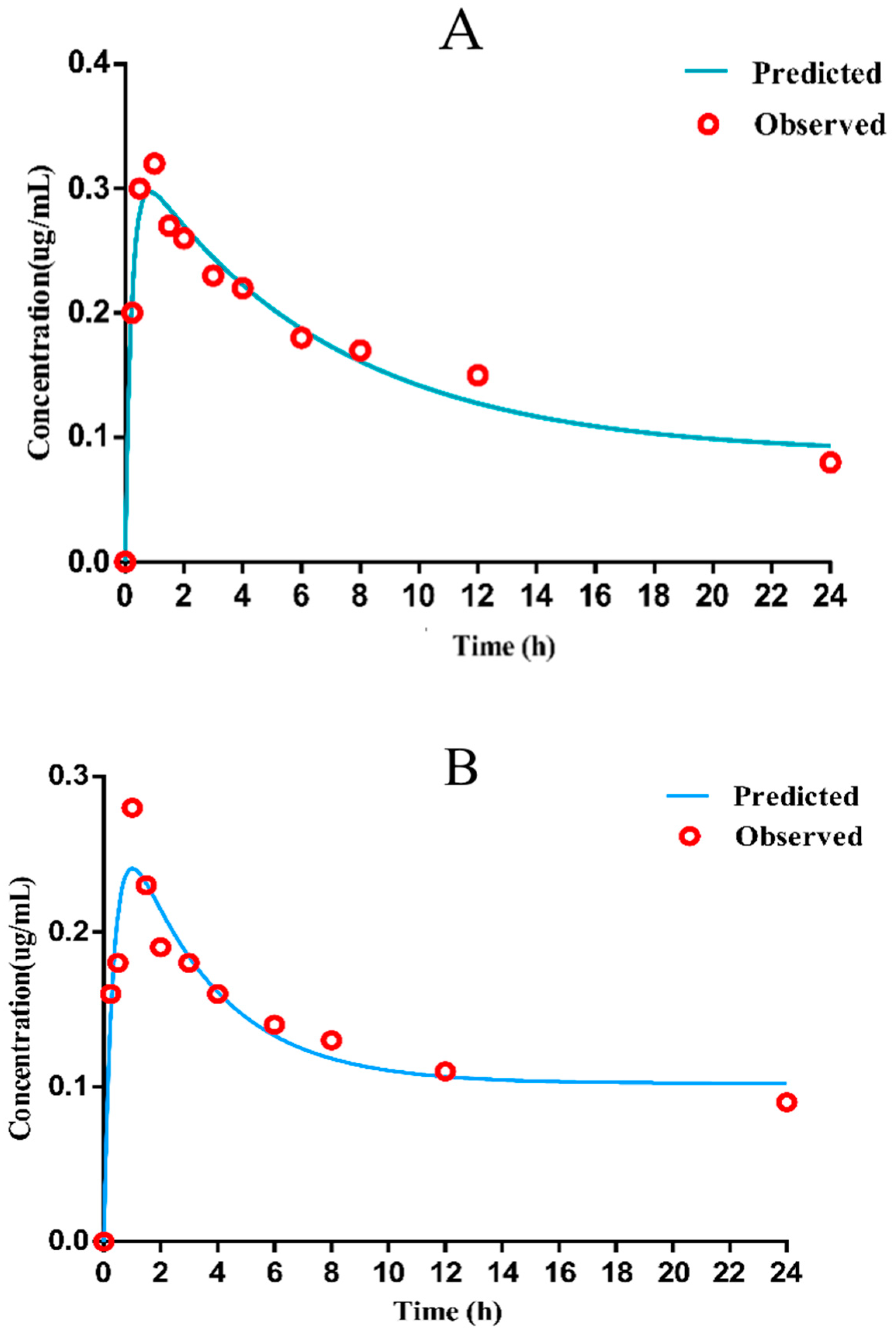
2.5.3. PK Studies of Danofloxacin in Chicken Plasma and Lung
2.5.4. High-Performance Liquid Chromatography (HPLC) Method for Danofloxacin Determination
2.5.5. The Protein Binding Rate
2.5.6. PK Analysis
2.5.7. PK–PD Integration and Modeling
2.5.8. The Monte Carlo Analysis and Establishment of COPD
2.6. Exploration of COCL
2.6.1. Dose Regimen Based on PK–PD Model
2.6.2. Infection Model and Clinical Trials
2.6.3. The Establishment of the Standard Curve of MG by Real-Time PCR
2.6.4. Statistical Analysis for the Establishment of COCL
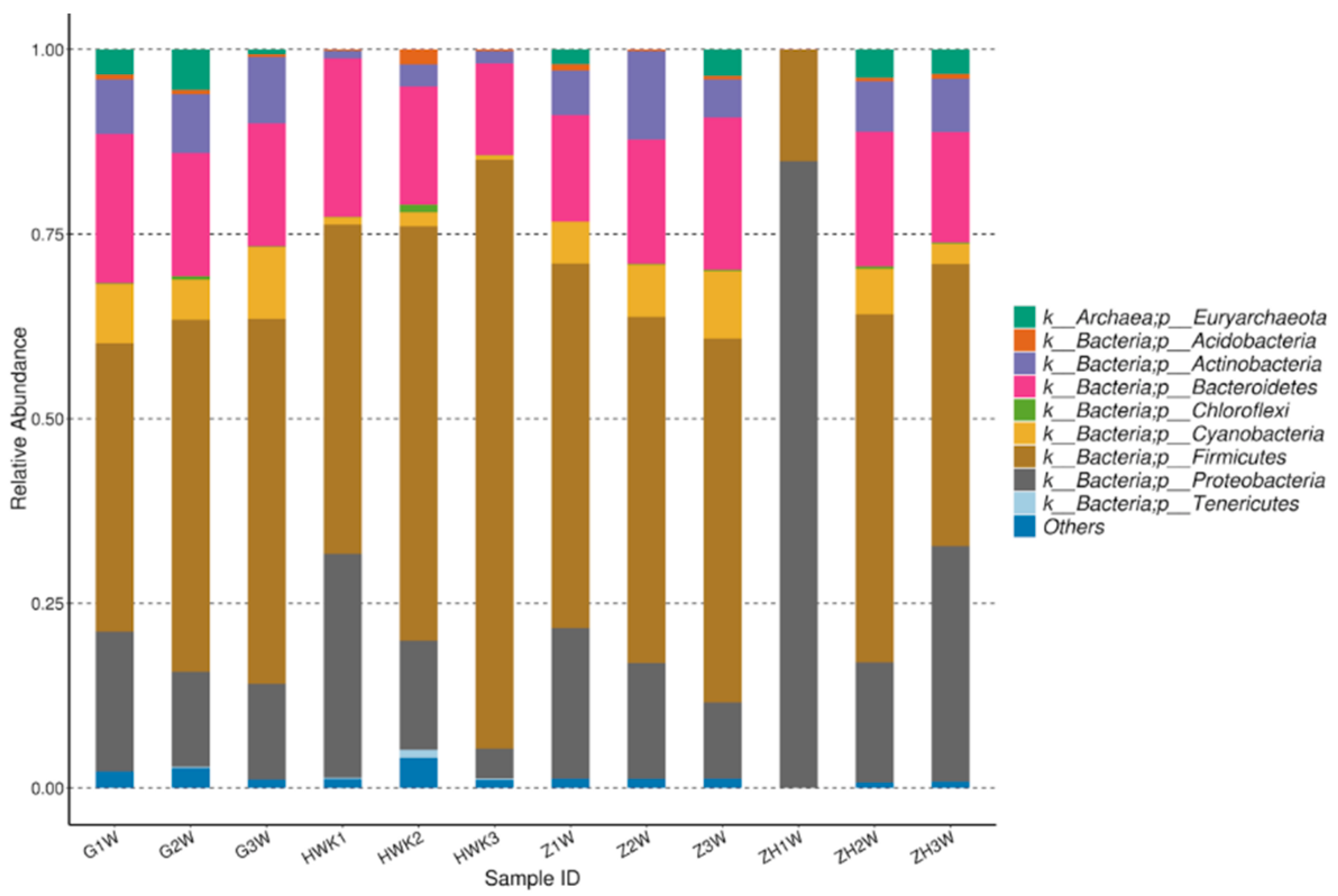
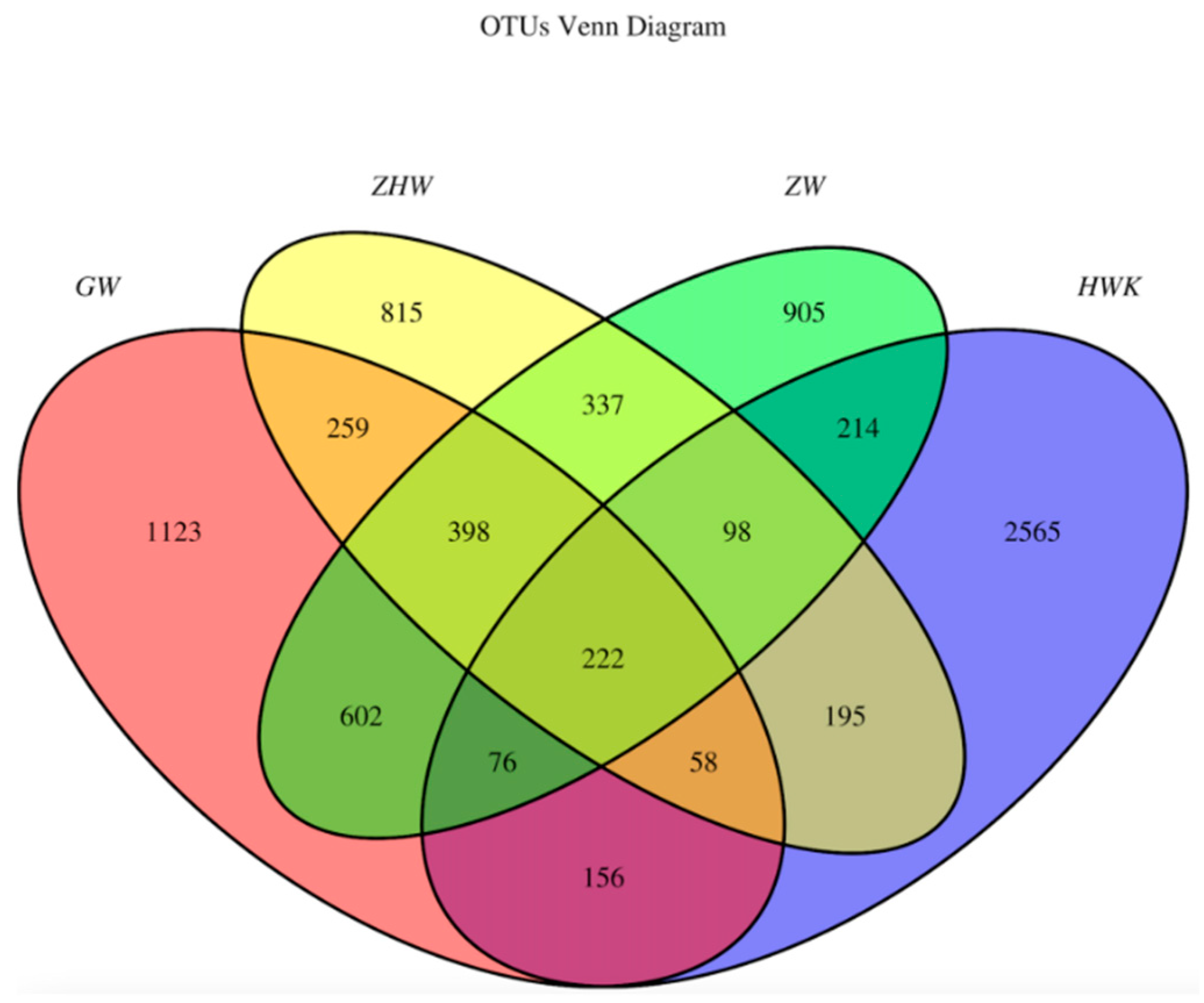
2.6.5. Evaluation of the Effect of Danofloxacin on the Lung Microbiota of MG Infected Chickens
3. Results
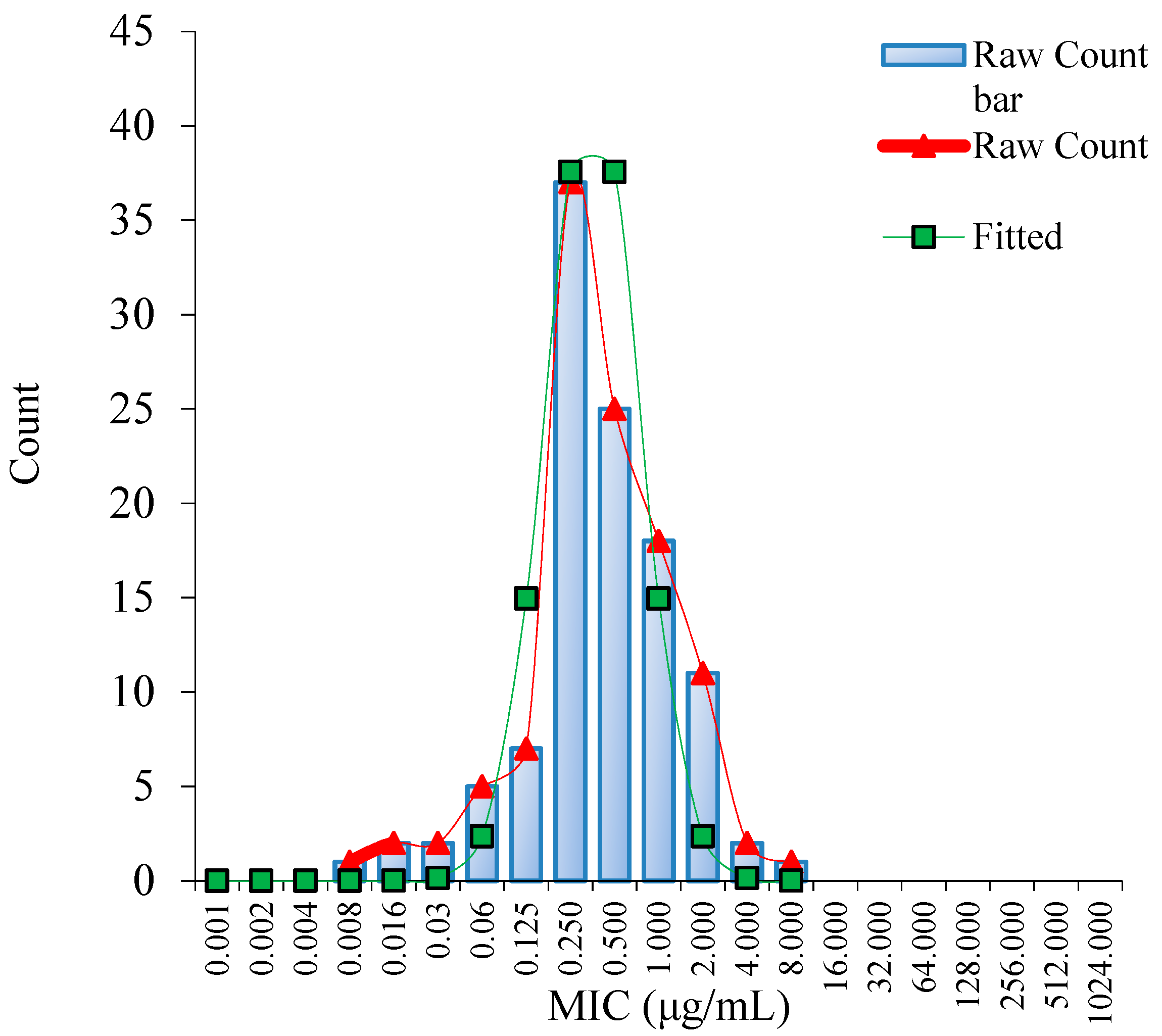
3.1. MIC Distribution and Establishment of the ECV
3.2. COPD for Danofloxacin against MG
3.2.1. Pharmacodynamics of Danofloxacin against MG in the Plasma and Lung
3.2.2. Verification of HPLC Method for Determination of Danofloxacin
3.2.3. PK Parameters of Danofloxacin in Plasma and Lung
3.2.4. Integration Modeling
3.2.5. Monte Carlo Simulation and COPD
3.3. COCL for Danofloxacin against MG
3.3.1. Dose Regiment Based on the PK–PD Model
3.3.2. Establishment of the Standard Curve of MG by Real-Time Fluorescence Quantitative PCR
3.3.3. Clinical Efficacy Assessment Indicators and Bacteriological Prognosis
3.3.4. Determination of COCL
3.3.5. Evaluation of the Effect of Danofloxacin on the Lung Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hawley, D.M.; Moyers, S.C.; Johanel, C.; Courtney, Y.; Adelman, J.S. Characterization of unilateral conjunctival inoculation with Mycoplasma gallisepticum in house finches. Avian Pathol. 2018, 47, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Bwala, D.G.; Ponman, S.; Duncan, N.; Wandrag, D.; Abolnik, C. Assessment of Mycoplasma gallisepticum vaccine efficacy in a co-infection challenge model with qx-like infectious bronchitis virus. Avian Pathol. 