Nanobody-Based Sandwich Immunoassay for Pathogenic Escherichia coli F17 Strain Detection

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Immunization, Construction of VHH Library and Biopanning
2.2. Subcloning, Nanobody Expression and Purification
2.3. VHH Binding by Cell-ELISA
2.4. Preparation of Nb1 Immunomagnetic Beads
2.5. Conjugation of Nb4 to the Horseradish Peroxidase (HRP)
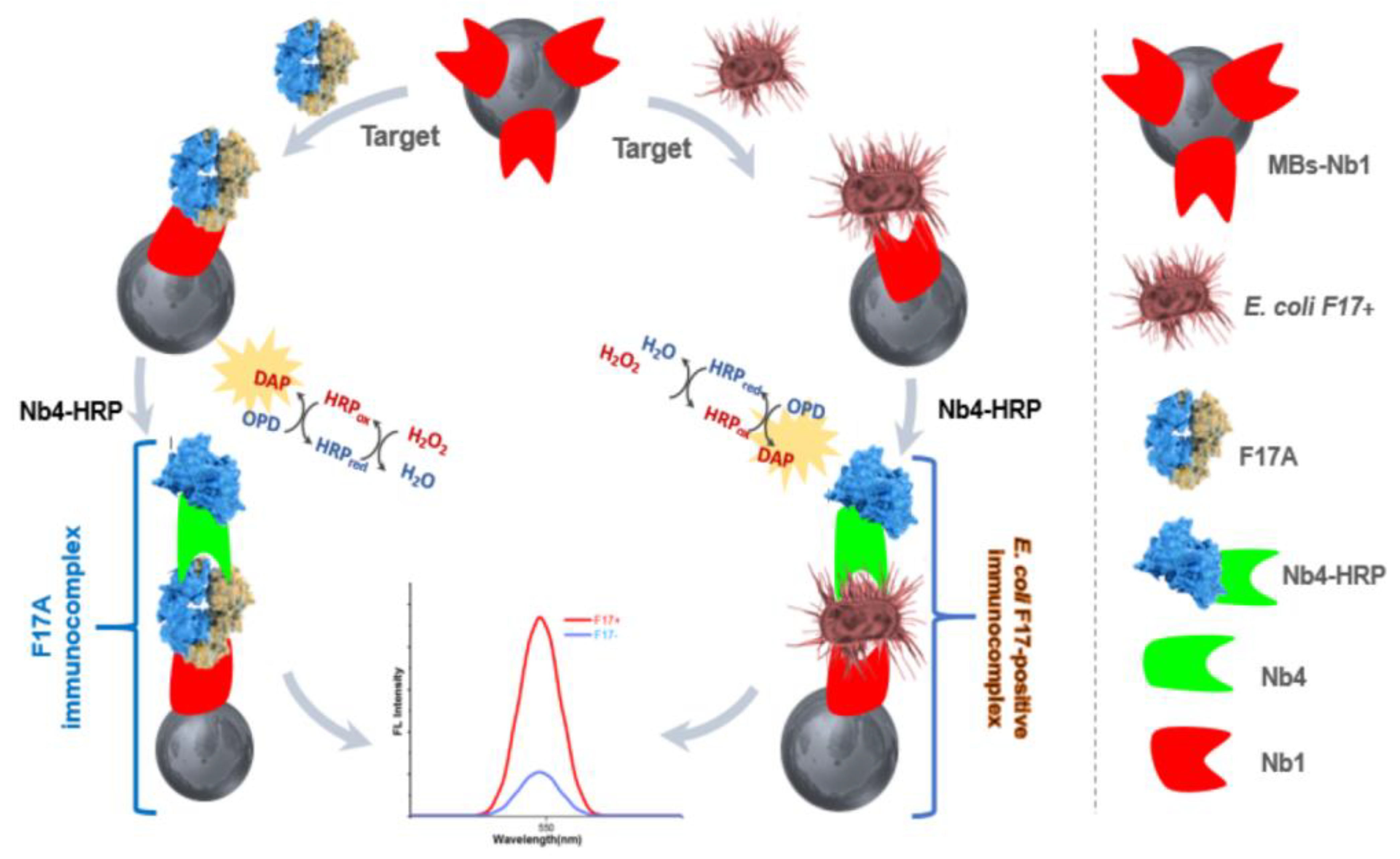
2.6. The Sandwich Immunoassays
2.7. Spike and Recovery Assay
3. Results
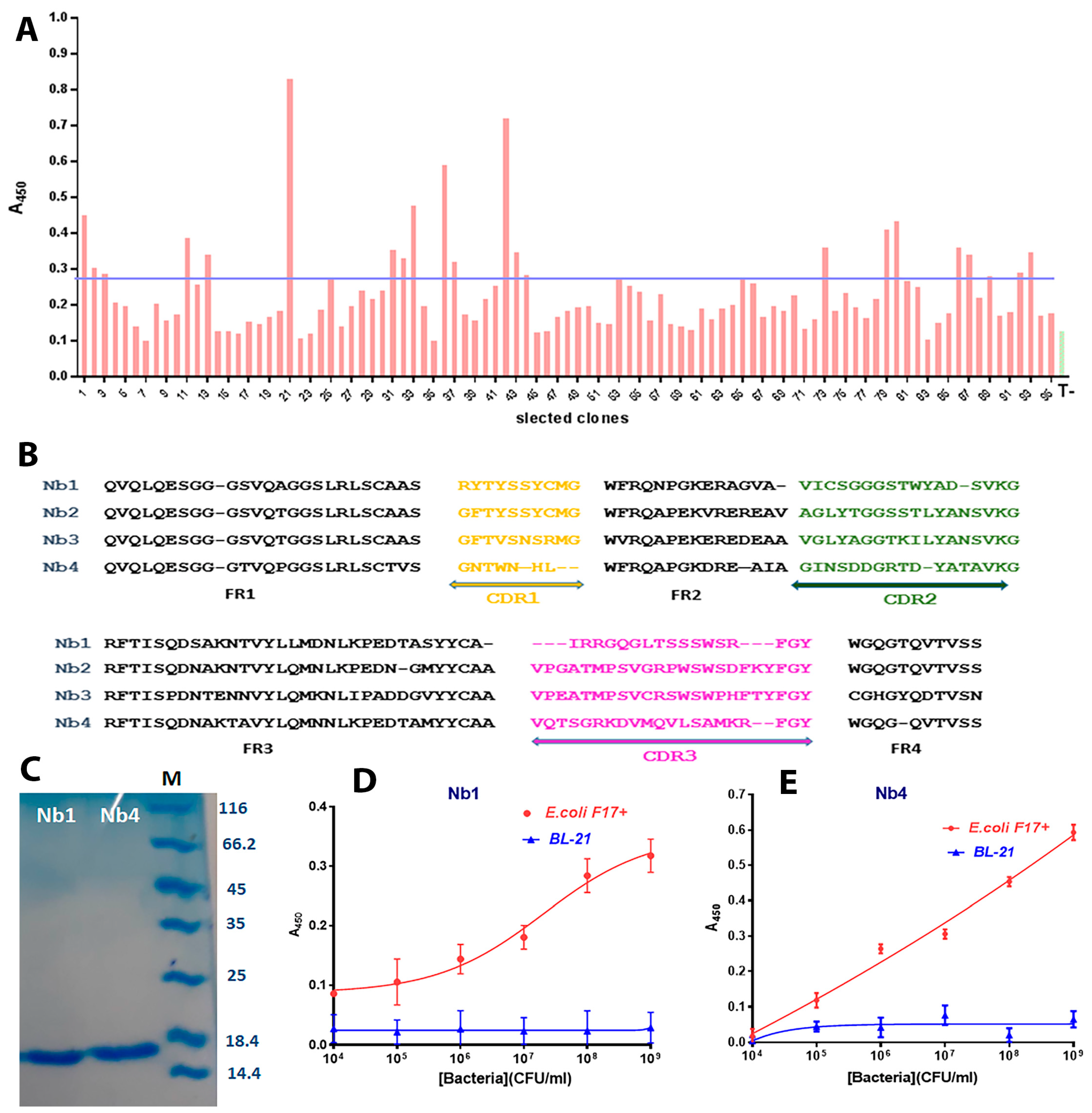
3.1. Library Construction and Selection of F17A-Specific Nanobodies
3.2. Design of the Nanobody-Sandwiched Immunoassay
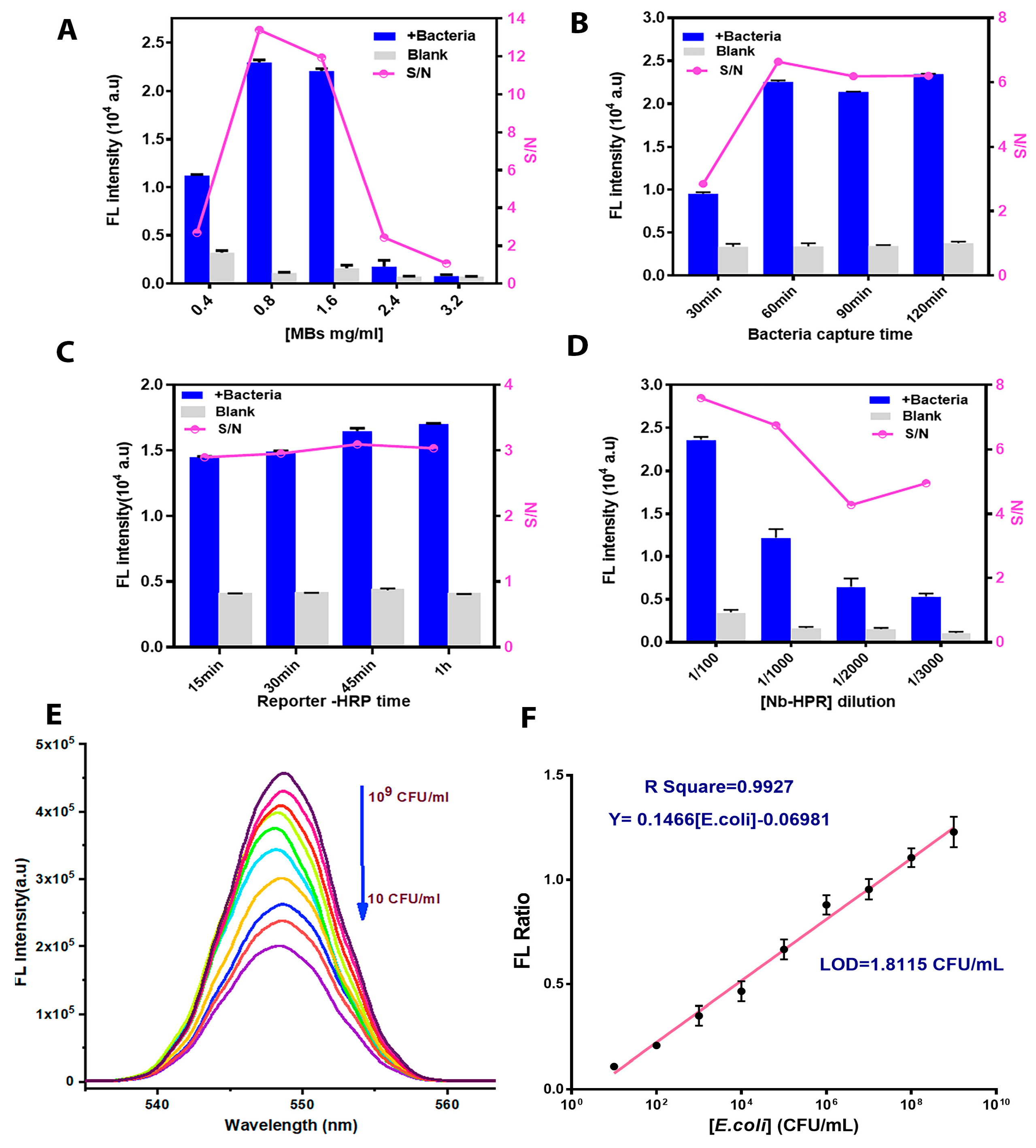
3.3. Immunoassay Parameter Optimization
3.4. Performance Analysis of the Immunoassay
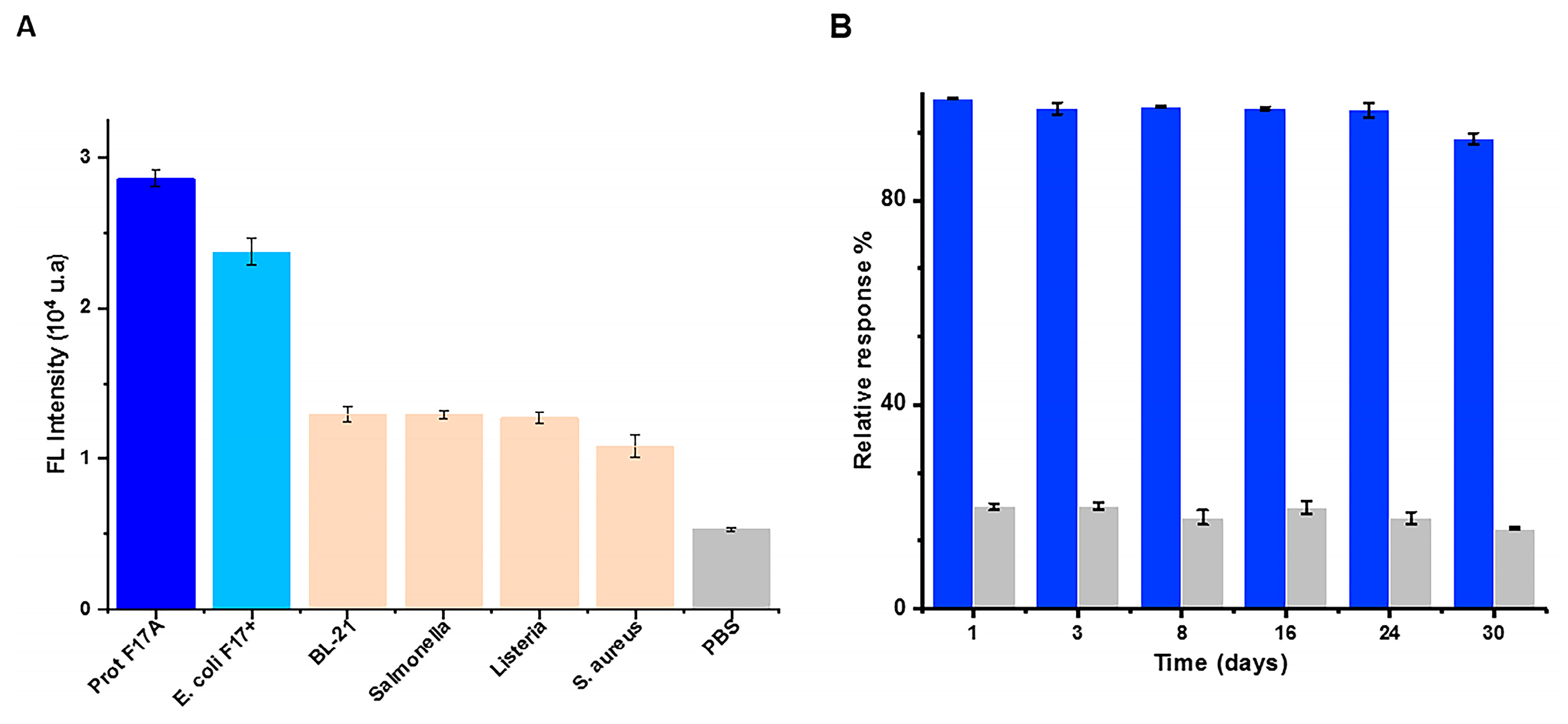
3.5. Specificity and Stability of the Sandwich Immunoassay
3.6. Spike and Recovery Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, D.; Banting, G.; Neumann, N.F. A review of the taxonomy, genetics, and biology of the genus Escherichia and the type species Escherichia coli . Can. J. Microbiol. 2021, 67, 553–571. [Google Scholar] [CrossRef] [PubMed]
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli . Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.G.; Zhou, X.; Kaper, J.B. Adherence of diarrheagenic Escherichia coli strains to epithelial cells. Infect. Immun. 2005, 73, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubreuil, J.D.; Isaacson, R.E.; Schifferli, D.M. Animal Enterotoxigenic Escherichia coli . EcoSal Plus 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Bessalah, S.; Fairbrother, J.M.; Salhi, I.; Vanier, G.; Khorchani, T.; Seddik, M.M.; Hammadi, M. Antimicrobial resistance and molecular characterization of virulence genes, phylogenetic groups of Escherichia coli isolated from diarrheic and healthy camel-calves in Tunisia. Comp. Immunol. Microbiol. Infect. Dis. 2016, 49, 1–7. [Google Scholar] [CrossRef]
