

Detection of Acquired Antibiotic Resistance Genes in Domestic Pig (Sus scrofa) and Common Carp (Cyprinus carpio) Intestinal Samples by Metagenomics Analyses in Hungary
 , ,
, ,  ,
,  , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Results
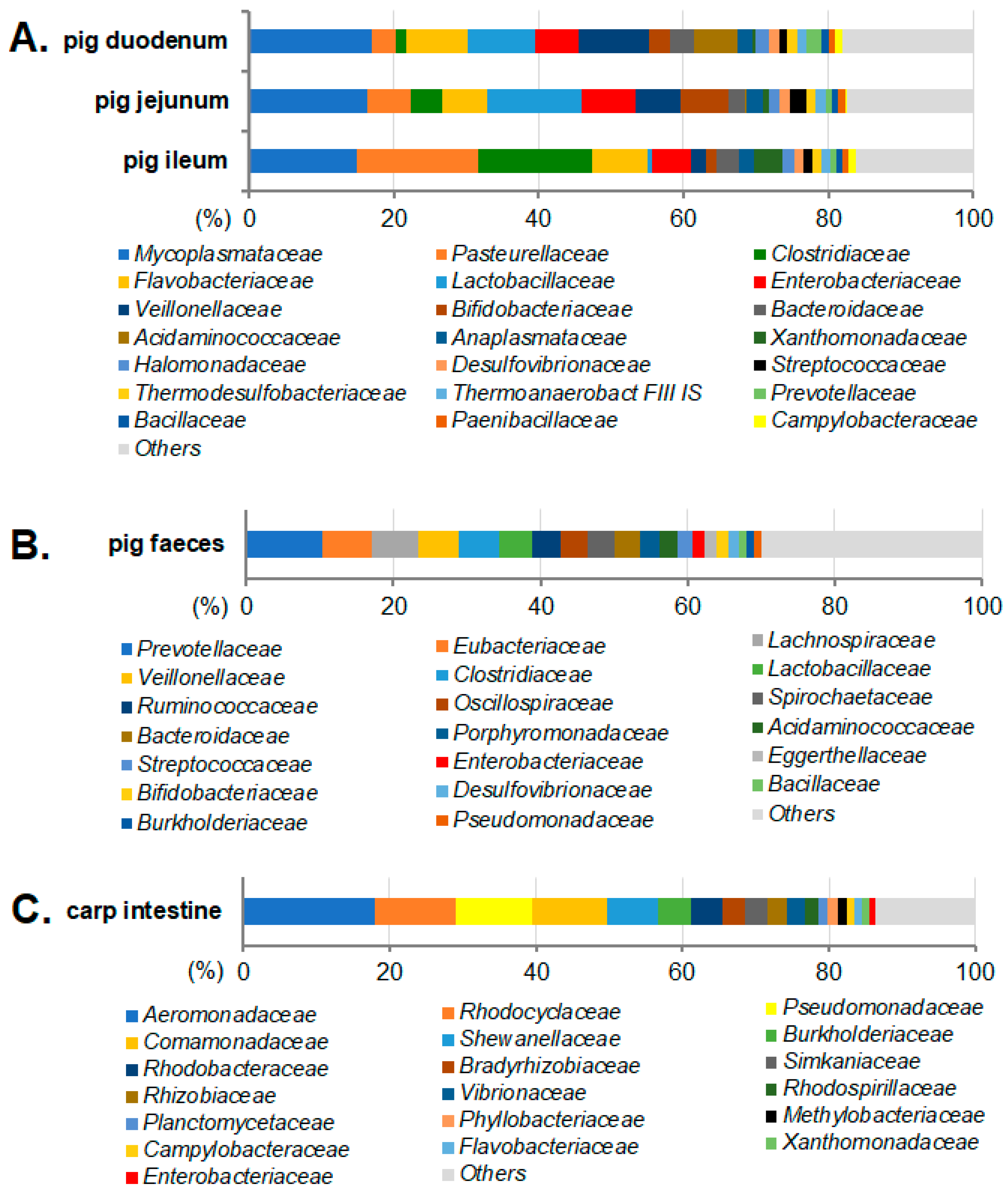
2.1. Taxonomic Assignment of the Metagenomic Contigs to Bacterial Families
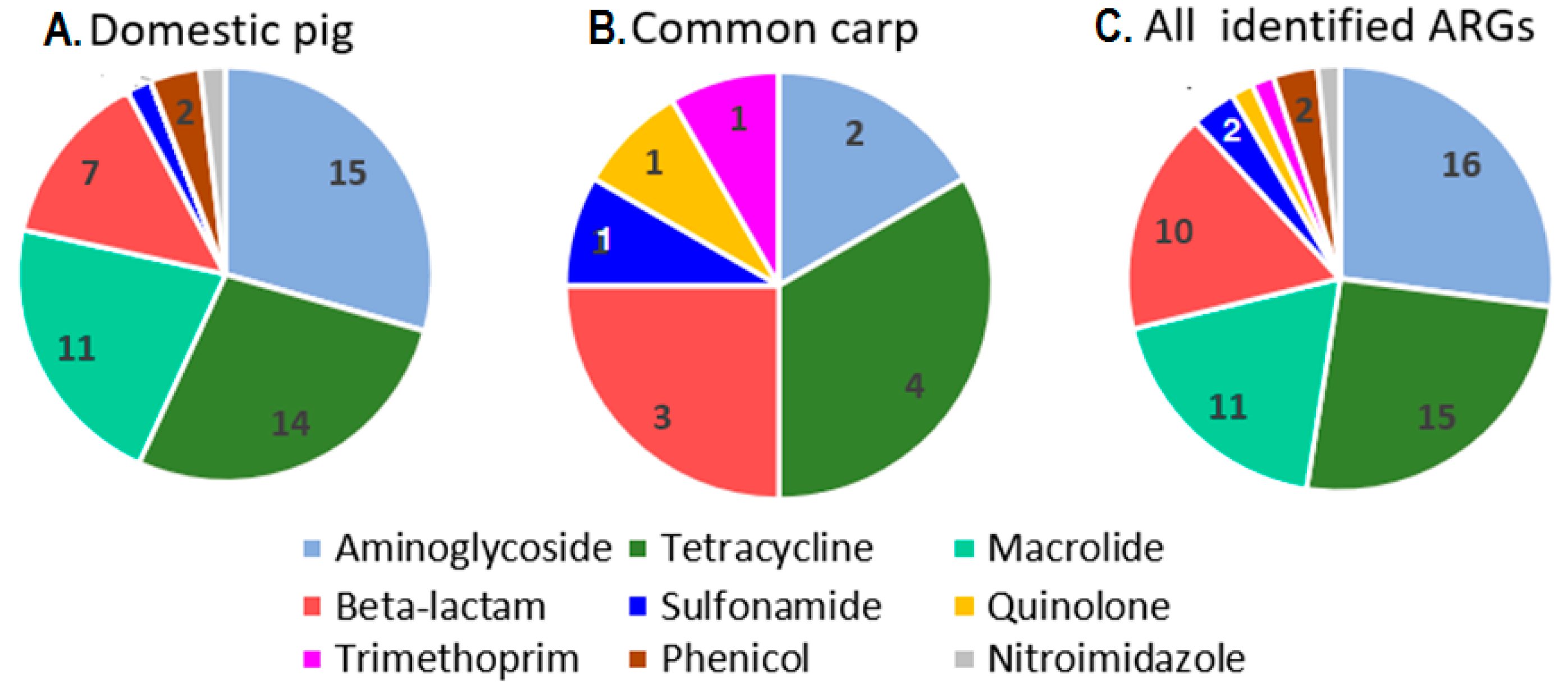
2.2. Identification of Acquired ARG Types in Domestic Pig Intestinal Microbiome
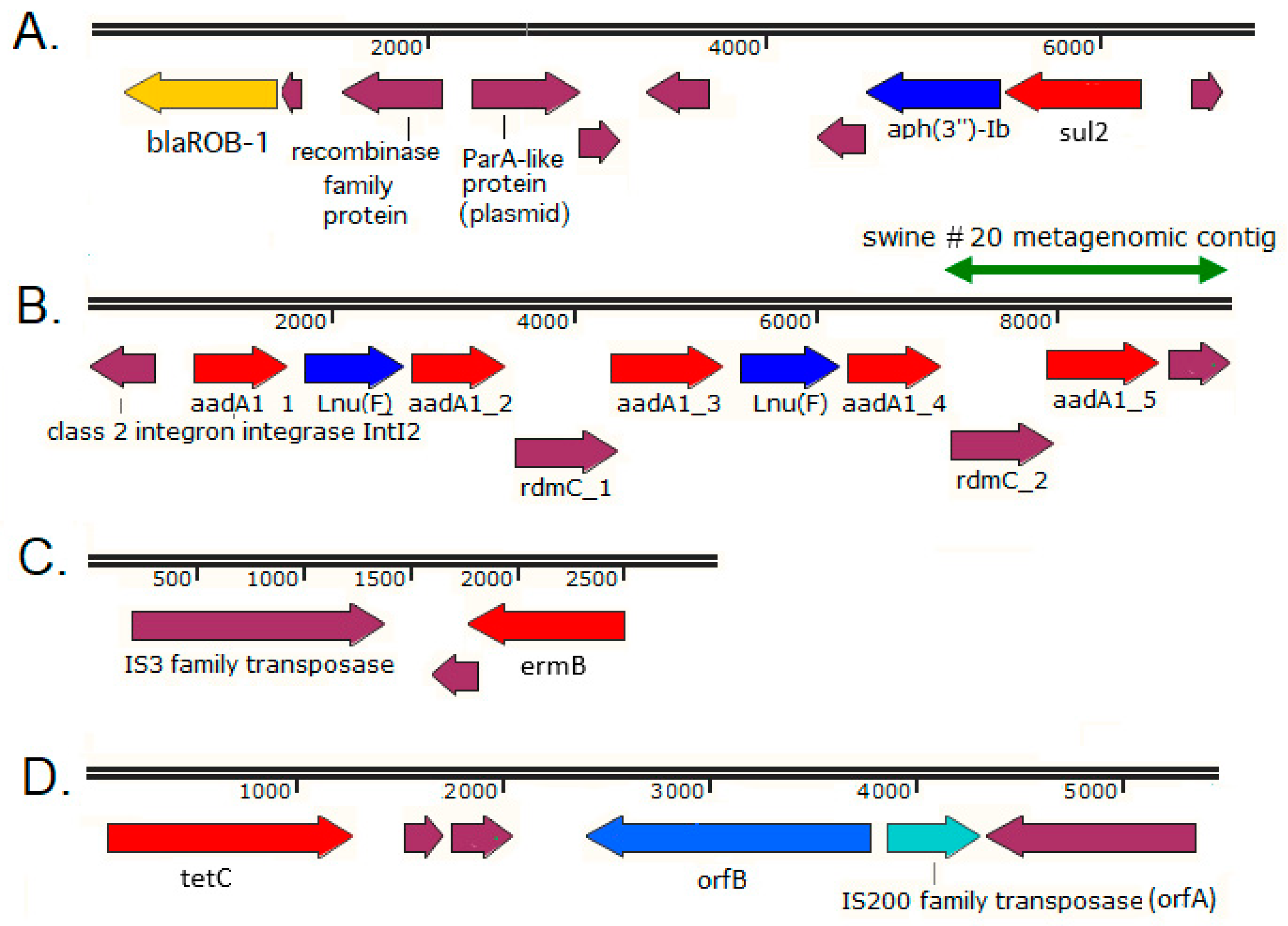
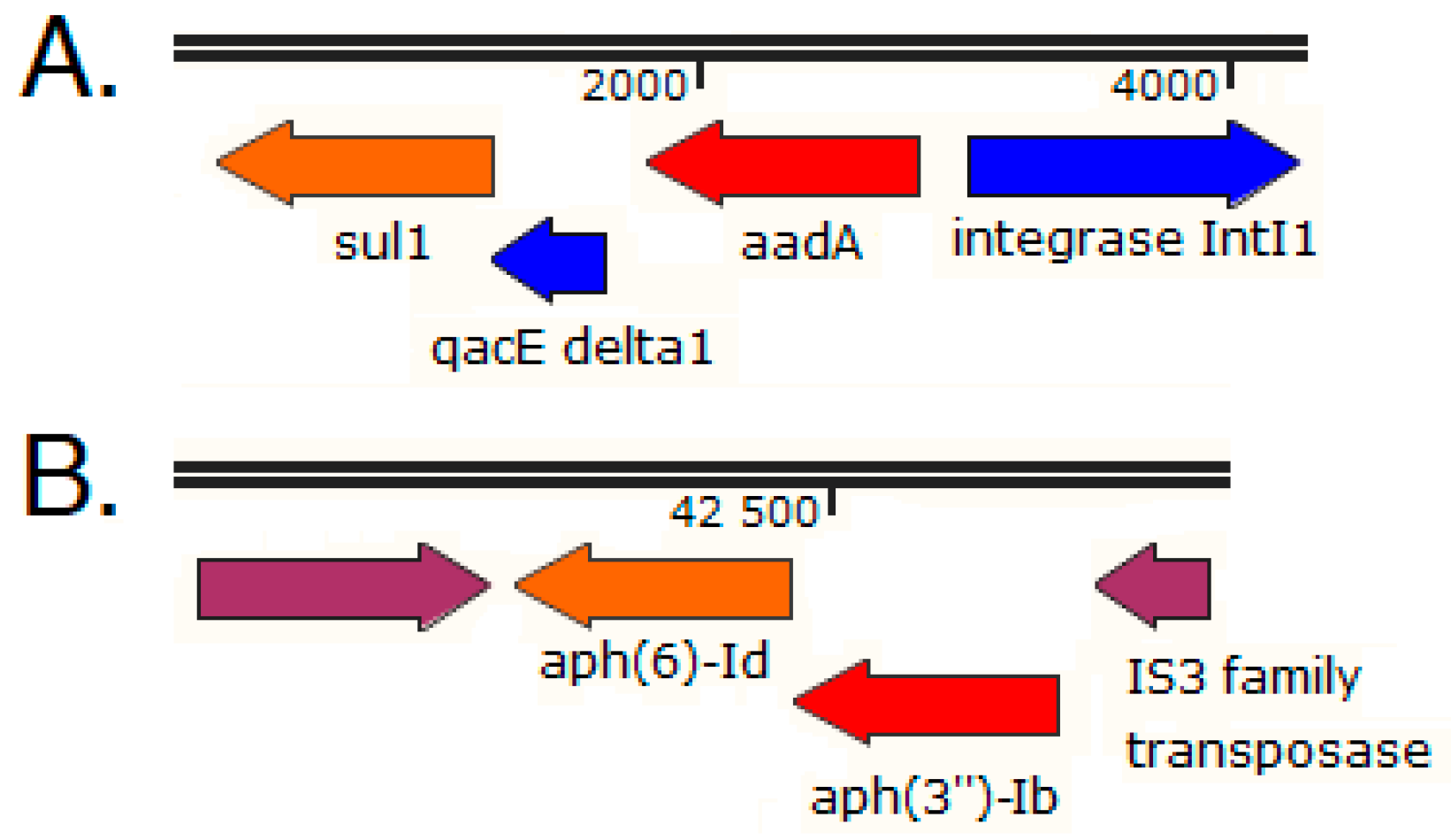
2.3. Immediate Genetic Environment of ARGs on Selected Swine Gut Metagenomic Contigs
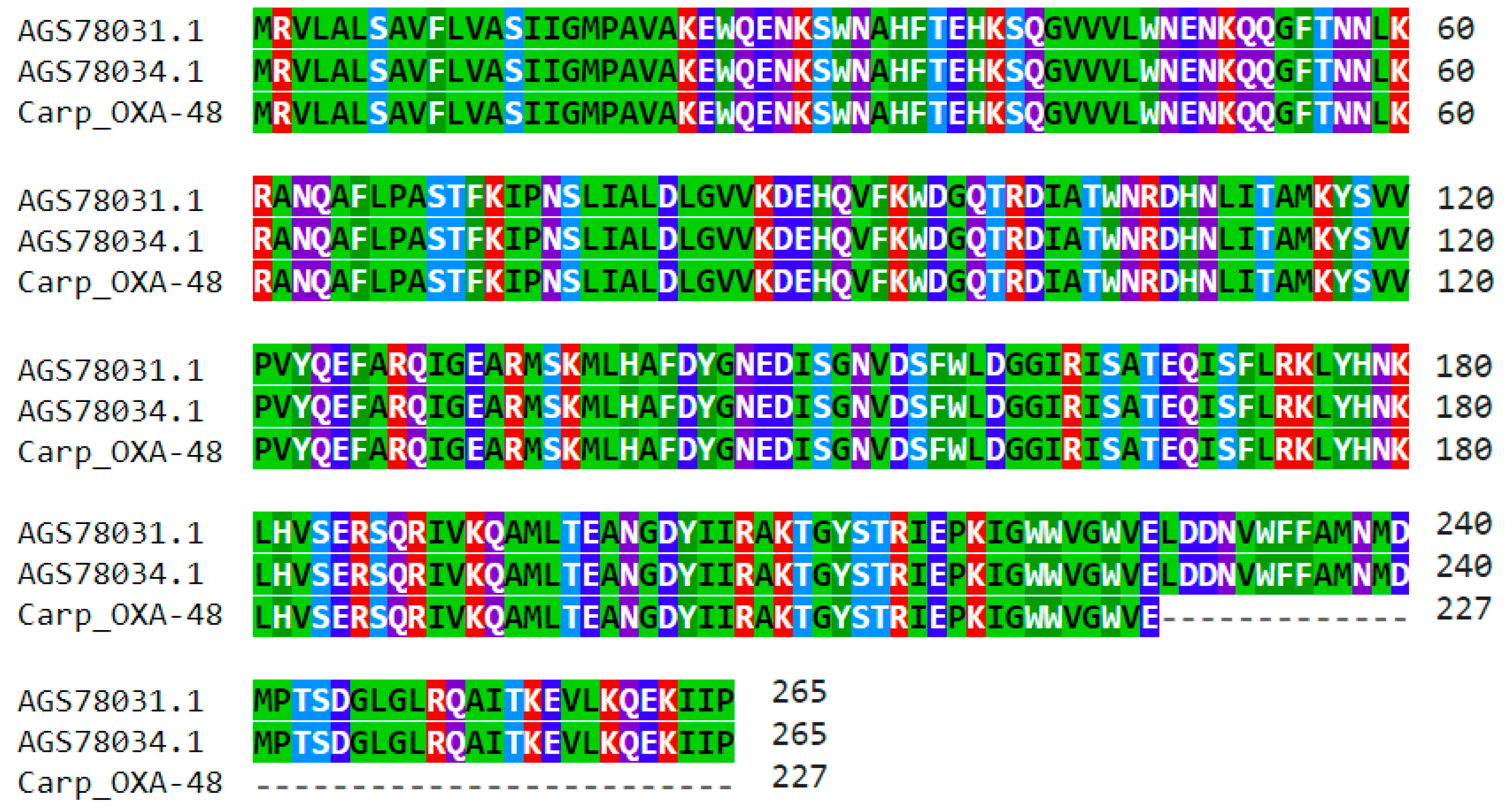
2.4. Identification of Acquired ARGs in the Carp Intestinal Microbiome
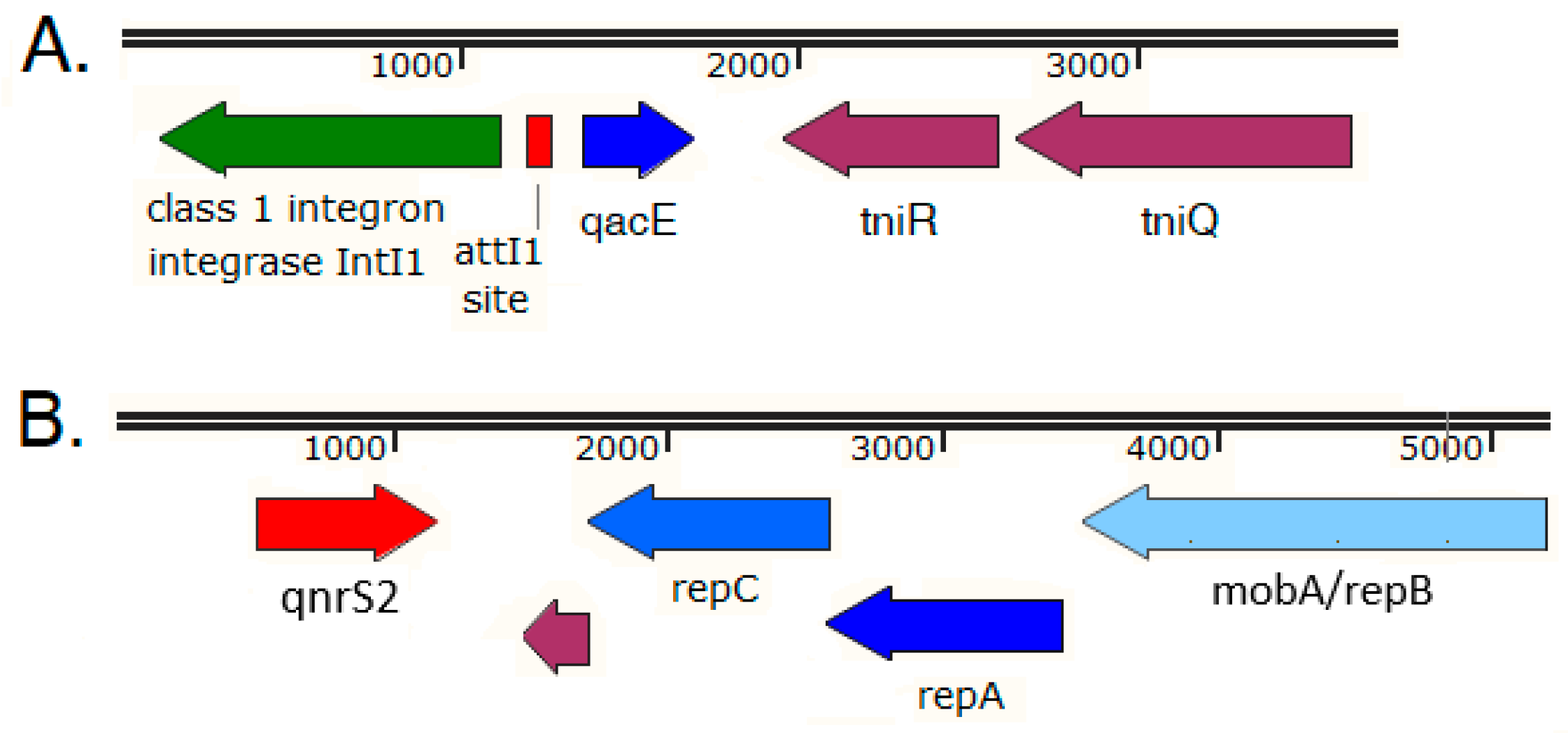
