
In Vitro Activity of Robenidine Analogues NCL259 and NCL265 against Gram-Negative Pathogens
 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
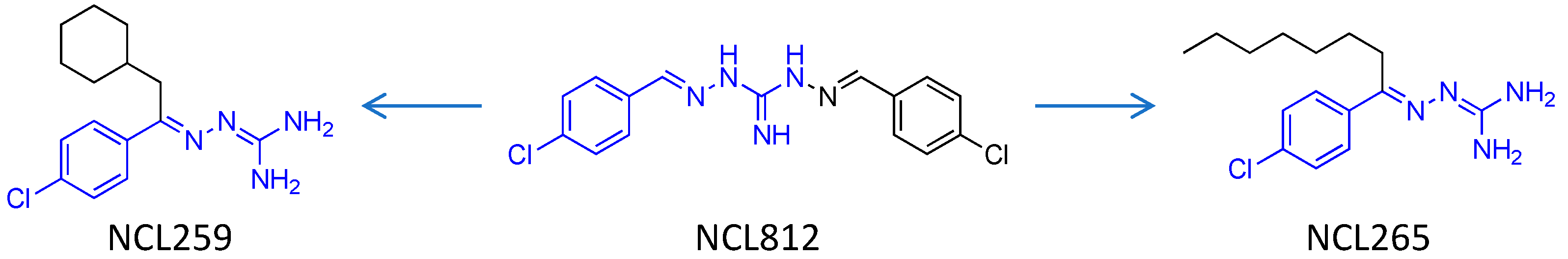
2.1. Antimicrobial Agents
2.2. Bacterial Strains and Growth Conditions
2.3. Antimicrobial Susceptibility Testing
2.4. Synergy Testing by a Checkerboard Assay
2.5. In Vitro Cytotoxicity and Hemolysis Assays
3. Results
3.1. NCL259 and NCL265 Show Antimicrobial Activity against Gram-Negative Reference Strains
3.2. Combination of NCL259 or NCL265 with PMB Increases Antimicrobial Activity against Gram-Negative Reference Strains
3.3. NCL259 and NCL265 Show Antimicrobial Activity against Gram-Negative Clinical Isolates from Animals
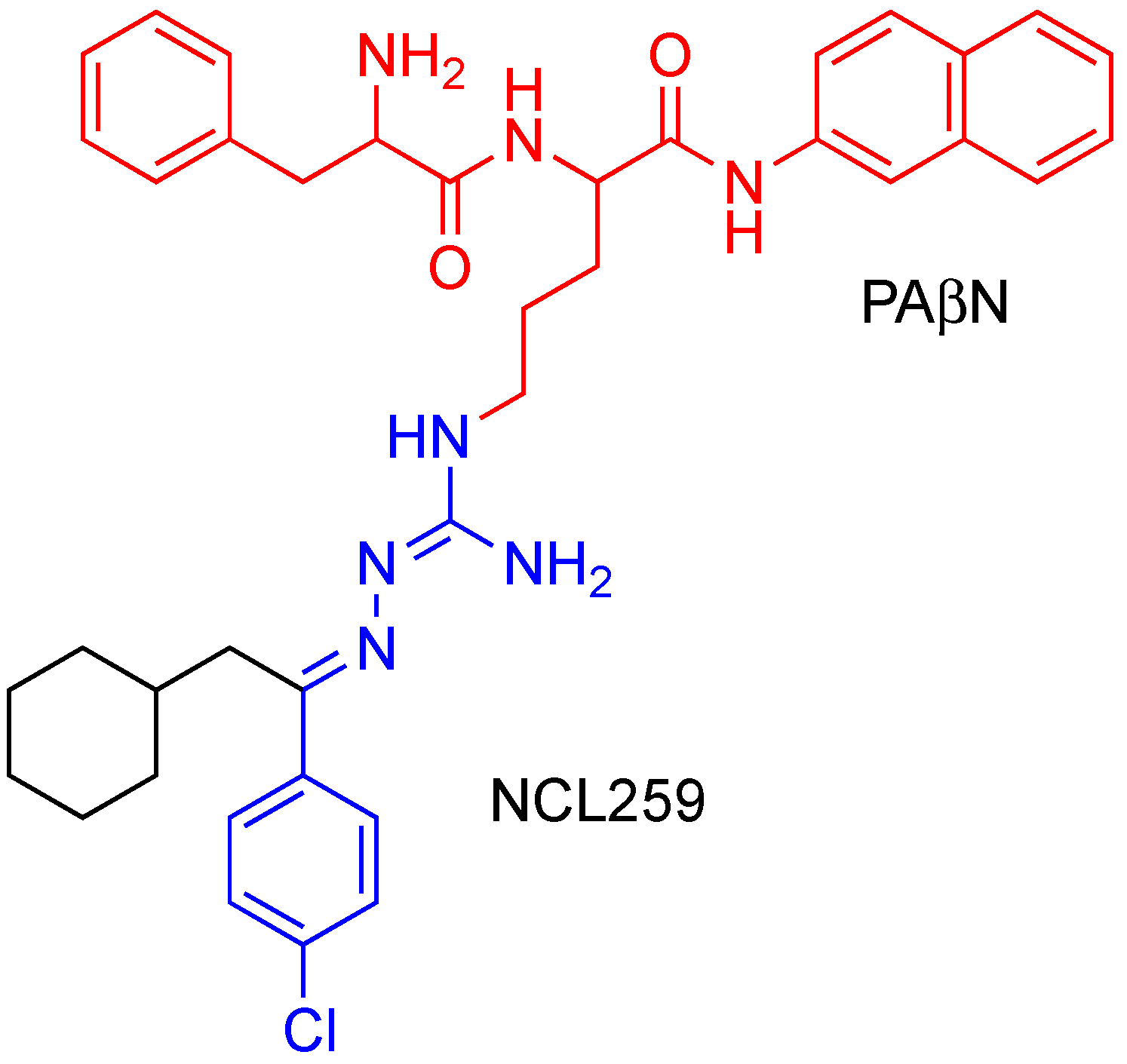
3.4. Efflux Pump Inhibitor (PAβN) Increases the Antimicrobial Activity of NCL259 and NCL265 against Resistant Klebsiella Isolates
3.5. The AcrA/AcrB/TolC Multidrug Efflux Pump Impacts the Activity of NCL259 and NCL265
3.6. NCL259 and NCL265 Are Toxic to Mammalian Cells and Hemolytic to Erythrocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis. 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Zilberberg, M.D.; Shorr, A.F.; Micek, S.T.; Vazquez-Guillamet, C.; Kollef, M.H. Multi-drug resistance, inappropriate initial antibiotic therapy and mortality in Gram-negative severe sepsis and septic shock: A retrospective cohort study. Crit. Care 2014, 18, 596. [Google Scholar] [CrossRef] [PubMed]
- Frieri, M.; Kumar, K.; Boutin, A. Antibiotic resistance. J. Infect. Public Health 2017, 10, 369–378. [Google Scholar] [CrossRef]
- Merker, M.; Tueffers, L.; Vallier, M.; Groth, E.E.; Sonnenkalb, L.; Unterweger, D.; Baines, J.F.; Niemann, S.; Schulenburg, H. Evolutionary Approaches to Combat Antibiotic Resistance: Opportunities and Challenges for Precision Medicine. Front. Immunol. 2020, 11, 1938. [Google Scholar] [CrossRef] [PubMed]
- Michael, C.A.; Dominey-Howes, D.; Labbate, M. The antimicrobial resistance crisis: Causes, consequences, and management. Front. Public Health 2014, 2, 145. [Google Scholar] [CrossRef]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert Rev. Anti Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of antimicrobial resistance in ESKAPE pathogens. BioMed Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef]
- Cardoso, T.; Ribeiro, O.; Aragão, I.C.; Costa-Pereira, A.; Sarmento, A.E. Additional risk factors for infection by multidrug-resistant pathogens in healthcare-associated infection: A large cohort study. BMC Infect. Dis. 2012, 12, 375. [Google Scholar] [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- Ventola, C.L. The Antibiotic Resistance Crisis: Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- Talbot, G.H.; Jezek, A.; Murray, B.E.; Jones, R.N.; Ebright, R.H.; Nau, G.J.; Rodvold, K.A.; Newland, J.G.; Boucher, H.W.; The