2018, 47, 261–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazakiewicz, D.; Karr, J.R.; Langner, K.M.; Plewczynski, D. A combined systems and structural modeling approach repositions antibiotics for Mycoplasma genitalium. Comput. Biol. Chem. 2015, 59, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Semashko, T.A.; Arzamasov, A.A.; Fisunov, G.Y.; Govorun, V.M. Transcription profiling data set of different states of Mycoplasma gallisepticum. Genom. Data 2016, 11, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Deguchi, T.; Nakane, K.; Yasuda, M.; Yokoi, S.; Ito, S.-I.; Nakano, M.; Ito, S.; Ishiko, H. Macrolide Resistance–associated 23S rRNA Mutation in Mycoplasma genitalium, Japan. Emerg. Infect. Dis. 2011, 17, 1148–1150. [Google Scholar] [CrossRef]
- Heleili, N.; Ayachi, A.; Mamache, B.; Chelihi, A. Seroprevalence of Mycoplasma synoviae and Mycoplasma gallisepticum at batna commercial poultry farms in algeria. Vet. World. 2012, 5, 709–712. [Google Scholar] [CrossRef]
- Vogl, G.; Plaickner, A.; Szathmary, S.; Stipkovits, L.; Rosengarten, R.; Szostak, M.P. Mycoplasma gallisepticum invades chicken erythrocytes during infection. Infect. Immun. 2008, 76, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Gharibi, D.; Ghadimipour, R.; Mayahi, M. Detection of Mycoplasma gallisepticum and Mycoplasma synoviae among Commercial Poultry in Khouzestan Province, Iran. Arch. Razi Inst. 2018, 73, 139–146. [Google Scholar]
- Gosal, N.S.; Rampai, S.; Goyal, S.; Chaudhary, R.K. Effect of paracetamol on the pharmacokinetics of danofloxacin in buffalo calves. Indian Vet. J. 2009, 86, 466–468. [Google Scholar]
- Mohammadi, G.R.; Ghazvini, K.; Abbaspanah, H. Antimicrobial susceptibility testing of Mannheimia haemolytica and Pasteurella multocida isolated from calves with dairy calf pneumonia. Arch. Razi Inst. 2006, 61, 91–96. [Google Scholar]
- Xiao, X.; Pei, L.; Jiang, L.J.; Lan, W.X.; Zhang, Z.Q. In vivo pharmacokinetic/pharmacodynamic profiles of danofloxacin in rabbits infected with Salmonella typhimurium after oral administration. Front. Pharmacol. 2018, 9, 391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Kang, Z.; Yao, L.; Gu, X.; Huang, Z.; Cai, Q.; Shen, X.; Ding, H. Pharmacokinetic/pharmacodynamic integration to evaluate the changes in susceptibility of Actinobacillus pleuropneumoniae after repeated administration of danofloxacin. Front. Microbiol. 2018, 9, 2445. [Google Scholar] [CrossRef]
