
Protective Effects of Glycyrrhiza Total Flavones on Liver Injury Induced by Streptococcus agalactiae in Tilapia (Oreochromis niloticus)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.1.1. Ethics Statement
2.1.2. Fish
2.1.3. Reagents and Drugs
2.1.4. Resuscitation and Culture of S. agalactiae
2.2. Experimental Design and Parameter Measurement
2.2.1. Test Groups
2.2.2. Sample Collection and Processing
2.2.3. Measurement of Biochemical Indexes
2.2.4. Growth Performance Indicators
2.2.5. Preparation and Observation of Liver Slices
2.2.6. Real-Time Quantitative RT-PCR
2.2.7. Statistical Processing
3. Results
3.1. Effects of GTF on Growth Performance of Tilapia
3.2. Effect of GTF on Liver Morphology of Tilapia
3.3. Effects of GTF on Serum Biochemical Indices of Tilapia
3.4. Effects of GTF on Antioxidant System of Tilapia
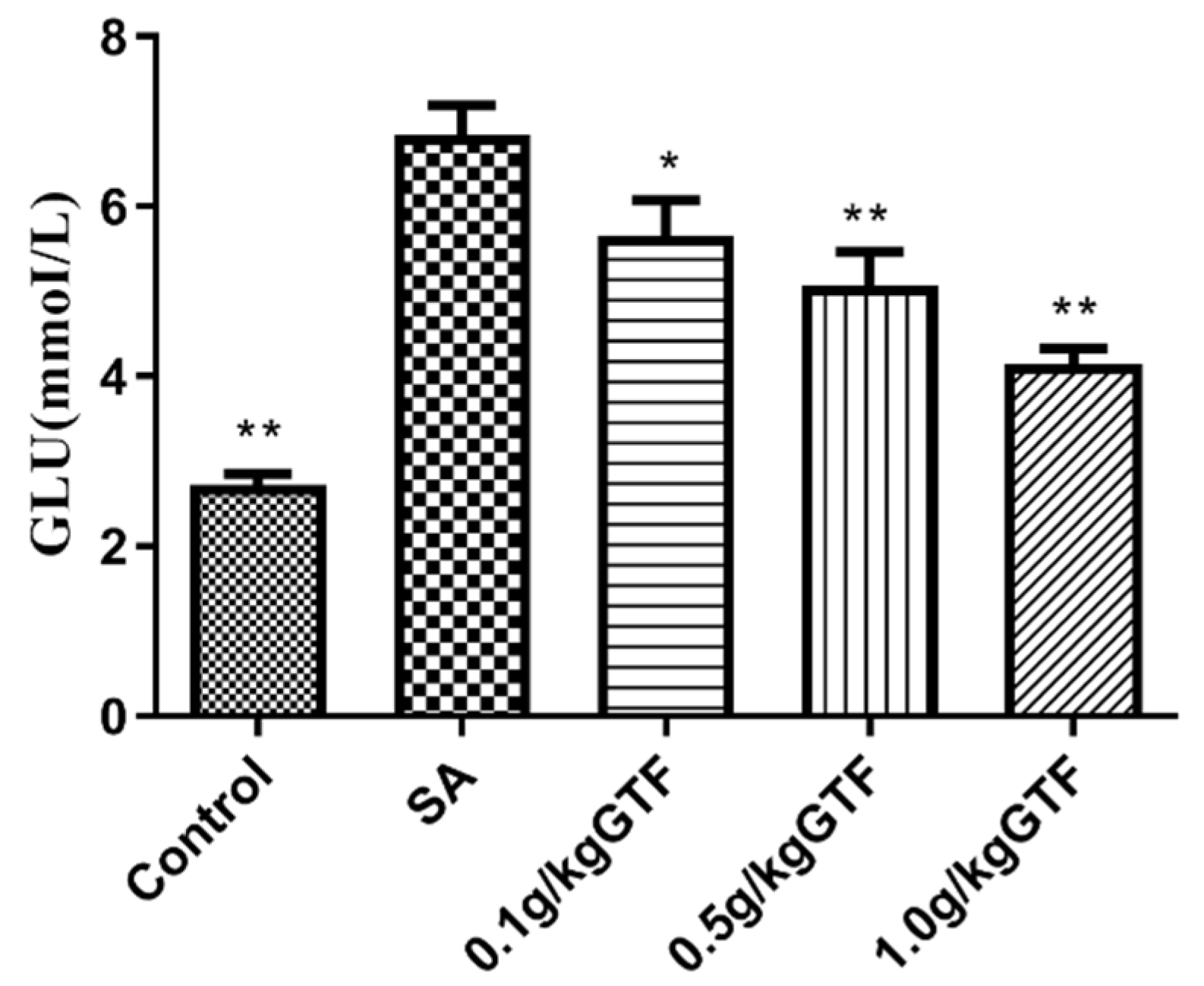
3.5. Effects of GTF on Glucose Metabolism Indexes of Tilapia
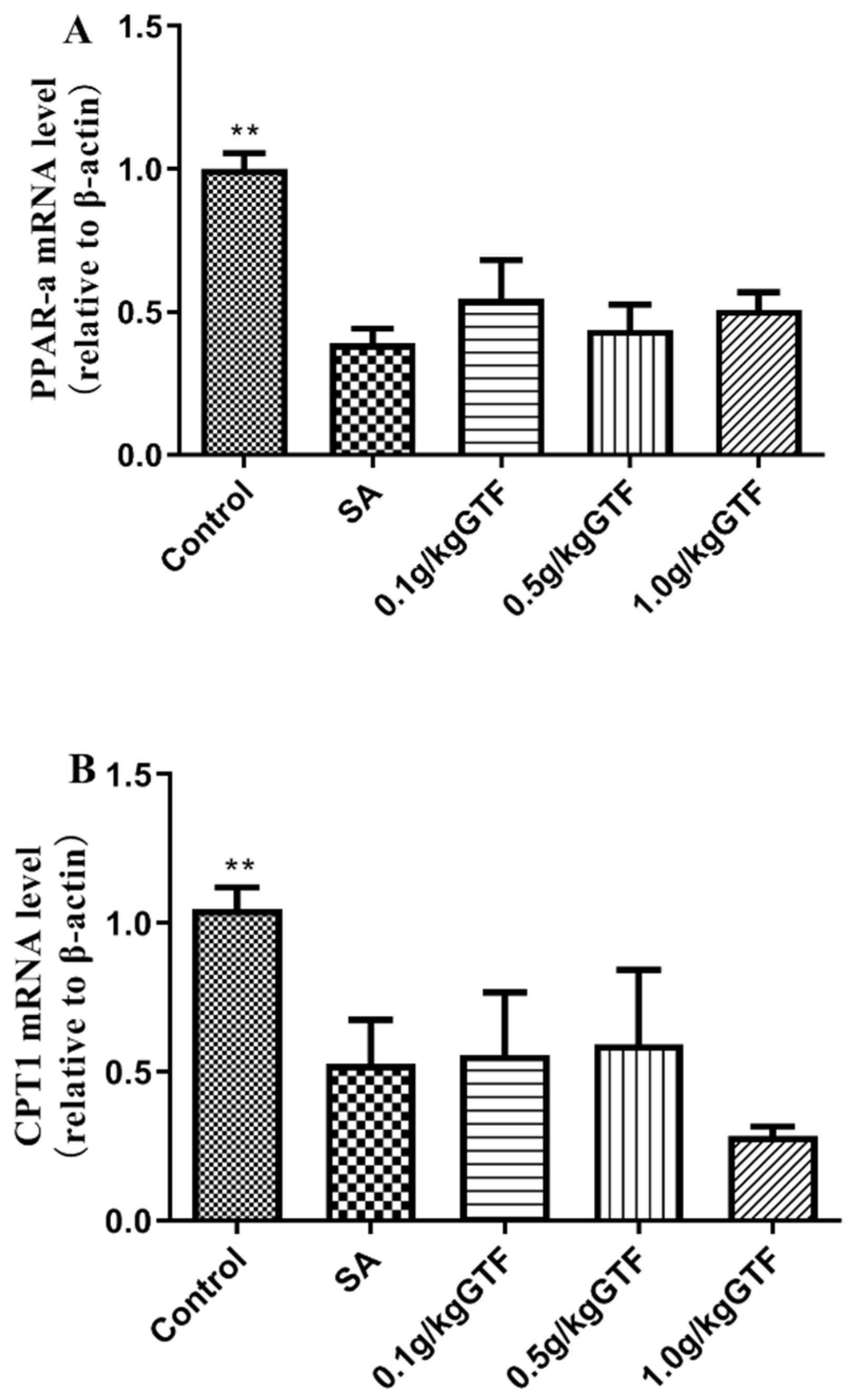
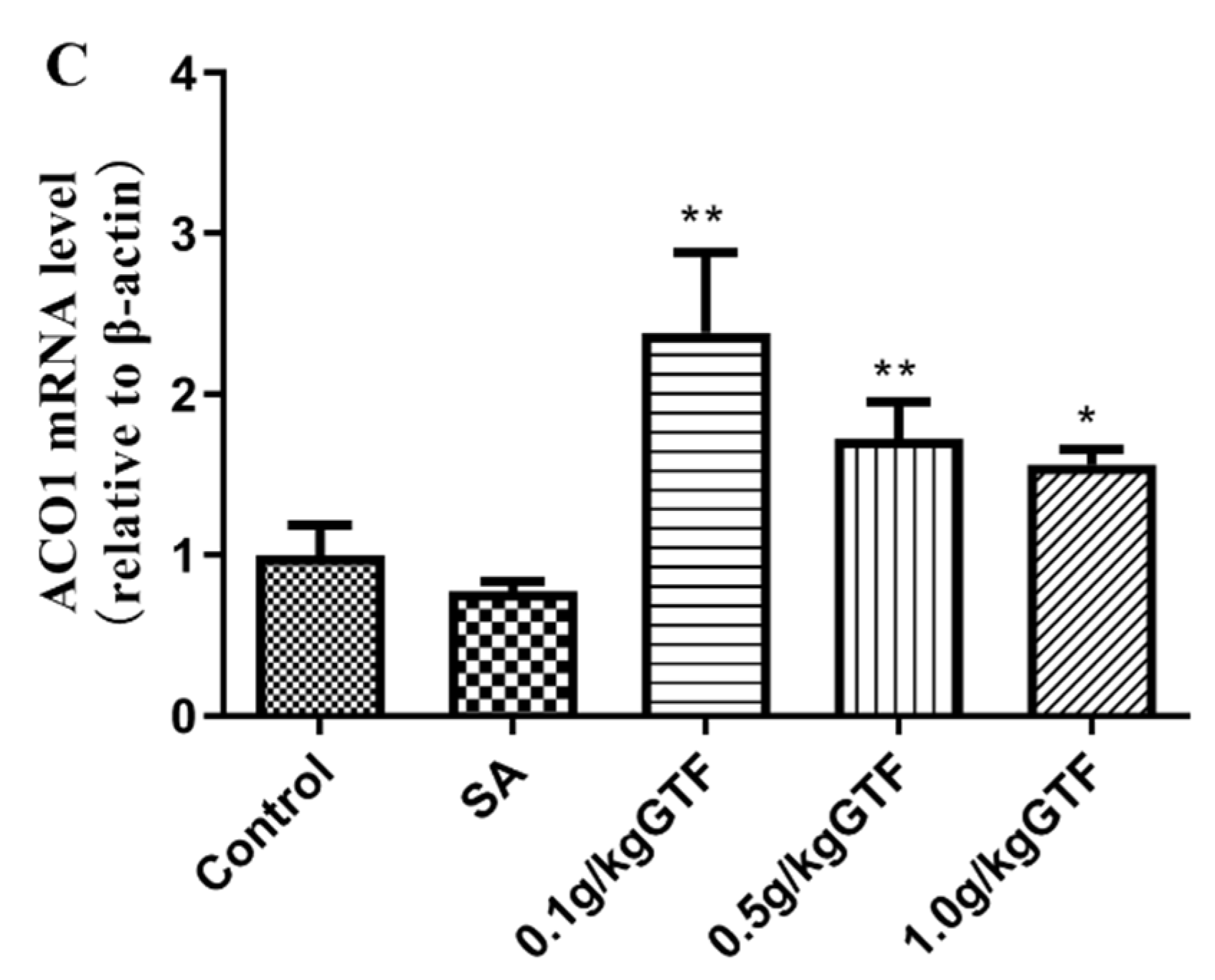
3.6. Effects of GTF on β-Oxidation-Related Gene Rxpression in Tilapia
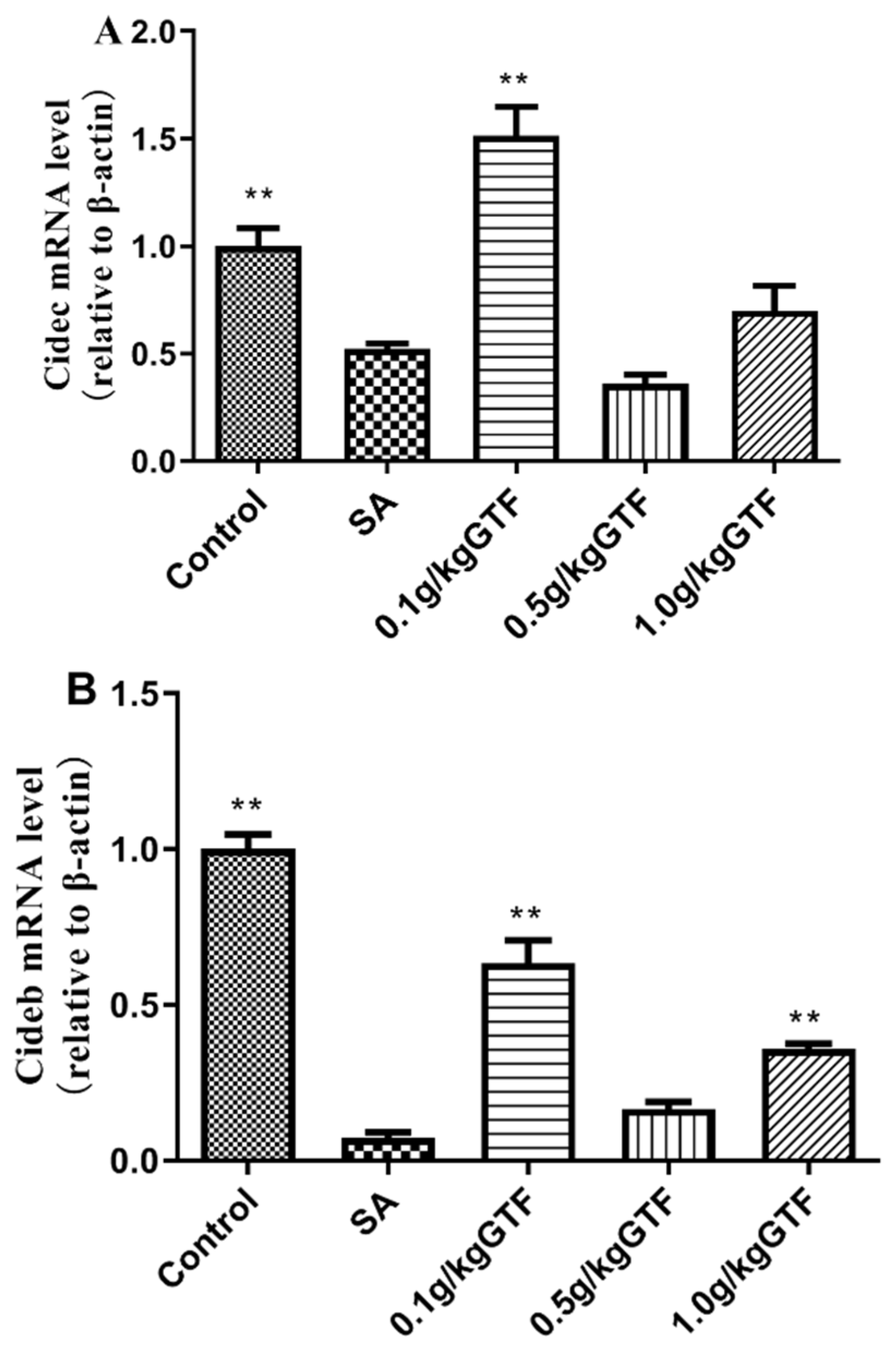
3.7. Effects of GTF on Expression of Genes Related to Lipid Droplet Formation in Tilapia
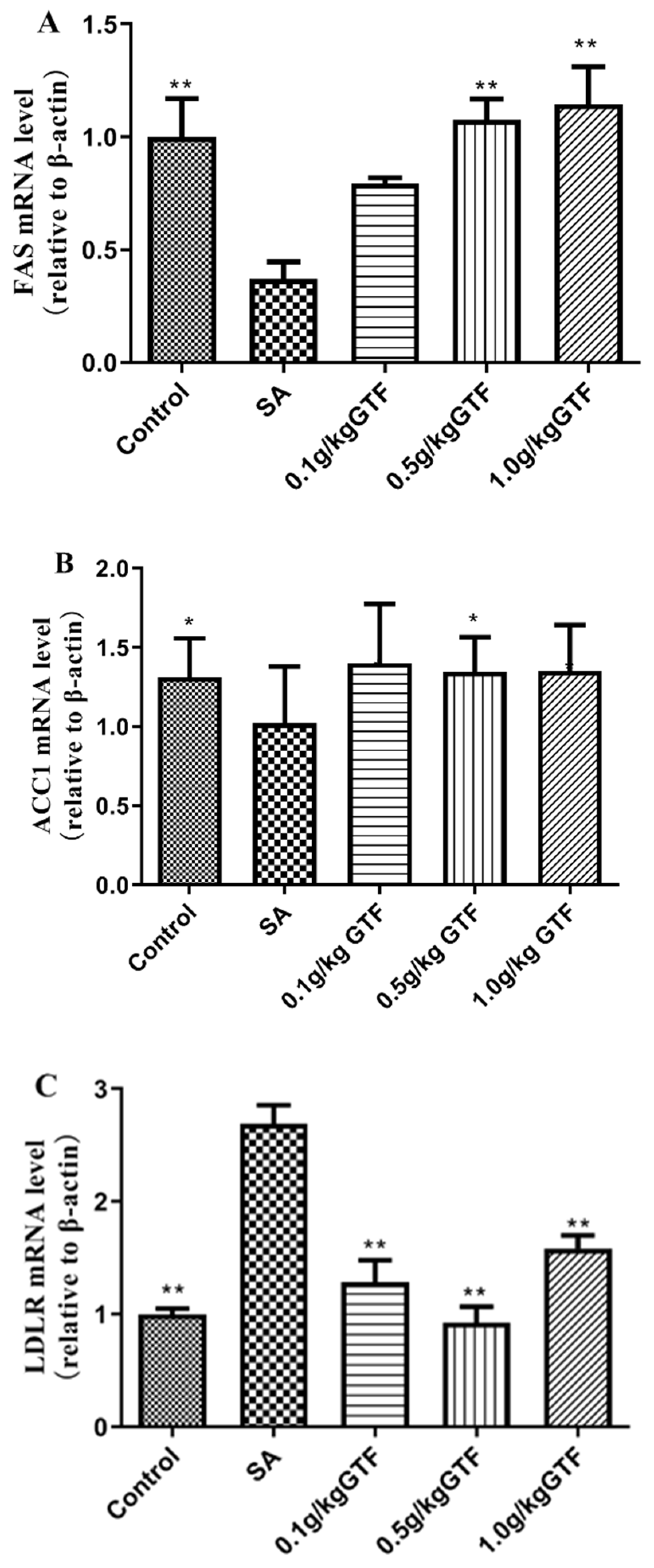
3.8. Effects of GTF on Genes Related to Fatty Acid Formation in Tilapia
3.9. Effects of GTF on the Expression of HSP70 and IgM Genes in Tilapia Liver Tissue
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salman, K.H.; Ali, F.A.Z.; Elhanbaly, R. Effect of cultured white soft cheese on the histopathological changes in the kidneys and liver of albino rats. Sci. Rep. 2022, 12, 2564. [Google Scholar] [CrossRef]
- Medvedeva, N.; Lazaruc, S.C.; Mirzaei, S.; Mikosch, P. Abnormal liver findings as the first manifestation in a patient with brucellosis: Case report. Wien. Med. Wochenschr. 2022, 172, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, H.; Panthee, S. Transcriptome change of Staphylococcus aureus in infected mouse liver. Commun. Biol. 2022, 5, 721. [Google Scholar] [CrossRef]
- Shi, C.; Wang, J.; Zhang, R.; Ishfaq, M.; Li, Y.; Zhang, R.; Si, C.; Li, R.; Li, C.; Liu, F. Dihydromyricetin alleviates Escherichia coli lipopolysaccharide-induced hepatic injury in chickens by inhibiting the NLRP3 inflammasome. Vet. Res. 2022, 53, 6. [Google Scholar] [CrossRef] [PubMed]
