

Prevalence and Genetic Analysis of Resistance Mechanisms of Linezolid-Nonsusceptible Enterococci in a Tertiary Care Hospital Examined via Whole-Genome Sequencing
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Evolution of Linezolid Nonsusceptibility in Severance Hospital over the Past Five Years
2.2. Linezolid Minimum Inhibitory Concentration (MIC) Distribution and Resistance Mechanisms
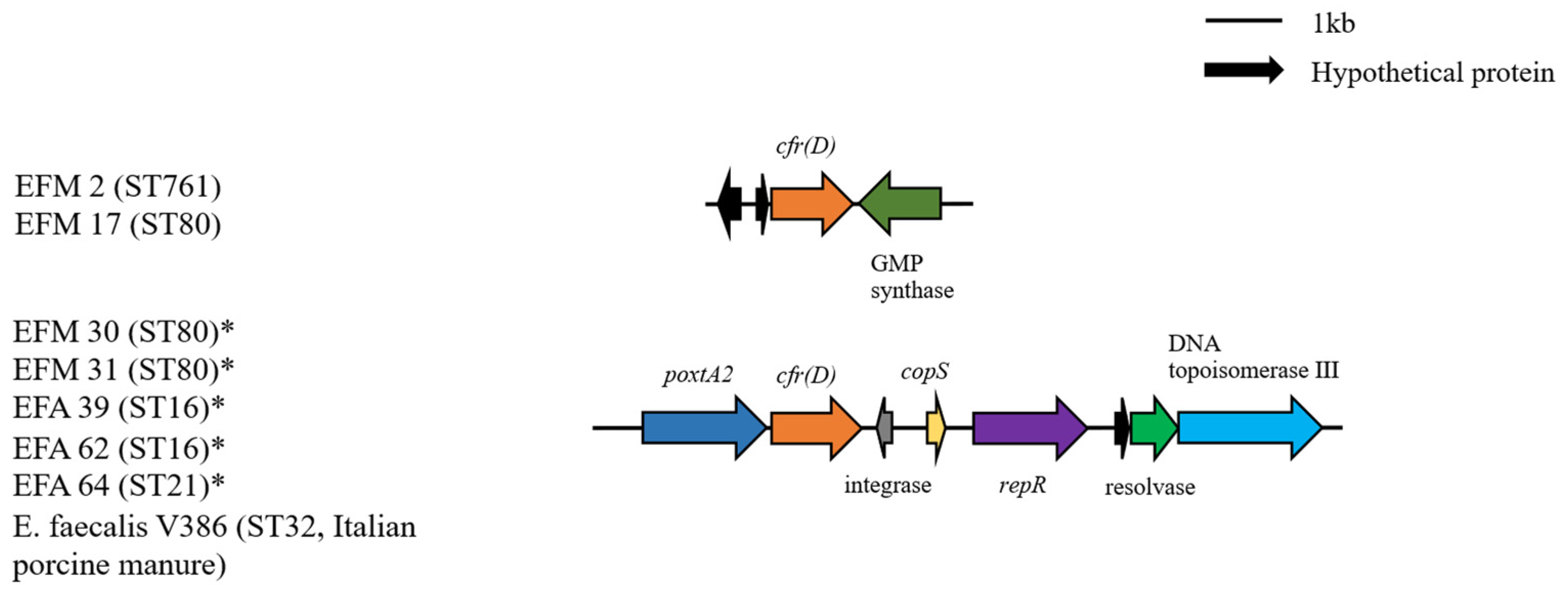
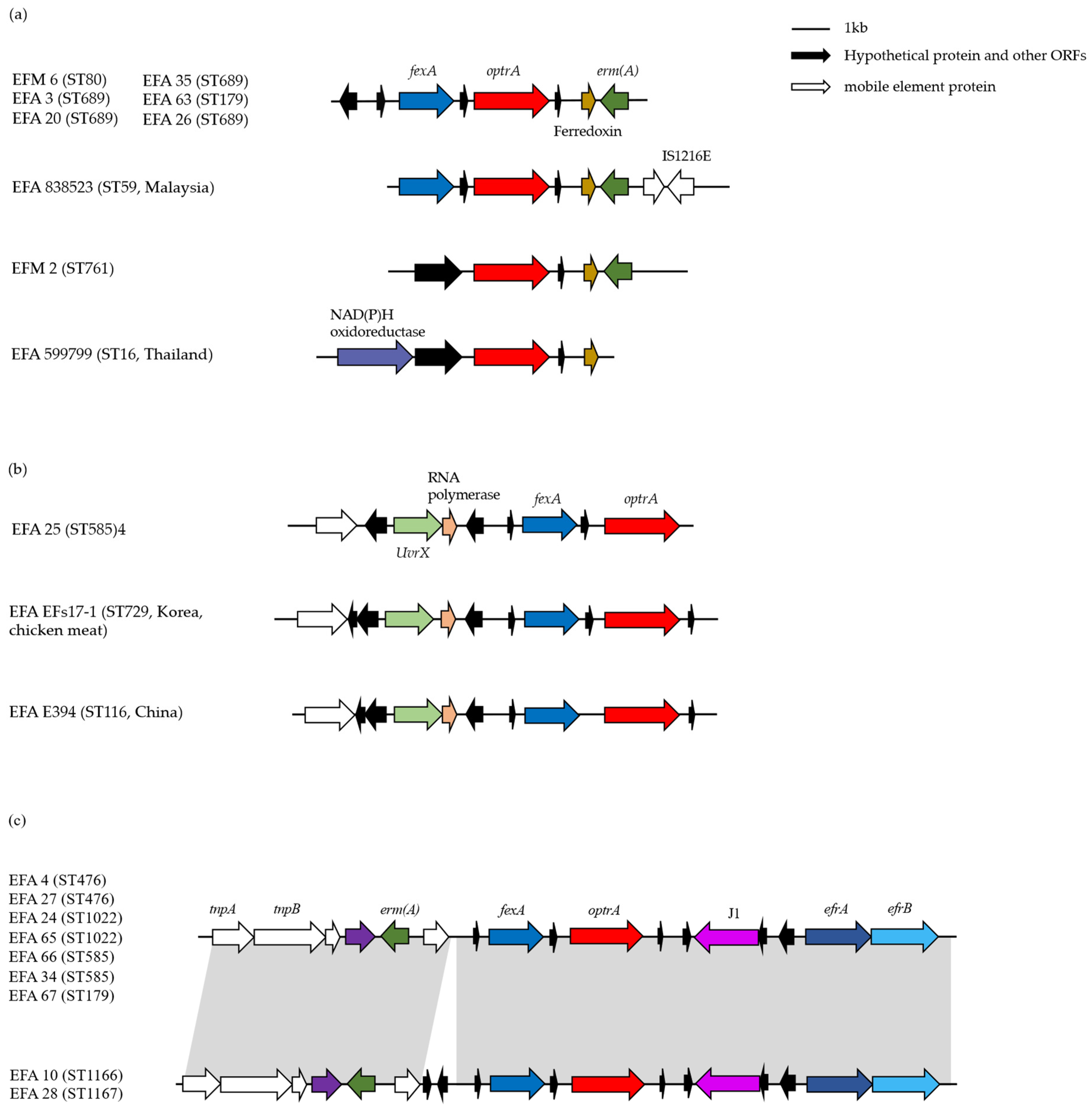
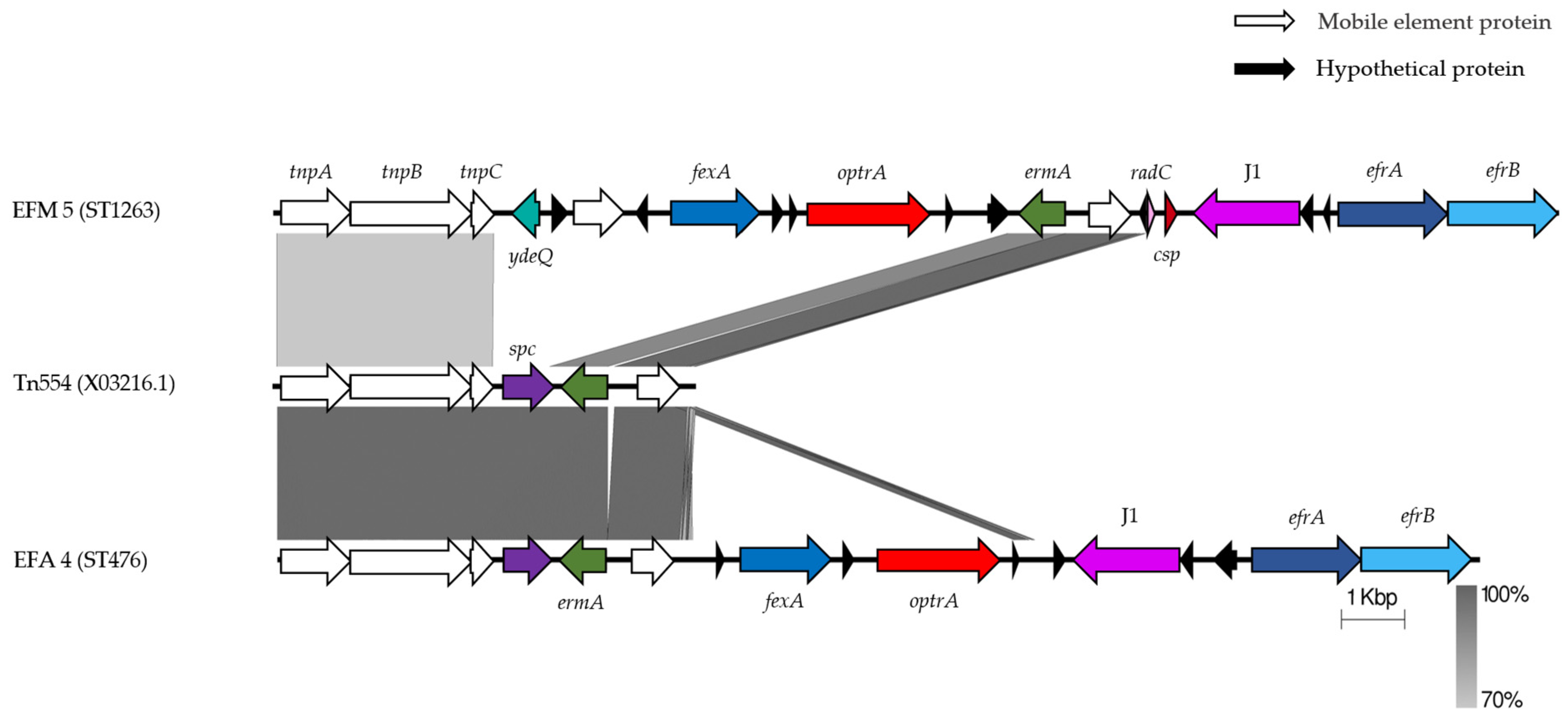
2.3. Genetic Environment of Different Resistance Determinants
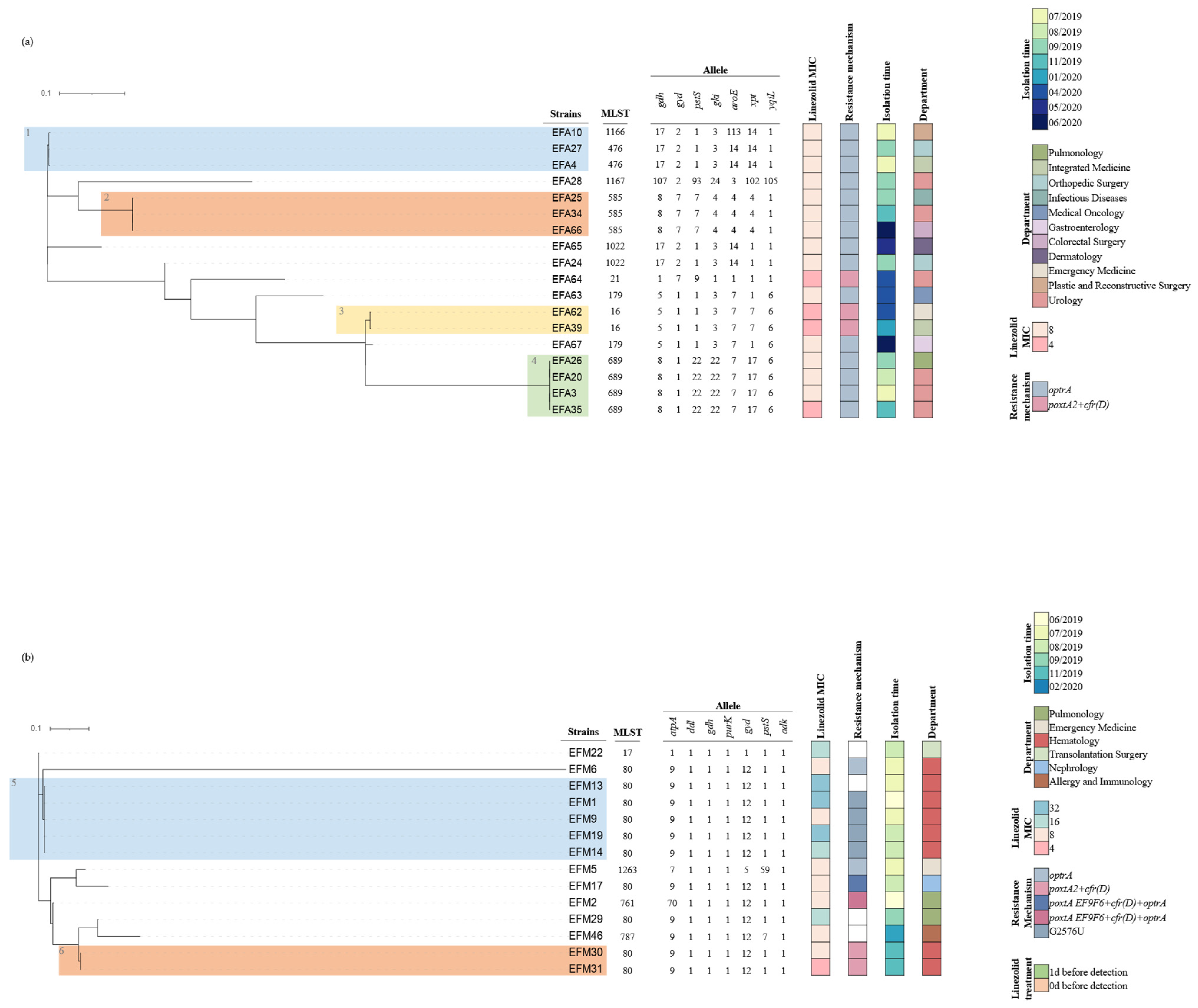
2.4. Clonal Relatedness and Other Molecular Features
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Linezolid Susceptibility Testing
4.3. Whole-Genome Sequencing and Bioinformatic Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Oliveira, D.M.P.; Forde, B.M.; Kidd, T.J.; Harris, P.N.A.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Qin, T.; Fan, W.; Ma, P.; Gu, B. The emerging problem of linezolid-resistant enterococci. J. Glob. Antimicrob. Resist. 2018, 13, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.E.; Deshpande, L.M.; Jones, R.N. Linezolid update: Stable in vitro activity following more than a decade of clinical use and summary of associated resistance mechanisms. Drug Resist. Updat. 2014, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Werckenthin, C.; Kehrenberg, C. Identification of a Plasmid-Borne Chloramphenicol-Florfenicol Resistance Gene in Staphylococcus sciuri. Antimicrob. Agents Chemother. 2000, 44, 2530–2533. [Google Scholar] [CrossRef] [Green Version]