- Xu, Z.H.; Huang, A.X.; Luo, X.; Zhang, P.; Huang, L.L.; Wang, X.; Mi, K.; Fang, S.W.; Huang, X.; Li, J.; et al. Exploration of Clinical Breakpoint of Danofloxacin for Glaesserella parasuis in Plasma and in PELF. Antibiotics 2021, 10, 808. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Wu, Y.; Huang, Z.; Zhang, C.; Zhang, L.; Cai, Q.; Shen, X.; Jiang, H.; Ding, H.; Browning, G.F. Relationship between danofloxacin pk/pd parameters and emergence and mechanism of resistance of Mycoplasma gallisepticum in In Vitro model. PLoS ONE 2018, 13, e0202070. [Google Scholar] [CrossRef] [PubMed]
- Migaki, T.T.; Avakian, A.P.; Barnes, H.J.; Ley, D.H.; Tanner, A.C.; Magonigle, R.A. Efficacy of danofloxacin and tylosin in the control of Mycoplasmosis in chicks infected with tylosin-susceptible or tylosin-resistant field isolates of Mycoplasma gallisepticum. Avian Dis. 1993, 37, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.C.; Fuller, J.R.; Fuller, M.K.; Whittlestone, P.; Wise, D.R. In vitro activity of danofloxacin, tylosin and oxytetracycline against Mycoplasmas of veterinary importance. Res. Vet. Sci. 1993, 54, 329–334. [Google Scholar] [CrossRef]
- Dong, H.; Xie, J.; Chen, L.; Wang, T.; Sun, J.; Zhao, Y.; Dong, Y. Developments in the pharmacokinetic-pharmacodynamic index of linezolid a step toward dose optimization using Monte Carlo simulation in critically ill patients. Int. J. Infect. Dis. 2014, 22, 35–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.F.; Peng, H.M.; Bu, M.X. Pharmacodynamic evaluation and pk/pd-based dose prediction of tulathromycin: A potential new indication for Streptococcus suis infection. Front. Pharmacol. 2017, 8, 684. [Google Scholar] [CrossRef]
- Mouton, J.W.; Theuretzbacher, U.; Craig, W.A.; Tulkens, P.M.; Derendorf, H.; Cars, O. Tissue concentrations: Do we ever learn? J. Antimicrob. Chemother. 2008, 61, 235–237. [Google Scholar] [CrossRef] [Green Version]
- Foster, D.M.; Martin, L.G.; Papich, M.G.; Torrelles, J.B. Comparison of active drug concentrations in the pulmonary epithelial lining fluid and interstitial fluid of calves injected with enrofloxacin, florfenicol, ceftiofur, or tulathromycin. PLoS ONE 2016, 11, e0149100. [Google Scholar] [CrossRef]
- Messenger, K.M.; Papich, M.G.; Blikslager, A.T. Distribution of enrofloxacin and its active metabolite, using an in vivo ultrafiltration sampling technique after the injection of enrofloxacin to pigs. J. Vet. Pharmacol. Ther. 2012, 35, 452–459. [Google Scholar] [CrossRef] [PubMed]