- Nur-Nazifah, M.; Sabri, M.Y.; Siti-Zahrah, A. Development and efficacy of feed-based recombinant vaccine encoding the cell wall surface anchor family protein of Streptococcus agalactiae against streptococcosis in Oreochromis sp. Fish Shellfish. Immunol. 2014, 37, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.L.; Li, D.L.; Zhou, Z.Y.; Xiao, W.; Yu, J.; Yang, H.; Xue, L.Y. The physiological response of hybrid tilapia (Oreochromis niloticus ♀ × Oreochromis aures ♂) and its parents infected with Streptococcus agalactiae. Acta Hydrobiol. Sin. 2021, 45, 1190–1200. [Google Scholar] [CrossRef]
- Luo, W.; Gan, X.; Zhu, J.J.; Ao, Q.G.; Tan, Y.; Chen, M.; Luo, Y.J. Pathological research of GIFT tilapia infected by Streptococcus agalactiae. J. South. Agric. 2018, 49, 375–382. [Google Scholar]
- Ao, Q.Z.; Luo, Y.J.; Lv, M.; Zhu, J.J. Effects of Streptococcus agalactiae Infection on Blood and Hepatopancreatic Tissue Biochemical Indices in Different Species of Tilapia. Prog. Fish. Sci. 2020, 41, 167–173. [Google Scholar] [CrossRef]
- Chen, R.R. Isolation, Identification and Pathogenic Biology of Streptococcus agalactiae from Tilapia in Guangzhou. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2012. [Google Scholar]
- Yi, M.M.; Wang, M.; Li, Z.W.; Zhang, D.F.; Liu, Z.G.; Gao, F.Y.; Lu, M.X. The impacts and mechanism of Streptococcus agalactiae on behavior of GIFT Tilapia (Oreochromis niloticus). J. Agric. Biotechnol. 2019, 27, 799–808. [Google Scholar]
- Su, Y.L.; Liu, C.; Deng, Y.Q.; Guo, Z.X.; Feng, J. Research on Streptococcus agalactiae disease in tilapia: A review. J. Dalian Fish. Univ. 2019, 34, 757–766. [Google Scholar] [CrossRef]
- Li, Y.; Wang, B.; Wang, Z.W.; Cai, X.H.; Tang, J.F.; lu, Y.S.; Jian, J.C. Transmission routes of Streptococcus agalactiae and the strategy of immune evasion in tilapia. Guangdong Agric. Sci. 2017, 44, 132–140. [Google Scholar] [CrossRef]
- Fu, Y.; Chen, J.; Li, Y.J.; Zheng, Y.F.; Li, P. Antioxidant and anti-inflammatory activities of six flavonoids separated from licorice. Food Chem. 2013, 141, 1063–1071. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Hua, S.; Ma, J.; Dong, L.; Xu, F. Spectrum-Effect Relationships of Flavonoids in Glycyrrhiza uralensis Fisch. J. Anal. Methods Chem. 2020, 2020, 8838290. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, L.; Hou, J.; Tian, S.; Liu, Y. Molecular mechanisms underlying the anticancer activities of licorice flavonoids. J. Ethnopharmacol. 2021, 267, 113635. [Google Scholar] [CrossRef]
