
Concordance between Antimicrobial Resistance Phenotype and Genotype of Staphylococcus pseudintermedius from Healthy Dogs
 , , , and
, , , and
Abstract
:1. Introduction
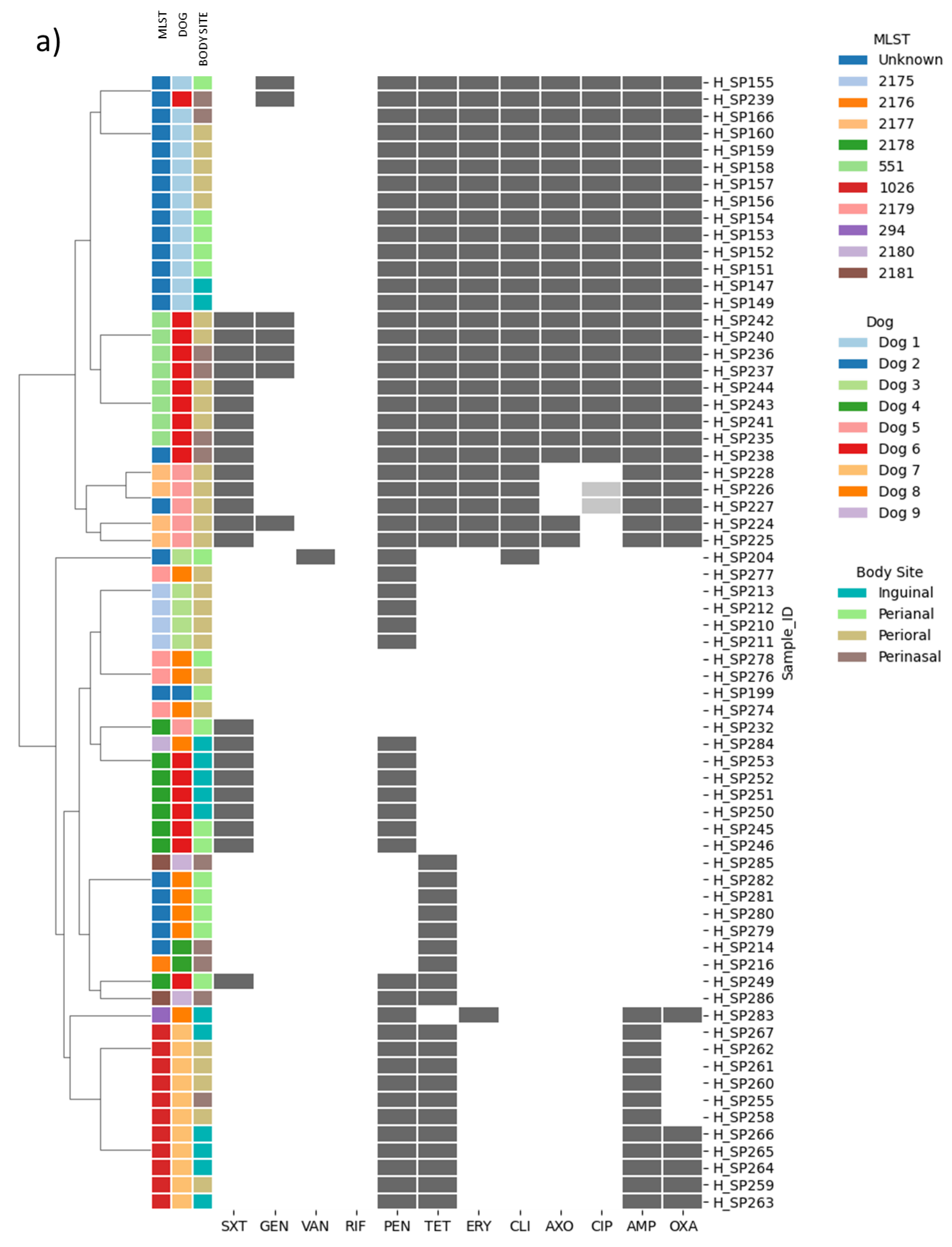
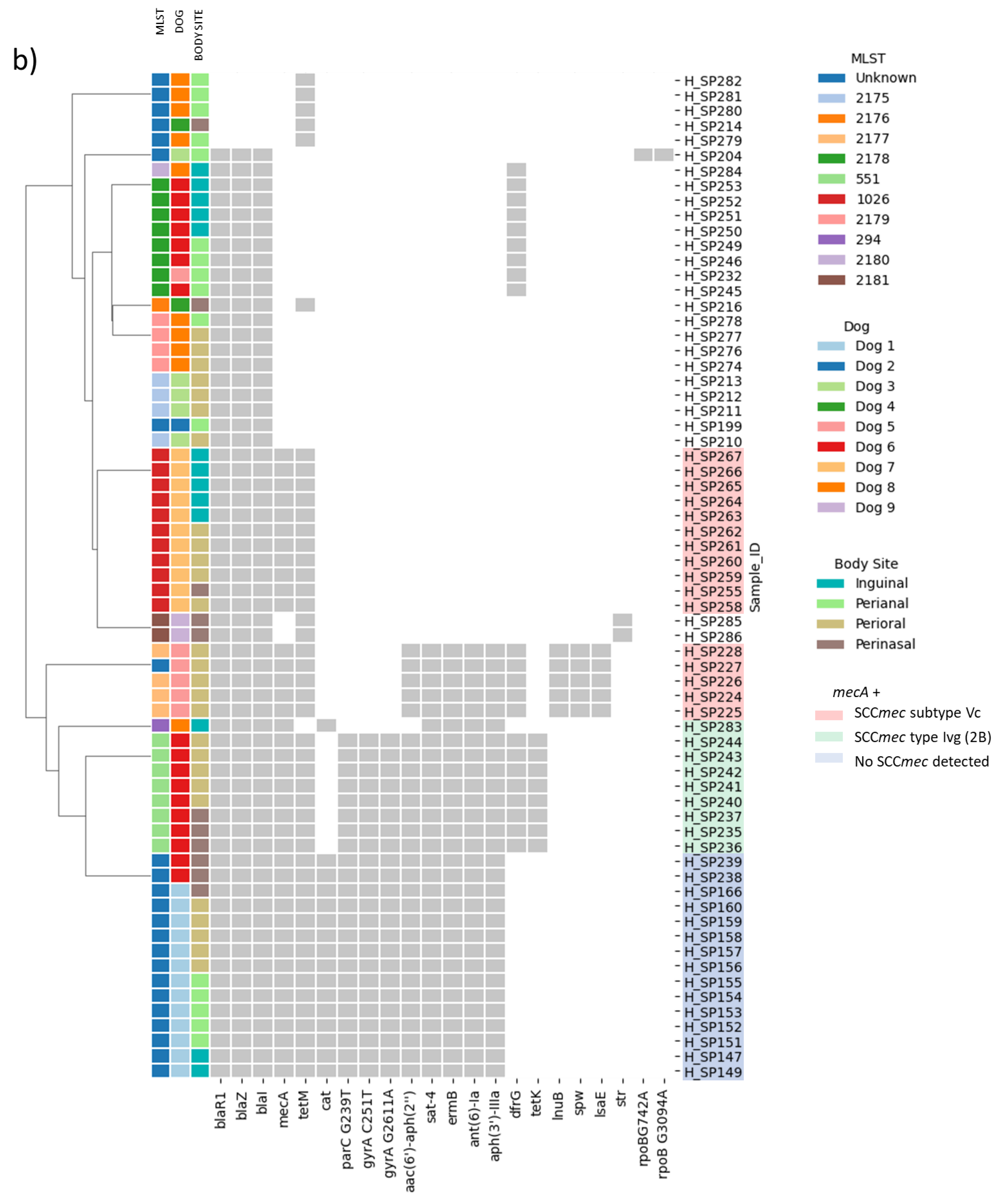
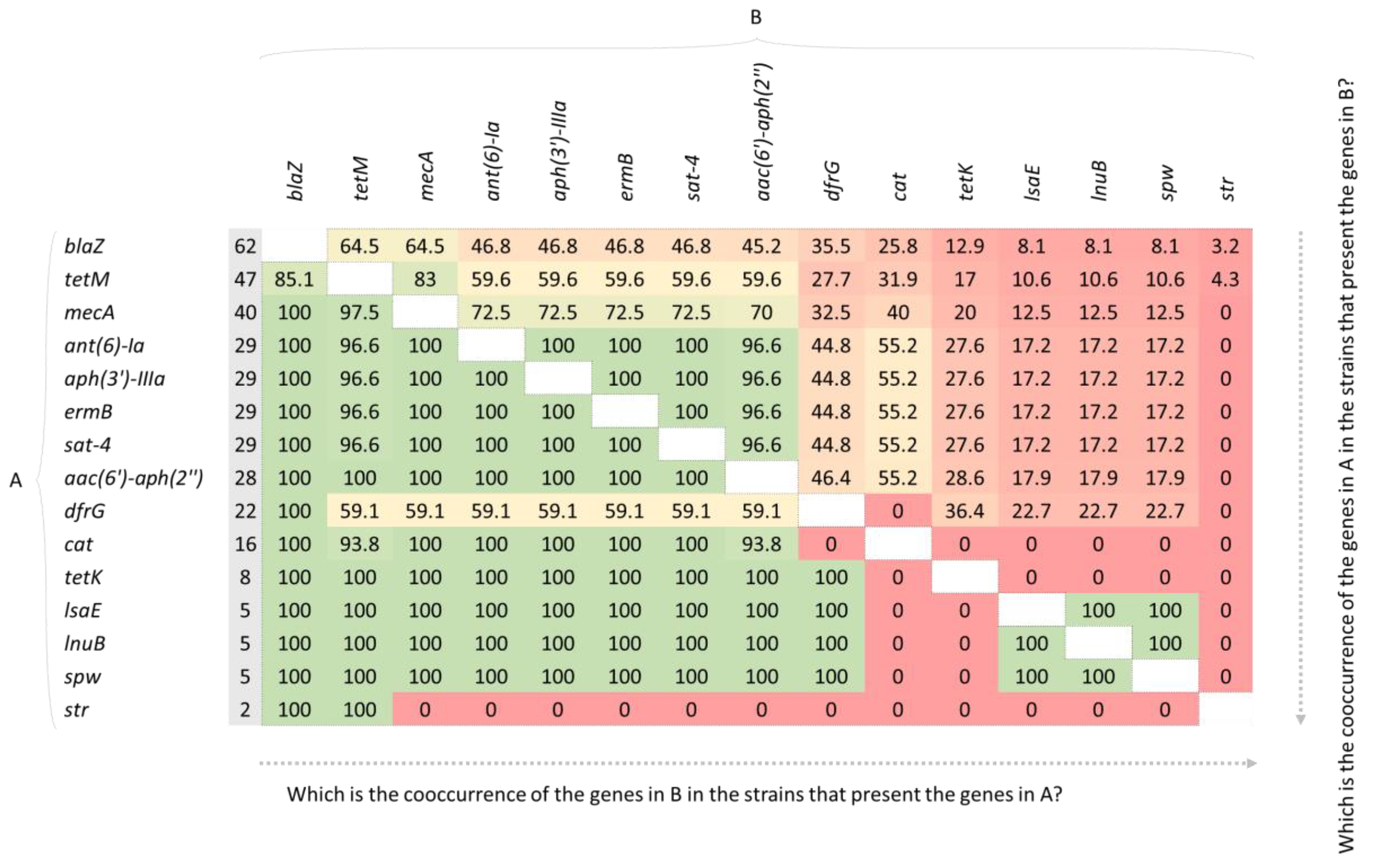
2. Results
2.1. Distribution of MRSP and MDR Strains
2.2. Genotype and Phenotype Concordance Varied Depending on the Antibiotic Family
2.3. Beta-Lactams and Cephalosporin Resistance
2.4. Fluoroquinolone Resistance
2.5. Vancomycin Resistance
2.6. Macrolide and Lincosamide Resistance
2.7. Tetracycline Resistance
2.8. Trimethoprim-Sulfamethoxazole (Cotrimoxazole) Resistance
2.9. Aminoglycoside Resistance
2.10. Genes Not Tested Phenotypically: sat-4 and cat
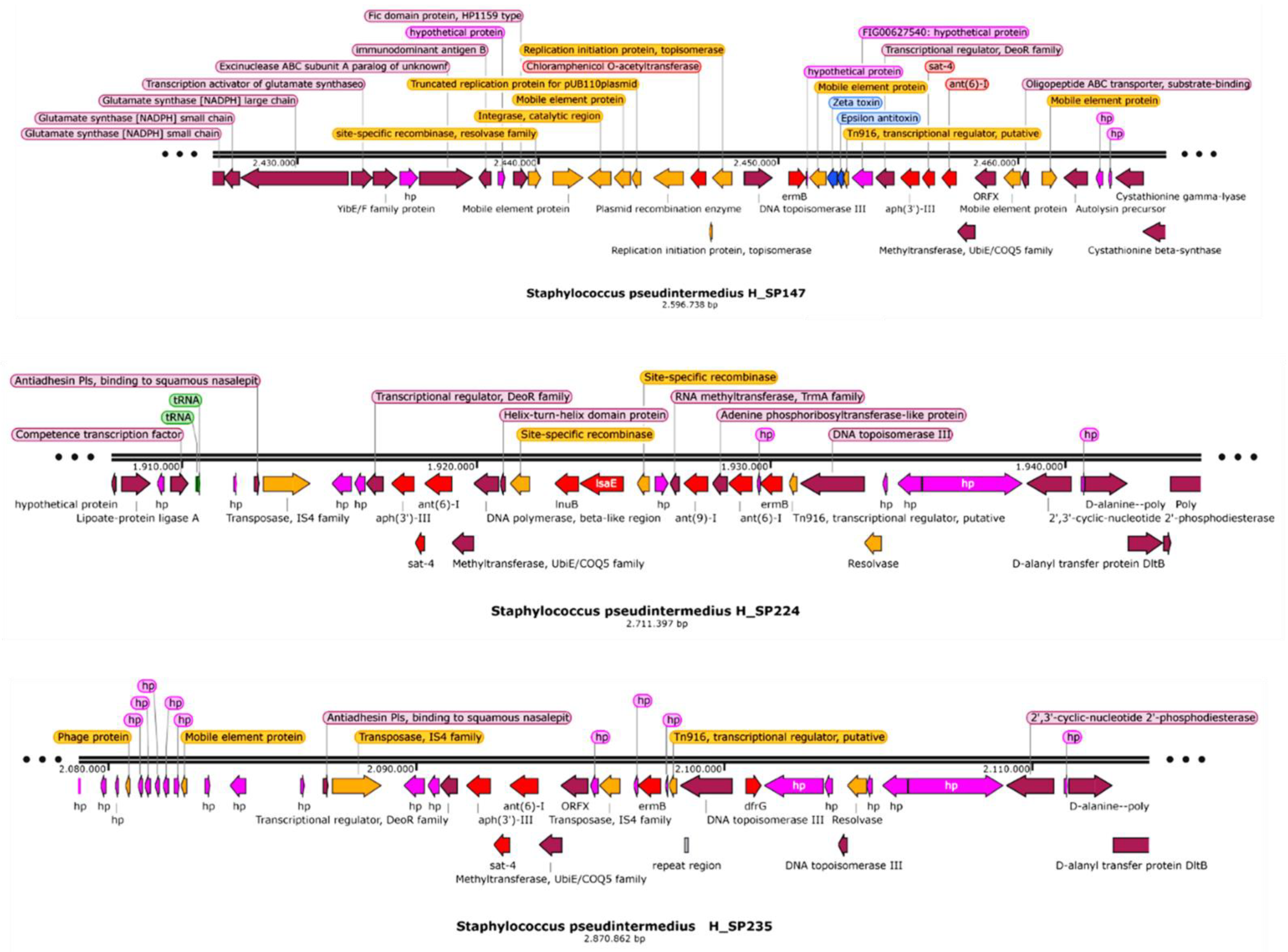
3. Discussion
4. Materials and Methods
4.1. Bacterial Cultures
4.2. Antibiotic Susceptibility
4.3. DNA Extraction and Nanopore Sequencing Library Preparation
4.4. Bioinformatic Genome Sequencing Analysis
4.5. Antibiotic Resistance Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bannoehr, J.; Guardabassi, L. Staphylococcus pseudintermedius in the dog: Taxonomy, diagnostics, ecology, epidemiology and pathogenicity. Vet. Dermatol. 2012, 23, 253-e52. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.C.; Bärgman, S.C.; Moodley, A.; Nielsen, S.S.; Guardabassi, L. Staphylococcus pseudintermedius colonization patterns and strain diversity in healthy dogs: A cross-sectional and longitudinal study. Vet. Microbiol. 2012, 160, 420–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubin, J.E.; Chirino-Trejo, M. Prevalence, Sites of Colonization, and Antimicrobial Resistance Among Staphylococcus pseudintermedius Isolated from Healthy Dogs in Saskatoon, Canada. J. Vet. Diagn. Investig. 2011, 23, 351–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Somayaji, R.; Priyantha, M.A.R.; Rubin, J.E.; Church, D. Human infections due to Staphylococcus pseudintermedius, an emerging zoonosis of canine origin: Report of 24 cases. Diagn. Microbiol. Infect. Dis. 2016, 85, 471–476. [Google Scholar] [CrossRef]