- Menge, M.; Rose, M.; Bohland, C.; Zschiesche, E.; Nürnberger, M. Pharmacokinetics of tildipirosin in bovine plasma, lung tissue, and bronchial fluid (from live, nonanesthetized cattle). J. Vet. Pharmacol. Ther. 2011, 35, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.L.; Foster, D.M.; Papich, M.G. Pharmacokinetics and tissue distribution of enrofloxacin and its active metabolite ciprofloxacin in calves. J. Vet. Pharmacol. Ther. 2010, 30, 564–571. [Google Scholar] [CrossRef] [PubMed]
- Toutain, P.L.; Sidhu, P.K.; Lees, P.; Rassouli, A.; Pelligand, L. VetCAST Method for determination of the pharmacokinetic-pharmacodynamic cut-off values of a long-acting formulation of florfenicol to support clinical breakpoints for florfenicol antimicrobial susceptibility testing in cattle. Front. Microbiol. 2019, 10, 1310. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Development of In Vitro Susceptibility Testing Criteria and Quality Control Parameters for Veterinary Antimicrobial Agents; Approved Standard, 3rd ed.; CLSI Document M37-A3 Wayne; Clinical and Laboratory Standards Institute: Pittsburgh, PA, USA, 2007; p. M37-A3. [Google Scholar]
- Toutain, P.L.; Bousquet-Mélou, A.; Damborg, P.; Ferran, A.A.; Mevius, D.; Pelligand, L.; Veldman, K.T.; Lees, P. En Route towards European Clinical Breakpoints for Veterinary Antimicrobial Susceptibility Testing: A Position Paper Explaining the VetCAST Approach. Front. Microbiol. 2017, 8, 2344. [Google Scholar] [CrossRef] [Green Version]
- Turnidge, J.D.; Martinez, M.N. Proposed method for estimating clinical cut-off (COCL) values: An attempt to address challenges encountered when setting clinical breakpoints for veterinary antimicrobial agents. Vet. J. 2017, 228, 33–37. [Google Scholar] [CrossRef]
- Turnidge, J.; Kahlmeter, G.; Kronvall, G. Statistical characterisation of bacterial wild-type MIC value distributions and the determination of epidemiological cut-off values. Clin. Microbiol. Infect. 2006, 12, 418–425. [Google Scholar] [CrossRef]
- European Medicines Agency. Guideline on the Use of Pharmacokinetics and Pharmacodynamics in the Development of Antimicrobial Medicinal Products; EMA: Amsterdam, The Netherlands, 2015.
