
In Vitro Antimicrobial Effects and Inactivation Mechanisms of 5,8-Dihydroxy-1,4-Napthoquinone

Abstract
:1. Introduction
2. Results
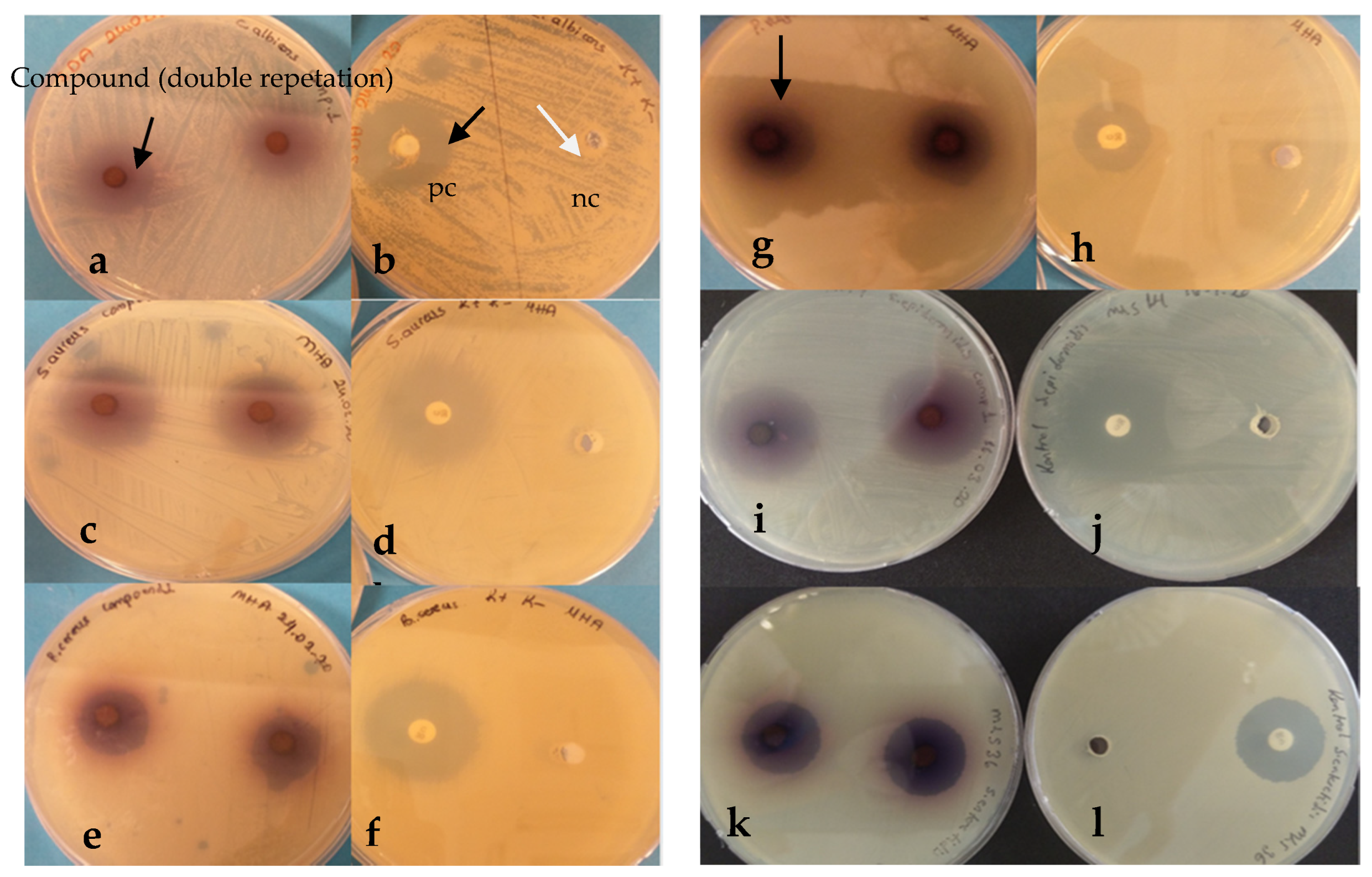
2.1. Antimicrobial Test
2.2. Minimum Inhibition Concentration (MIC50)
2.3. Crystal Violet Assay
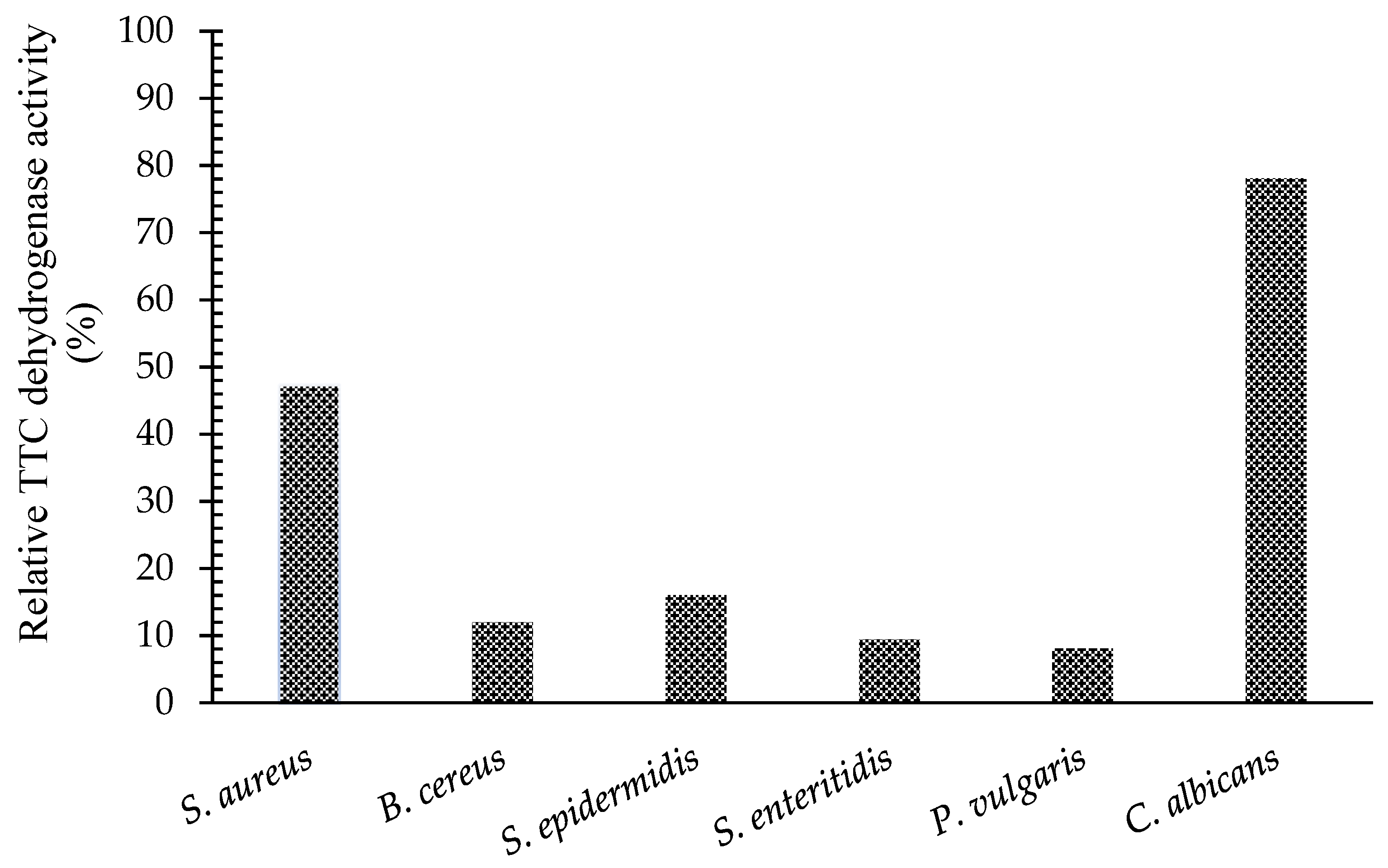
2.4. Measurement of TTC Dehydrogenase Activity
2.5. DNA Leakage
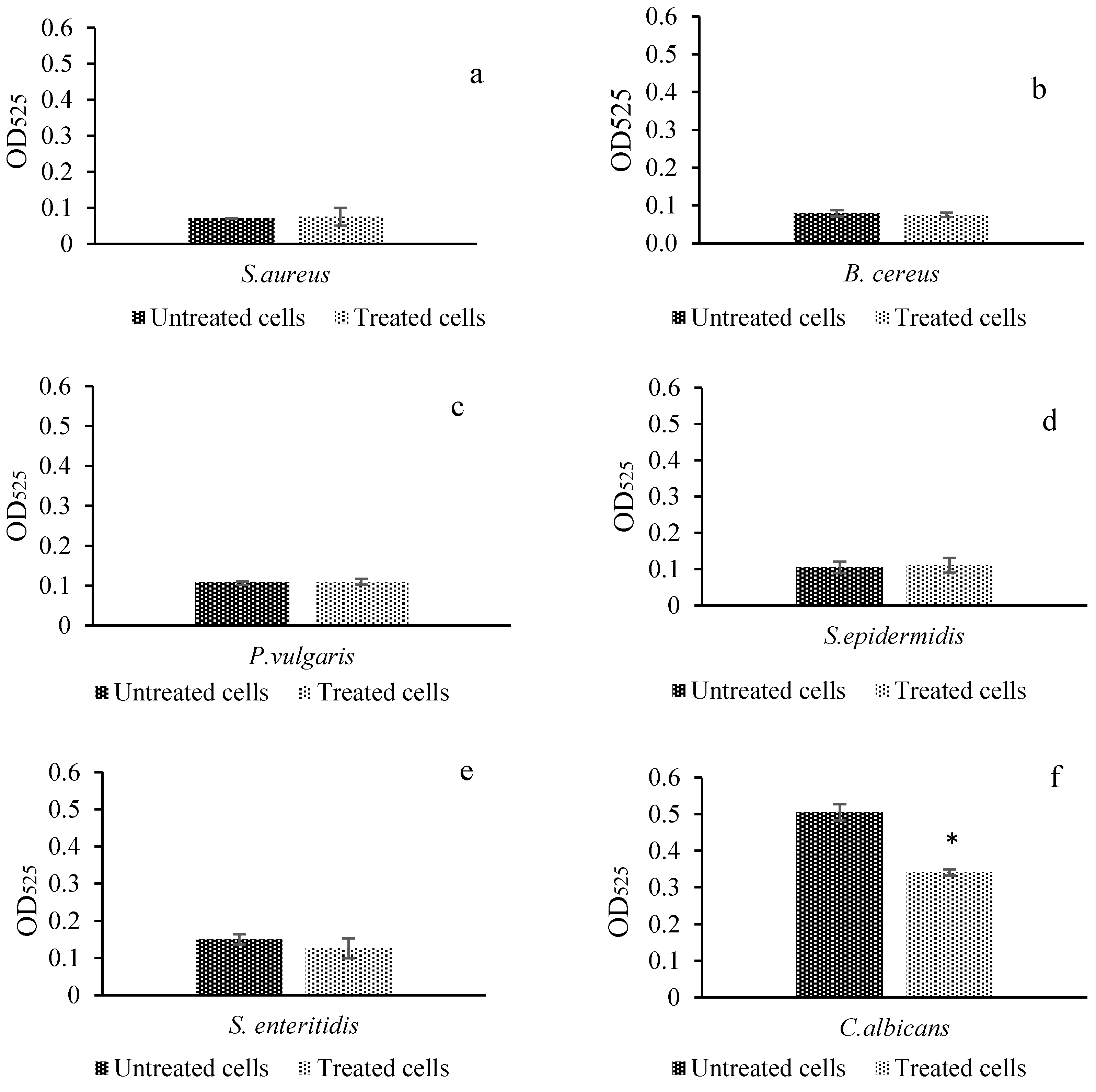
2.6. Protein Leakage
2.7. SDS-PAGE Analysis
2.8. DNA Damage
2.9. FT–IR Analysis
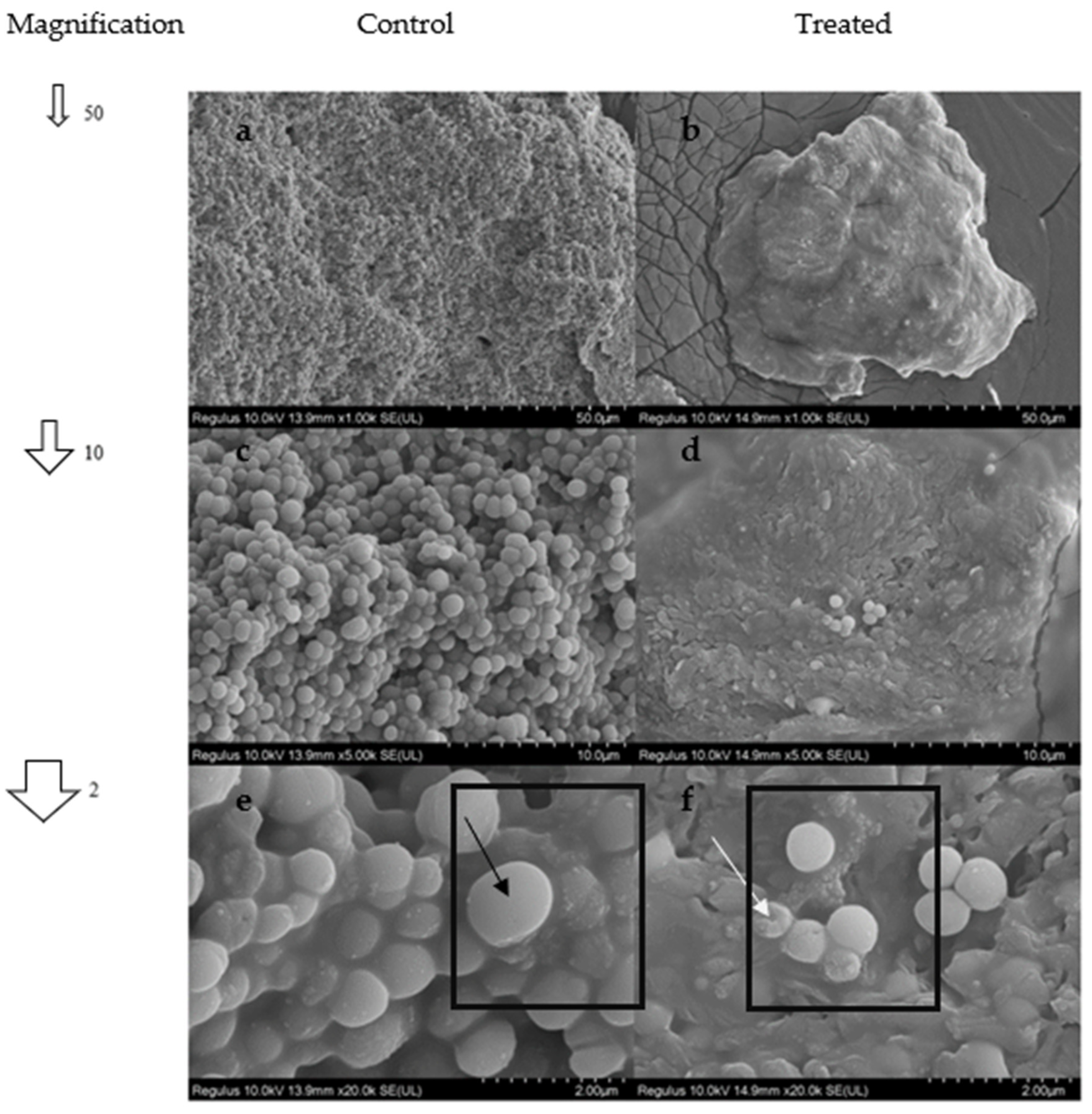
2.10. SEM Analysis
3. Discussion
4. Materials and Methods
4.1. Screening of In Vitro Antimicrobial Effect
4.2. Minimum Inhibition Concentration (MIC50) Assay
4.3. Mechanism of Action Assays
4.3.1. Preliminary Preparation of Experiments
4.3.2. Crystal Violet Assay
4.3.3. TTC Dehydrogenase Activity
4.3.4. DNA Leakage
4.3.5. Protein Leakage
4.3.6. SDS PAGE Analysis
4.3.7. DNA Damage
4.3.8. FT–IR Analysis
4.3.9. SEM Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Mone, N.S.; Bhagwat, S.A.; Sharma, D.; Chaskar, M.; Patil, R.H.; Zamboni, P.; Nawani, N.N.; Satpute, S.K. Naphthoquinones and Their Derivatives: Emerging Trends in Combating Microbial Pathogens. Coatings 2021, 11, 434. [Google Scholar] [CrossRef]
- Sánchez-Calvo, J.M.; Barbero, G.R.; Guerrero-Vásquez, G.; Durán, A.G.; Macías, M.; Rodríguez-Iglesias, M.A.; Molinillo, J.M.G.; Macías, F.A. Synthesis, antibacterial and antifungal activities of naphthoquinone derivatives: A structure–activity relationship study. Med. Chem. Res. 2016, 25, 1274–1285. [Google Scholar] [CrossRef]
