Development of a Microfluidic Chip Powered by EWOD for In Vitro Manipulation of Bovine Embryos

 , , , , , ,
, , , , , ,
Abstract
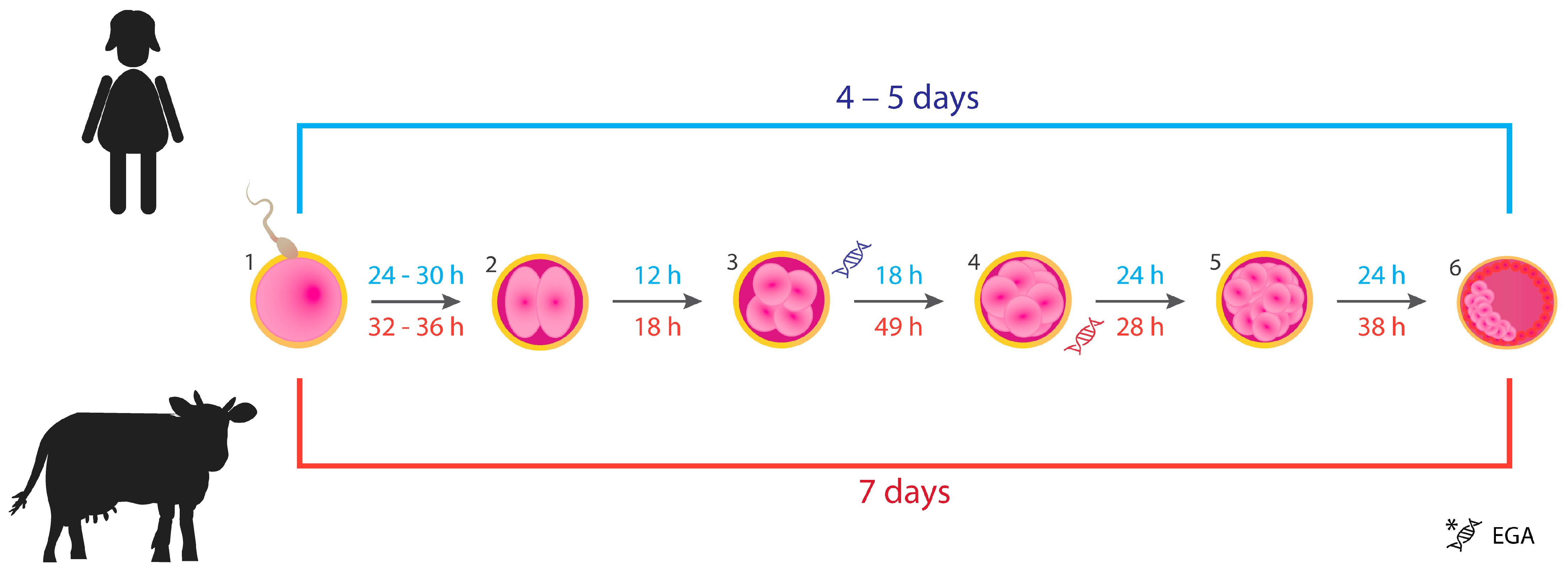
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
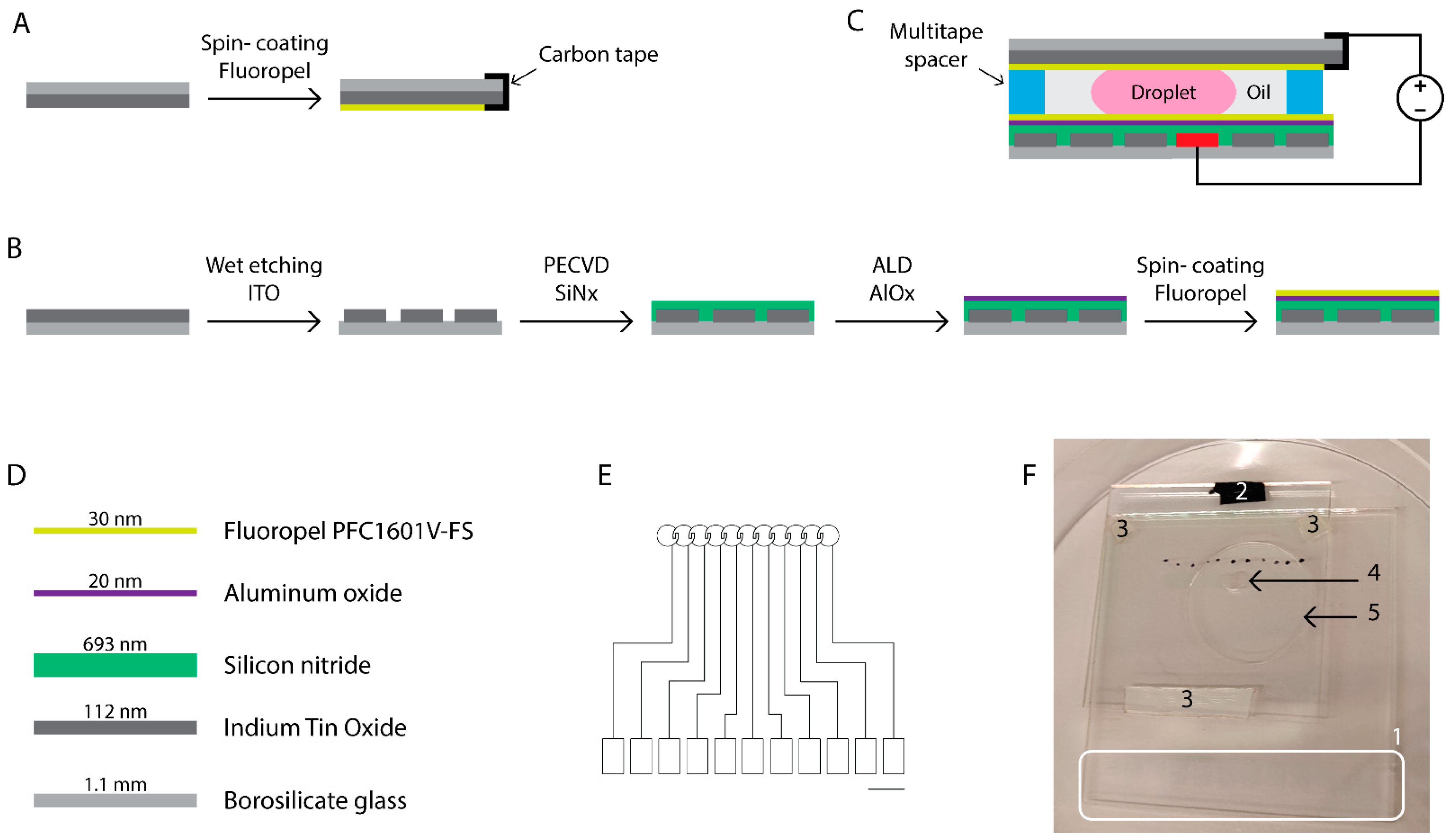
2.1.1. Microdevice Fabrication
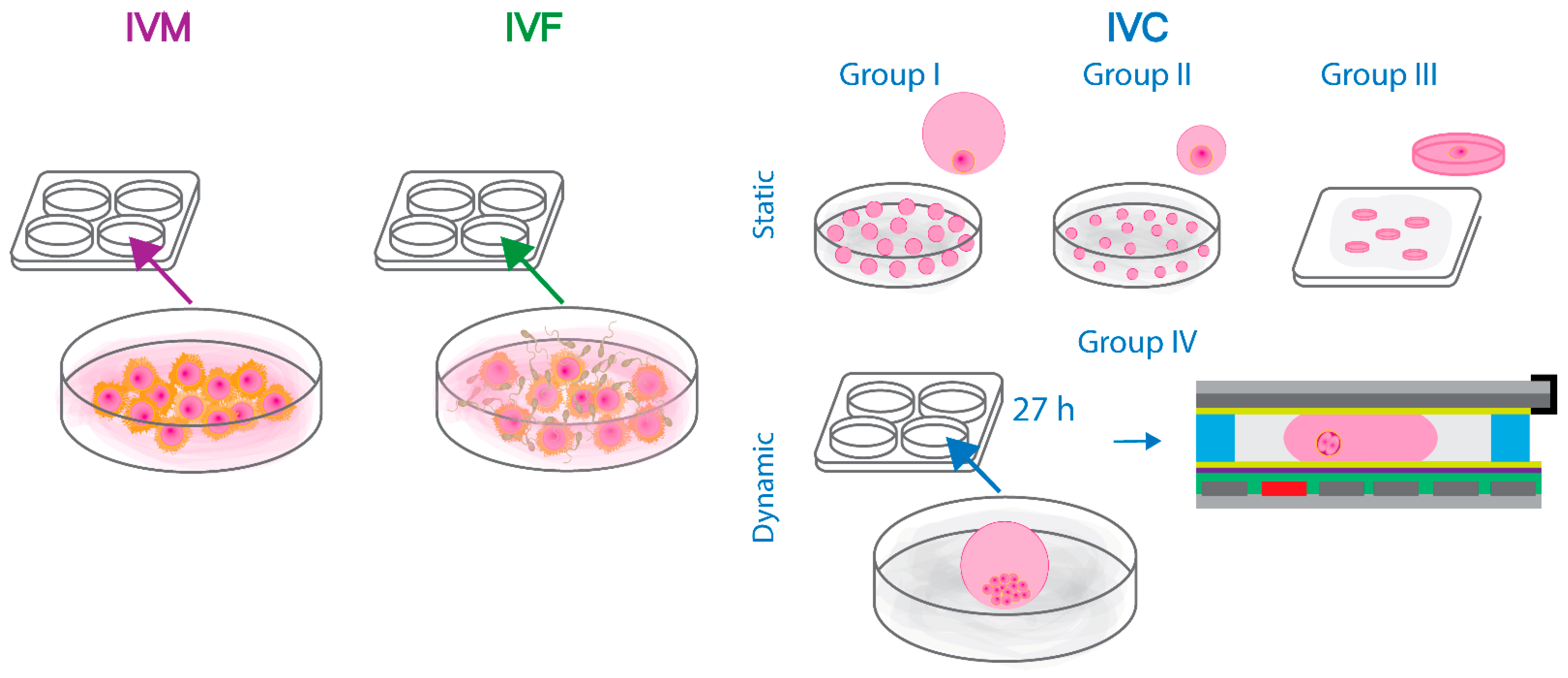
2.1.2. In Vitro Production of Bovine Embryos
2.2. Methods
2.2.1. Fabrication of EWOD Chips
2.2.2. In Vitro Production of Bovine Embryos
3. Results
3.1. Development of EWOD Microdevices with Multilayered Insulators
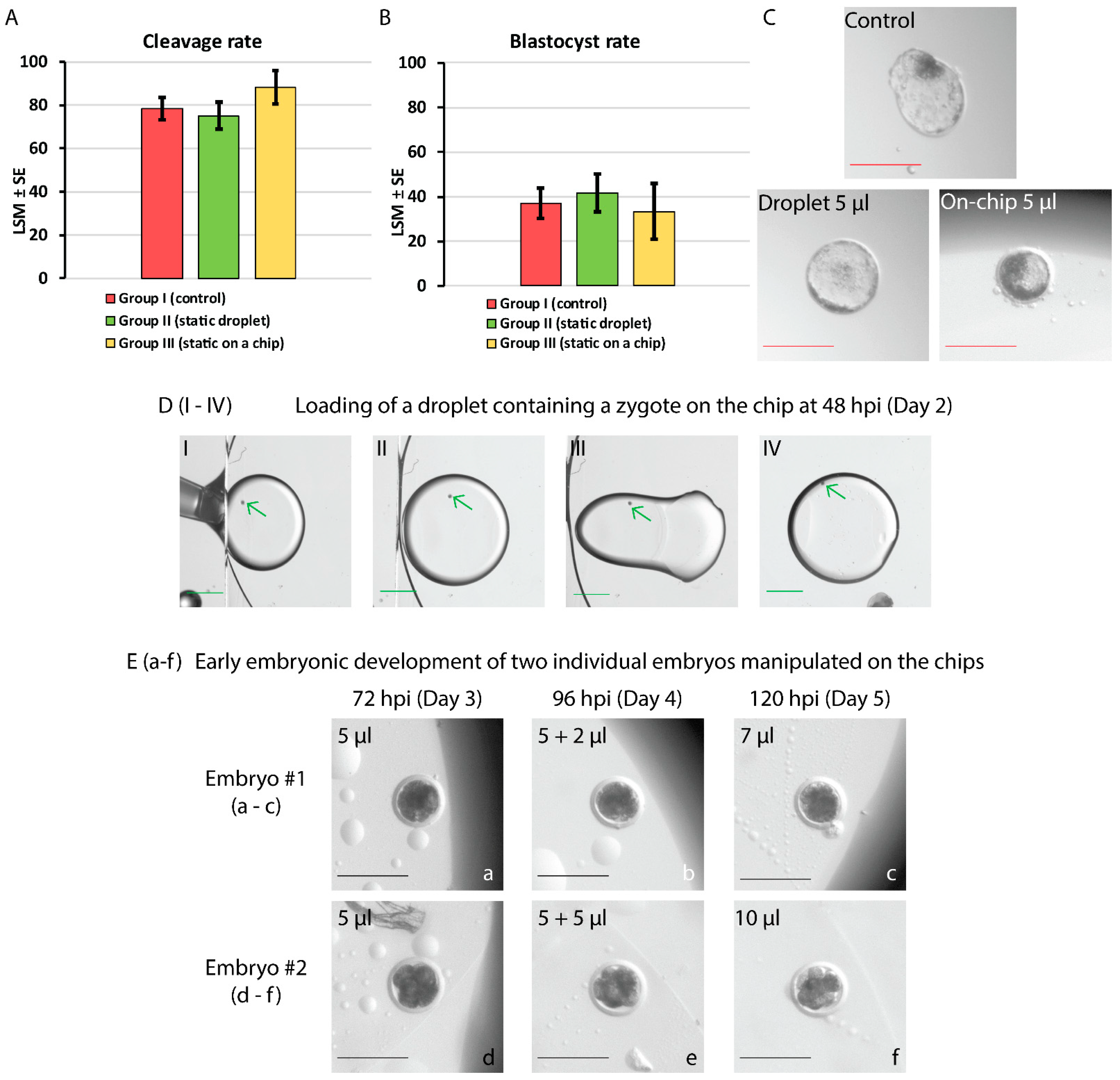
3.2. In Vitro Development of Bovine Embryos: Static Culture and Dynamic Manipulation on the Developed Chips
4. Discussion
4.1. Development of EWOD Chips Using Multilayered Dielectric Approach
4.2. Culture Medium Modification with Tween 80
4.3. In Vitro Culture and On-Chip Manipulation of Bovine Embryos