2.5. Analysis of the Immediate Genetic Environment of ARGs on Selected Carp Intestinal Metagenomic Contigs
3. Discussion
4. Materials and Methods
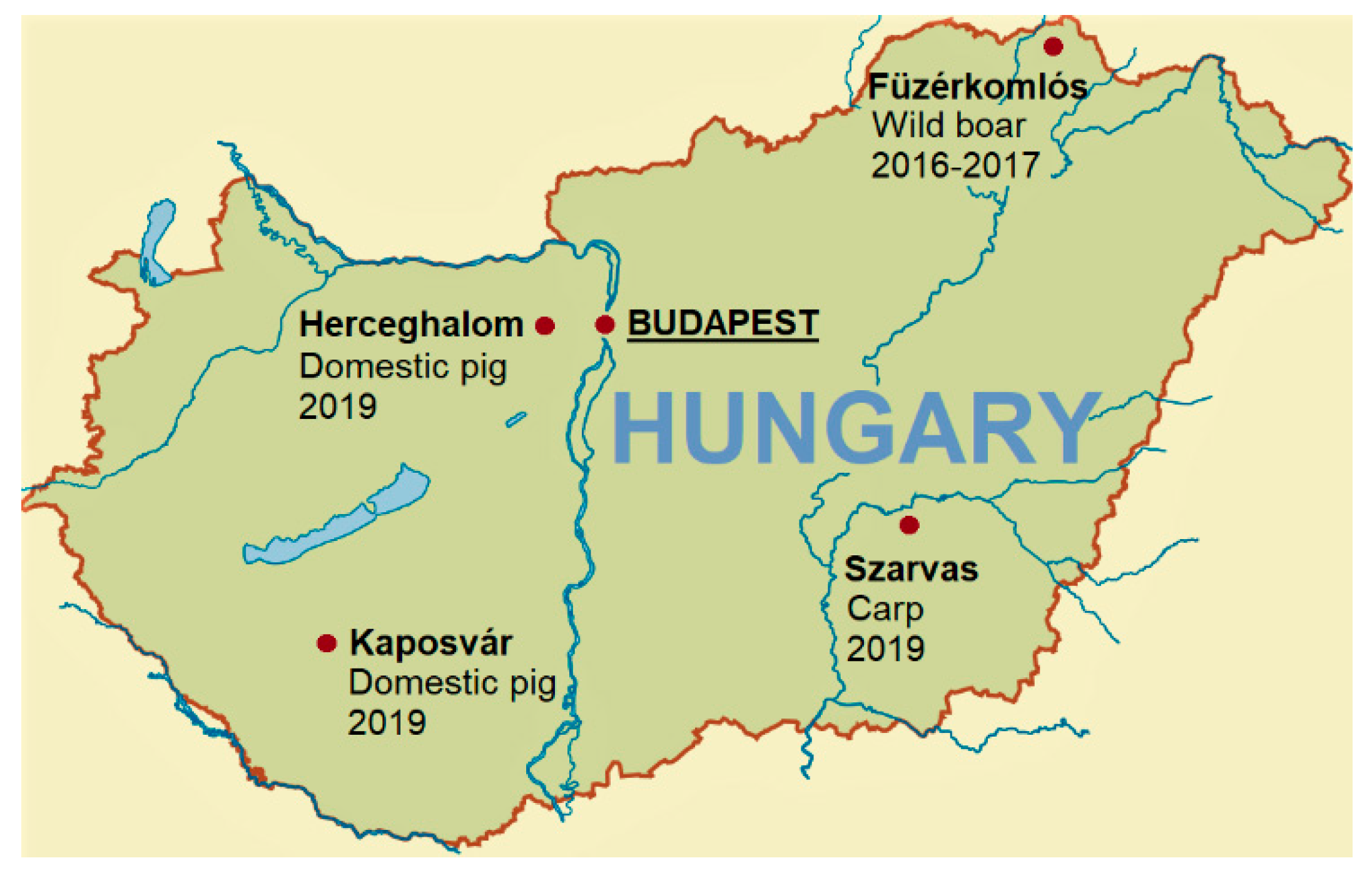
4.1. Sample Collection and Processing
4.2. DNA Purification and Metagenomic Sequencing
4.3. Metagenomic Contig Assembly
4.4. Identification of ARGs Harbored by the Metagenomic Contigs
4.5. Annotation and Functional Analysis of Selected Metagenomic Contigs
4.6. Taxonomic Classification of the Assembled Metagenomic Contigs
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carvalho, I.T.; Santos, L. Antibiotics in the aquatic environments: A review of the European scenario. Environ. Int. 2016, 94, 736–757. [Google Scholar] [CrossRef] [PubMed]