- Ravichandiran, P.; Sheet, S.; Premnath, D.; Kim, A.R.; Yoo, D.J. 1,4-Naphthoquinone Analogues: Potent Antibacterial Agents and Mode of Action Evaluation. Molecules 2019, 24, 1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuck, D.C.; Ferreira, S.B.; Cruz, L.N.; da Rocha, D.R.; Moraes, M.S.; Nakabashi, M.; Rosenthal, P.J.; Ferreira, V.F.; Garcia, C.R. Biological evaluation of hydroxynaphthoquinones as anti-malarials. Malar. J. 2013, 12, 234–236. [Google Scholar] [CrossRef] [Green Version]
- Salas, C.O.; Faúndez, M.; Morello, A.; Maya, J.D.; Tapia, R. Natural and synthetic naphthoquinones active against Trypanosoma cruzi: An initial step towards new drugs for Chagas disease. Curr. Med. Chem. 2011, 18, 144–161. [Google Scholar] [CrossRef]
- Tandon, V.K.; Singh, R.V.; Yadav, D.B. Synthesis and evaluation of novel 1,4-naphthoquinone derivatives as antiviral, antifungal and anticancer agents. Bioorganic Med. Chem. Lett. 2004, 14, 2901–2904. [Google Scholar] [CrossRef]
- Wellington, K.W.; Nyoka, N.B.P.; McGaw, L.J. Investigation of the antibacterial and antifungal activity of thiolated naphthoquinones. Drug Dev. Res. 2019, 80, 386–394. [Google Scholar] [CrossRef]
- Lall, N.; Weiganand, O.; Hussein, A.; Meyer, J. Antifungal activity of naphthoquinones and triterpenes isolated from the root bark of Euclea natalensis. S. Afr. J. Bot. 2006, 72, 579–583. [Google Scholar] [CrossRef] [Green Version]
- Widhalm, J.R.; Rhodes, D. Biosynthesis and molecular actions of specialized 1,4-naphthoquinone natural products produced by horticultural plants. Hortic. Res. 2016, 3, 16046. [Google Scholar] [CrossRef]
- Hughes, L.M.; Lanteri, C.A.; O’Neil, M.T.; Johnson, J.D.; Gribble, G.W.; Trumpower, B.L. Design of anti-parasitic and anti-fungal hydroxy-naphthoquinones that are less susceptible to drug resistance. Mol. Biochem. Parasitol. 2011, 177, 12–19. [Google Scholar] [CrossRef]