- Zhou, H.; Zhao, X.; Sun, L.; Liu, Y.; Lv, Y.; Gang, X. Gut Microbiota Profile in Patients with Type 1 Diabetes Based on 16S rRNA Gene Sequencing: A Systematic Review. Dis. Markers 2020, 27, 3936247. [Google Scholar] [CrossRef]
- Taylor, J.D.; Doyle, D.J.; Blackall, P.J.; Confer, A.W. Use of REP-PCR and 16S rRNA gene sequencing for comparison of Mannheimia haemolytica isolates obtained from fatal cases of bovine respiratory disease in the USA and Australia. Aust. Vet. J. 2014, 92, 15–23. [Google Scholar] [CrossRef]
- Kinoshita, Y.; Niwa, H.; Katayama, Y.; Hariu, K. Dominant obligate anaerobes revealed in lower respiratory tract infection in horses by 16S rRNA gene sequencing. J. Vet. Med. Sci. 2014, 76, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Allali, I.; Arnold, J.W.; Roach, J.; Cadenas, M.B.; Butz, N.; Hassan, H.M.; Koci, M.; Ballou, A.; Mendoza, M.; Ali, R.; et al. A comparison of sequencing platforms and bioinformatics pipelines for compositional analysis of the gut microbiome. BMC Microbiol. 2017, 17, 194. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, M.Z.; Malys, T.; Ivanov, Y.V.; Park, J.; Shabbir, M.A.; Rabbani, M.; Yaqub, T.; Harvill, E.T. Microbial communities present in the lower respiratory tract of clinically healthy birds in Pakistan. Poult. Sci. 2015, 94, 612–620. [Google Scholar] [CrossRef] [PubMed]
- Glendinning, L.; McLachlan, G.; Vervelde, L. Age-related differences in the respiratory microbiota of chickens. PLoS ONE 2017, 12, e0188455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhang, M.; Liu, Q.; Feng, J. The alterations of tracheal microbiota and inflammation caused by different levels of ammonia exposure in broiler chickens. Poult. Sci. 2020, 100, 685–696. [Google Scholar] [CrossRef]
- Khatoon, H.; Afzal, F.; Tahir, M.F.; Hussain, M.; Khan, S.U. Prevalence of mycoplasmosis and antibiotic susceptibility of Mycoplasma gallisepticum in commercial chicken flocks of Rawalpindi division, Pakistan. Pak. Vet. J. 2018, 38, 446–448. [Google Scholar] [CrossRef]
- Xiao, X.; Sun, J.; Chen, Y.; Zou, M.; Zhao, D.H.; Liu, Y.H. Ex vivo pharmacokinetic and pharmacodynamic analysis of valnemulin against Mycoplasma gallisepticum s6 in Mycoplasma gallisepticum and Escherichia coli co-infected chickens. Vet. J. 2015, 204, 54–59. [Google Scholar] [CrossRef]
- Kronvall, G. Normalized Resistance Interpretation as a Tool for Establishing Epidemiological MIC Susceptibility Breakpoints. J. Clin. Microbiol. 2010, 48, 4445–4452. [Google Scholar] [CrossRef] [Green Version]
- Canton, E.; Peman, J.; Hervas, D.; Iniguez, C.; Navarro, D.; Echeverria, J.; Martinez-Alarcon, J.; Fontanals, D.; Gomila-Sard, B.; Buendia, B. Comparison of Three Statistical Methods for Establishing Tentative Wild-Type Population and Epidemiological Cutoff Values for Echinocandins, Amphotericin B, Flucytosine, and Six Candida Species as Determined by the Colorimetric Sensititre YeastOne Method. J. Clin. Microbiol. 2012, 50, 3921–3926. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhang, C.; Aragon, V.; Zhou, X.; Zou, M.; Wu, C.; Shen, Z. Investigation of Haemophilus parasuis from healthy pigs in China. Vet. Microbiol. 2019, 231, 40–44. [Google Scholar] [CrossRef] [Green Version]
- Zain, Z.M.; Bradbury, J.M. The influence of type of swab and laboratory method on the recovery of Mycoplasma gallisepticum and Mycoplasma synoviae in broth medium. Avian Pathol. 1995, 24, 707–716. [Google Scholar] [CrossRef]
- Tian, E.; Chen, C.; Hu, W.; Miao, Y.; Muhammad, I.; Zhang, Q.; Liu, Y.; Xu, L.; Bao, J.; Ding, L.; et al. Population pharmacokinetics for danofloxacin in the intestinal contents of healthy and infected chickens. J. Vet. Pharmacol. Ther. 2019, 42, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Geornaras, I.; von-Holy, A. Antimicrobial susceptibilities of isolates of Staphylococcus aureus, Listeria species and Salmonella serotypes associated with poultry processing. Int. J. Food Microbiol. 2001, 70, 29–35. [Google Scholar] [CrossRef]
- Yadav, J.P.; Tomar, P.; Singh, Y.; Khurana, S.K. Insights on Mycoplasma gallisepticum and Mycoplasma synoviae infection in poultry: A systematic review. Anim. Biotechnol. 2021, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Marouf, S.; Khalf, M.A.; Alorabi, M.; El-Shehawi, A.M.; El-Tahan, A.M.; El-Hack, M.E.A.; El-Saadony, M.T.; Salem, H.M. Mycoplasma gallisepticum: A devastating organism for the poultry industry in Egypt. Poult. Sci. 2022, 101, 101658. [Google Scholar] [CrossRef] [PubMed]
- Feberwee, A.; de-Wit, S.; Dijkman, R. Clinical expression, epidemiology, and monitoring of Mycoplasma gallisepticum and Mycoplasma synoviae: An update. Avian Pathol. 2022, 51, 2–18. [Google Scholar] [CrossRef]