- Hackmann, C.; Gastmeier, P.; Schwarz, S.; Lübke-Becker, A.; Bischoff, P.; Leistner, R. Pet husbandry as a risk factor for colonization or infection with MDR organisms: A systematic meta-analysis. J. Antimicrob. Chemother. 2021, 76, 1392–1405. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortázar, C.; Herskin, M.; Michel, V.; et al. Assessment of listing and categorisation of animal diseases within the framework of the Animal Health Law (Regulation (EU) No 2016/429): Antimicrobial-resistant Staphylococcus pseudintermedius in dogs and cats. EFSA J. 2022, 20, e07080. [Google Scholar] [CrossRef]
- Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar Schmidt, C.; Herskin, M.; Michel, V.; et al. Assessment of animal diseases caused by bacteria resistant to antimicrobials: Dogs and cats. EFSA J. 2021, 19, e06680. [Google Scholar] [CrossRef]
- Bannoehr, J.; Ben Zakour, N.L.; Waller, A.S.; Guardabassi, L.; Thoday, K.L.; Van Den Broek, A.H.M.; Fitzgerald, J.R. Population Genetic Structure of the Staphylococcus intermedius Group: Insights into agr Diversification and the Emergence of Methicillin-Resistant Strains. J. Bacteriol. 2007, 189, 8685–8692. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, F.A.; White, D.G.; West, S.E.H.; Walker, R.D.; DeBoer, D.J. Molecular characterization of Staphylococcus intermedius carriage by healthy dogs and comparison of antimicrobial susceptibility patterns to isolates from dogs with pyoderma. Vet. Microbiol. 2005, 108, 119–131. [Google Scholar] [CrossRef]
- Marí-Almirall, M.; Cosgaya, C.; Pitart, C.; Viñes, J.; Muñoz, L.; Campo, I.; Cuscó, A.; Rodríguez-Serna, L.; Santana, G.; Del Río, A.; et al. Dissemination of NDM-producing Klebsiella pneumoniae and Escherichia coli high-risk clones in Catalan healthcare institutions. J. Antimicrob. Chemother. 2020, 76, 345–354. [Google Scholar] [CrossRef]
- Viñes, J.; Cuscó, A.; Napp, S.; Alvarez, J.; Saez-Llorente, J.L.; Rosàs-Rodoreda, M.; Francino, O.; Migura-Garcia, L. Transmission of Similar Mcr-1 Carrying Plasmids among Different Escherichia coli Lineages Isolated from Livestock and the Farmer. Antibiotics 2021, 10, 313. [Google Scholar] [CrossRef]