- Everaert, N.; Van Cruchten, S.; Weström, B.; Bailey, M.; Van Ginneken, C.; Thymann, T.; Pieper, R. A review on early gut maturation and colonization in pigs, including biological and dietary factors affecting gut homeostasis. Anim. Feed Sci. Technol. 2017, 233, 89–103. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.A.; Looft, T.; Severin, A.J.; Bayles, D.O.; Nasko, D.J.; Wommack, K.E.; Howe, A.; Allen, H.K. The in-feed antibiotic carbadox induces phage gene transcription in the swine gut microbiome. mBio 2017, 8, e00709-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control (ECDC); European Food Safety Authority (EFSA); European Medicines Agency (EMA). Third joint inter-agency report on integrated analysis of consumption of antimicrobial agents and occurrence of antimicrobial resistance in bacteria from humans and food-producing animals in the EU/EEA: JIACRA III 2016–2018. EFSA J. 2021, 19, e06712. [Google Scholar] [CrossRef]
- Dunay, A.; Vinkler-Rajcsányi, K. Hungarian pig sector: Actual problems and prospects for the future development. Acta Univ. Agric. Silvic. Mendel. Brun. 2016, 64, 1879–1888. [Google Scholar] [CrossRef] [Green Version]
- Karnai, L.; Szűcs, I. Outlooks and perspectives of the common carp production. Rocz. Nauk. Stow. Ekon. Rol. Agrobiz. 2018, XX, 64–72. [Google Scholar] [CrossRef]
- Pennone, V.; Cobo-Díaz, J.F.; Prieto, M.; Alvarez-Ordóñez, A. Application of genomics and metagenomics to improve food safety based on an enhanced characterisation of antimicrobial resistance. Curr. Opin. Food Sci. 2022, 43, 183–188. [Google Scholar] [CrossRef]
- European Medicines Agency. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2019 and 2020 (11th ESVAC Report); European Medicines Agency: Amsterdam, The Netherlands, 2021.