Infectious Diseases Society of America. The Infectious Diseases Society of America’s 10 × ′20 Initiative (10 New Systemic Antibacterial Agents US Food and Drug Administration Approved by 2020): Is 20 × ′20 a Possibility? Clin. Infect. Dis. 2019, 69, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DiMasi, J.A.; Grabowski, H.G.; Hansen, R.W. Innovation in the pharmaceutical industry: New estimates of R&D costs. J. Health Econ. 2016, 47, 20–33. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, J.; Davies, S.; Rex, J.; White, L.; Murray, R. Review on Antimicrobial Resistance, Tackling Drug-Resistant Infections Globally: Final Report and Recommendations; Wellcome Trust: London, UK, 2016. [Google Scholar]
- Lepore, C.; Silver, L.; Theuretzbacher, U.; Thomas, J.; Visi, D. The small-molecule antibiotics pipeline: 2014-2018. Nat. Rev. Drug Discov. 2019, 18, 739. [Google Scholar] [CrossRef]
- Mulani, M.S.; Kamble, E.E.; Kumkar, S.N.; Tawre, M.S.; Pardesi, K.R. Emerging Strategies to Combat ESKAPE Pathogens in the Era of Antimicrobial Resistance: A Review. Front. Microbiol. 2019, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41. [Google Scholar] [CrossRef]
- Theuretzbacher, U. Global antimicrobial resistance in Gram-negative pathogens and clinical need. Curr. Opin. Microbiol. 2017, 39, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Arzanlou, M.; Chai, W.C.; Venter, H. Intrinsic, adaptive and acquired antimicrobial resistance in Gram-negative bacteria. Essays Biochem. 2017, 61, 49–59. [Google Scholar] [CrossRef]
- Kantor, S.; Kennett, R.L.; Waletzky, E.; Tomcufcik, A.S. 1,3-Bis(p-chlorobenzylideneamino)guanidine Hydrochloride (Robenzidene): New Poultry Anticoccidial Agent. Science 1970, 168, 373–374. [Google Scholar] [CrossRef]
- Abraham, R.J.; Stevens, A.J.; Young, K.A.; Russell, C.; Qvist, A.; Khazandi, M.; Wong, H.S.; Abraham, S.; Ogunniyi, A.D.; Page, S.W.; et al. Robenidine Analogues as Gram-Positive Antibacterial Agents. J. Med. Chem. 2016, 59, 2126–2138. [Google Scholar] [CrossRef]
- Ogunniyi, A.D.; Khazandi, M.; Stevens, A.J.; Sims, S.K.; Page, S.W.; Garg, S.; Venter, H.; Powell, A.; White, K.; Petrovski, K.R.; et al. Evaluation of robenidine analog NCL195 as a novel broad-spectrum antibacterial agent. PLoS ONE 2017, 12, e0183457. [Google Scholar] [CrossRef]
- Khazandi, M.; Pi, H.; Chan, W.Y.; Ogunniyi, A.D.; Sim, J.X.F.; Venter, H.; Garg, S.; Page, S.W.; Hill, P.B.; McCluskey, A. In vitro Antimicrobial Activity of Robenidine, Ethylenediaminetetraacetic Acid and Polymyxin B Nonapeptide Against Important Human and Veterinary Pathogens. Front. Microbiol. 2019, 10, 837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venter, H. Reversing resistance to counter antimicrobial resistance in the World Health Organisation’s critical priority of most dangerous pathogens. Biosci. Rep. 2019, 39, BSR20180474. [Google Scholar] [CrossRef] [PubMed]