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The International Glossary on Infertility and Fertility Care, 2017. Fertil. Steril. 2017, 108, 393–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inhorn, M.C.; Patrizio, P. Infertility around the Globe: New Thinking on Gender, Reproductive Technologies and Global Movements in the 21st Century. Hum. Reprod. Update 2014, 21, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Krausz, C.; Riera-Escamilla, A. Genetics of Male Infertility. Nat. Rev. Urol. 2018, 15, 369–384. [Google Scholar] [CrossRef]
- Yatsenko, S.A.; Rajkovic, A. Genetics of Human Female Infertility. Biol. Reprod. 2019, 101, 549–566. [Google Scholar] [CrossRef]
- Thurston, L.; Abbara, A.; Dhillo, W.S. Investigation and Management of Subfertility. J. Clin. Pathol. 2019, 72, 579–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vander Borght, M.; Wyns, C. Fertility and Infertility: Definition and Epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Gelbaya, T.; Potdar, N.; Jeve, Y.N.L. CME Review Article. Pediatr. Emerg. Care 2017, 33, 792–793. [Google Scholar] [CrossRef]
- Choy, J.T.; Eisenberg, M.L. Male Infertility as a Window to Health. Fertil. Steril. 2018, 110, 810–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanson, B.; Johnstone, E.; Dorais, J.; Silver, B.; Peterson, C.M.; Hotaling, J. Female Infertility, Infertility-Associated Diagnoses, and Comorbidities: A Review. J. Assist. Reprod. Genet. 2017, 34, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Rooney, K.L.; Domar, A.D. The Relationship between Stress and Infertility. Dialogues Clin. Neurosci. 2018, 20, 41–47. [Google Scholar] [CrossRef]
- Fact Sheet on ART. Available online: https://www.eshre.eu/Europe/Factsheets-and-infographics (accessed on 12 January 2022).