- Descloux, S.; Rossano, A.; Perreten, V. Characterization of new staphylococcal cassette chromosome mec (SCCmec) and topoisomerase genes in fluoroquinolone- and methicillin-resistant Staphylococcus pseudintermedius. J. Clin. Microbiol. 2008, 46, 1818–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, J.; Evers, S.; Bischoff, K.; Carlier, C.; Courvalin, P. Characterization of the sat 4 gene encoding a streptothricin acetyltransferase in Campylobacter coli BE/G4. FEMS Microbiol. Lett. 1994, 120, 13–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fàbregas, N.; Pérez, D.; Viñes, J.; Fonticoba, R.; Cuscó, A.; Migura-García, L.; Ferrer, L.; Francino, O. Whole-Genome Sequencing and De Novo Assembly of 67 Staphylococcus pseudintermedius Strains Isolated from the Skin of Healthy Dogs. Microbiol. Resour. Announc. 2022, 11, e00039-22. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.H.; Ceric, O.; Guag, J.; Nemser, S.; Borenstein, S.; Slavic, D.; Lippert, S.; McDowell, R.; Krishnamurthy, A.; Korosec, S.; et al. Genomics accurately predicts antimicrobial resistance in Staphylococcus pseudintermedius collected as part of Vet-LIRN resistance monitoring. Vet. Microbiol. 2021, 254, 109006. [Google Scholar] [CrossRef]
- Wegener, A.; Broens, E.M.; Zomer, A.; Spaninks, M.; Wagenaar, J.A.; Duim, B. Comparative genomics of phenotypic antimicrobial resistances in methicillin-resistant Staphylococcus pseudintermedius of canine origin. Vet. Microbiol. 2018, 225, 125–131. [Google Scholar] [CrossRef]
- Walther, B.; Hermes, J.; Cuny, C.; Wieler, L.H.; Vincze, S.; Abou Elnaga, Y.; Stamm, I.; Kopp, P.A.; Kohn, B.; Witte, W.; et al. Sharing More than Friendship—Nasal Colonization with Coagulase-Positive Staphylococci (CPS) and Co-Habitation Aspects of Dogs and Their Owners. PLoS ONE 2012, 7, e35197. [Google Scholar] [CrossRef]
- Ferrer, L.; García-Fonticoba, R.; Pérez, D.; Viñes, J.; Fàbregas, N.; Madroñero, S.; Meroni, G.; Martino, P.A.; Martínez, S.; Maté, M.L.; et al. Whole genome sequencing and de novo assembly of Staphylococcus pseudintermedius: A pangenome approach to unravelling pathogenesis of canine pyoderma. Vet. Dermatol. 2021, 32, 654–663. [Google Scholar] [CrossRef]