- Long, K.S.; Poehlsgaard, J.; Kehrenberg, C.; Schwarz, S.; Vester, B. The Cfr rRNA Methyltransferase Confers Resistance to Phenicols, Lincosamides, Oxazolidinones, Pleuromutilins, and Streptogramin A Antibiotics. Antimicrob. Agents Chemother. 2006, 50, 2500–2505. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, L.M.; Ashcraft, D.S.; Kahn, H.P.; Pankey, G.; Jones, R.N.; Farrell, D.J.; Mendes, R.E. Detection of a new cfr-like gene, cfr(B), in Enterococcus faecium isolates recovered from human specimens in the United States as part of the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents Chemother. 2015, 59, 6256–6261. [Google Scholar] [CrossRef] [Green Version]
- Bender, J.K.; Fleige, C.; Klare, I.; Fiedler, S.; Mischnik, A.; Mutters, N.T.; Dingle, K.E.; Werner, G. Detection of a cfr(B) Variant in German Enterococcus faecium Clinical Isolates and the Impact on Linezolid Resistance in Enterococcus spp. PLoS ONE 2016, 11, e0167042. [Google Scholar] [CrossRef] [Green Version]
- Guerin, F.; Sassi, M.; Dejoies, L.; Zouari, A.; Schutz, S.; Potrel, S.; Auzou, M.; Collet, A.; Lecointe, D.; Auger, G.; et al. Molecular and functional analysis of the novel cfr(D) linezolid resistance gene identified in Enterococcus faecium. J. Antimicrob. Chemother. 2020, 75, 1699–1703. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hu, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [Green Version]
- Cavaco, L.; Bernal, J.F.; Zankari, E.; Léon, M.; Hendriksen, R.S.; Perez-Gutierrez, E.; Aarestrup, F.; Donado-Godoy, P. Detection of linezolid resistance due to the optrA gene in Enterococcus faecalis from poultry meat from the American continent (Colombia). J. Antimicrob. Chemother. 2016, 72, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Bender, J.K.; Fleige, C.; Lange, D.; Klare, I.; Werner, G. Rapid emergence of highly variable and transferable oxazolidinone and phenicol resistance gene optrA in German Enterococcus spp. clinical isolates. Int. J. Antimicrob. Agents 2018, 52, 819–827. [Google Scholar] [CrossRef] [PubMed]
- Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Linezolid-resistant enterococci in Polish hospitals: Species, clonality and determinants of linezolid resistance. Eur. J. Clin. Microbiol. 2017, 36, 1279–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonelli, A.; D’Andrea, M.M.; Brenciani, A.; Galeotti, C.L.; Morroni, G.; Pollini, S.; Varaldo, P.E.; Rossolini, G.M. Characterization of poxtA, a novel phenicol–oxazolidinone–tetracycline resistance gene from an MRSA of clinical origin. J. Antimicrob. Chemother. 2018, 73, 1763–1769. [Google Scholar] [CrossRef] [Green Version]
- Elghaieb, H.; Freitas, A.; Abbassi, M.S.; Novais, C.; Zouari, M.; Hassen, A.; Peixe, L. Dispersal of linezolid-resistant enterococci carrying poxtA or optrA in retail meat and food-producing animals from Tunisia. J. Antimicrob. Chemother. 2019, 74, 2865–2869. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.R.; Tedim, A.P.; Duarte, B.; Elghaieb, H.; Abbassi, M.S.; Hassen, A.; Read, A.; Alves, V.; Novais, C.; Peixe, L. Linezolid-resistant (Tn6246::fexB-poxtA) Enterococcus faecium strains colonizing humans and bovines on different continents: Similarity without epidemiological link. J. Antimicrob. Chemother. 2020, 75, 2416–2423. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Shan, X.; Li, D.; Schwarz, S.; Zhang, S.-M.; Li, X.-S.; Du, X.-D. Analysis of a poxtA- and optrA-co-carrying conjugative multiresistance plasmid from Enterococcus faecalis. J. Antimicrob. Chemother. 2019, 74, 1771–1775. [Google Scholar] [CrossRef]