- Wang, C.H.; Wang, H.J. The antibacterial action of five traditional Chinese herbs including radix glycyrrhiza and rhizomacoptidis. Chin. J. Vet. Med. 2014, 50, 44–46. [Google Scholar]
- Liu, Y. Study on Antibacterial Activity of Extracts and Active Constituents of Licorice. Master’s Thesis, Yanbian University, Yanbian, China, 2013. [Google Scholar]
- Zhou, H.; Dai, C.; Cui, X.; Zhang, T.; Che, Y.; Duan, K.; Yi, L.; Nguyen, A.D.; Li, N.; De Souza, C.; et al. Immunomodulatory and antioxidant effects of Glycyrrhiza uralensis polysaccharide in Lohmann Brown chickens. Front. Vet. Sci. 2022, 9, 959449. [Google Scholar] [CrossRef]
- Chang, H.J.; Kim, Y.H.; Kang, Y.H.; Choi, M.H.; Lee, J.H. Antioxidant and antibacterial effects of medicinal plants and their stick-type medicinal concentrated beverages. Food Sci. Biotechnol. 2020, 29, 1413–1423. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Wang, W.; Zhang, Y.; Yang, Z.; Jin, Y.; Ge, H.M.; Li, E.; Yang, G. Glycyrrhizic acid as the antiviral component of Glycyrrhiza uralensis Fisch. against coxsackievirus A16 and enterovirus 71 of hand foot and mouth disease. J. Ethnopharmacol. 2013, 147, 114–121. [Google Scholar] [CrossRef]
- Cao, L.P.; Du, J.L.; Jia, R.; Ding, W.D.; Xu, P.; Yin, G.J.; Zhang, T. Effects of cyclophosphamide on antioxidative and immune functions of Nile tilapia (Oreochromis niloticus) via the TLR-NF-κB signaling pathway. Aquat. Toxicol. 2021, 239, 105956. [Google Scholar] [CrossRef]
- Zhang, C.Y.; Cao, L.P.; Du, J.L.; Jia, R.; Gu, Z.Y.; Yin, G.J.; Jeney, G. Nrf2 mediate protective effects of curcumin against carbon tetrachloride (CCl4 )-induced liver injury in Carassius auratus. Jiangsu J. Agr. Sci. 2018, 34, 114–121. [Google Scholar] [CrossRef]
- Cao, L.; Du, J.; Nie, Z.; Jia, R.; Yin, G.; Xu, P.; Ding, W.; Xu, G. Alteration of endoplasmic reticulum stress, inflammation and anti-oxidative status in cyclophosphamide-damaged liver of Nile tilapia (Oreochromis niloticus). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2022, 254, 109271. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, R.; Marino, F.; Salvaggio, A.; Capparucci, F.; Di Caro, G.; Iaria, C.; Salvo, A.; Rotondo, A.; Tibullo, D.; Guerriero, G.; et al. Evaluation of Chronic Nanosilver Toxicity to Adult Zebrafish. Front. Physiol. 