- Biggers, J.D. IVF and Embryo Transfer: Historical Origin and Development. Reprod. Biomed. Online 2012, 25, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speyer, B.; O’Neill, H.; Saab, W.; Seshadri, S.; Cawood, S.; Heath, C.; Gaunt, M.; Serhal, P. In Assisted Reproduction by IVF or ICSI, the Rate at Which Embryos Develop to the Blastocyst Stage Is Influenced by the Fertilization Method Used: A Split IVF/ICSI Study. J. Assist. Reprod. Genet. 2019, 36, 647–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulte, K.; Ehmcke, J.; Schlatt, S.; Boiani, M.; Nordhoff, V. Lower Total Cell Numbers in Mouse Preimplantation Embryos Cultured in Human Assisted Reproductive Technique (ART) Media Are Not Induced by Apoptosis. Theriogenology 2015, 84, 1620–1630. [Google Scholar] [CrossRef] [PubMed]
- Isobe, T.; Ikebata, Y.; Do, L.T.K.; Tanihara, F.; Taniguchi, M.; Otoi, T. In Vitro Development of OPU-derived Bovine Embryos Cultured Either Individually or in groups with the silk protein sericin and the viability of frozen-thawed embryos after transfer. Anim. Sci. J. 2015, 86, 661–665. [Google Scholar] [CrossRef]
- Lacham-Kaplan, O.; Trounson, A. Reduced Developmental Competence of Immature, In-Vitro Matured and Postovulatory Aged Mouse Oocytes Following IVF and ICSI. Reprod. Biol. Endocrinol. 2008, 6, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ferré, L.B.; Kjelland, M.E.; Strøbech, L.B.; Hyttel, P.; Mermillod, P.; Ross, P.J. Review: Recent Advances in Bovine In Vitro Embryo Production: Reproductive Biotechnology History and Methods. Animal 2020, 14, 991–1004. [Google Scholar] [CrossRef] [Green Version]
- Coy, P.; García-Vázquez, F.A.; Visconti, P.E.; Avilés, M. Roles of the Oviduct in Mammalian Fertilization. Reproduction 2012, 144, 649–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, S.; Wang, Z.; Peng, H.; Ward, S.M.; Hennig, G.W.; Zheng, H.; Yan, W. Oviductal Motile Cilia Are Essential for Oocyte Pickup but Dispensable for Sperm and Embryo Transport. Proc. Natl. Acad. Sci. USA 2021, 118, e2102940118. [Google Scholar] [CrossRef]
- Mirihagalle, S.; Hughes, J.R.; Miller, D.J. Progesterone-Induced Sperm Release from the Oviduct Sperm Reservoir. Cells 2022, 11, 1622. [Google Scholar] [CrossRef]
- Kölle, S.; Hughes, B.; Steele, H. Early Embryo-Maternal Communication in the Oviduct: A Review. Mol. Reprod. Dev. 2020, 87, 650–662. [Google Scholar] [CrossRef]
- Rossant, J.; Tam, P.P.L. Exploring Early Human Embryo Development. Science 2018, 360, 1075–1076. [Google Scholar] [CrossRef]
- Kocyigit, A.; Cevik, M. Correlation between the Cryosurvival, Cell Number and Diameter in Bovine in Vitro Produced Embryos. Cryobiology 2016, 73, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.S.; Harris, D.C.; Daneshmand, S.T.; Shapiro, B.S. Quantitative Grading of a Human Blastocyst: Optimal Inner Cell Mass Size and Shape. Fertil. Steril. 2001, 76, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- An, L.; Liu, Y.; Li, M.; Liu, Z.; Wang, Z.; Dai, Y.; Presicce, G.A.; Du, F. Site Specificity of Blastocyst Hatching Significantly Influences Pregnancy Outcomes in Mice. FASEB J. 2021, 35, e21812. [Google Scholar] [CrossRef]
- Svoboda, P. Seminars in Cell & Developmental Biology Mammalian Zygotic Genome Activation. Semin. Cell Dev. Biol. 2018, 84, 118–126. [Google Scholar]
- Niakan, K.K.; Han, J.; Pedersen, R.A.; Simon, C.; Pera, R.A.R. Human Pre-Implantation Embryo Development. Development 2012, 139, 829–841. [Google Scholar] [CrossRef] [Green Version]
- Santos, R.R.; Schoevers, E.J.; Roelen, B.A.J. Usefulness of Bovine and Porcine IVM/IVF Models for Reproductive Toxicology. Reprod. Biol. Endocrinol. 2014, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asami, M.; Lam, B.Y.H.; Ma, M.K.; Rainbow, K.; Braun, S.; VerMilyea, M.D.; Yeo, G.S.H.; Perry, A.C.F. Human Embryonic Genome Activation Initiates at the One-Cell Stage. Cell Stem Cell 2022, 29, 209–216.e4. [Google Scholar] [CrossRef]
- Lonergan, P.; Fair, T. The ART of Studying Early Embryo Development: Progress and Challenges in Ruminant Embryo Culture. Theriogenology 2014, 81, 49–55. [Google Scholar] [CrossRef]
- Zhai, J.; Xiao, Z.; Wang, Y.; Wang, H. Human Embryonic Development: From Peri-Implantation to Gastrulation. Trends Cell Biol. 2022, 32, 18–29. [Google Scholar] [CrossRef]