- Worthing, K.A.; Schwendener, S.; Perreten, V.; Saputra, S.; Coombs, G.W.; Pang, S.; Davies, M.R.; Abraham, S.; Trott, D.J.; Norris, J.M. Characterization of Staphylococcal Cassette Chromosome mec Elements from Methicillin-Resistant Staphylococcus pseudintermedius Infections in Australian Animals. mSphere 2018, 3, e00491-18. [Google Scholar] [CrossRef] [Green Version]
- Foglia Manzillo, V.; Paola Nocera, F.; Gizzarelli, M.; Oliva, G. A Successful Vancomycin Treatment Of Multidrug-resistant MRSA-associated Canine Pyoderma. J. Dermatol. Res. Ther. 2015, 1, 19–25. [Google Scholar] [CrossRef]
- Aizawa, J.; de Souza-Filho, A.F.; Cortez, A.; Vasconcelos, C.G.C.; Biotto, J.; Heinemann, M.B. Staphylococcus pseudintermedius multirresistente isolado do cão: Relato de caso. Braz. J. Vet. Res. Anim. Sci. 2017, 54, 430–433. [Google Scholar] [CrossRef] [Green Version]
- Novick, R.P.; Morse, S.I. In vivo transmission of drug resistance factors between strains of Staphylococcus aureus. J. Exp. Med. 1967, 125, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Jiang, L.; Han, W.; Xie, X.; Jin, Y.; He, X.; Wu, R. Novel Mutation Sites in the Development of Vancomycin- Intermediate Resistance in Staphylococcus aureus. Front. Microbiol. 2017, 7, 2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoji, M.; Cui, L.; Iizuka, R.; Komoto, A.; Neoh, H.; Watanabe, Y.; Hishinuma, T.; Hiramatsu, K. walK and clpP Mutations Confer Reduced Vancomycin Susceptibility in Staphylococcus aureus. Antimicrob. Agents Chemother. 2011, 55, 3870–3881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, Y.; Cui, L.; Katayama, Y.; Kozue, K.; Hiramatsu, K. Impact of rpoB Mutations on Reduced Vancomycin Susceptibility in Staphylococcus aureus. J. Clin. Microbiol. 2011, 49, 2680–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, L.; Ma, X.; Sato, K.; Okuma, K.; Tenover, F.C.; Mamizuka, E.M.; Gemmell, C.G.; Kim, M.-N.; Ploy, M.-C.; El Solh, N.; et al. Cell Wall Thickening Is a Common Feature of Vancomycin Resistance in Staphylococcus aureus. J. Clin. Microbiol. 2003, 41, 5–14. [Google Scholar] [CrossRef] [Green Version]
- Vitali, L.A.; Daniela, B.; Massimo, B.; Dezemona, P. Draft genome of an extremely-drug resistant st551 Staphylococcus pseudintermedius from an Italian dog with otitis externa. J. Glob. Antimicrob. Resist. 2021, 25, 107–109. [Google Scholar] [CrossRef]