- Siuce, J.; Maturrano, L.; Wheeler, J.C.; Rosadio, R. Diarrheagenic Escherichia coli isolates from neonatal alpacas mainly display F17 fimbriae adhesion gene. Trop. Anim. Health Prod. 2020, 52, 3917–3921. [Google Scholar] [CrossRef]
- Stordeur, P.; Marlier, D.; Blanco, J.; Oswald, E.; Biet, F.; Dho-Moulin, M.; Mainil, J. Examination of Escherichia coli from poultry for selected adhesin genes important in disease caused by mammalian pathogenic E. coli . Vet. Microbiol. 2002, 84, 231–241. [Google Scholar] [CrossRef]
- Van Duijkeren, E.; van Asten, A.J.A.M.; Gaastra, W. Characterization of Escherichia coli isolated from adult horses with and without enteritis. Vet. Q. 2000, 22, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Su, Z.; Li, W.; Ren, J. Recent Progresses on Biosensors for Escherichia coli Detection. Food Anal. Methods 2021, 15, 338–366. [Google Scholar] [CrossRef]
- Cesewski, E.; Johnson, B.N. Electrochemical biosensors for pathogen detection. Biosens. Bioelectron. 2020, 159, 112214. [Google Scholar] [CrossRef]
- Salhi, I.; Rabti, A.; Dhehibi, A.; Raouafi, N. Sandwich-Based Immunosensor for Dual-Mode Detection of Pathogenic F17-Positive Escherichia coli Strains. Int. J. Mol. Sci. 2022, 23, 6028. [Google Scholar] [CrossRef]
- Patris, S.; Vandeput, M.; Kauffmann, J.-M. Antibodies as target for affinity biosensors. TrAC Trends Anal. Chem. 2016, 79, 239–246. [Google Scholar] [CrossRef]
- Arbabi Ghahroudi, M.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett. 1997, 414, 521–526. [Google Scholar] [CrossRef] [Green Version]