- Na, S.-H.; Moon, D.-C.; Kim, M.-H.; Kang, H.-Y.; Kim, S.-J.; Choi, J.-H.; Mechesso, A.-F.; Yoon, S.-S.; Lim, S.-K. Detection of the Phenicol–Oxazolidinone Resistance Gene poxtA in Enterococcus faecium and Enterococcus faecalis from Food-Producing Animals during 2008–2018 in Korea. Microorganisms 2020, 8, 1839. [Google Scholar] [CrossRef]
- Baccani, I.; Antonelli, A.; Di Pilato, V.; Coppi, M.; Di Maggio, T.; Spinicci, M.; Villagran, A.L.; Revollo, C.; Bartoloni, A.; Rossolini, G.M. Detection of poxtA2, a Presumptive poxtA Ancestor, in a Plasmid from a Linezolid-Resistant Enterococcus gallinarum Isolate. Antimicrob. Agents Chemother. 2021, 65, AAC0069521. [Google Scholar] [CrossRef] [PubMed]
- Cinthi, M.; Coccitto, S.N.; Fioriti, S.; Morroni, G.; Simoni, S.; Vignaroli, C.; Magistrali, C.F.; Albini, E.; Brenciani, A.; Giovanetti, E. Occurrence of a plasmid co-carrying cfr(D) and poxtA2 linezolid resistance genes in Enterococcus faecalis and Enterococcus casseliflavus from porcine manure, Italy. J. Antimicrob. Chemother. 2021, 77, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Crowe-McAuliffe, C.; Murina, V.; Turnbull, K.J.; Huch, S.; Kasari, M.; Takada, H.; Nersisyan, L.; Sundsfjord, A.; Hegstad, K.; Atkinson, G.C.; et al. Structural basis for PoxtA-mediated resistance to phenicol and oxazolidinone antibiotics. Nat. Commun. 2022, 13, 1860. [Google Scholar] [CrossRef]
- Park, K.; Jeong, Y.S.; Chang, J.; Sung, H.; Kim, A.M.-N. Emergence of optrA-Mediated Linezolid-Nonsusceptible Enterococcus faecalis in a Tertiary Care Hospital. Ann. Lab. Med. 2020, 40, 321–325. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.-M.; Huh, H.J.; Song, D.J.; Shim, H.J.; Park, K.S.; Kang, C.-I.; Ki, C.-S.; Lee, N.Y. Resistance mechanisms of linezolid-nonsusceptible enterococci in Korea: Low rate of 23S rRNA mutations in Enterococcus faecium. J. Med. Microbiol. 2017, 66, 1730–1735. [Google Scholar] [CrossRef]