- Loiacono, M.; Martino, P.A.; Albonico, F.; Dell’Orco, F.; Ferretti, M.; Zanzani, S.; Mortarino, M. High-resolution melting analysis of gyrA codon 84 and grlA codon 80 mutations conferring resistance to fluoroquinolones in Staphylococcus pseudintermedius isolates from canine clinical samples. J. Vet. Diagn. Investig. 2017, 29, 711–715. [Google Scholar] [CrossRef] [Green Version]
- Sanfilippo, C.M.; Hesje, C.K.; Haas, W.; Morris, T.W. Topoisomerase Mutations That Are Associated with High-Level Resistance to Earlier Fluoroquinolones in Staphylococcus aureus Have Less Effect on the Antibacterial Activity of Besifloxacin. Chemotherapy 2011, 57, 363–371. [Google Scholar] [CrossRef]
- Yamada, M.; Yoshida, J.; Hatou, S.; Yoshida, T.; Minagawa, Y. Mutations in the quinolone resistance determining region in Staphylococcus epidermidis recovered from conjunctiva and their association with susceptibility to various fluoroquinolones. Br. J. Ophthalmol. 2008, 92, 848–851. [Google Scholar] [CrossRef]
- Gold, R.M.; Lawhon, S.D. Incidence of Inducible Clindamycin Resistance in Staphylococcus pseudintermedius from Dogs. J. Clin. Microbiol. 2013, 51, 4196–4199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.-M.; Wang, J.; Tao, X.-X.; Jia, H.-B.; Meng, F.-L.; Yang, H.; You, Y.-H.; Zheng, B.; Hu, Y.; Bu, X.-X.; et al. A Conjugative MDR pMG1-Like Plasmid Carrying the lsa(E) Gene of Enterococcus faecium with Potential Transmission to Staphylococcus aureus. Front. Microbiol. 2021, 12, 667415. [Google Scholar] [CrossRef] [PubMed]
- Berbel, D.; Càmara, J.; García, E.; Tubau, F.; Guérin, F.; Giard, J.-C.; Domínguez, M.Á.; Cattoir, V.; Ardanuy, C. A novel genomic island harbouring lsa(E) and lnu(B) genes and a defective prophage in a Streptococcus pyogenes isolate resistant to lincosamide, streptogramin A and pleuromutilin antibiotics. Int. J. Antimicrob. Agents 2019, 54, 647–651. [Google Scholar] [CrossRef] [PubMed]
- Phumthanakorn, N.; Schwendener, S.; Donà, V.; Chanchaithong, P.; Perreten, V.; Prapasarakul, N. Genomic insights into methicillin-resistant Staphylococcus pseudintermedius isolates from dogs and humans of the same sequence types reveals diversity in prophages and pathogenicity islands. PLoS ONE 2021, 16, e0254382. [Google Scholar] [CrossRef] [PubMed]