2017, 8, 1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rairat, T.; Chi, Y.; Hsieh, C.Y.; Liu, Y.K.; Chuchird, N.; Chou, C.C. Determination of Optimal Doses and Minimum Effective Concentrations of Tricaine Methanesulfonate, 2-Phenoxyethanol and Eugenol for Laboratory Managements in Nile Tilapia (Oreochromis niloticus). Animals 2021, 11, 1521. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Gu, Z.; He, Q.; Du, J.; Cao, L.; Jeney, G.; Xu, P.; Yin, G. Anti-oxidative, anti-inflammatory and hepatoprotective effects of Radix Bupleuri extract against oxidative damage in tilapia (Oreochromis niloticus) via Nrf2 and TLRs signaling pathway. Fish Shellfish. Immunol. 2019, 93, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lu, Y.J.; Lai, D.G.; Tian, X.J.; Yu, H. Isolation, identification and drug sensitivity test of Tilapia Streptococcus iniae. Guangdong Agric. Sci. 2014, 41, 110–113. [Google Scholar] [CrossRef]
- Huang, Y.H.; Ma, S.; Han, S.Y.; Huang, W.D.; Wei, M.L.; Meng, L.L.; Lv, X.L.; Tan, H.M.; Li, D.Z.; Liang, J.Z.; et al. Antibiogram types of Streptococcus agalactiae strains isolated from Oreochromis spp. in Guangxi during 2011–2016. J. South. Agric. 2018, 49, 1023–1031. [Google Scholar] [CrossRef]
- Zhou, M.; Yu, S.; Hong, B.; Li, J.; Han, H.; Qie, G. Antibiotics control in aquaculture requires more than antibiotic-free feeds: A tilapia farming case. Environ. Pollut. 2021, 268, 115854. [Google Scholar] [CrossRef]
- Batool, R.; Khan, M.R.; Majid, M. Euphorbia dracunculoides L. abrogates carbon tetrachloride induced liver and DNA damage in rats. BMC Complement. Altern. Med. 2017, 17, 223. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yang, L.; Han, Y.; Wu, Z.; Chen, P.; Zhang, H.; Zhou, J. Protective effects of hepatocyte-specific glycyrrhetic derivatives against carbon tetrachloride-induced liver damage in mice. Bioorg. Chem. 2017, 72, 42–50. [Google Scholar] [CrossRef]
- Qiang, J.; Yang, H.; Wang, H.; Xu, P.; Qi, S.L.; He, J. Studies on blood biochemical indices and expression of hepatic HSP70 mRNA of different tilapia strains artificially challenged with Streptococcus iniae. J. Fish. China 2012, 36, 958–968. [Google Scholar] [CrossRef]