- De-Jong, A.; Youala, M.; Klein, U.; El-Garch, F.; Simjee, S.; Moyaert, H.; Rose, M.; Gautier-Bouchardon, A.V.; Catania, S.; Ganapathy, K.; et al. Minimal inhibitory concentration of seven antimicrobials to Mycoplasma gallisepticum and Mycoplasma synoviae isolates from six European countries. Avian Pathol. 2021, 50, 161–173. [Google Scholar] [CrossRef]
- Rodriguez-Tudela, J.L.; Alcazar-Fuoli, L.; Mellado, E.; Alastruey-Izquierdo, A.; Monzon, A.; Cuenca-Estrella, M. Epidemiological cutoffs and cross-resistance to azole drugs in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2008, 52, 2468–2472. [Google Scholar] [CrossRef] [Green Version]
- Arendrup, M.C.; Garcia-Effron, G.; Lass-Florl, C.; Lopez, A.G.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Perlin, D.S. Echinocandin susceptibility testing of Candida species: Comparison of EUCAST EDef 7.1, CLSI M27-A3, Etest, disk diffusion, and agar dilution methods with RPMI and isosensitest media. Antimicrob. Agents Chemother. 2010, 54, 426–439. [Google Scholar] [CrossRef] [Green Version]
- Turnidge, J.; Paterson, D.L. Setting and revising antibacterial susceptibility breakpoints. Clin. Microbiol. Rev. 2007, 20, 391–408. [Google Scholar] [CrossRef] [Green Version]
- Hannan, P. Guidelines and recommendations for antimicrobial minimum inhibitory concentration (MIC) testing against veterinary Mycoplasma species. Vet. Res. 2000, 31, 373–395. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, R.A.; Gebriel, M.G.; Kadry, H.M.; Ahmed, M. Carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa: Characterization of carbapenemase genes and E-test evaluation of colistin-based combinations. Infect. Drug. Resist. 2018, 11, 1261–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohner, P.C.; Patel, R.; Uhl, J.R.; Garin, K.M.; Hopkins, M.K.; Wegener, L.T.; Cockerill, F.R. Comparison of agar dilution, broth microdilution, e-test, disk diffusion, and automated vitek methods for testing susceptibilities of Enterococcus spp. to vancomycin. J. Clin. Microbiol. 1997, 35, 3258–3263. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Ye, X.; Wu, Y.; Huang, Z.; Gu, X.; Cai, Q.; Shen, Q.; Jiang, H.; Ding, H.; Balish, M.F. Determination of the mutant selection window and evaluation of the killing of Mycoplasma gallisepticum by danofloxacin, doxycycline, tilmicosin, tylvalosin and valnemulin. PLoS ONE 2017, 12, e0169134. [Google Scholar] [CrossRef] [PubMed]
- Tatay-Dualde, J.; Prats-van der Ham, M.; de la Fe, C.; Paterna, A.; Sánchez, A.; Corrales, J.C.; Contreras, A.; Tola, S.; Gómez-Martin, Á. Antimicrobial susceptibility and multilocus sequence typing of Mycoplasma capricolum subsp. capricolum. PLoS ONE 2017, 12, e0174700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jelinski, M.; Kinnear, A.; Gesy, K.; Andrés-Lasheras, S.; Zaheer, R.; Weese, S.; McAllister, T.A. Antimicrobial Sensitivity Testing of Mycoplasma bovis Isolates Derived from Western Canadian Feedlot Cattle. Microorganisms 2020, 8, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.; Li, J.; Liu, J.; Guan, X.; Gong, J.; Liu, L.; He, L.; Meng, F.; Zhang, J. Antimicrobial susceptibility and molecular characteristics of Mycoplasma pneumoniae isolates across different regions of China. Antimicrob. Resist. Infect. Control 2019, 8, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantón, R. Aspectos microbiológicos actuales de la infección respiratoria comunitaria más allá de la COVID-19 [Current microbiological aspects of community respiratory infection beyond COVID-19]. Rev. Esp. Quimioter. 2021, 34, 81–92. [Google Scholar] [CrossRef]
- Huang, Z.; Wu, Y.; Zhou, Z.; Xia, X.; Gu, X.; Cai, Q.; Shen, X.; Yang, H.; Ding, H. Pharmacokinetic and Pharmacodynamic Integration and Resistance Analysis of Tilmicosin against Mycoplasma gallisepticum in an In Vitro Dynamic Model. Front. Pharmacol. 2019, 10, 670. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Gu, X.; Ye, X.; Wu, X.; Zhang, B.; Zhang, L. The PK/PD Interactions of Doxycycline against Mycoplasma gallisepticum. Front. Microbiol. 2016, 7, 653. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Sun, J.; Yang, T.; Fang, X.; Wu, D.; Xiong, Y.Q.; Cheng, J.; Chen, Y.; Shi, W.; Liu, Y.H. In vivo pharmacokinetic/pharmacodynamic profiles of valnemulin in an experimental intratracheal Mycoplasma gallisepticum infection model. Antimicrob. Agents Chemother. 2015, 59, 3754–3760. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Sun, J.; Yang, T.; Fang, X.; Cheng, J.; Xiong, Y.Q.; Liu, Y.H. Pharmacokinetic/Pharmacodynamic Profiles of Tiamulin in an Experimental Intratracheal Infection Model of Mycoplasma gallisepticum. Front. Vet. Sci. 2016, 3, 75. [Google Scholar] [CrossRef] [Green Version]