- Wegener, A.; Broens, E.M.; van der Graaf-van Bloois, L.; Zomer, A.L.; Visser, C.E.; van Zeijl, J.; van der Meer, C.; Kusters, J.G.; Friedrich, A.W.; Kampinga, G.A.; et al. Absence of Host-Specific Genes in Canine and Human Staphylococcus pseudintermedius as Inferred from Comparative Genomics. Antibiotics 2021, 10, 854. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Torres, C.; Lozano, C.; Sáenz, Y.; Zarazaga, M. Detection and characterization of methicillin-resistant Staphylococcus pseudintermedius in healthy dogs in La Rioja, Spain. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 447–453. [Google Scholar] [CrossRef]
- Kang, J.-H.; Hwang, C.-Y. First detection of multiresistance pRE25-like elements from Enterococcus spp. in Staphylococcus pseudintermedius isolated from canine pyoderma. J. Glob. Antimicrob. Resist. 2020, 20, 304–308. [Google Scholar] [CrossRef]
- Loeffler, A.; Lloyd, D.H. What has changed in canine pyoderma? A narrative review. Vet. J. 2018, 235, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Viñes, J.; Cuscó, A.; Francino, O. Hybrid assembly from a pathogenic Methicillin and Multi-Drug Resistant Staphylococcus pseudintermedius isolated from a canine otitis, Spain. Microbiol. Resour. Announc. 2019, 9, e01121-19. [Google Scholar]
- Khan, S.A.; Novick, R.P. Complete nucleotide sequence of pT181, a tetracycline-resistance plasmid from Staphylococcus aureus. Plasmid 1983, 10, 251–259. [Google Scholar] [CrossRef]
- Côrtes, M.F.; Botelho, A.M.N.; Almeida, L.G.P.; Souza, R.C.; Cunha, O.d.L.; Nicolás, M.F.; Vasconcelos, A.T.R.; Figueiredo, A.M.S. Community-acquired methicillin-resistant Staphylococcus aureus from ST1 lineage harboring a new SCCmec IV subtype (SCCmec IVm) containing the tetK gene. Infect. Drug Resist. 2018, 11, 2583–2592. [Google Scholar] [CrossRef] [PubMed]
- Dale, G.E.; Broger, C.; D’Arcy, A.; Hartman, P.G.; DeHoogt, R.; Jolidon, S.; Kompis, I.; Labhardt, A.M.; Langen, H.; Locher, H.; et al. A single amino acid substitution in Staphylococcus aureus dihydrofolate reductase determines trimethoprim resistance. J. Mol. Biol. 1997, 266, 23–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fowler, P.W.; Cole, K.; Gordon, N.C.; Kearns, A.M.; Llewelyn, M.J.; Peto, T.E.A.; Crook, D.W.; Walker, A.S. Robust Prediction of Resistance to Trimethoprim in Staphylococcus aureus. Cell Chem. Biol. 2018, 25, 339–349.e4. [Google Scholar] [CrossRef] [Green Version]