- Rajput, S.A.; Liang, S.J.; Wang, X.Q.; Yan, H.C. Lycopene Protects Intestinal Epithelium from Deoxynivalenol-Induced Oxidative Damage via Regulating Keap1/Nrf2 Signaling. Antioxidants 2021, 10, 1493. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.H.; Long, C.G.; Wang, X.M.; Li, Q.R.; Zheng, Q.X.; Hu, C.L. The scavenging effect of flavoniods of glycyrrhiza on hydroxyl radical studied in vitro. J. North Sichuan Med. Coll. 2001, 16, 3–5. [Google Scholar] [CrossRef]
- Gou, S.H.; He, M.; Li, B.B.; Zhu, N.Y.; Ni, J.M. Hepatoprotective effect of total flavonoids from Glycyrrhiza uralensis Fisch in liver injury mice. Nat. Prod. Res. 2021, 35, 6083–6087. [Google Scholar] [CrossRef]
- Du, J.L.; Cao, L.P.; Jia, R.; Gu, Z.Y.; He, Q.; Xu, P.; Jeney, G.; Ma, Y.Z.; Yin, G.J. Protective effects of Glycyrrhiza total flavones on liver injury of tilapia (Oreochromis niloticus) under high fat condition. Acta Agriculturae Zhejiangensis 2021, 33, 1826–1835. [Google Scholar] [CrossRef]
- Liu, J.F.; Guo, X.F. Effects of Glycyrrhiza flavone on the activity of GSH-Px and T-AOC in mice induced by ethanol oxidation. Heilongjiang Anim. Sci. Vet. Med. 2015, 176–178. [Google Scholar] [CrossRef]
- Li, M.H.; Tang, J.; Wang, C.L. Protective Effect of Glycyrrhizic Flavone on Myocardial Injury Induced by Long-term Exhaustive Exercised Rats. Genom. Appl. Biol. 2015, 34, 290–295. [Google Scholar] [CrossRef]
- Zhao, Q.; Tang, P.; Zhang, T.; Huang, J.F.; Xiao, X.R.; Zhu, W.F.; Gonzalez, F.J.; Li, F. Celastrol ameliorates acute liver injury through modulation of PPARα. Biochem. Pharmacol. 2020, 178, 114058. [Google Scholar] [CrossRef]
- Xu, W.J. PPARa involved in lipid metabolism pathway and nonalcoholic fatty liver disease. Chin. Hepatol. 2020, 25, 894–896. [Google Scholar] [CrossRef]
- Li, Y.J.; Hou, W.; Chen, W.H.; Long, Y.F.; Wu, J.Z. Effects of Anthraquinone Glycosides from Juemingzion the Expression of Fatty Acid β-Oxidation Related Enzyme Protein in Rats with Nonalcoholic Fatty Liver Disease. West. J. Tradit. Chin. Med. 2021, 34, 6–10. [Google Scholar] [CrossRef]
- Misheva, M.; Kotzamanis, K. Oxylipin metabolism is controlled by mitochondrial β-oxidation during bacterial inflammation. Nat. Commun. 2022, 13, 139. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Xu, M.; Qin, M.; Zhang, D.; Wu, S.; Liu, X.; Zhang, Y. Study on the role and mechanism of lncRNA in the remodeling of atrial energy metabolism in rabbits with atrial fibrillation based on nano sensor technology. Bioengineered 2022, 13, 863–875. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.L.; Luo, M. Research progress of carnitine palmitoyl transferase-I. Int. J. Endocrinol. Metab. 2002, 22, 166–169. [Google Scholar] [CrossRef]