- D’Costa, V.M.; Griffiths, E.; Wright, G.D. Expanding the soil antibiotic resistome: Exploring environmental diversity. Curr. Opin. Microbiol. 2007, 10, 481–489. [Google Scholar] [CrossRef]
- Founou, L.L.; Founou, R.C.; Essack, S.Y. Antibiotic Resistance in the Food Chain: A Developing Country-Perspective. Front. Microbiol. 2016, 7, 1881. [Google Scholar] [CrossRef]
- Looft, T.; Johnson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef]
- Van den Meersche, T.; Rasschaert, G.; Vanden Nest, T.; Haesebrouck, F.; Herman, L.; Van Coillie, E.; Van Weyenberg, S.; Daeseleire, E.; Heyndrickx, M. Longitudinal screening of antibiotic residues, antibiotic resistance genes and zoonotic bacteria in soils fertilized with pig manure. Environ. Sci. Pollut. Res. Int. 2020, 27, 28016–28029. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, P.J.; Townsend, J.P.; Bøhn, T.; Simonsen, G.S.; Sundsfjord, A.; Nielsen, K.M. Factors affecting the reversal of antimicrobial-drug resistance. Lancet Infect. Dis. 2009, 9, 357–364. [Google Scholar] [CrossRef]
- Yang, D.; Heederik, D.; Mevius, D.J.; Scherpenisse, P.; Luiken, R.; Van Gompel, L.; Skarżyńska, M.; Wadepohl, K.; Chauvin, C.; Van Heijnsbergen, E.; et al. Risk factors for the abundance of antimicrobial resistance genes aph(3′)-III, erm(B), sul2 and tet(W) in pig and broiler faeces in nine European countries. J. Antimicrob. Chemother. 2022, 77, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.; Ramos, F. Antimicrobial resistance in aquaculture: Current knowledge and alternatives to tackle the problem. Int. J. Antimicrob. Agents 2018, 52, 135–143. [Google Scholar] [CrossRef]
- Preena, P.G.; Swaminathan, T.R.; Kumar, V.J.R.; Singh, I.S.B. Antimicrobial resistance in aquaculture: A crisis for concern. Biologia 2020, 75, 1497–1517. [Google Scholar] [CrossRef]
- Sándor, Z.J.; Papp, Z.G.; Kosáros, T.J.; Hegedűs, R.; Csengeri, I. Potential effects of pharmaceuticals and their residues in aquatic environment. Stud. Univ. Vasile Goldis Arad Ser. Stiint. Vietii Life Sci. Ser. 2012, 22, 247–255. [Google Scholar]
- Burridge, L.; Weis, J.S.; Cabello, F.; Pizarro, J.; Bostick, K. Chemical use in salmon aquaculture: A review of current practices and possible environmental effects. Aquaculture 2010, 306, 7–23. [Google Scholar] [CrossRef]
- European Commission. A European One Health Action Plan against Antimicrobial Resistance; European Commission: Brussels, Belgium, 2017.
- Huygens, J.; Rasschaert, G.; Heyndrickx, M.; Dewulf, J.; Van Coillie, E.; Quataert, P.; Daeseleire, E.; Becue, I. Impact of fertilization with pig or calf slurry on antibiotic residues and resistance genes in the soil. Sci. Total Environ. 2022, 822, 153518. [Google Scholar] [CrossRef]
- Rasschaert, G.; Elst, D.V.; Colson, L.; Herman, L.; Ferreira, H.; Dewulf, J.; Decrop, J.; Meirlaen, J.; Heyndrickx, M.; Daeseleire, A.E. Antibiotic Residues and Antibiotic-Resistant Bacteria in Pig Slurry used to Fertilize Agricultural Fields. Antibiotics 2020, 9, 34. [Google Scholar] [CrossRef] [Green Version]
- Libisch, B.; Keresztény, T.; Kerényi, Z.; Kocsis, R.; Sipos, R.; Papp, P.P.; Olasz, F. Metagenomic analysis of acquired antibiotic resistance determinants in the gut microbiota of wild boars (Sus scrofa)—Preliminary results. J. Vet. Res. 2020, 64, 111–118. [Google Scholar] [CrossRef]
- Yang, D.; Heederik, D.; Scherpenisse, P.; Van Gompel, L.; Luiken, R.; Wadepohl, K.; Skarżyńska, M.; Van Heijnsbergen, E.; Wouters, I.M.; Greve, G.D.; et al. Antimicrobial resistance genes aph (3′)-III, erm(B), sul2 and tet(W) abundance in animal faeces, meat, production environments and human faeces in Europe. J. Antimicrob. Chemother. 2022, 77, 1883–1893. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); Aerts, M.; Battisti, A.; Hendriksen, R.; Kempf, I.; Teale, C.; Tenhagen, B.A.; Veldman, K.; Wasyl, D.; Guerra, B.; et al. Technical specifications on harmonised monitoring of antimicrobial resistance in zoonotic and indicator bacteria from food-producing animals and food. EFSA J. 2019, 17, e05709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto-Maradona, M.; Alvarez-Ordóñez, A. Identification of risk factors and hotspots of antibiotic resistance along the food chain using next-generation sequencing. EFSA J. 2020, 18 (Suppl. S1), e181107. [Google Scholar] [CrossRef] [PubMed]
- Lal Gupta, C.; Kumar Tiwari, R.; Cytryn, E. Platforms for elucidating antibiotic resistance in single genomes and complex metagenomes. Environ. Int. 2020, 138, 105667. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (EFSA BIOHAZ Panel); Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al. Whole genome sequencing and metagenomics for outbreak investigation, source attribution and risk assessment of food-borne microorganisms. EFSA J. 2019, 17, e05898. [Google Scholar] [CrossRef] [Green Version]
- Libisch, B.; Picot, C.; Ceballos-Garzon, A.; Moravkova, M.; Klimesová, M.; Telkes, G.; Chuang, S.T.; Le Pape, P. Prototheca Infections and Ecology from a One Health Perspective. Microorganisms 2022, 10, 938. [Google Scholar] [CrossRef]
- Hendriksen, R.S.; Bortolaia, V.; Tate, H.; Tyson, G.H.; Aarestrup, F.M.; McDermott, P.F. Using Genomics to Track Global Antimicrobial Resistance. Front. Public Health 2019, 7, 242. [Google Scholar] [CrossRef] [Green Version]
- Munk, P.; Knudsen, B.E.; Lukjancenko, O.; Duarte, A.; Van Gompel, L.; Luiken, R.; Smit, L.; Schmitt, H.; Garcia, A.D.; Hansen, R.B.; et al. Abundance and diversity of the faecal resistome in slaughter pigs and broilers in nine European countries. Nat. Microbiol. 2018, 3, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Van Gompel, L.; Luiken, R.; Sarrazin, S.; Munk, P.; Knudsen, B.E.; Hansen, R.B.; Bossers, A.; Aarestrup, F.M.; Dewulf, J.; Wagenaar, J.A.; et al. The antimicrobial resistome in relation to antimicrobial use and biosecurity in pig farming, a metagenome-wide association study in nine European countries. J. Antimicrob. Chemother. 2019, 74, 865–876. [Google Scholar] [CrossRef]
- Doyle, R.M.; O’Sullivan, D.M.; Aller, S.D.; Bruchmann, S.; Clark, T.; Coello Pelegrin, A.; Cormican, M.; Diez Be-navente, E.; Ellington, M.J.; McGrath, E.; et al. Discordant bioinformatic predictions of antimicrobial resistance from whole-genome sequencing data of bacterial isolates: An inter-laboratory study. Microb. Genom. 2020, 6, e000335. [Google Scholar] [CrossRef]
- Matter, D.; Rossano, A.; Sieber, S.; Perreten, V. Small multidrug resistance plasmids in Actinobacillus porcitonsillarum. Plasmid 2008, 59, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Funnell, B.E. Partition-mediated plasmid pairing. Plasmid 2005, 53, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Schuster, C.F.; Weber, R.E.; Weig, M.; Werner, G.; Pfeifer, Y. Ultra-deep long-read sequencing detects IS-mediated gene duplications as a potential trigger to generate arrays of resistance genes and a mechanism to induce novel gene variants such as blaCTX-M-243. J. Antimicrob. Chemother. 2022, 77, 381–390. [Google Scholar] [CrossRef]