- Griffith, E.C.; Wallace, M.J.; Wu, Y.; Kumar, G.; Gajewski, S.; Jackson, P.; Phelps, G.A.; Zheng, Z.; Rock, C.O.; Lee, R.E.; et al. The Structural and Functional Basis for Recurring Sulfa Drug Resistance Mutations in Staphylococcus aureus Dihydropteroate Synthase. Front. Microbiol. 2018, 9, 1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues Hoffmann, A.; Patterson, A.P.; Diesel, A.; Lawhon, S.D.; Ly, H.J.; Stephenson, C.E.; Mansell, J.; Steiner, J.M.; Dowd, S.E.; Olivry, T.; et al. The Skin Microbiome in Healthy and Allergic Dogs. PLoS ONE 2014, 9, e83197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuscó Martí, A.; Sanchez Bonastre, A.; Altet, L.; Ferrer, L.; Francino, O. Individual Signatures Define Canine Skin Microbiota Composition and Variability. Front. Vet. Sci. 2017, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- CLSI Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Disk and Dilution Susceptibility Tests for Bacteria Isolated from Animals, 4th ed.; CLSI Supplement VET08; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- CLSI Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 28th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Schwarz, S.; Silley, P.; Simjee, S.; Woodford, N.; van Duijkeren, E.; Johnson, A.P.; Gaastra, W. Assessing the antimicrobial susceptibility of bacteria obtained from animals. Vet. Microbiol. 2010, 141, 1–4. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of neural network basecalling tools for Oxford Nanopore sequencing. Genome Biol. 2019, 20, 129. [Google Scholar] [CrossRef] [Green Version]
- De Coster, W.; D’Hert, S.; Schultz, D.T.; Cruts, M.; Van Broeckhoven, C. NanoPack: Visualizing and processing long-read sequencing data. Bioinformatics 2018, 34, 2666–2669. [Google Scholar] [CrossRef] [Green Version]