- De Vlieger, D.; Ballegeer, M.; Rossey, I.; Schepens, B.; Saelens, X. Single-Domain Antibodies and Their Formatting to Combat Viral Infections. Antibodies 2018, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Bastos-Soares, E.A.; Sousa, R.M.O.; Gómez, A.F.; Alfonso, J.; Kayano, A.M.; Zanchi, F.B.; Funes-Huacca, M.E.; Stábeli, R.G.; Soares, A.M.; Pereira, S.S.; et al. Single domain antibodies in the development of immunosensors for diagnostics. Int. J. Biol. Macromol. 2020, 165, 2244–2252. [Google Scholar] [CrossRef]
- Liu, X.; Wang, D.; Chu, J.; Xu, Y.; Wang, W. Sandwich pair nanobodies, a potential tool for electrochemical immunosensing serum prostate-specific antigen with preferable specificity. J. Pharm. Biomed. Anal. 2018, 158, 361–369. [Google Scholar] [CrossRef]
- El-Moghazy, A.Y.; Huo, J.; Amaly, N.; Vasylieva, N.; Hammock, B.D.; Sun, G. An Innovative Nanobody-Based Electrochemical Immunosensor Using Decorated Nylon Nanofibers for Point-of-Care Monitoring of Human Exposure to Pyrethroid Insecticides. ACS Appl. Mater. Interfaces 2020, 12, 6159–6168. [Google Scholar] [CrossRef] [PubMed]
- Morales-Yánez, F.; Trashin, S.; Hermy, M.; Sariego, I.; Polman, K.; Muyldermans, S.; De Wael, K. Fast One-Step Ultrasensitive Detection of Toxocara canis Antigens by a Nanobody-Based Electrochemical Magnetosensor. Anal. Chem. 2019, 91, 11582–11588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Shi, L.; Feng, H.; Zhou, H.S. Single domain antibody coated gold nanoparticles as enhancer for Clostridium difficile toxin detection by electrochemical impedance immunosensors. Bioelectrochemistry 2015, 101, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baral, T.N.; MacKenzie, R.; Arbabi Ghahroudi, M. Single-Domain Antibodies and Their Utility. Curr. Protoc. Immunol. 