- Cho, S.Y.; Kim, H.M.; Chung, D.R.; Kim, S.H.; Huh, H.J.; Kang, C.-I.; Peck, K.R.; Lee, N.Y.; Song, J.-H. Resistance mechanisms and clinical characteristics of linezolid-resistant Enterococcus faecium isolates: A single-centre study in South Korea. J. Glob. Antimicrob. Resist. 2018, 12, 44–47. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yoon, E.-J.; Kim, D.; Shin, J.H.; Shin, J.H.; Shin, K.S.; Kim, Y.A.; Uh, Y.; Kim, H.S.; Kim, Y.R.; et al. Antimicrobial resistance in South Korea: A report from the Korean global antimicrobial resistance surveillance system (Kor-GLASS) for 2017. J. Infect. Chemother. 2019, 25, 845–859. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Hwang, H.; Kim, J.; Lee, M.-J.; Pai, H. Ten-year trends in antibiotic usage at a tertiary care hospital in Korea, 2004 to 2013. Korean J. Intern. Med. 2020, 35, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, L.M.; Castanheira, M.; Flamm, R.K.; Mendes, R.E. Evolving oxazolidinone resistance mechanisms in a worldwide collection of enterococcal clinical isolates: Results from the SENTRY Antimicrobial Surveillance Program. J. Antimicrob. Chemother. 2018, 73, 2314–2322. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.; Son, S.H.; Bin Kim, Y.; Seo, K.W.; Lee, Y.J. Molecular characteristics of optrA-carrying Enterococcus faecalis from chicken meat in South Korea. Poult. Sci. 2020, 99, 6990–6996. [Google Scholar] [CrossRef]
- He, T.; Shen, Y.; Schwarz, S.; Cai, J.; Lv, Y.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J.; et al. Genetic environment of the transferable oxazolidinone/phenicol resistance gene optrA in Enterococcus faecalisisolates of human and animal origin. J. Antimicrob. Chemother. 2016, 71, 1466–1473. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Moon, D.; Mechesso, A.; Choi, J.-H.; Kim, S.-J.; Song, H.-J.; Kim, M.; Yoon, S.-S.; Lim, S.-K. Emergence of cfr-Mediated Linezolid Resistance in Staphylococcus aureus Isolated from Pig Carcasses. Antibiotics 2020, 9, 769. [Google Scholar] [CrossRef]