- Devi, S.P.; Kumaria, S.; Rao, S.R.; Tandon, P. Carnivorous Plants as a Source of Potent Bioactive Compound: Naphthoquinones. Trop. Plant Biol. 2016, 9, 267–279. [Google Scholar] [CrossRef]
- Rudnicka, M.; Ludynia, M.; Karcz, W. The Effect of Naphthazarin on the Growth, Electrogenicity, Oxidative Stress, and Microtubule Array in Z. mays Coleoptile Cells Treated with IAA. Front. Plant Sci. 2019, 9, 1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitagawa, R.R.; Bonacorsi, C.; Da Fonseca, L.M.; Vilegas, W.; Raddi, M.S.G. Anti-Helicobacter pylori activity and oxidative burst inhibition by the naphthoquinone 5-methoxy-3,4-dehydroxanthomegnin from Paepalanthus latipes. Rev. Bras. Farm. 2011, 22, 53–59. [Google Scholar] [CrossRef] [Green Version]
- Induli, M.; Cheloti, M.; Wasuna, A.; Wekesa, I.; Wanjohi, J.M.; Byamukama, R.; Heydenrich, M.; Makayoto, M.; Yenesew, A. Naphthoquinones from the roots of Aloe secundiflora. Phytochem. Lett. 2012, 5, 506–509. [Google Scholar] [CrossRef]
- Rahmoun, N.; Boucherit-Otmani, Z.; Boucherit, K.; Benabdallah, M.; Villemin, D.; Choukchou-Braham, N. Antibacterial and antifungal activity of lawsone and novel naphthoquinone derivatives. Med. Mal. Infect. 2012, 42, 270–275. [Google Scholar] [CrossRef]
- Ibis, C.; Tuyun, A.F.; Ozsoy-Gunes, Z.; Ayla, S.S.; Stasevych, M.; Musyanovych, R.; Komarovska-Porokhnyavets, O.; Novikov, V. Synthesis and Antibacterial and Antifungal Properties of Novel S-, N-, N,S-, and S,O-Substituted 1,4-Naphthoquinone Derivatives. Phosphorus Sulfur Silicon Relat. Elem. 2013, 188, 955–966. [Google Scholar] [CrossRef]
- Chadar, D.; Rao, S.S.; Gejji, S.P.; Ugale, B.; Nagaraja, C.M.; Nikalje, M.; Salunke-Gawali, S. Regioselective synthesis of a vitamin K3 based dihydrobenzophenazine derivative: Its novel crystal structure and DFT studies. RSC Adv. 2015, 5, 76419–76423. [Google Scholar] [CrossRef]