- Strøman, P.; Müller, C.C.; Sørensen, K.I. Heat shock treatment increases the frequency of loss of an erythromycin resistance-encoding transposable element from the chromosome of Lactobacillus crispatus CHCC3692. Appl. Environ. Microbiol. 2003, 69, 7173–7180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dugan, J.; Andersen, A.A.; Rockey, D.D. Functional characterization of IScs605, an insertion element carried by tetracycline-resistant Chlamydia suis. Microbiol. 2007, 153, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Dugan, J.; Rockey, D.D.; Jones, L.; Andersen, A.A. Tetracycline resistance in Chlamydia suis mediated by genomic islands inserted into the chlamydial inv-like gene. Antimicrob. Agents Chemother. 2004, 48, 3989–3995. [Google Scholar] [CrossRef] [Green Version]
- Seth-Smith, H.M.; Wanninger, S.; Bachmann, N.; Marti, H.; Qi, W.; Donati, M.; di Francesco, A.; Polkinghorne, A.; Borel, N. The Chlamydia suis Genome Exhibits High Levels of Diversity, Plasticity, and Mobile Antibiotic Resistance: Comparative Genomics of a Recent Livestock Cohort Shows Influence of Treatment Regimes. Genome Biol. Evol. 2017, 9, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Betteridge, T.; Partridge, S.R.; Iredell, J.R.; Stokes, H.W. Genetic context and structural diversity of class 1 integrons from human commensal bacteria in a hospital intensive care unit. Antimicrob. Agents Chemother. 2011, 55, 3939–3943. [Google Scholar] [CrossRef] [Green Version]
- Collis, C.M.; Recchia, G.D.; Kim, M.J.; Stokes, H.W.; Hall, R.M. Efficiency of recombination reactions catalyzed by class 1 integron integrase IntI1. J. Bacteriol. 2001, 183, 2535–2542. [Google Scholar] [CrossRef] [Green Version]
- Partridge, S.R.; Recchia, G.D.; Scaramuzzi, C.; Collis, C.M.; Stokes, H.W.; Hall, R.M. Definition of the attI1 site of class 1 integrons. Microbiology 2000, 146, 2855–2864. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, E.; Sela, N.; Doron-Faigenboim, A.; Navon-Venezia, S.; Jurkevitch, E.; Cytryn, E. Genomic and Functional Characterization of qnr-Encoding Plasmids from Municipal Wastewater Biosolid Klebsiella pneumoniae Isolates. Front. Microbiol. 2015, 6, 1354. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Dziewit, L.; Ostrowski, R.; Chmielowska, C.; Popowska, M. Molecular Characterization and Comparative Genomics of IncQ-3 Plasmids Conferring Resistance to Various Antibiotics Isolated from a Wastewater Treatment Plant in Warsaw (Poland). Antibiotics 2020, 9, 613. [Google Scholar] [CrossRef]
- Loftie-Eaton, W.; Rawlings, D.E. Diversity, biology and evolution of IncQ-family plasmids. Plasmid 2012, 67, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Tacão, M.; Correia, A.; Henriques, I. Environmental Shewanella xiamenensis strains that carry blaOXA-48 or blaOXA-204 genes: Additional proof for blaOXA-48-like gene origin. Antimicrob. Agents Chemother. 2013, 57, 6399–6400. [Google Scholar] [CrossRef] [Green Version]
- De Luca, F.; Benvenuti, M.; Carboni, F.; Pozzi, C.; Rossolini, G.M.; Mangani, S.; Docquier, J.D. Evolution to carbapenem-hydrolyzing activity in noncarbapenemase class D β-lactamase OXA-10 by rational protein design. Proc. Natl. Acad. Sci. USA 2011, 108, 18424–18429. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Animal Health and Welfare (AHAW); Nielsen, S.S.; Bicout, D.J.; Calistri, P.; Canali, E.; Drewe, J.A.; Garin-Bastuji, B.; Gonzales Rojas, J.L.; Gortazar Schmidt, C.; Herskin, M.; et al. Assessment of animal diseases caused by bacteria resistant to antimicrobials: Swine. EFSA J. 2021, 19, e07113. [Google Scholar] [CrossRef]
- de Been, M.; Lanza, V.F.; de Toro, M.; Scharringa, J.; Dohmen, W.; Du, Y.; Hu, J.; Lei, Y.; Li, N.; Tooming-Klunderud, A.; et al. Dissemination of cephalosporin resistance genes between Escherichia coli strains from farm animals and humans by specific plasmid lineages. PLoS Genet. 2014, 10, e1004776. [Google Scholar] [CrossRef] [Green Version]
- Guillard, T.; Pons, S.; Roux, D.; Pier, G.B.; Skurnik, D. Antibiotic resistance and virulence: Understanding the link and its consequences for prophylaxis and therapy. Bioessays 2016, 38, 682–693. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Epidemiology of β-Lactamase-Producing Pathogens. Clin. Microbiol. Rev. 2020, 33, e00047-19. [Google Scholar] [CrossRef]
- Bertran, X.; Rubio, M.; Gómez, L.; Llovet, T.; Muñoz, C.; Navarro, F.; Miro, E. Taxonomic Identification of Different Species of the Genus Aeromonas by Whole-Genome Sequencing and Use of Their Species-Specific β-Lactamases as Phylogenetic Markers. Antibiotics 2021, 10, 354. [Google Scholar] [CrossRef]
- Galán, J.C.; Reig, M.; Navas, A.; Baquero, F.; Blázquez, J. ACI-1 from Acidaminococcus fermentans: Characterization of the first β-lactamase in Anaerobic cocci. Antimicrob. Agents Chemother. 2000, 44, 3144–3149. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, G.; Galán, J.C.; Rodríguez-Alcayna, M.; Moya, A.; Baquero, F.; Latorre, A. Complete genome sequence of Acidaminococcus intestini RYC-MR95, a Gram-negative bacterium from the phylum Firmicutes. J. Bacteriol. 2011, 193, 7008–7009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Liu, C.; Zhang, D.; Wang, R.; Qin, L.; Xu, Q.; Che, L.; Gao, F. Effects of Low-Dose Antibiotics on Gut Immunity and Antibiotic Resistomes in Weaned Piglets. Front. Immunol. 2020, 11, 903. [Google Scholar] [CrossRef]
- Holman, D.B.; Chénier, M.R. Antimicrobial use in swine production and its effect on the swine gut microbiota and antimicrobial resistance. Can. J. Microbiol. 2015, 61, 785–798. [Google Scholar] [CrossRef]
- Szmolka, A.; Anjum, M.F.; La Ragione, R.M.; Kaszanyitzky, E.J.; Nagy, B. Microarray based comparative genotyping of gentamicin resistant Escherichia coli strains from food animals and humans. Vet. Microbiol. 2012, 156, 110–118. [Google Scholar] [CrossRef]
- Szmolka, A.; Lestár, B.; Pászti, J.; Fekete, P.; Nagy, B. Conjugative IncF and IncI1 plasmids with tet(A) and class 1 integron conferring multidrug resistance in F18(+) porcine enterotoxigenic E. coli. Acta Vet. Hung. 2015, 63, 425–443. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Wang, W.; Zhu, X.; Sun, X.; Xiao, J.; Li, D.; Cui, Y.; Wang, C.; Shi, Y. Response of Gut Microbiota to Dietary Fiber and Metabolic Interaction With SCFAs in Piglets. Front. Microbiol. 2018, 9, 2344. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Yu, B.; Chen, D.; Zheng, P.; Luo, Y.; Huang, Z.; Luo, J.; Mao, X.; Yu, J.; He, J. Changes of porcine gut microbiota in response to dietary chlorogenic acid supplementation. Appl. Microbiol. Biotechnol. 2019, 103, 8157–8168. [Google Scholar] [CrossRef]
- Zeng, Q.; Liao, C.; Terhune, J.; Wang, L. Impacts of florfenicol on the microbiota landscape and resistome as revealed by metagenomic analysis. Microbiome 2019, 7, 155. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Lv, Z.; Shen, Y.; Liu, D.; Fu, Y.; Zhou, L.; Liu, W.; Chen, K.; Ye, H.; Xia, X.; et al. Metagenomic insights into differences in environmental resistome profiles between integrated and monoculture aquaculture farms in China. Environ. Int. 2020, 144, 106005. [Google Scholar] [CrossRef]