- el Azhar, Y.; Sonnen, K.F. Development in a Dish—In Vitro Models of Mammalian Embryonic Development. Front. Cell Dev. Biol. 2021, 9, 655993. [Google Scholar] [CrossRef]
- Holm, P.; Booth, P.J.; Callesen, H. Kinetics of Early In Vitro Development of Bovine In Vivo- and In Vitro-Derived Zygotes Produced and/or Cultured in Chemically Defined or Serum-Containing Media. Reproduction 2002, 123, 553–565. [Google Scholar] [CrossRef]
- Sirard, M.A. 40 Years of Bovine IVF in the New Genomic Selection Context. Reproduction 2018, 156, R1–R7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ealy, A.D.; Wooldridge, L.K.; McCoski, S.R. Board Invited Review: Post-Transfer Consequences of In Vitro-Produced Embryos in Cattle. J. Anim. Sci. 2019, 97, 2555–2568. [Google Scholar] [CrossRef]
- Ramos-Ibeas, P.; Gimeno, I.; Cañón-Beltrán, K.; Gutiérrez-Adán, A.; Rizos, D.; Gómez, E. Senescence and Apoptosis during In Vitro Embryo Development in a Bovine Model. Front. Cell Dev. Biol. 2020, 8, 619902. [Google Scholar] [CrossRef] [PubMed]
- Wydooghe, E.; Vandaele, L.; Heras, S.; De Sutter, P.; Deforce, D.; Peelman, L.; De Schauwer, C.; Van Soom, A. Autocrine Embryotropins Revisited: How Do Embryos Communicate with Each Other In Vitro When Cultured in Groups? Biol. Rev. 2017, 92, 505–520. [Google Scholar] [CrossRef]
- Asaadi, A.; Dolatabad, N.A.; Atashi, H.; Raes, A.; Van Damme, P.; Hoelker, M.; Hendrix, A.; Pascottini, O.B.; Van Soom, A.; Kafi, M.; et al. Extracellular Vesicles from Follicular and Ampullary Fluid Isolated by Density Gradient Ultracentrifugation Improve Bovine Embryo Development and Quality. Int. J. Mol. Sci. 2021, 22, 578. [Google Scholar] [CrossRef] [PubMed]
- Stone, H.A.; Stroock, A.D.; Ajdari, A. Engineering Flows in Small Devices: Microfluidics toward a Lab-on-a-Chip. Annu. Rev. Fluid Mech. 2004, 36, 381–411. [Google Scholar] [CrossRef] [Green Version]
- Ahmadkhani, N.; Hosseini, M.; Saadatmand, M.; Abbaspourrad, A. The Influence of the Female Reproductive Tract and Sperm Features on the Design of Microfluidic Sperm-Sorting Devices. J. Assist. Reprod. Genet. 2022, 39, 19–36. [Google Scholar] [CrossRef]
- Le Gac, S.; Nordhoff, V. Microfluidics for Mammalian Embryo Culture and Selection: Where Do We Stand Now? Mol. Hum. Reprod. 2017, 23, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Shuchat, S.; Yossifon, G.; Huleihel, M. Perfusion in Organ-on-Chip Models and Its Applicability to the Replication of Spermatogenesis In Vitro. Int. J. Mol. Sci. 2022, 23, 5402. [Google Scholar] [CrossRef]
- Alias, A.; Huang, H.; Yao, D.-J. A Review on Microfluidics: An Aid to Assisted. Molecules 2021, 26, 4354. [Google Scholar] [CrossRef]
- Karcz, A.; Van Soom, A.; Smits, K.; Verplancke, R.; Van Vlierberghe, S.; Vanfleteren, J. Electrically-Driven Handling of Gametes and Embryos: Taking a Step towards the Future of ARTs. Lab A Chip 2022, 22, 1852–1875. [Google Scholar] [CrossRef] [PubMed]
- Kothamachu, V.B.; Zaini, S.; Muffatto, F. Role of Digital Microfluidics in Enabling Access to Laboratory Automation and Making Biology Programmable. SLAS Technol. 2020, 25, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Alistar, M. Mobile Microfluidics. Bioengineering 2019, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- Suman Chakraborty. Electrocapillary. In Encyclopedia of Microfluidics and Nanofluidics; Li, D., Ed.; Springer: Boston, MA, USA, 2008; pp. 460–469. [Google Scholar]
- Lin, Y.Y.; Welch, E.R.F.; Fair, R.B. Low Voltage Picoliter Droplet Manipulation Utilizing Electrowetting-on- Dielectric Platforms. Sensors Actuators B Chem. 2012, 173, 338–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikapitiya, N.Y.J.B.; Nahar, M.M.; Moon, H. Accurate, Consistent, and Fast Droplet Splitting and Dispensing in Electrowetting on Dielectric Digital Microfluidics. Micro Nano Syst. Lett. 2017, 5, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nahar, M.M.; Nikapitiya, J.B.; You, S.M.; Moon, H. Droplet Velocity in an Electrowetting on Dielectric Digital Microfluidic Device. Micromachines 2016, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.P.; Wang, Y. Fundamentals and Applications of Electrowetting: A Critical Review. Rev. Adhes. Adhes. 2013, 1, 114–174. [Google Scholar] [CrossRef]