- Liao, W.Y.; Xu, D. Study on the improvement effect of Ginsenoside Rg1 on nonalcoholic fatty liver phenotype by regulation of β-oxidation. Chongqing Med. 2016, 45, 1179–1182. [Google Scholar] [CrossRef]
- Li, P.; Zhang, R.; Wang, M.; Chen, Y.; Chen, Z.; Ke, X.; Zuo, L.; Wang, J. Baicalein Prevents Fructose-Induced Hepatic Steatosis in Rats: In the Regulation of Fatty Acid De Novo Synthesis, Fatty Acid Elongation and Fatty Acid Oxidation. Front Pharmacol. 2022, 13, 917329. [Google Scholar] [CrossRef]
- Chen, F.J.; Yin, Y.; Chua, B.T.; Li, P. CIDE family proteins control lipid homeostasis and the development of metabolic diseases. Traffic 2020, 21, 94–105. [Google Scholar] [CrossRef]
- Xu, W.; Wu, L.; Yu, M.; Chen, F.J.; Arshad, M.; Xia, X.; Ren, H.; Yu, J.; Xu, L.; Xu, D.; et al. Differential Roles of Cell Death-inducing DNA Fragmentation Factor-α-like Effector (CIDE) Proteins in Promoting Lipid Droplet Fusion and Growth in Subpopulations of Hepatocytes. J. Biol. Chem. 2016, 291, 4282–4293. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Li, J.Z.; Liu, Y.; Li, X.; Yang, T.; Ma, X.; Li, Q.; Yao, Z.; Li, P. Cideb, an ER- and lipid droplet-associated protein, mediates VLDL lipidation and maturation by interacting with apolipoprotein B. Cell Metab. 2009, 9, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Guo, K.; Toh, S.Y.; Zhou, Z.; Li, P. Mitochondria localization and dimerization are required for CIDE-B to induce apoptosis. J. Biol. Chem. 2000, 275, 22619–22622. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Kang, H.; Chu, Y.; Jin, Y.; Zhang, L.; Yang, R.; Zhang, Z.; Zhao, S.; Zhou, L. Cidec differentially regulates lipid deposition and secretion through two tissue-specific isoforms. Gene 2018, 641, 265–271. [Google Scholar] [CrossRef]
- Zheng, G.S.; Tan, Y.M.; Shang, Y.Y.; Liu, Y.P.; Hu, B.A.; Wang, D.; Han, L.; Wang, Z.H.; Zhang, W.; Ti, Y.; et al. CIDEC silencing attenuates diabetic nephropathy via inhibiting apoptosis and promoting autophagy. J. Diabetes Investig. 2021, 12, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Cao, L.P.; Du, J.L.; He, Q.; Yin, G.J. Effects of High-Fat Diet on Steatosis, Endoplasmic Reticulum Stress and Autophagy in Liver of Tilapia (Oreochromis niloticus). Front. Mar. Sci. 2020, 7, 363. [Google Scholar] [CrossRef]