- Pi, H.; Nguyen, H.T.; Venter, H.; Boileau, A.R.; Woolford, L.; Garg, S.; Page, S.W.; Russell, C.C.; Baker, J.R.; McCluskey, A.; et al. In vitro Activity of Robenidine Analog NCL195 in Combination With Outer Membrane Permeabilizers Against Gram-Negative Bacterial Pathogens and Impact on Systemic Gram-Positive Bacterial Infection in Mice. Front. Microbiol. 2020, 11, 1556. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Venter, H.; Veltman, T.; Williams, R.; O’Donovan, L.A.; Russell, C.C.; McCluskey, A.; Page, S.W.; Ogunniyi, A.D.; Trott, D.J. In vitro synergistic activity of NCL195 in combination with colistin against Gram-negative bacterial pathogens. Int. J. Antimicrob. Agents 2021, 57, 106323. [Google Scholar] [CrossRef]
- Nguyen, H.T.; O’Donovan, L.A.; Venter, H.; Russell, C.C.; McCluskey, A.; Page, S.W.; Trott, D.J.; Ogunniyi, A.D. Comparison of Two Transmission Electron Microscopy Methods to Visualize Drug-Induced Alterations of Gram-Negative Bacterial Morphology. Antibiotics 2021, 10, 307. [Google Scholar] [CrossRef]
- Russell, C.C.; Stevens, A.; Pi, H.; Khazandi, M.; Ogunniyi, A.D.; Young, K.A.; Baker, J.R.; McCluskey, S.N.; Page, S.W.; Trott, D.J.; et al. Gram-Positive and Gram-Negative Antibiotic Activity of Asymmetric and Monomeric Robenidine Analogues. ChemMedChem 2018, 13, 2573–2580. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.C.; Stevens, A.; Young, K.A.; Baker, J.R.; McCluskey, S.N.; Khazandi, M.; Pi, H.; Ogunniyi, A.; Page, S.W.; Trott, D.J.; et al. Discovery of 4,6-bis(2-((E)-benzylidene)hydrazinyl)pyrimidin-2-Amine with Antibiotic Activity. ChemistryOpen 2019, 8, 896–907. [Google Scholar] [CrossRef]
- Saputra, S.; Jordan, D.; Mitchell, T.; Wong, H.S.; Abraham, R.J.; Kidsley, A.; Turnidge, J.; Trott, D.J.; Abraham, S. Antimicrobial resistance in clinical Escherichia coli isolated from companion animals in Australia. Vet. Microbiol. 2017, 211, 43–50. [Google Scholar] [CrossRef]
- Abraham, S.; Jordan, D.; Wong, H.S.; Johnson, J.R.; Toleman, M.A.; Wakeham, D.L.; Gordon, D.M.; Turnidge, J.D.; Mollinger, J.L.; Gibson, J.S.; et al. First detection of extended-spectrum cephalosporin- and fluoroquinolone-resistant Escherichia coli in Australian food-producing animals. J. Glob. Antimicrob. Resist. 2015, 3, 273–277. [Google Scholar] [CrossRef]
- Wang, Y.; Alenazy, R.; Gu, X.; Polyak, S.W.; Zhang, P.; Sykes, M.J.; Zhang, N.; Venter, H.; Ma, S. Design and structural optimization of novel 2H-benzo[h]chromene derivatives that target AcrB and reverse bacterial multidrug resistance. Eur. J. Med. Chem. 2021, 213, 113049. [Google Scholar] [CrossRef]