- Choi, K.; Ng, A.H.C.; Fobel, R.; Wheeler, A.R. Digital Microfluidics. Annu. Rev. Anal. Chem. 2012, 5, 413–440. [Google Scholar] [CrossRef] [Green Version]
- Samiei, E.; Tabrizian, M.; Hoorfar, M. A Review of Digital Microfluidics as Portable Platforms for Lab-on a-Chip Applications. Lab Chip 2016, 16, 2376–2396. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Nguyen, N.T. Magnetic Digital Microfluidics-a Review. Lab Chip 2017, 17, 994–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.; Liu, H.; Li, W.; Li, M. Recent Advances in Magnetic Digital Microfluidic Platforms. Electrophoresis 2021, 42, 2329–2346. [Google Scholar] [CrossRef] [PubMed]
- Rui, X.; Song, S.; Wang, W.; Zhou, J. Applications of Electrowetting-on-Dielectric (EWOD) Technology for Droplet Digital PCR. Biomicrofluidics 2020, 14, 061503. [Google Scholar] [CrossRef]
- Barman, S.R.; Khan, I.; Chatterjee, S.; Saha, S.; Choi, D.; Lee, S.; Lin, Z.H. Electrowetting-on-Dielectric (EWOD): Current Perspectives and Applications in Ensuring Food Safety. J. Food Drug Anal. 2020, 28, 595–621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y. Advances in Integrated Digital Microfluidic Platforms for Point-of-Care Diagnosis: A Review. Sens. Diagn. 2022, 1, 648–672. [Google Scholar] [CrossRef]
- Bender, B.F.; Garrell, R.L. Digital Microfluidic System with Vertical Functionality. Micromachines 2015, 6, 1655–1674. [Google Scholar] [CrossRef] [Green Version]
- George, S.M.; Moon, H. Alginate Hydrogel Based 3-Dimensional Cell Culture and Chemical Screening Platform Using Digital Microfluidics. In Proceedings of the 2015 28th IEEE International Conference on Micro Electro Mechanical Systems (MEMS), Estoril, Portugal, 18–22 January 2015; pp. 443–446. [Google Scholar] [CrossRef]
- Alias, A.B.; Chiang, C.E.; Huang, H.Y.; Lin, K.T.; Lu, P.J.; Wang, Y.W.; Wu, T.H.; Jiang, P.S.; Chen, C.A.; Yao, D.J. Extraction of Cell-Free Dna from An Embryo-Culture Medium Using Micro-Scale Bio-Reagents on Ewod. Sci. Rep. 2020, 10, 9708. [Google Scholar] [CrossRef]
- Chiang, C.E.; Huang, H.Y.; Lin, K.T.; Alias, A.B.; Lu, P.J.; Wang, Y.W.; Wu, T.H.; Jiang, P.S.; Chen, C.A.; Yao, D.J. A Medical Innovation: A New and Improved Method of DNA Extraction with Electrowetting-on-Dielectric of Genetic Testing in-Vitro Fertilization (IVF). Microfluid. Nanofluidics 2020, 24, 1–9. [Google Scholar] [CrossRef]
- Lee, M.S.; Hsu, W.; Huang, H.Y.; Tseng, H.Y.; Lee, C.T.; Hsu, C.Y.; Shieh, Y.C.; Wang, S.H.; Yao, D.J.; Liu, C.H. Simultaneous Detection of Two Growth Factors from Human Single-Embryo Culture Medium by a Bead-Based Digital Microfluidic Chip. Biosens. Bioelectron. 2020, 150, 111851. [Google Scholar] [CrossRef]
- Huang, H.Y.; Shen, H.H.; Tien, C.H.; Li, C.J.; Fan, S.K.; Liu, C.H.; Hsu, W.S.; Yao, D.J. Digital Microfluidic Dynamic Culture of Mammalian Embryos on an Electrowetting on Dielectric (EWOD) Chip. PLoS ONE 2015, 10, e0124196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karcz, A.A.; Schaubroeck, D.; Verplancke, R.; Van Soom, A.; Vanfleteren, J. Introduction of a High Quality Nanofilm of Aluminum Oxide Enhances the Performance of Ewod Microfluidic Platforms. In Proceedings of the 25th International Conference on Miniaturized Systems for Chemistry and Life Sciences, Palm Springs, CA, USA, 10–14 October 2021; pp. 1395–1396. [Google Scholar]
- Wydooghe, E.; Vaele, L.; Piepers, S.; Dewulf, J.; Van Abbeel, E.D.; De Sutter, P.; Van Soom, A. Individual Commitment to a Group Effect: Strengths and Weaknesses of Bovine Embryo Group Culture. Reproduction 2014, 148, 519–529. [Google Scholar] [CrossRef] [Green Version]
- Yota, J.; Shen, H.; Ramanathan, R. Characterization of Atomic Layer Deposition HfO2, Al2O3, and Plasma-Enhanced Chemical Vapor Deposition Si3N4 as Metal–Insulator–Metal Capacitor Dielectric for GaAs HBT Technology. J. Vac. Sci. Technol. A Vacuum Surfaces Film. 2013, 31, 01A134. [Google Scholar] [CrossRef] [Green Version]
- Robertson, J. High Dielectric Constant Oxides. Eur. Phys. J. Appl. Phys. 2004, 28, 265–291. [Google Scholar] [CrossRef] [Green Version]
- Available online: https://Cytonix.Com/Products/1601v (accessed on 21 October 2022).