- Zheng, G.D.; Yang, L.C.; Lin, L.Z.; Zhang, Q.F. Effects of caffeine and catechins combination on gene and protein expression levels of hepatic fatty acid synthase in mice. J. Chin. Inst. Food Sci. Technol. 2016, 16, 21–25. [Google Scholar] [CrossRef]
- Brownsey, R.W.; Boone, A.N.; Elliott, J.E.; Kulpa, J.E.; Lee, W.M. Regulation of acetyl-CoA carboxylase. Biochem. Soc. Trans. 2006, 34, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.Y.; Huang, H.L.; Li, J.; Cao, M.Y.; Zhang, Y.; Zhao, Y.Y.; Xue, L.Y. Effects of fasting on the mRNA levels and enzymatic activities of hormone sensitive lipase and acetyl-CoA carboxylase in the large yellow croaker Larimichthys crocea. Acta Oceanol. Sin. 2018, 40, 31–39. [Google Scholar] [CrossRef]
- Ye, Q.; Lei, H.; Zheng, W.B.; Zheng, S.Z.; Chen, Y.X. Macrophages converted into foam cells via LPS-induced LDLr: Another important pathway of foam-cell formation under inflammatory stress. Chin. Pharmacol. Bull. 2013, 29, 1064–1070. [Google Scholar] [CrossRef]
- Tashiro, S.; Miyake, H.; Rokutan, K. Role of geranylgeranylacetone as non-toxic HSP70 inducer in liver surgery: Clinical application. J. Hepato-Biliary-Pancreat. Sci. 2018, 25, 269–274. [Google Scholar] [CrossRef]
- Liu, X.Z.; Li, S.Q.; Ren, J.T.; Xi, S.M.; Xu, Z.S.; Sun, Y.J. Expression and significance of HSP70, IL-6 and TNF-α in the tissue of acute liver enjury induced by CCl 4. Shandong Med. J. 2011, 51, 17–19. [Google Scholar]
- Ma, Y.J.; Li, S.Q. Expression of HSP70 in Acetaminophen Induced Acute Liver Injury Tissue. J. Henan Univ. Sci. Technol. (Med. Sci.) 2012, 30, 8–9. [Google Scholar] [CrossRef]
- Liu, Y.J.; Du, J.L.; Jia, R.; Cao, L.P.; Wang, J.H.; Yin, G.J. Hepatoprotective effects of Lycium barbarum polysaccharides on carbon tetrachloride induced primary hepatocyte injury in Jian Carp. J. Shanghai Ocean. Univ. 2014, 23, 718–725. [Google Scholar]
- Guo, X.Y.; Yin, Y.W.; Mao, A.T.; Dai, J.; Zhang, L.; Zhang, P.J.; Li, Y.H. Effects of lead exposure on antioxidation and immunity of Carassius aurous. Chin. Vet. Sci. 2018, 48, 222–227. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Gene | Primer Sequence (5′–3′) | GenBank Number/References |
|---|---|---|---|
| Fatty acid β-oxidation | PPAR-α | F: CTGATAAAGCTTCGGGCTTCCA | NM_001290066.1 |