- Kim, E.; Shin, S.-W.; Kwak, H.-S.; Cha, M.-H.; Yang, S.-M.; Gwak, Y.-S.; Woo, G.-J.; Kim, H.-Y. Prevalence and Characteristics of Phenicol-Oxazolidinone Resistance Genes in Enterococcus faecalis and Enterococcus faecium Isolated from Food-Producing Animals and Meat in Korea. Int. J. Mol. Sci. 2021, 22, 11335. [Google Scholar] [CrossRef]
- Lazaris, A.; Coleman, D.; Kearns, A.M.; Pichon, B.; Kinnevey, P.; Earls, M.R.; Boyle, B.; O’Connell, B.; I Brennan, G.; Shore, A.C. Novel multiresistance cfr plasmids in linezolid-resistant methicillin-resistant Staphylococcus epidermidis and vancomycin-resistant Enterococcus faecium (VRE) from a hospital outbreak: Co-location of cfr and optrA in VRE. J. Antimicrob. Chemother. 2017, 72, 3252–3257. [Google Scholar] [CrossRef]
- Morroni, G.; Brenciani, A.; Antonelli, A.; D’Andrea, M.M.; Di Pilato, V.; Fioriti, S.; Mingoia, M.; Vignaroli, C.; Cirioni, O.; Biavasco, F.; et al. Characterization of a Multiresistance Plasmid Carrying the optrA and cfr Resistance Genes From an Enterococcus faecium Clinical Isolate. Front. Microbiol. 2018, 9, 2189. [Google Scholar] [CrossRef] [Green Version]
- Cinthi, M.; Coccitto, S.N.; D’Achille, G.; Morroni, G.; Simoni, S.; Fioriti, S.; Magistrali, C.F.; Brenciani, A.; Giovanetti, E. Characterization of a novel cfr(D)/poxtA-carrying plasmid in an oxazolidinone-resistant Enterococcus casseliflavus isolate from swine manure, Italy. J. Glob. Antimicrob. Resist. 2022, 30, 308–310. [Google Scholar] [CrossRef]