- Schultz, A.; Chevalliot, S.; Kuiper, S.; Heikenfeld, J. Detailed Analysis of Defect Reduction in Electrowetting Dielectrics through a Two-Layer “barrier” Approach. Thin Solid Films 2013, 534, 348–355. [Google Scholar] [CrossRef]
- Gisby, T.A.; Xie, S.Q.; Calius, E.P.; Anderson, I.A. Leakage Current as a Predictor of Failure in Dielectric Elastomer Actuators. Electroact. Polym. Actuators Devices 2010, 7642, 319–329. [Google Scholar] [CrossRef]
- Popova, E.; Bader, M.; Krivokharchenko, A. Effects of Electric Field on Early Preimplantation Development In Vitro in Mice and Rats. Hum. Reprod. 2011, 26, 662–670. [Google Scholar] [CrossRef] [Green Version]
- Cross, M.H.; Cross, P.C.; Brinster, R.L. Changes in Membrane Potential during Mouse Egg Development. Dev. Biol. 1973, 416, 412–416. [Google Scholar] [CrossRef]
- Funk, R.H.W. Endogenous Electric Fields as Guiding Cue for Cell Migration. Front. Physiol. 2015, 6, 143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, G.; Tai, M.; Zhao, M. Electrically Stimulated Cell Migration and Its Contribution to Wound Healing. Burn. Trauma 2018, 6, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Nuccitelli, R. Endogenous Electric Fields in Embryos during Development, Regeneration and Wound Healing. Radiat. Prot. Dosimetry 2003, 106, 375–383. [Google Scholar] [CrossRef]
- Hotary, K.B.; Robinson, K.R. Endogenous Electrical Currents and Voltage Gradients in Xenopus Embryos and Consequences of Their Disruption. Dev. Biol. 1994, 166, 789–800. [Google Scholar] [CrossRef]
- Koo, J.; Lee, J.; Kim, S.; Kim, Y.; Jeon, H.; Kim, D.; Kim, Y. Characteristics of Hafnium-Aluminum-Oxide Thin Films Deposited by Using Atomic Layer Deposition with Various Aluminum Compositions. J. Korean Phys. Soc. 2005, 47, 501–507. [Google Scholar]
- Ylivaara, O.M.E.; Liu, X.; Kilpi, L.; Lyytinen, J.; Schneider, D.; Laitinen, M.; Julin, J.; Ali, S.; Sintonen, S.; Berdova, M.; et al. Aluminum Oxide from Trimethylaluminum and Water by Atomic Layer Deposition: The Temperature Dependence of Residual Stress, Elastic Modulus, Hardness and Adhesion. Thin Solid Films 2014, 552, 124–135. [Google Scholar] [CrossRef]
- Wydooghe, E.; Heras, S.; Dewulf, J.; Piepers, S.; Van Den Abbeel, E.; De Sutter, P.; Vandaele, L.; Van Soom, A. Replacing Serum in Culture Medium with Albumin and Insulin, Transferrin and Selenium Is the Key to Successful Bovine Embryo Development in Individual Culture. Reprod. Fertil. Dev. 2014, 26, 717–724. [Google Scholar] [CrossRef]
- Francis, G.L. Albumin and Mammalian Cell Culture: Implications for Biotechnology Applications. Cytotechnology 2010, 62, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Jeyachandran, Y.L.; Mielczarski, E.; Rai, B.; Mielczarski, J.A. Quantitative and Qualitative Evaluation of Adsorption/Desorption of Bovine Serum Albumin on Hydrophilic and Hydrophobic Surfaces. Langmuir 2009, 25, 11614–11620. [Google Scholar] [CrossRef] [PubMed]
- Luk, V.N.; Mo, G.C.; Wheeler, A.R. Pluronic Additives: A Solution to Sticky Problems in Digital Microfluidics. Langmuir 2008, 24, 6382–6389. [Google Scholar] [CrossRef]
- Zadymova, N.M.; Yampol’skaya, G.P.; Filatova, L.Y. Interaction of Bovine Serum Albumin with Nonionic Surfactant Tween 80 in Aqueous Solutions: Complexation and Association. Colloid J. 2006, 68, 162–172. [Google Scholar] [CrossRef]
- Ruiz-Peña, M.; Oropesa-Nuñez, R.; Pons, T.; Louro, S.R.W.; Pérez-Gramatges, A. Physico-Chemical Studies of Molecular Interactions between Non-Ionic Surfactants and Bovine Serum Albumin. Colloids Surfaces B Biointerfaces 2010, 75, 282–289. [Google Scholar] [CrossRef]
- Delgado-Magnero, K.H.; Valiente, P.A.; Ruiz-Peña, M.; Pérez-Gramatges, A.; Pons, T. Unraveling the Binding Mechanism of Polyoxyethylene Sorbitan Esters with Bovine Serum Albumin: A Novel Theoretical Model Based on Molecular Dynamic Simulations. Colloids Surfaces B Biointerfaces 2014, 116, 720–726. [Google Scholar] [CrossRef]