2013, 103, 2–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pardon, E.; Laeremans, T.; Triest, S.; Rasmussen, S.G.F.; Wohlkönig, A.; Ruf, A.; Muyldermans, S.; Hol, W.G.J.; Kobilka, B.K.; Steyaert, J. A general protocol for the generation of Nanobodies for structural biology. Nat. Protoc. 2014, 9, 674–693. [Google Scholar] [CrossRef] [PubMed]
- Bessalah, S.; Fairbrother, J.M.; Salhi, I.; Vanier, G.; Khorchani, T.; Seddik, M.-M.; Hammadi, M. Characterization and antimicrobial susceptibility of Escherichia coli isolated from healthy farm animals in Tunisia. Anim. Biotechnol. 2020, 32, 748–757. [Google Scholar] [CrossRef] [PubMed]
- Campuzano, S.; Salema, V.; Moreno-Guzmán, M.; Gamella, M.; Yáñez-Sedeño, P.; Fernández, L.A.; Pingarrón, J.M. Disposable amperometric magnetoimmunosensors using nanobodies as biorecognition element. Determination of fibrinogen in plasma. Biosens. Bioelectron. 2014, 52, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Zayani, R.; Rezig, D.; Fares, W.; Marrakchi, M.; Essafi, M.; Raouafi, N. Multiplexed Magnetofluorescent Bioplatform for the Sensitive Detection of SARS-CoV-2 Viral RNA without Nucleic Acid Amplification. Anal. Chem. 2021, 93, 11225–11232. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sun, Y.; Gu, J.; Yang, F.; Wu, S.; Zhang, C.; Ji, X.; Lv, H.; Muyldermans, S.; Wang, S. Selection of specific nanobodies to develop an immuno-assay detecting Staphylococcus aureus in milk. Food Chem. 2021, 353, 129481. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Chen, L.; Wang, Y.; Zhang, K.; Wu, H.; Liu, Y.; Wang, Y.; Wang, J. Development of a Double Nanobody-Based Sandwich Immunoassay for the Detecting Staphylococcal Enterotoxin C in Dairy Products. Foods 2021, 10, 2426. [Google Scholar] [CrossRef]