- Ceccarelli, D.; van Essen-Zandbergen, A.; Veldman, K.T.; Tafro, N.; Haenen, O.; Mevius, D.J. Chromosome-Based blaOXA-48-Like Variants in Shewanella Species Isolates from Food-Producing Animals, Fish, and the Aquatic Environment. Antimicrob. Agents Chemother. 2017, 61, e01013-16. [Google Scholar] [CrossRef] [PubMed]
- Kovács, K.; Nyul, A.; Mestyán, G.; Melegh, S.; Fenyvesi, H.; Jakab, G.; Szabó, H.; Jánvári, L.; Damjanova, I.; Tóth, Á. Emergence and interhospital spread of OXA-48-producing Klebsiella pneumoniae ST395 clone in Western Hungary. Infect. Dis. 2017, 49, 231–233. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M.; Ábrók, M.; Lázár, A.; Jánvári, L.; Tóth, Á.; Terhes, G.; Burián, K. Detection of VIM, NDM and OXA-48 producing carbapenem resistant Enterobacterales among clinical isolates in Southern Hungary. Acta Microbiol. Immunol. Hung. 2020, 67, 209–215. [Google Scholar] [CrossRef]
- Dabos, L.; Oueslati, S.; Bernabeu, S.; Bonnin, R.A.; Dortet, L.; Naas, T. To Be or Not to Be an OXA-48 Carbapenemase. Microorganisms 2022, 10, 258. [Google Scholar] [CrossRef]
- Boyd, S.E.; Holmes, A.; Peck, R.; Livermore, D.M.; Hope, W. OXA-48-Like β-Lactamases: Global Epidemiology, Treatment Options, and Development Pipeline. Antimicrob. Agents Chemother. 2022, 66, e0021622. [Google Scholar] [CrossRef] [PubMed]
- Irrgang, A.; Pauly, N.; Tenhagen, B.A.; Grobbel, M.; Kaesbohrer, A.; Hammerl, A. Spill-Over from Public Health? First Detection of an OXA-48-Producing Escherichia coli in a German Pig Farm. Microorganisms 2020, 8, 855. [Google Scholar] [CrossRef]
- Szmolka, A.; Fortini, D.; Villa, L.; Carattoli, A.; Anjum, M.F.; Nagy, B. First report on IncN plasmid-mediated quinolone resistance gene qnrS1 in porcine Escherichia coli in Europe. Microb. Drug Resist. 2011, 17, 567–573. [Google Scholar] [CrossRef]
- Szabó, D.; Kocsis, B.; Rókusz, L.; Szentandrássy, J.; Katona, K.; Kristóf, K.; Nagy, K. First detection of plasmid-mediated, quinolone resistance determinants qnrA, qnrB, qnrS and aac(6′)-Ib-cr in extended-spectrum beta-lactamase (ESBL)-producing Enterobacteriaceae in Budapest, Hungary. J. Antimicrob. Chemother. 2008, 62, 630–632. [Google Scholar] [CrossRef] [Green Version]
- Domokos, J.; Kristóf, K.; Szabó, D. Plasmid-mediated quinolone resistance among extended-spectrum beta-lactamase producing Enterobacteriaceae from bloodstream infections. Acta Microbiol. Immunol. Hung. 2016, 63, 313–323. [Google Scholar] [CrossRef] [Green Version]
- Hendriksen, R.S.; Munk, P.; Njage, P.; van Bunnik, B.; McNally, L.; Lukjancenko, O.; Röder, T.; Nieuwenhuijse, D.; Pedersen, S.K.; Kjeldgaard, J.; et al. Global monitoring of antimicrobial resistance based on metagenomics analyses of urban sewage. Nat. Commun. 2019, 10, 1124. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Martínez, L.; Pascual, A.; Jacoby, G.A. Quinolone resistance from a transferable plasmid. Lancet 1998, 351, 797–799. [Google Scholar] [CrossRef]
- Cattoir, V.; Poirel, L.; Aubert, C.; Soussy, C.J.; Nordmann, P. Unexpected occurrence of plasmid-mediated quinolone resistance determinants in environmental Aeromonas spp. Emerg. Infect. Dis. 2008, 14, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Ligetvári, F.; Zsabokorszky, F.; Kovács, K.; Zsirai, I. Wastewater Treatment and Sludge Utilisation in Hungary. J. Environ. Sci. Eng. B 2015, 4, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Serwecińska, L. Antimicrobials and Antibiotic-Resistant Bacteria: A Risk to the Environment and to Public Health. Water 2020, 12, 3313. [Google Scholar] [CrossRef]
- Libisch, B.; Uzinger, N.; Biró, B.; Anton, A. Isolation and characterisation of potentially pathogenic antibiotic-resistant bacterial strains from sewage sludge samples in Hungary. In Book of Abstracts, Proceedings of the 11th European Meeting on Environmental Chemistry, Portorož, Slovenia, 8–11 December 2010; Trebše, P., Petrič, M., Lavtižar, V., Eds.; University of Nova Gorica: Nova Gorica, Slovenia, 2010; p. 193. [Google Scholar]
- Sørum, H.; L’Abée-Lund, T.M.; Solberg, A.; Wold, A. Integron-containing IncU R plasmids pRAS1 and pAr-32 from the fish pathogen Aeromonas salmonicida. Antimicrob. Agents Chemother. 2003, 47, 1285–1290. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, A.S.; Bruun, M.S.; Larsen, J.L.; Dalsgaard, I. Characterization of class 1 integrons associated with R-plasmids in clinical Aeromonas salmonicida isolates from various geographical areas. J. Antimicrob. Chemother. 2001, 47, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, G.; Parkhill, J.; Bird, C.; Ambrose, K.; Jones, M.C.; Huys, G.; Swings, J.; Pickup, R.W. Complete nucleotide sequence of the conjugative tetracycline resistance plasmid pFBAOT6, a member of a group of IncU plasmids with global ubiquity. Appl. Environ. Microbiol. 2004, 70, 7497–7510. [Google Scholar] [CrossRef] [Green Version]
- Chang, Y.C.; Shih, D.Y.C.; Wang, J.Y.; Yang, S.S. Molecular characterization of class 1 integrons and antimicrobial resistance in Aeromonas strains from foodborne outbreak-suspect samples and environmental sources in Taiwan. Diagn. Microbiol. Infect. Dis. 2007, 59, 191–197. [Google Scholar] [CrossRef]
- Libisch, B.; Balogh, B.; Füzi, M. Identification of two multidrug-resistant Pseudomonas aeruginosa clonal lineages with a countrywide distribution in Hungary. Curr. Microbiol. 2009, 58, 111–116. [Google Scholar] [CrossRef]
- Libisch, B.; Giske, C.G.; Kovács, B.; Tóth, T.G.; Füzi, M. Identification of the first VIM metallo-β-lactamase-producing multiresistant Aeromonas hydrophila strain. J. Clin. Microbiol. 2008, 46, 1878–1880. [Google Scholar] [CrossRef] [Green Version]
- Libisch, B.; Watine, J.; Balogh, B.; Gacs, M.; Muzslay, M.; Szabó, G.; Füzi, M. Molecular typing indicates an important role for two international clonal complexes in dissemination of VIM-producing Pseudomonas aeruginosa clinical isolates in Hungary. Res. Microbiol. 2008, 159, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Campos Calero, G.; Caballero Gómez, N.; Benomar, N.; Pérez Montoro, B.; Knapp, C.W.; Gálvez, A.; Abriouel, H. Deciphering Resistome and Virulome Diversity in a Porcine Slaughterhouse and Pork Products Through Its Production Chain. Front. Microbiol. 2018, 9, 2099. [Google Scholar] [CrossRef] [Green Version]
- Jalili, V.; Afgan, E.; Gu, Q.; Clements, D.; Blankenberg, D.; Goecks, J.; Taylor, J.; Nekrutenko, A. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2020 update. Nucleic Acids Res. 2020, 48, W395–W402. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seemann, T.; Gladman, S. Fasta Statistics: Display Summary Statistics for a Fasta File. 2012. Available online: https://github.com/galaxyproject/tools-iuc (accessed on 30 September 2022).