- Wang, Y.; Mowla, R.; Ji, S.; Guo, L.; De Barros Lopes, M.A.; Jin, C.; Song, D.; Ma, S.; Venter, H. Design, synthesis and biological activity evaluation of novel 4-subtituted 2-naphthamide derivatives as AcrB inhibitors. Eur. J. Med. Chem. 2018, 143, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Ohene-Agyei, T.; Mowla, R.; Rahman, T.; Venter, H. Phytochemicals increase the antibacterial activity of antibiotics by acting on a drug efflux pump. Microbiologyopen 2014, 3, 885–896. [Google Scholar] [CrossRef] [PubMed]
- CLSI Standard M100; Performance Standards for Antimicrobial Susceptibility Testing. 27th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017.
- CLSI Standard VET01; Performance Standards for Antimicrobial Disk and Dilution Susceptbility Tests for Bacteria Isolated from Animals. 5th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018.
- Hamoud, R.; Reichling, J.; Wink, M. Synergistic antibacterial activity of the combination of the alkaloid sanguinarine with EDTA and the antibiotic streptomycin against multidrug resistant bacteria. J. Pharm. Pharmacol. 2015, 67, 264–273. [Google Scholar] [CrossRef] [PubMed]
- Palomino, J.C.; Martin, A.; Camacho, M.; Guerra, H.; Swings, J.; Portaels, F. Resazurin microtiter assay plate: Simple and inexpensive method for detection of drug resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2002, 46, 2720–2722. [Google Scholar] [CrossRef]
- Pi, H.; Ogunniyi, A.D.; Savaliya, B.; Nguyen, H.T.; Page, S.W.; Lacey, E.; Venter, H.; Trott, D.J. Repurposing of the Fasciolicide Triclabendazole to Treat Infections Caused by Staphylococcus spp. and Vancomycin-Resistant Enterococci. Microorganisms 2021, 9, 1697. [Google Scholar] [CrossRef]
- Lin, Y.-W.; Han, M.-L.; Zhao, J.; Zhu, Y.; Rao, G.; Forrest, A.; Song, J.; Kaye, K.; Hertzong, P.; Purcell, A.W. Synergistic Combination of Polymyxin B and Enrofloxacin Induced Metabolic Perturbations in Extensive Drug-resistant Pseudomonas aeruginosa. Front. Pharmacol. 2019, 10, 1146. [Google Scholar] [CrossRef]
- Lin, Y.-W.; Heidi, H.Y.; Zhao, J.; Han, M.-L.; Zhu, Y.; Akter, J.; Wickremasinghe, H.; Walpola, H.; Wirth, V.; Rao, G.G. Polymyxin B in combination with enrofloxacin exerts synergistic killing against extensively drug-resistant Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, e00028-18. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Venter, H.; Woolford, L.; Young, K.; McCluskey, A.; Garg, S.; Page, S.W.; Trott, D.J.; Ogunniyi, A.D. Impact of a Novel Anticoccidial Analogue on Systemic Staphylococcus aureus Infection in a Bioluminescent Mouse Model. Antibiotics 2022, 11, 65. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, L.; Molin, S. Synergistic activities of an efflux pump inhibitor and iron chelators against Pseudomonas aeruginosa growth and biofilm formation. Antimicrob. Agents Chemother 2010, 54, 3960–3963. [Google Scholar] [CrossRef]