- Tomozane, H.; Takeuchi, Y.; Choshi, T.; Kishida, S.; Yamato, M. Syntheses and antifungal activities of dl-griseofulvin and its congeners. I. Chem. Pharm. Bull. 1990, 38, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Oogose, K.; Hafuri, Y.; Takemori, E.; Nakata, E.; Inouye, Y.; Nakamura, S.; Kubo, A. Mechanism of Inhibition of Reverse Transcriptase by Quinone Antibiotics. J. Antibiot. 1987, 40, 1778–1781. [Google Scholar] [CrossRef] [Green Version]
- McDonnell, G.; Russell, A.D. Antiseptics and Disinfectants: Activity, Action, and Resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef]
- Ramos-Peralta, L.; Lopez-Lopez, L.I.; Silva-Belmares, S.Y.; Zugasti-Cruz, A.; Rodriguez-Herrera, R.; Anguilar-Gonzalez, C.N. Naphthoquinone: Bioactivity and green synthesis. In Basic Science, Technological Advances and Educational Programs; Formatex: Copenhagen, Denmark, 2015; pp. 542–550. [Google Scholar] [CrossRef]
- Yap, J.K.Y.; Tan, S.Y.Y.; Tang, S.Q.; Thien, V.K.; Chan, E.W.L. Synergistic Antibacterial Activity between 1,4-Naphthoquinone and β-Lactam Antibiotics against Methicillin-Resistant Staphylococcus aureus. Microb. Drug Resist. 2021, 27, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Yu, B.; Friedrich, D.; Li, J.; Shen, H.; Krautscheid, H.; Huang, S.D.; Kim, M.-H. Naphthoquinone-derivative as a synthetic compound to overcome the antibiotic resistance of methicillin-resistant S. aureus. Commun. Biol. 2020, 3, 529. [Google Scholar] [CrossRef] [PubMed]
- Brandelli, A.D.B.; Martinelli, M.; Stefani, V.; Gerbase, A.E. Antimicrobial activity of 1,4-naphthoquinones by metal complexation. Braz. J. Pharm. Sci. 2004, 40, 247–251. [Google Scholar] [CrossRef]