- Ye, L.; Zhao, G.; Dou, W. An electrochemical immunoassay for Escherichia coli O157:H7 using double functionalized Au@Pt/SiO2 nanocomposites and immune magnetic nanoparticles. Talanta 2018, 182, 354–362. [Google Scholar] [CrossRef]
- Kumar, S.; Guo, Z.; Singh, R.; Wang, Q.; Zhang, B.; Cheng, S.; Liu, F.Z.; Marques, C.; Kaushik, B.K.; Jha, R. MoS2 Functionalized Multicore Fiber Probes for Selective Detection of Shigella Bacteria Based on Localized Plasmon. J. Light. Technol. 2021, 39, 4069–4081. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, J.; Ma, H.; Jiang, Y.; Huang, C.; Han, E.; Yao, B.; He, Y. Optimized dendrimer-encapsulated gold nanoparticles and enhanced carbon nanotube nanoprobes for amplified electrochemical immunoassay of E. coli in dairy product based on enzymatically induced deposition of polyaniline. Biosens. Bioelectron. 2016, 80, 666–673. [Google Scholar] [CrossRef]
- Wilson, D.; Materón, E.M.; Ibáñez-Redín, G.; Faria, R.C.; Correa, D.S.; Oliveira, O.N. Electrical detection of pathogenic bacteria in food samples using information visualization methods with a sensor based on magnetic nanoparticles functionalized with antimicrobial peptides. Talanta 2019, 194, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, R.; Li, Y. Rapid detection of Escherichia coli O157:H7 and Salmonella Typhimurium in foods using an electrochemical immunosensor based on screen-printed interdigitated microelectrode and immunomagnetic separation. Talanta 2016, 148, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, S.; Nejad, M.A.F.; Parolo, C.; Shahrokhian, S.; Merkoçi, A. Smart Chip for Visual Detection of Bacteria Using the Electrochromic Properties of Polyaniline. Anal. Chem. 2019, 91, 14960–14966. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Ryu, J.; Seol, I.-H.; Jeong, K.-B.; You, S.-M.; Kim, Y.-R. Paper-Based Radial Chromatographic Immunoassay for the Detection of Pathogenic Bacteria in Milk. ACS Appl. Mater. Interfaces 2019, 11, 46472–46478. [Google Scholar] [CrossRef] [PubMed]
- Malvano, F.; Pilloton, R.; Albanese, D. Sensitive Detection of Escherichia coli O157:H7 in Food Products by Impedimetric Immunosensors. Sensors 2018, 18, 2168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Güner, A.; Çevik, E.; Şenel, M.; Alpsoy, L. An electrochemical immunosensor for sensitive detection of Escherichia coli O157:H7 by using chitosan, MWCNT, polypyrrole with gold nanoparticles hybrid sensing platform. Food Chem. 2017, 229, 358–365. [Google Scholar] [CrossRef]
- Sun, X.; Lei, C.; Guo, L.; Zhou, Y. Separable detecting of Escherichia coli O157H:H7 by a giant magneto-resistance-based bio-sensing system. Sens. Actuators B Chem. 2016, 234, 485–492. [Google Scholar] [CrossRef]
- Cimafonte, M.; Fulgione, A.; Gaglione, R.; Papaianni, M.; Capparelli, R.; Arciello, A.; Bolletti Censi, S.; Borriello, G.; Velotta, R.; Della Ventura, B. Screen Printed Based Impedimetric Immunosensor for Rapid Detection of Escherichia coli in Drinking Water. Sensors 2020, 20, 274. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Platforms | Antibodies | Linear Range (CFU/mL) | LOD (CFU/mL) | Spiked Real Sample | References |
|---|---|---|---|---|---|
| Straptavidin-coated magnetic beads | mAb | 102–106 | 3.9 × 102 | Chicken rinse water | [32] |
| Electropolymerizing PANI on an ITO screen-printed a electrode | pAb | 102–106 | 10 | Tap and River water | [33] |
| Gold surface | mAb | 100–105 | 103 | Milk | [34] |
| Dendrimer | mAb | 3 × 100–3 × 105 | 3 | Meat and Milk | [35] |