- Arzey, G.G.; Arzey, K.E. Successful treatment of mycoplasmosis in layer chickens with single dose therapy. Aust. Vet. J. 1992, 69, 126–128. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.; Scaglione, F.; Derendorf, H. Class-dependent relevance of tissue distribution in the interpretation of anti-infective pharmacokinetic/pharmacodynamic indices. Int. J. Antimicrob. Agents 2010, 35, 431–438. [Google Scholar] [CrossRef]
- Huang, A.X.; Wang, S.G.; Guo, J.L.; Gu, Y.F.; Li, J.; Huang, L.L.; Wang, X.; Tao, Y.F.; Liu, Z.L.; Yuan, Z.H.; et al. Prudent Use of Tylosin for Treatment of Mycoplasma gallisepticum Based on Its Clinical Breakpoint and Lung Microbiota Shift. Front. Microbiol. 2021, 12, 712473. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cheng, P.; Xiao, T.; Ulzikhutag, J.; Zhang, X. Pharmacokinetics and pharmacodynamics integration of danofloxacin against Eschrichia coli in piglet ileum ultrafiltration probe model. Sci. Rep. 2021, 11, 681. [Google Scholar] [CrossRef]
- Bonassa, K.P.D.; Miragliotta, M.Y.; Simas, R.C.; Eberlin, M.N.; Anadón, A.; Moreno, R.A.; Reyes, F.G.R. Pharmacokinetics, Pharmacodynamic Efficacy Prediction Indexes and Monte Carlo Simulations of Enrofloxacin Hydrochloride against Bacterial Strains That Induce Common Clinical Diseases in Broiler Chickens. Front. Vet. Sci. 2021, 7, 606872. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Varón, E.; García-Romero, E.; Serrano-Rodríguez, J.M.; Cárceles, C.M.; García-Galán, A.; Cárceles-García, C.; Fernández, R.; Muñoz, C.; de-la-Fe, C. PK/PD Analysis of Marbofloxacin by Monte Carlo Simulation against Mycoplasma agalactiae in Plasma and Milk of Lactating Goats after IV, SC and SC-Long Acting Formulations Administration. Animals 2021, 11, 1104. [Google Scholar] [CrossRef]
- Zhang, N.; Wu, Y.; Huang, Z.; Yao, L.; Zhang, L.; Cai, Q.; Shen, X.; Jiang, H.; Ding, H. The PK-PD Relationship and Resistance Development of Danofloxacin against Mycoplasma gallisepticum in an In Vivo Infection Model. Front. Microbiol. 2017, 8, 926. [Google Scholar] [CrossRef] [Green Version]
- Kum, C.; Gokbulut, C.; Sekkin, S.; Boyacioglu, M. Pharmacokinetics of danofloxacin following intravenous and intramuscular administration in donkeys. J. Vet. Pharmacol.Ther. 2009, 32, 105–108. [Google Scholar] [CrossRef]
- Ehtisham-Ul-Haque, S.; Rahman, S.U.; Khan, M.I.; Younus, M.; Nasir, A. A simplified duplex real-time pcr incorporating taqman minor groove binder (mgb) probes and an exogenous internal positive control for the simultaneous detection of Mycoplasma gallisepticum and Mycoplasma synoviae cultures. Vet. Med. 2015, 60, 268–273. [Google Scholar] [CrossRef] [Green Version]
- Toutain, P.L. Setting Clinical Breakpoint Methodological Aspects; European Committee on Antimicrobial Susceptibility Testing: Toulouse, France, 2015. [Google Scholar]
- Cuesta, I.; Bielza, C.; Cuenca-Estrella, M.; Larranaga, P.; Rodriguez-Tudela, J.L. Evaluation by data mining techniques of fluconazole breakpoints established by the Clinical and Laboratory Standards Institute (CLSI) and comparison with those of the European Committee on Antimicrobial Susceptibility Testing (EUCAST). Antimicrob. Agents Chemother. 2010, 54, 1541–1546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, S.V.; Peleg, A.Y.; Lodise, T.P., Jr.; Shutt, K.A.; Capitano, B.; Potoski, B.A.; Paterson, D.L. Failure of current cefepime breakpoints to predict clinical outcomes of bacteremia caused by gram-negative organisms. Antimicrob. Agents Chemother. 2007, 51, 4390–4395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abundo, M.E.C.; Ngunjiri, J.M.; Taylor, K.J.M.; Ji, H.; Ghorbani, A.; Kc, M.; Elaish, M.; Jang, H.; Weber, B.; Johnson, T.J.; et al. Evaluation of Sampling Methods for the Study of Avian Respiratory Microbiota. Avian Dis. 2020, 64, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ishfaq, M.; Fan, Q.; Chen, C.; Li, J. A respiratory commensal bacterium acts as a risk factor for Mycoplasma gallisepticum infection in chickens. Vet. Immunol. Immunopathol. 2020, 230, 110127. [Google Scholar] [CrossRef] [PubMed]
- Abundo, M.E.C.; Ngunjiri, J.M.; Taylor, K.J.M.; Ji, H.; Ghorbani, A.; Mahesh, K.C.; Weber, B.P.; Johnson, T.J.; Lee, C.W. Assessment of two DNA extraction kits for profiling poultry respiratory microbiota from multiple sample types. PLoS ONE 2021, 16, e0241732. [Google Scholar] [CrossRef]
- Budden, K.F.; Shukla, S.D.; Rehman, S.F.; Bowerman, K.L.; Keely, S.; Hugenholtz, P.; Armstrong-James, D.P.H.; Adcock, I.M.; Chotirmall, S.H.; Chung, K.F.; et al. Functional effects of the microbiota in chronic respiratory disease. Lancet Respir. Med. 2019, 7, 907–920. [Google Scholar] [CrossRef]