- Futuro, D.O.; Ferreira, P.G.; Nicoletti, C.D.; Borba, L.P. The Antifungal Activity of Naphthoquinones: An Integrative Review. An. Acad. Bras. Ciênc. 2018, 90, 1187–1214. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, J.P.; Baker, C.; Kawasaki, Y.; Subedi, Y.P.; de Paul, N.N.V.; Takemoto, J.Y.; Chang, C.-W.T. Synthesis and bioactivity investigation of quinone-based dimeric cationic triazolium amphiphiles selective against resistant fungal and bacterial pathogens. Eur. J. Med. Chem. 2017, 126, 696–704. [Google Scholar] [CrossRef]
- Do Perpetuo Socorro Borges Carriço Ferreira, M.; do Carmo Cardoso, M.F.; Da Silva, F.D.C.; Ferreira, V.F.; Lima, E.S.; Souza, J.V.B. Antifungal Activity of Synthetic Naphthoquinones against Dermatophytes and Opportunistic Fungi: Preliminary Mechanism-of-Action Tests. Ann. Clin. Microbiol. Antimicrob. 2014, 13, 26. [Google Scholar] [CrossRef] [Green Version]
- Du, T.; Chen, S.; Zhang, J.; Li, T.; Li, P.; Liu, J.; Du, X.; Wang, S. Antibacterial Activity of Manganese Dioxide Nanosheets by ROS-Mediated Pathways and Destroying Membrane Integrity. Nanomaterials 2020, 10, 1545. [Google Scholar] [CrossRef]
- Tang, W.; Zeng, X.; Zhao, Y.; Ye, G.; Gui, W.; Ni, Y. Disinfection Effect and Its Mechanism of Electrolyzed Oxidizing Water on Spores of Bacillus Subtilis Var. Niger. Food Sci. Biotechnol. 2011, 20, 889–895. [Google Scholar] [CrossRef]
- Ersoy, Z.G.; Dinc, O.; Cinar, B.; Gedik, S.T.; Dimoglo, A. Comparative Evaluation of Disinfection Mechanism of Sodium Hypochlorite, Chlorine Dioxide and Electroactivated Water on Enterococcus faecalis. Lebensm.-Wiss. Technol. 2019, 102, 205–213. [Google Scholar] [CrossRef]
- Ward, T.H.; Marples, B. Technical report: SYBR Green I andthe improved sensitivity of the single-cell electrophoresis assay. Int. J. Radiat. Biol. 2000, 76, 61–65. [Google Scholar] [CrossRef]