- Seemann, T. ABRicate: Mass Screening of Contigs for Antiobiotic Resistance Genes. 2016. Available online: https://github.com/tseemann/abricate (accessed on 30 September 2022).
- Hernández-García, M.; García-Fernández, S.; García-Castillo, M.; Melo-Cristino, J.; Pinto, M.F.; Gonçalves, E.; Alves, V.; Costa, E.; Ramalheira, E.; Sancho, L.; et al. Confronting Ceftolozane-Tazobactam Susceptibility in Multidrug-Resistant Enterobacterales Isolates and Whole-Genome Sequencing Results (STEP Study). Int. J. Antimicrob. Agents 2021, 57, 106259. [Google Scholar] [CrossRef] [PubMed]
- Carroll, L.M.; Buehler, A.J.; Gaballa, A.; Siler, J.D.; Cummings, K.J.; Cheng, R.A.; Wiedmann, M. Monitoring the Microevolution of Salmonella enterica in Healthy Dairy Cattle Populations at the Individual Farm Level Using Whole-Genome Sequencing. Front. Microbiol. 2021, 12, 763669. [Google Scholar] [CrossRef] [PubMed]
- Bulens, S.N.; Reses, H.E.; Ansari, U.A.; Grass, J.E.; Carmon, C.; Albrecht, V.; Lawsin, A.; McAllister, G.; Daniels, J.; Lee, Y.K.; et al. Carbapenem-Resistant enterobacterales in individuals with and without health care risk factors—Emerging infections program, United States, 2012–2015. Am. J. Infect. Control, 2022; in press. [Google Scholar] [CrossRef]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Päivärinta, M.; Latvio, S.; Fredriksson-Ahomaa, M.; Heikinheimo, A. Whole genome sequence analysis of antimicrobial resistance genes, multilocus sequence types and plasmid sequences in ESBL/AmpC Escherichia coli isolated from broiler caecum and meat. Int. J. Food Microbiol. 2020, 315, 108361. [Google Scholar] [CrossRef]
- de Man, T.J.; Limbago, B.M. SSTAR, a Stand-Alone Easy-To-Use Antimicrobial Resistance Gene Predictor. mSphere 2016, 1, e00050-15. [Google Scholar] [CrossRef] [Green Version]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, D.E.; Salzberg, S.L. Kraken: Ultrafast metagenomic sequence classification using exact alignments. Genome Biol. 2014, 15, R46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.X.; Qin, Y.; Chen, T.; Lu, M.; Qian, X.; Guo, X.; Bai, Y. A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 2021, 12, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Steinegger, M.; Breitwieser, F.; Söding, J.; Levy Karin, E. Fast and sensitive taxonomic assignment to metagenomic contigs. Bioinformatics 2021, 37, 3029–3031. [Google Scholar] [CrossRef]
- Tamames, J.; Puente-Sánchez, F. SqueezeMeta, A Highly Portable, Fully Automatic Metagenomic Analysis Pipeline. Front. Microbiol 2019, 9, 3349. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aminoglycoside | Tetracycline | β-Lactam | Macrolide | Others | |
|---|---|---|---|---|---|
| Domestic pig | aac(6′)-Im aac(6′)-aph(2″) ant(3″)-Ia (aadA1) ant(6)-Ia (aadE) ant(6)-Ib ant(9)-Ia aph(2″)-Ib aph(2″)-Ic aph(2″)-If aph(2″)-Ih aph(3′)-Ia aph(3″)-Ib (strA) aph(6)-Id (strB) aph(3′)-III rmtF | tet(40) tet(44) tet(A) tet(B) tet(C) tet(H) tet(L) tet(M) tet(O) tet(Q) tet(W) tet(X) tetA(P) tetB(P) | blaACI-1 OXA-61 family β-lactamase blaROB-1 cfxA3 cfxA4 cfxA5 cfxA6 | lnu(B) lnu(C) lnu(P) lsa(E) mef(A) msr(D) erm(B) erm(F) erm(G) erm(Q) vatE | catP cfr(C) nimJ sul2 |
| Common carp | ant(3″)-Ia (aadA2) aph(3″)-Ib (strA) | tet(A) tet(B) tet(E) tet(X) | ampS cphA4 OXA-48-type carbapenemase | dfrA3 qnrS2 sul1 |
| Aminoglycosides | Tetracyclines | β-Lactams | Other Classes | In Total | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No. c | (%) | No. | (%) | No. | (%) | No. | (%) | No. | (%) | |
| Small intestine | 11 | 21.57 | 12 | 23.53 | 3 | 5.88 | 12 | 23.53 | 38 | 74.50 |
| Small intestine only a | 2 | 3.92 | 2 | 3.92 | 1 | 1.96 | 1 | 1.96 | 6 | 11.76 |
| Fecal samples | 13 | 25.49 | 11 | 21.57 | 6 | 11.76 | 14 | 27.45 | 44 | 86.27 |
| Fecal samples only b | 4 | 7.84 | 2 | 3.92 | 4 | 7.84 | 2 | 3.92 | 12 | 23.52 |
| In total | 15 | 29.41 | 14 | 27.45 | 7 | 13.73 | 15 | 29.41 | 51 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Libisch, B.; Abdulkadir, S.; Keresztény, T.; Papp, P.P.; Olasz, F.; Fébel, H.; Sándor, Z.J.; Rasschaert, G.; Lambrecht, E.; Heyndrickx, M.; et al. Detection of Acquired Antibiotic Resistance Genes in Domestic Pig (Sus scrofa) and Common Carp (Cyprinus carpio) Intestinal Samples by Metagenomics Analyses in Hungary. Antibiotics 2022, 11, 1441. https://doi.org/10.3390/antibiotics11101441
Libisch B, Abdulkadir S, Keresztény T, Papp PP, Olasz F, Fébel H, Sándor ZJ, Rasschaert G, Lambrecht E, Heyndrickx M, et al. Detection of Acquired Antibiotic Resistance Genes in Domestic Pig (Sus scrofa) and Common Carp (Cyprinus carpio) Intestinal Samples by Metagenomics Analyses in Hungary. Antibiotics. 2022; 11(10):1441. https://doi.org/10.3390/antibiotics11101441
Chicago/Turabian StyleLibisch, Balázs, Sahabi Abdulkadir, Tibor Keresztény, Péter P. Papp, Ferenc Olasz, Hedvig Fébel, Zsuzsanna J. Sándor, Geertrui Rasschaert, Ellen Lambrecht, Marc Heyndrickx, and et al. 2022. "Detection of Acquired Antibiotic Resistance Genes in Domestic Pig (Sus scrofa) and Common Carp (Cyprinus carpio) Intestinal Samples by Metagenomics Analyses in Hungary" Antibiotics 11, no. 10: 1441. https://doi.org/10.3390/antibiotics11101441