- Nang, S.C.; Azad, M.A.K.; Velkov, T.; Zhou, Q.T.; Li, J. Rescuing the Last-Line Polymyxins: Achievements and Challenges. Pharm. Rev 2021, 73, 679–728. [Google Scholar] [CrossRef]
- Van Bambeke, F.; Balzi, E.; Tulkens, P.M. Antibiotic efflux pumps. Biochem. Pharmacol. 2000, 60, 457–470. [Google Scholar] [CrossRef]
- Venter, H.; Mowla, R.; Ohene-Agyei, T.; Ma, S. RND-type drug efflux pumps from Gram-negative bacteria: Molecular mechanism and inhibition. Front. Microbiol. 2015, 6, 377. [Google Scholar] [CrossRef] [PubMed]
- Yu, E.W.; Aires, J.R.; Nikaido, H. AcrB Multidrug Efflux Pump of Escherichia coli: Composite Substrate-Binding Cavity of Exceptional Flexibility Generates Its Extremely Wide Substrate Specificity. J. Bacteriol. 2003, 185, 5657–5664. [Google Scholar] [CrossRef] [PubMed]
- Abraham, R.J.; Abraham, S.; Stevens, A.J.; Page, S.W.; McCluskey, A.; Trott, D.J.; O’Handley, R.M. Aminoguanidines: New leads for treatment of Giardia duodenalis infection. Int. J. Parasitol. Drugs Drug Resist. 2019, 10, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Jiang, T.; Zou, Y.; Wang, Y.; Zhou, J.; Li, J.; Liu, L.; Tan, J.; Wei, L.; Li, J.; et al. FDA Approved Drug Library Screening Identifies Robenidine as a Repositionable Antifungal. Front. Microbiol. 2020, 11, 996. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Advisory Group on Integrated Surveillance of Antimicrobial Resistance (AGISAR): Critically Important Antimicrobials for Human Medicine 6th Revision 2018; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Willyard, C. The drug-resistant bacteria that pose the greatest health threats. Nature 2017, 543, 15. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isolates | 1 MIC (μg/mL) | 2 Combination Effect (FICI) | 3 Dose Reduction Index (DRI) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Single Drug | Combination | |||||||||
| PMB | NCL259 | NCL265 | PMB | NCL259 | NCL265 | NCL259 | NCL265 | PMB:NCL259 | PMB:NCL265 | |
| E. coli ATCC 25922 | 0.5 | 16 | 4 | 0.25 | 1 | 1 | Additivity (0.563) | Additivity (0.75) | 2:16 | 2:4 |
| E. coli ATCC 10763 | 1 | 64 | 64 | 0.5 | 0.25 | 0.25 | Additivity (0.504) | Synergism (0.254) | 2:256 | 4:256 |
| P. aeruginosa PAO1 | 1 | 64 | 64 | 0.5 | 1 | 0.25 | Additivity (0.516) | Additivity (0.504) | 2:64 | 2:256 |
| P. aeruginosa ATCC 27853 | 1 | 64 | 32 | 0.5 | 2 | 0.5 | Additivity (0.531) | Additivity (0.516) | 2:32 | 2:64 |
| A.baumannii ATCC 12457 | 1 | 8 | 2 | 0.5 | 1 | 0.5 | Additivity (0.625) | Additivity (0.75) | 2:8 | 2:4 |
| A.baumannii ATCC 19606 | 1 | 8 | 2 | 0.5 | 1 | 0.5 | Additivity (0.625) | Additivity (0.75) | 2:8 | 2:4 |
| K. pneumoniae ATCC 4352 | 1 | 8 | 2 | 0.25 | 1 | 0.5 | Synergism (0.375) | Synergism (0.5) | 4:8 | 4:4 |