- Baltz, J.M.; Tartia, A.P. Cell Volume Regulation in Oocytes and Early Embryos: Connecting Physiology to Successful Culture Media. Hum. Reprod. Update 2009, 16, 166–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, D.K.; Hamilton, R.; McCallie, B.; Schoolcraft, W.B.; Katz-Jaffe, M.G. Human and Mouse Embryonic Development, Metabolism and Gene Expression Are Altered by an Ammonium Gradient In Vitro. Reproduction 2013, 146, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammon, D.S.; Wang, S.; Holyoak, G.R. Effects of Ammonia during Different Stages of Culture on Development of in Vitro Produced Bovine Embryos. Anim. Reprod. Sci. 2000, 59, 23–30. [Google Scholar] [CrossRef]
- Gardner, D.K.; Kelley, R.L. Impact of the IVF Laboratory Environment on Human Preimplantation Embryo Phenotype. J. Dev. Orig. Health Dis. 2017, 8, 418–435. [Google Scholar] [CrossRef] [PubMed]
- Lopes, A.S.; Lane, M.; Thompson, J.G. Oxygen Consumption and ROS Production Are Increased at the Time of Fertilization and Cell Cleavage in Bovine Zygotes. Hum. Reprod. 2010, 25, 2762–2773. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Wang, L. Oxidative Stress in Oocytes and Embryo Development: Implications for In Vitro Systems. Antioxidants Redox Signal. 2021, 34, 1394–1406. [Google Scholar] [CrossRef]
- Nagao, Y.; Iijima, R.; Saeki, K. Interaction between Embryos and Culture Conditions during In Vitro Development of Bovine Early Embryos. Zygote 2008, 16, 127–133. [Google Scholar] [CrossRef]
- Ottosen, L.D.M.; Hindkjær, J.; Ingerslev, J. Light Exposure of the Ovum and Preimplantation Embryo during ART Procedures. J. Assist. Reprod. Genet. 2007, 24, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Pomeroy, K.O.; Reed, M.L. The Effect of Light on Embryos and Embryo Culture. J. Reprod. Stem Cell Biotechnol. 2012, 3, 46–54. [Google Scholar] [CrossRef] [Green Version]
- Swain, J.E.; Carrell, D.; Cobo, A.; Meseguer, M.; Rubio, C.; Smith, G.D. Optimizing the Culture Environment and Embryo Manipulation to Help Maintain Embryo Developmental Potential. Fertil. Steril. 2016, 105, 571–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kermack, A.J.; Fesenko, I.; Christensen, D.R.; Parry, K.L.; Lowen, P.; Wellstead, S.J.; Harris, S.F.; Calder, P.C.; Macklon, N.S.; Houghton, F.D. Incubator Type Affects Human Blastocyst Formation and Embryo Metabolism: A Randomized Controlled Trial. Hum. Reprod. 2022, 37, 2757–2767. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Chen, P.; Zhou, Q. Adhesion Promotion between PDMS and Glass by Oxygen Plasma Pre-Treatment. J. Adhes. Sci. Technol. 2014, 28, 1046–1054. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coating | Material | Thickness | Dielectric Constant εr * | |
|---|---|---|---|---|
| Open Platform (Figure 2) | Chip for Embryo Manipulation (Figure 3) | |||
| Dielectric 1 | Silicon nitride | 590 nm | 693 nm | 6–8 [67] |
| Dielectric 2 | Aluminum oxide | 13 nm | 20 nm | 9 [68] |
| Hydrophobic ** | Fluoropel PFC1601V-FS | 30 nm | 30 nm | 1.9 [69] |
| Morphological Assessment | 48 hpi | 72 hpi | 96 hpi | 120 hpi | 144 hpi | |
|---|---|---|---|---|---|---|
| Corresponding Cell Cycles (Timing in Hpi) | 1st (32–36) 1-Cell to 2-Cell | 2nd (36–54) 2-Cell to 4-Cell | 3rd (54–103) 4-Cell to 8-Cell | 4th (103–131) 8-Cell to Morula | Blastocyst Formation (131–192) | |
| Developmental stage | 2-cell | 7 (47%) | ||||
| 4-cell | 8 (53%) | 12 (80%) | 10 (67%) | |||
| 8-cell | 2 (13%) | 4 (27%) | 2 (13%) | |||
| Morula | 2 (13%) | 2 (13%) | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karcz, A.; Van Soom, A.; Smits, K.; Van Vlierberghe, S.; Verplancke, R.; Pascottini, O.B.; Van den Abbeel, E.; Vanfleteren, J. Development of a Microfluidic Chip Powered by EWOD for In Vitro Manipulation of Bovine Embryos. Biosensors 2023, 13, 419. https://doi.org/10.3390/bios13040419
Karcz A, Van Soom A, Smits K, Van Vlierberghe S, Verplancke R, Pascottini OB, Van den Abbeel E, Vanfleteren J. Development of a Microfluidic Chip Powered by EWOD for In Vitro Manipulation of Bovine Embryos. Biosensors. 2023; 13(4):419. https://doi.org/10.3390/bios13040419
Chicago/Turabian StyleKarcz, Adriana, Ann Van Soom, Katrien Smits, Sandra Van Vlierberghe, Rik Verplancke, Osvaldo Bogado Pascottini, Etienne Van den Abbeel, and Jan Vanfleteren. 2023. "Development of a Microfluidic Chip Powered by EWOD for In Vitro Manipulation of Bovine Embryos" Biosensors 13, no. 4: 419. https://doi.org/10.3390/bios13040419