- Holmström, K.M.; Finkel, T. Cellular Mechanisms and Physiological Consequences of Redox-Dependent Signalling. Nat. Rev. Mol. Cell Biol. 2014, 15, 411. [Google Scholar] [CrossRef] [PubMed]
- Fieser, L.F.; Fieser, M. The reduction potentials of various naphthoquinones. J. Am. Chem. Soc. 1935, 57, 491–494. [Google Scholar] [CrossRef]
- De Castro, S.L.; Emery, F.S.; Da Silva Júnior, E.N. Synthesis of Quinoidal Molecules: Strategies towards Bioactive Compounds with an Emphasis on Lapachones. Eur. J. Med. Chem. 2013, 69, 678–700. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, Y.; Shinkai, Y.; Miura, T.; Arthur, K. Cho2 The chemical biology of naphthoquinones and its environmental implications. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 221–247. [Google Scholar] [CrossRef]
- Chan, K.L.A.; Kazarian, S.G. Detection of Trace Materials with Fourier Transform Infrared Spectroscopy Using a Multi-Channel Detector. Analyst 2006, 131, 126–131. [Google Scholar] [CrossRef]
- Prince, A.; Sandhu, P.; Ror, P.; Dash, E.; Sharma, S.; Arakha, M.; Jha, S.; Akhter, Y.; Saleem, M. Lipid-II Independent Antimicrobial Mechanism of Nisin Depends on Its Crowding and Degree of Oligomerization. Sci. Rep. 2016, 6, 37908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valgas, C.; De Souza, S.M.; Smânia, E.F.A.; Smânia, A. Screening Methods to Determine Antibacterial Activity of Natural Products. Braz. J. Microbiol. 2007, 38, 369–380. [Google Scholar] [CrossRef] [Green Version]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Vaara, M.; Vaara, T. Outer Membrane Permeability Barrier Disruption by Polymyxin in Polymyxin-Susceptible and -Resistant Salmonella Typhimurium. Antimicrob. Agents Chemother. 1981, 19, 578–583. [Google Scholar] [CrossRef]