| K. pneumoniae ATCC 33495 | 1 | 64 | 64 | 0.5 | 1 | 2 | Additivity (0.516) | Synergism (0.281) | 2:64 | 4:32 |
| Isolates | NCL259 | NCL265 | ||||
|---|---|---|---|---|---|---|
| 1 MIC Values (μg/mL) | MIC Values (μg/mL) | |||||
| MIC Range | MIC50 | MIC90 | MIC Range | MIC50 | MIC90 | |
| Companion animal 2 ExPEC (n = 51) | 16–64 | 32 | 32 | 4–16 | 32 | 32 |
| Porcine/bovine 3 ETEC (n = 83) | 4–64 | 8 | 8 | 2–16 | 8 | 8 |
| Isolates | 1 MIC (μg/mL) | Combination Effect (2 FICI) | 3 DRI PAβN:NCL259 | |||
|---|---|---|---|---|---|---|
| Single Drug | Combination | |||||
| PAβN | NCL259 | PAβN | NCL259 | |||
| K. pneumoniae 13GNB–429 | >32 | >256 | 16 | 16 | Synergism (0.281) | 4:16 |
| K. pneumoniae 13GNB–550 | >32 | >256 | 32 | 8 | Additivity (0.531) | 2:32 |
| K. oxytoca 13GNB–582 | >32 | >256 | 16 | 8 | Synergism (0.281) | 4:32 |
| Isolates | 1 MIC (μg/mL) | Combination Effect (2 FICI) | 3 DRI PAβN:NCL265 | |||
|---|---|---|---|---|---|---|
| Single Drug | Combination | |||||
| PAβN | NCL265 | PAβN | NCL265 | |||
| K. pneumoniae 13GNB–429 | >32 | >256 | 32 | 4 | Additivity (0.516) | 2:64 |
| K. pneumoniae 13GNB–550 | >32 | >256 | 32 | 4 | Additivity (0.516) | 2:64 |
| K. oxytoca 13GNB–582 | >32 | >256 | 32 | 2 | Additivity (0.508) | 2:128 |
| Isolates | 1 MIC Values (μg/mL) | ||
|---|---|---|---|
| NCL259 | NCL265 | Erythromycin | |
| Wild-type (E. coli BW 25113) | 64 | 16 | >128 |
| E. coli BW 25113 (∆AcrB) | 4 | 1 | 2 |
| Cell Line | 1 IC50 or 2 HC50 Values (μg/mL) 3 for: | |||
|---|---|---|---|---|
| NCL812 | NCL259 | NCL265 | Ampicillin | |
| 1 Caco-2 | 8, 12 | <1, 3 | <1, <2 | >128, >128 |
| 1 HEL 299 | 8, 6 | <2, <2 | <2, <2 | >128, >128 |
| 1 Hep G2 | 6, 8 | <1, <1 | <1, <1 | >128, >128 |
| 1 MCF-7 | 8, 12 | 3, 6 | <1, 6 | >128, >128 |
| 1 MDBK | 8, 12 | 2, 2 | 3, 2 | >128, >128 |
| 1 Vero | 8, 4 | 6, 3 | 6, 3 | >128, >128 |
| 2 SRBC | 32, 32 | 16, 32 | 16, 32 | >128, >128 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pi, H.; Venter, H.; Russell, C.C.; Young, K.A.; McCluskey, A.; Page, S.W.; Ogunniyi, A.D.; Trott, D.J. In Vitro Activity of Robenidine Analogues NCL259 and NCL265 against Gram-Negative Pathogens. Antibiotics 2022, 11, 1301. https://doi.org/10.3390/antibiotics11101301
Pi H, Venter H, Russell CC, Young KA, McCluskey A, Page SW, Ogunniyi AD, Trott DJ. In Vitro Activity of Robenidine Analogues NCL259 and NCL265 against Gram-Negative Pathogens. Antibiotics. 2022; 11(10):1301. https://doi.org/10.3390/antibiotics11101301
Chicago/Turabian StylePi, Hongfei, Henrietta Venter, Cecilia C. Russell, Kelly A. Young, Adam McCluskey, Stephen W. Page, Abiodun D. Ogunniyi, and Darren J. Trott. 2022. "In Vitro Activity of Robenidine Analogues NCL259 and NCL265 against Gram-Negative Pathogens" Antibiotics 11, no. 10: 1301. https://doi.org/10.3390/antibiotics11101301