| PPy/AuNPs/MWCNTs/Chitosane hybrid modified pencil graphite electrode (PGE) b | mAb | 3 × 101–3 × 107 | 30 | NR | [36] |
| Poly(amidoamine)dendrimer-encapsulated AuNPs | pAb | 102–106 | 50 | Dairy products (Milk, yogurt…) | [30] |
| Streptavidin-functionalized dynabeads | mAb and pAb | 0–500 | 102 | Orange juice | [37] |
| Gold surface | pAb | 101–108 | 30 | Drinking water | [38] |
| AuNPs/SPCE c | pAb | 102–109 | 37 | NR | [12] |
| Carboxyl Magnetic beads | Nb | 101–109 | 1.8 | Fecal sample | This work |
| E. coli Concentration (CFU/mL) | |||
|---|---|---|---|
| Spiked | Detected | Recovery(%) | |
| 1 | 1 × 105 1 × 106 1 × 107 1 × 108 | (1.02 ± 0.01) × 105 (0.89 ± 0.001) × 106 (9.19 ± 2.0) × 106 (0.99 ± 0.0008) × 108 | 102 89 91.9 99 |
| 2 | 1 × 105 1 × 106 1 × 107 1 × 108 | (0.99 ± 0.04) × 105 (0.09 ± 1.6) × 107 (0.944 ± 8.8) × 107 (9.4 ± 0.0001) × 107 | 99 90 94.4 94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhehibi, A.; Allaoui, A.; Raouafi, A.; Terrak, M.; Bouhaouala-Zahar, B.; Hammadi, M.; Raouafi, N.; Salhi, I. Nanobody-Based Sandwich Immunoassay for Pathogenic Escherichia coli F17 Strain Detection. Biosensors 2023, 13, 299. https://doi.org/10.3390/bios13020299
Dhehibi A, Allaoui A, Raouafi A, Terrak M, Bouhaouala-Zahar B, Hammadi M, Raouafi N, Salhi I. Nanobody-Based Sandwich Immunoassay for Pathogenic Escherichia coli F17 Strain Detection. Biosensors. 2023; 13(2):299. https://doi.org/10.3390/bios13020299
Chicago/Turabian StyleDhehibi, Asma, Abdelmounaaim Allaoui, Amal Raouafi, Mohammed Terrak, Balkiss Bouhaouala-Zahar, Mohamed Hammadi, Noureddine Raouafi, and Imed Salhi. 2023. "Nanobody-Based Sandwich Immunoassay for Pathogenic Escherichia coli F17 Strain Detection" Biosensors 13, no. 2: 299. https://doi.org/10.3390/bios13020299