- Normark, S.; Westling, B. Nature of the Penetration Barrier in Escherichia coli K-12: Effect of Macromolecular Inhibition of Penetrability in Strains Containing the EnvA Gene. J. Bacteriol. 1971, 108, 45–50. [Google Scholar] [CrossRef] [Green Version]
- Ding, T.; Dong, Q.L.; Rahman, S.M.E.; Oh, D.H. Response Surface Modeling of Listeria Monocytogenes Inactivation on Lettuce Treated with Electrolyzed Oxidizing Water. J. Food Process Eng. 2011, 34, 1729–1745. [Google Scholar] [CrossRef]
- Zeng, X.; Tang, W.; Ye, G.; Ouyang, T.; Tian, L.; Ni, Y.; Li, P. Studies on Disinfection Mechanism of Electrolyzed Oxidizing Water on E. Coli and Staphylococcus aureus. J. Food Sci. 2010, 75, M253–M260. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; Kim, J.; Kim, J.Y.; Yoon, J.; Kim, J.H. Mechanisms of Escherichia coli Inactivation by Several Disinfectants. Water Res. 2010, 44, 3410–3418. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, R.; Yang, Z.; Zhan, Y.; Ma, Y.; Ping, S.; Zhang, L.; Lin, M.; Yan, Y. 1-Aminocyclopropane-1-Carboxylate Deaminase from Pseudomonas stutzeri A1501 Facilitates the Growth of Rice in the Presence of Salt or Heavy Metals. J. Microbiol. Biotechnol. 2015, 25, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Turkay, O.; Seker, M.G.; Ramadhan, S.S.; Dimoglo, A. Electro Activated Water Mechanism on Escherichia coli and Enterococcus faecalis and Optimization of Operating Parameters. J. Food Process. Preserv. 2018, 42, e13345. [Google Scholar] [CrossRef]
- Farkas, A.; Maróti, G.; Kereszt, A.; Kondorosi, É.; Luiz Franco, O.; Nascimento Silva, O.; Bosco, D.; Diana Gaspar, B.; Maróti, G.; Farkas, A.; et al. Comparative Analysis of the Bacterial Membrane Disruption Effect of Two Natural Plant Antimicrobial Peptides. Front. Microbiol. 2017, 8, 51. [Google Scholar] [CrossRef]