| CPT-1 | R: CGCTCACACTTATCATACTCCAGCT F: TTTCCAGGCCTCCTTACCCA | XM_013268638.3 | |
| R: TGTACTGCTCATTGTCCAGCAGA | |||
| ACO-1 | F: GGTCAAAGGCAACAATCAGGAG | NM_001290199.1 | |
| R: GACTCTGCCAAAGGCAACCA | |||
| Lipid droplet formation | Cideb | F: CTACCCGACAACGTCATGCT | XM_013266878.3 |
| R: CCTTGAAATGTGGCCTGCAC | |||
| Cidec | F: TGGAGCCCACATCCTTACAA | XM_005450237.4 | |
| R: TTTTTGGCAGCATAACAGCG | |||
| Fatty acid synthesis | ACC1 | F: GCGGTGTTCGGCTTGTTTTT | XM_025910662.1 |
| R: CAAGTCCACCTTCCCTTGGT | |||
| FAS | F: TTTGAGATGTGCTCACAGCTGCA | XM_003454056.5 | |
| R: TCAGCCAGTGAGCTGTGGATGAT | |||
| Fatty acid uptake | LDLR | F: TACGGCTTACCAGTCCTCCA | XM_003443172.5 |
| R: AGGTGACTGGAGCTTGTGTG | |||
| Detoxification | HSP70 | F:ATTTCAGACGGAGGGAAGCC | XM_019357557.1 |
| R:CAGCGTTGGACACCTTTTGG | |||
| Immune | IgM | F: ACCGAATCGAAAAATGCGGC | KJ676389.1 |
| R: AACACAACCAGGACATTGGTTC | |||
| Internal reference | β-actin | F:CCTGAGCGTAAATACTCCGTCTG | KJ126772.1 |
| R:AAGCACTTGCGGTGGACGAT |
| Index | Dietary GTF Level/(g/kg) | |||
|---|---|---|---|---|
| 0 | 0.1 | 0.5 | 1.0 | |
| Initial weight/g | 20.00 ± 0.57 | 19.33 ± 0.88 | 20.67 ± 0.33 | 19.67 ± 0.67 |
| Final weight/g | 80.19 ± 1.79 | 82.34 ± 2.12 | 85.16 ± 2.05 | 85.90 ± 2.59 |
| WGR, % | 3.02 ± 0.21 | 3.29 ± 0.30 | 3.12 ± 0.05 | 3.38 ± 0.20 |
| SGR/(%/d) | 1.00 ± 0.04 | 1.05 ± 0.05 | 1.07 ± 0.03 | 1.10 ± 0.04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, J.; Cao, L.; Gao, J.; Jia, R.; Zhu, H.; Nie, Z.; Xi, B.; Yin, G.; Ma, Y.; Xu, G. Protective Effects of Glycyrrhiza Total Flavones on Liver Injury Induced by Streptococcus agalactiae in Tilapia (Oreochromis niloticus). Antibiotics 2022, 11, 1648. https://doi.org/10.3390/antibiotics11111648
Du J, Cao L, Gao J, Jia R, Zhu H, Nie Z, Xi B, Yin G, Ma Y, Xu G. Protective Effects of Glycyrrhiza Total Flavones on Liver Injury Induced by Streptococcus agalactiae in Tilapia (Oreochromis niloticus). Antibiotics. 2022; 11(11):1648. https://doi.org/10.3390/antibiotics11111648
Chicago/Turabian StyleDu, Jinliang, Liping Cao, Jiancao Gao, Rui Jia, Haojun Zhu, Zhijuan Nie, Bingwen Xi, Guojun Yin, Yuzhong Ma, and Gangchun Xu. 2022. "Protective Effects of Glycyrrhiza Total Flavones on Liver Injury Induced by Streptococcus agalactiae in Tilapia (Oreochromis niloticus)" Antibiotics 11, no. 11: 1648. https://doi.org/10.3390/antibiotics11111648