- Nuesch-Inderbinen, M.; Haussmann, A.; Treier, A.; Zurfluh, K.; Biggel, M.; Stephan, R. Fattening Pigs Are a Reservoir of Florfenicol-Resistant Enterococci Harboring Oxazolidinone Resistance Genes. J. Food Prot. 2022, 85, 740–746. [Google Scholar] [CrossRef]
- Lewis, J.S., II. CLSI M100. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022. [Google Scholar]
- Weinstein, M.P. CLSI M7. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 11th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Clausen, P.; Kaya, H.; Hansen, F.; Knudsen, J.D.; Wang, M.; Holzknecht, B.J.; Samulioniené, J.; Røder, B.L.; Frimodt-Møller, N.; et al. LRE-Finder, a Web tool for detection of the 23S rRNA mutations and the optrA, cfr, cfr(B) and poxtA genes encoding linezolid resistance in enterococci from whole-genome sequences. J. Antimicrob. Chemother. 2019, 74, 1473–1476. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | E. faecalis | E. faecium | ||||||
|---|---|---|---|---|---|---|---|---|
| No. Isoltaes | S a | I b | R c | No. Isoltaes | S | I | R | |
| 2016 | 1536 | 96.1% | 0.5% | 3.4% | 2121 | 96.4% | 0.1% | 3.5% |
| 2017 | 1505 | 95.3% | 1.1% | 3.7% | 2407 | 97.6% | 0.2% | 2.2% |
| 2018 | 1716 | 95.4% | 1.4% | 3.2% | 2558 | 96.9% | 0.4% | 2.7% |
| 2019 | 1544 | 94.2% | 1.2% | 4.7% | 2606 | 95.6% | 0.7% | 3.8% |
| 2020 | 1366 | 90.7% | 2.6% | 6.7% | 2559 | 94.6% | 0.4% | 5.0% |
| MIC (μg/mL) | 4 | 8 | 16 | 32 |
|---|---|---|---|---|
| E. faecalis (n = 18) | 4 (22.2) | 14 (77.8) | 0 | 0 |
| optrA (n = 15, 83.3%) | 1 (6.7) | 14 (93.3) | ||
| cfr(D) + poxtA2 (n = 3, 16.7%) | 3 (100) | |||
| E. faecium (n = 14) | 1 (7.1) | 7 (50.0) | 3 (21.4) | 3 (21.4) |
| optrA (n = 2, 14.3%) | 2 (100) | |||
| optrA+cfr(D) + poxtA-EF9F6 (n = 1, 7.1%) | 1 (100) | |||
| cfr(D) + poxtA-EF9F6 (n = 1, 7.1%) | 1 (100) | |||
| cfr(D) + poxtA2 (n = 2, 14.3%) | 1 (50.00) | 1 (50.0) | ||