{kind=link}
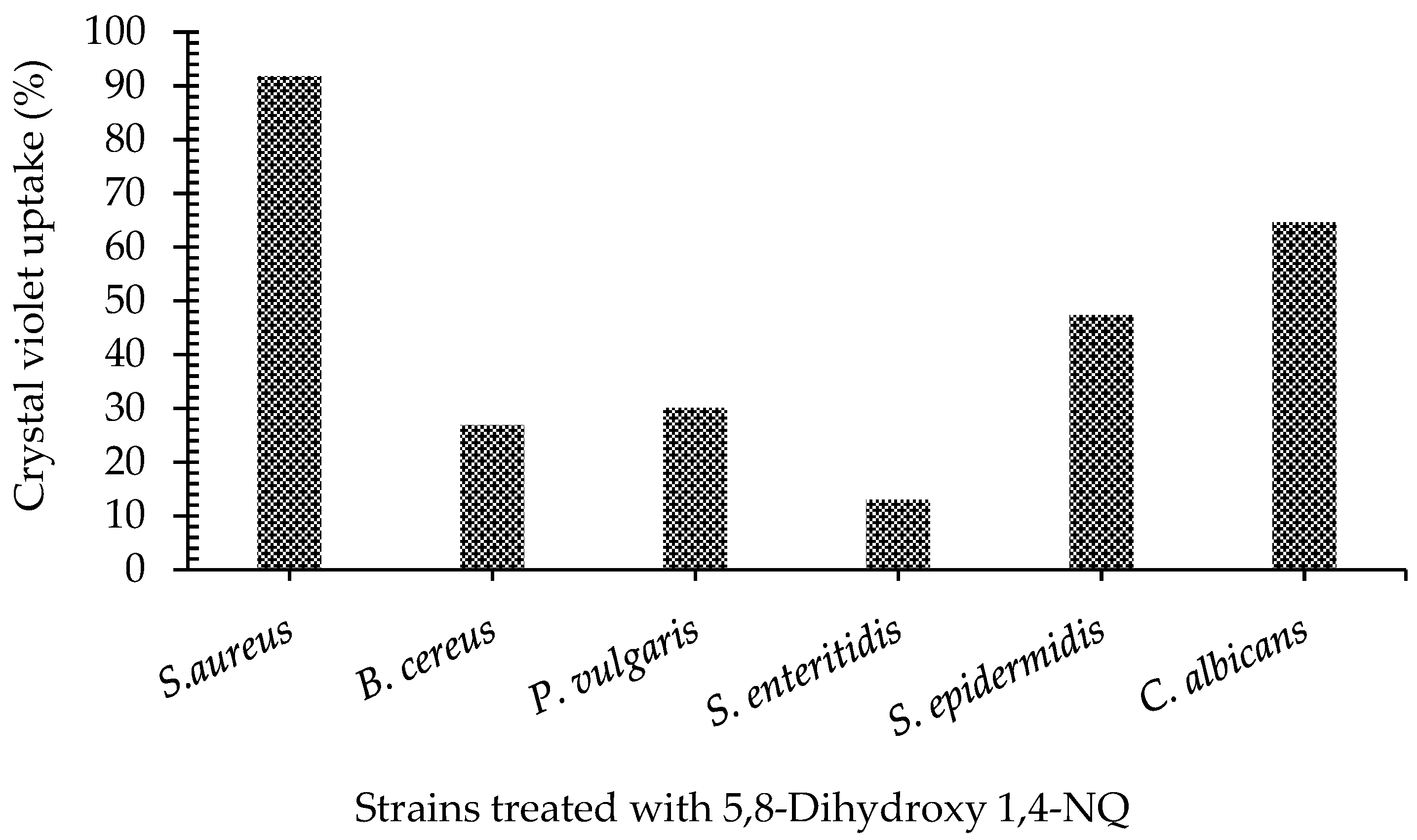{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Zone Diameter (mm) * | Negative Control DMSO | Positive Control Chloramphenicol (C30), Nystatin ** |
|---|---|---|---|
| S. aureus | 24 | - | 26 cm |
| S. epidermidis | 30 | - | 26 cm |
| E. faecalis | 11 | - | 19 cm |
| B. cereus | 18 | - | 26 cm |
| P. vulgaris | 19 | - | 18 cm |
| E. coli | 8 | - | 25 cm |
| K. pneumoniae | - | - | 22 cm |
| P. aeruginosa | - | - | 15 cm |
| S. enteritidis | 22 | - | 23 cm |
| C. albicans | 18 | - | 25 cm |
| Microorganism Name | MIC50 Value |
|---|---|
| S. aureus | 1.2 μg/mL |
| S. enteritidis | 6 μg/mL |
| S. epidermidis | 2.4 μg/mL |
| B. cereus | 14 μg/mL |
| P. vulgaris | 10 μg/mL |
| C. albicans | <0.6 μg/mL |
| Microorganisms | DNA Leakage (µg/mL) |
|---|---|
| S. enteritidis | 4.375 |
| C. albicans | 3.25 |
| S. epidermidis | 2.875 |
| S. aureus | ND * |
| B. cereus | ND |
| P. vulgaris | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Topçu, S.; Şeker, M.G. In Vitro Antimicrobial Effects and Inactivation Mechanisms of 5,8-Dihydroxy-1,4-Napthoquinone. Antibiotics 2022, 11, 1537. https://doi.org/10.3390/antibiotics11111537
Topçu S, Şeker MG. In Vitro Antimicrobial Effects and Inactivation Mechanisms of 5,8-Dihydroxy-1,4-Napthoquinone. Antibiotics. 2022; 11(11):1537. https://doi.org/10.3390/antibiotics11111537
Chicago/Turabian StyleTopçu, Seray, and Mine Gül Şeker. 2022. "In Vitro Antimicrobial Effects and Inactivation Mechanisms of 5,8-Dihydroxy-1,4-Napthoquinone" Antibiotics 11, no. 11: 1537. https://doi.org/10.3390/antibiotics11111537