- Juul, S.; Izquierdo, F.; Hurst, A.; Dai, X.; Wright, A.; Kulesha, E.; Pettett, R.; Turner, D. What’s in my pot? Real-time species identification on the MinION. bioRxiv 2015, 030742. [Google Scholar] [CrossRef] [Green Version]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef] [PubMed]
- Oxford Nanopore Technologies Medaka. Available online: https://github.com/nanoporetech/medaka (accessed on 21 October 2022).
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, M.; De Silva, N.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: Automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total-genome-sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Waskom, M. Seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Seemann, T. Abricate. Available online: https://github.com/tseemann/abricate (accessed on 21 October 2022).
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding data and analysis capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dog | Inguinal | Perianal | Perioral | Nasal | Total |
|---|---|---|---|---|---|
| 1 | 2 | 5 | 5 | 1 | 13 |
| 2 | 0 | 1 | 0 | 0 | 1 |
| 3 | 0 | 1 | 4 | 0 | 5 |
| 4 | 0 | 0 | 0 | 2 | 2 |
| 5 | 0 | 1 | 5 | 0 | 6 |
| 6 | 4 | 3 | 5 | 5 | 17 |
| 7 | 5 | 0 | 5 | 1 | 11 |
| 8 | 2 | 5 | 3 | 0 | 10 |
| 9 | 0 | 0 | 0 | 2 | 2 |
| Total | 13 | 16 | 27 | 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viñes, J.; Fàbregas, N.; Pérez, D.; Cuscó, A.; Fonticoba, R.; Francino, O.; Ferrer, L.; Migura-Garcia, L. Concordance between Antimicrobial Resistance Phenotype and Genotype of Staphylococcus pseudintermedius from Healthy Dogs. Antibiotics 2022, 11, 1625. https://doi.org/10.3390/antibiotics11111625
Viñes J, Fàbregas N, Pérez D, Cuscó A, Fonticoba R, Francino O, Ferrer L, Migura-Garcia L. Concordance between Antimicrobial Resistance Phenotype and Genotype of Staphylococcus pseudintermedius from Healthy Dogs. Antibiotics. 2022; 11(11):1625. https://doi.org/10.3390/antibiotics11111625
Chicago/Turabian StyleViñes, Joaquim, Norma Fàbregas, Daniel Pérez, Anna Cuscó, Rocío Fonticoba, Olga Francino, Lluís Ferrer, and Lourdes Migura-Garcia. 2022. "Concordance between Antimicrobial Resistance Phenotype and Genotype of Staphylococcus pseudintermedius from Healthy Dogs" Antibiotics 11, no. 11: 1625. https://doi.org/10.3390/antibiotics11111625