| G2576T (n = 4, 28.6%) | 1 (25.0) | 1 (25.0) | 2 (50.0) | |
| unknown mechanism (n = 4, 28.6%) | 1 (25.0) | 2 (50.0) | 1 (25.0) | |
| E. gallinarum (n = 2) | 0 | 2 (100) | 0 | 0 |
| poxtA-EF9F6 (n = 2, 100%) | 2 (100) | |||
| Total (n = 34) | 5 (14.7) | 23 (67.6) | 3 (8.8) | 3 (8.8) |
| optrA (n = 17, 50.0%) | 1 (5.9) | 16 (94.1) | ||
| optrA+cfr(D) + poxtA-EF9F6 (n = 1, 2.9%) | 1 (100) | |||
| cfr(D) + poxtA-EF9F6 (n = 1, 2.9%) | 1 (100) | |||
| cfr(D) + poxtA2 (n = 5, 14.7%) | 4 (80.0) | 1 (20.0) | ||
| poxtA-EF9F6 (n = 2, 5.9%) | 2 (100) | |||
| G2576T (n = 4, 11.8%) | 1 (25.0) | 1 (25.0) | 2 (50.0) | |
| unknown mechanism (n = 4, 11.8%) | 1 (25.0) | 2 (50.0) | 1 (25.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Won, D.; Nguyen, L.P.; Osei, K.M.; Seo, Y.; Kim, J.; Lee, Y.; Lee, H.; Yong, D.; Choi, J.R.; et al. Prevalence and Genetic Analysis of Resistance Mechanisms of Linezolid-Nonsusceptible Enterococci in a Tertiary Care Hospital Examined via Whole-Genome Sequencing. Antibiotics 2022, 11, 1624. https://doi.org/10.3390/antibiotics11111624
Hu Y, Won D, Nguyen LP, Osei KM, Seo Y, Kim J, Lee Y, Lee H, Yong D, Choi JR, et al. Prevalence and Genetic Analysis of Resistance Mechanisms of Linezolid-Nonsusceptible Enterococci in a Tertiary Care Hospital Examined via Whole-Genome Sequencing. Antibiotics. 2022; 11(11):1624. https://doi.org/10.3390/antibiotics11111624
Chicago/Turabian StyleHu, Yuxin, Dongju Won, Le Phuong Nguyen, Kennedy Mensah Osei, Younghee Seo, Junglim Kim, Yoonhee Lee, Hyukmin Lee, Dongeun Yong, Jong Rak Choi, and et al. 2022. "Prevalence and Genetic Analysis of Resistance Mechanisms of Linezolid-Nonsusceptible Enterococci in a Tertiary Care Hospital Examined via Whole-Genome Sequencing" Antibiotics 11, no. 11: 1624. https://doi.org/10.3390/antibiotics11111624