- Mazel, D.; Mobashery, S. Antibiotics as physiological stress inducers and bacterial response to the challenge. Curr. Opin. Microbiol. 2012, 15, 553–554. [Google Scholar] [CrossRef] [Green Version]
- Martínez, J.L.; Rojo, F. Metabolic regulation of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 768–789. [Google Scholar] [CrossRef]
- Poole, K. Bacterial stress responses as determinants of antimicrobial resistance. J. Antimicrob. Chemother. 2012, 67, 2069–2089. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.A.; Hoffmann, C.; Abt, M.C.; Du, Y.; Kobuley, D.; Kirn, T.J.; Bushman, F.D.; Artis, D. Metagenomic analyses reveal antibiotic-induced temporal and spatial changes in intestinal microbiota with associated alterations in immune cell homeostasis. Mucosal. Immunol. 2010, 3, 148–158. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chen, X.P.; Li, J.C.; Ishfaq, M. Gut Microbiota Dysbiosis Aggravates Mycoplasma gallisepticum Colonization in the Chicken Lung. Front. Vet. Sci. 2021, 8, 788811. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence | Amplicon Size |
|---|---|---|
| MG-13R | GAGCTAATCTGTAAAGTTGGTC | 185 bp |
| MG-14F | GCTTCCTTGCGGTTAGCAAC |
| Parameter | Units | Plasma | Lung | ||
|---|---|---|---|---|---|
| Healthy Group | Infected Group | Healthy Group | Infected Group | ||
| Cmax | μg/mL | 0.32 ± 0.00 | 0.28 ± 0.01 | 4.28 ± 0.17 | 4.71 ± 0.29 |
| Tmax | h | 0.82 ± 0.04 | 0.99 ± 0.10 | 1.28 ± 0.03 | 1.30 ± 0.01 |
| α | 1/h | 1.80 ± 0.02 | 1.32 ± 0.03 | 0.75 ± 0.02 | 0.76 ± 0.02 |
| β | 1/h | 0.05 ± 0.02 | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.07 ± 0.01 |
| T1/2α | h | 0.38 ± 0.02 | 0.53 ± 0.13 | 0.92 ± 0.03 | 0.91 ± 0.03 |
| T1/2β | h | 14.55 ± 0.83 | 25.21 ± 0.72 | 13.77 ± 0.43 | 9.77 ± 0.72 |
| CL/F | (mg/kg)/(h·μg/mL) | 0.93 ± 0.08 | 0.82 ± 0.06 | 0.23 ± 0.02 | 0.22 ± 0.05 |
| AUC24 | h·μg/mL | 5.40 ± 0.46 | 6.12 ± 0.24 | 21.97 ± 2.80 | 23.67 ± 2.24 |
| Parameters | Units | Diseased Group |
|---|---|---|
| Emax | LgCCU/mL | 1.85 |
| E0 | LgCCU/mL | −6 |
| EC50 | h | 31.22 |
| N | − | 2.67 |
| Emax − E0 | LgCCU/mL | 7.85 |
| (AUC24h/MIC)exE = 0 | h | 20.09 |
| (AUC24h/MIC)exE = −3 | h | 37.34 |
| (AUC24h/MIC)exE = −4 | h | 46.67 |
| Group | MIC (μg/mL) | Total | Valid | Cure | Effective Rate (%) | Cure Rate (%) | Average Gain | Air Sac Average Damage Score | Air Sac Injury Reduction Rate (%) |
|---|---|---|---|---|---|---|---|---|---|
| Blank group | - | 15 | - | - | - | - | 290.00 ± 68.10 | 0.00 ± 0.00 | 100 |
| M57 Test group Control group | 4 | 15 | 13 | 8 | 86.7 | 53.3 | 185.10 ± 50.20 | 2.57 ± 0.50 | 58 |
| 15 | - | - | - | - | 100.21 ± 28.20 | 2.90 ± 0.80 | 0 | ||
| M19 Test group Control group | 2 | 15 | 12 | 9 | 80 | 60 | 230.10 ± 25.20 | 1.73 ± 0.30 | 62 |
| 15 | - | - | - | - | 97.10 ± 35.20 | 2.87 ± 0.77 | 0 | ||
| M73 Test group Control group | 1 | 15 | 11 | 10 | 73.3 | 66.7 | 255.50 ± 29.22 | 0.71 ± 0.70 | 64 |
| 15 | - | - | - | - | 80.30 ± 19.30 | 3.00 ± 0.52 | 0 | ||
| M24 Test group Control group | 0.25 | 15 | 14 | 14 | 93.3 | 93.3 | 247.20 ± 32.35 | 0.56 ± 0.72 | 79 |
| 15 | - | - | - | - | 59.10 ± 14.00 | 2.93 ± 0.69 | 0 | ||
| M2 Test group Control group | 0.016 | 15 | 14 | 15 | 93.3 | 100 | 268.10 ± 18.23 | 0.53 ± 0.73 | 85 |
| 15 | - | - | - | - | 80.30 ± 16.39 | 3.2 ± 0.93 | 0 |
| Strain | MIC (μg/mL) | Success Treatment | %Success ≤ MIC | %Success > MIC | MaxDiff | AUCSucc | AUCTotal | CAR |
|---|---|---|---|---|---|---|---|---|
| M2 | 0.016 | 15 | 100.00 A | 68.33 B | 31.67 | 0.12 | 0.12 | 1 |
| M24 | 0.25 | 14 | 93.30 | 60.00 | 33.30 C | 3.51 | 3.63 | 0.967 |
| M73 | 1 | 10 | 66.67 | 56.67 | 10.00 | 12.513 | 14.88 | 0.841 |
| M19 | 2 | 9 | 60.00 | 53.33 | 6.67 | 22.013 | 29.88 | 0.737 |
| M57 | 4 | 8 | 53.33 | 53.33 | 0.00 | 39.093 | 59.88 | 0.653 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Huang, A.; Gu, Y.; Li, J.; Huang, L.; Wang, X.; Tao, Y.; Liu, Z.; Wu, C.; Yuan, Z.; et al. Rational Use of Danofloxacin for Treatment of Mycoplasma gallisepticum in Chickens Based on the Clinical Breakpoint and Lung Microbiota Shift. Antibiotics 2022, 11, 403. https://doi.org/10.3390/antibiotics11030403
Wang S, Huang A, Gu Y, Li J, Huang L, Wang X, Tao Y, Liu Z, Wu C, Yuan Z, et al. Rational Use of Danofloxacin for Treatment of Mycoplasma gallisepticum in Chickens Based on the Clinical Breakpoint and Lung Microbiota Shift. Antibiotics. 2022; 11(3):403. https://doi.org/10.3390/antibiotics11030403
Chicago/Turabian StyleWang, Shuge, Anxiong Huang, Yufeng Gu, Jun Li, Lingli Huang, Xu Wang, Yanfei Tao, Zhenli Liu, Congming Wu, Zonghui Yuan, and et al. 2022. "Rational Use of Danofloxacin for Treatment of Mycoplasma gallisepticum in Chickens Based on the Clinical Breakpoint and Lung Microbiota Shift" Antibiotics 11, no. 3: 403. https://doi.org/10.3390/antibiotics11030403