Comparative Analysis of Cell Metabolic Activity Sensing by Escherichia coli rrnB P1-lux and Cd Responsive-Lux Biosensors: Time-Resolved Experiments and Mechanistic Modelling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Luminescent Whole-Cell Bacterial Reporters
2.2. Cell Growth Condition, and Nutritive Medium Composition in Bioluminescence Assays
2.3. Bioluminescence Measurements
3. Theory
3.1. Kinetics of Bioluminescence Emission by Non-Specific Lux-Biosensors Involving a Constitutive Promoter
3.2. Comparison with Bioluminescence Expression for Metal-Inducible Whole-Cell Lux-Biosensor
3.3. Inferring Time-Dependent Photoactivity of Constitutive Cells from Their Bioluminescence Signal
4. Results and Discussion
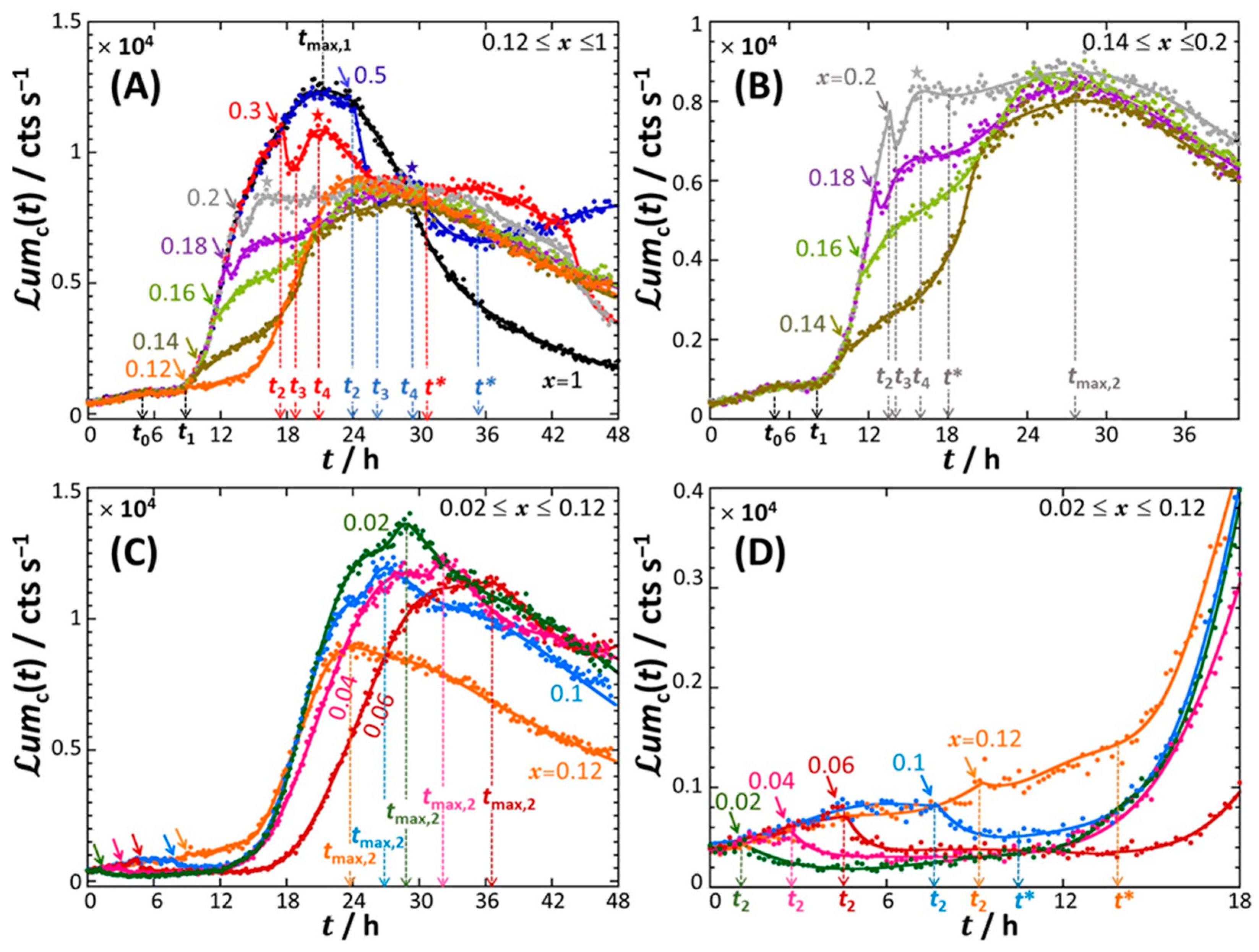
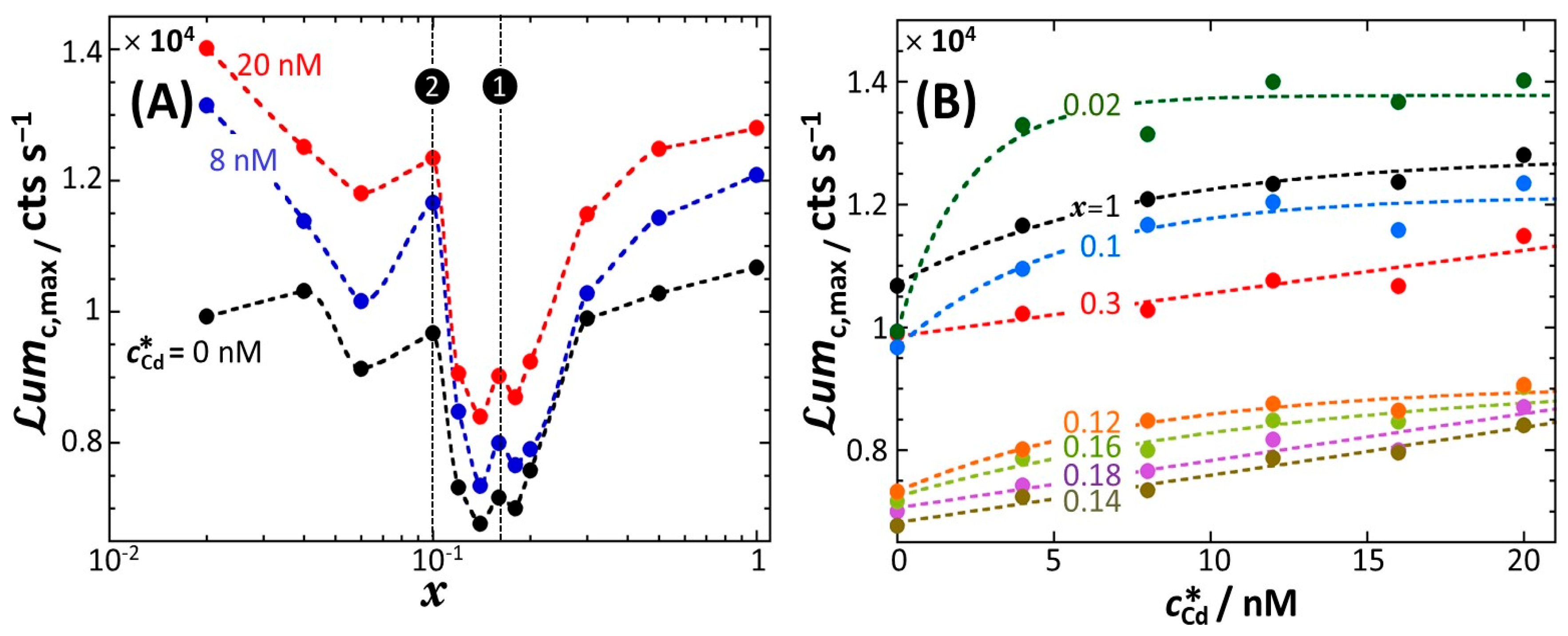
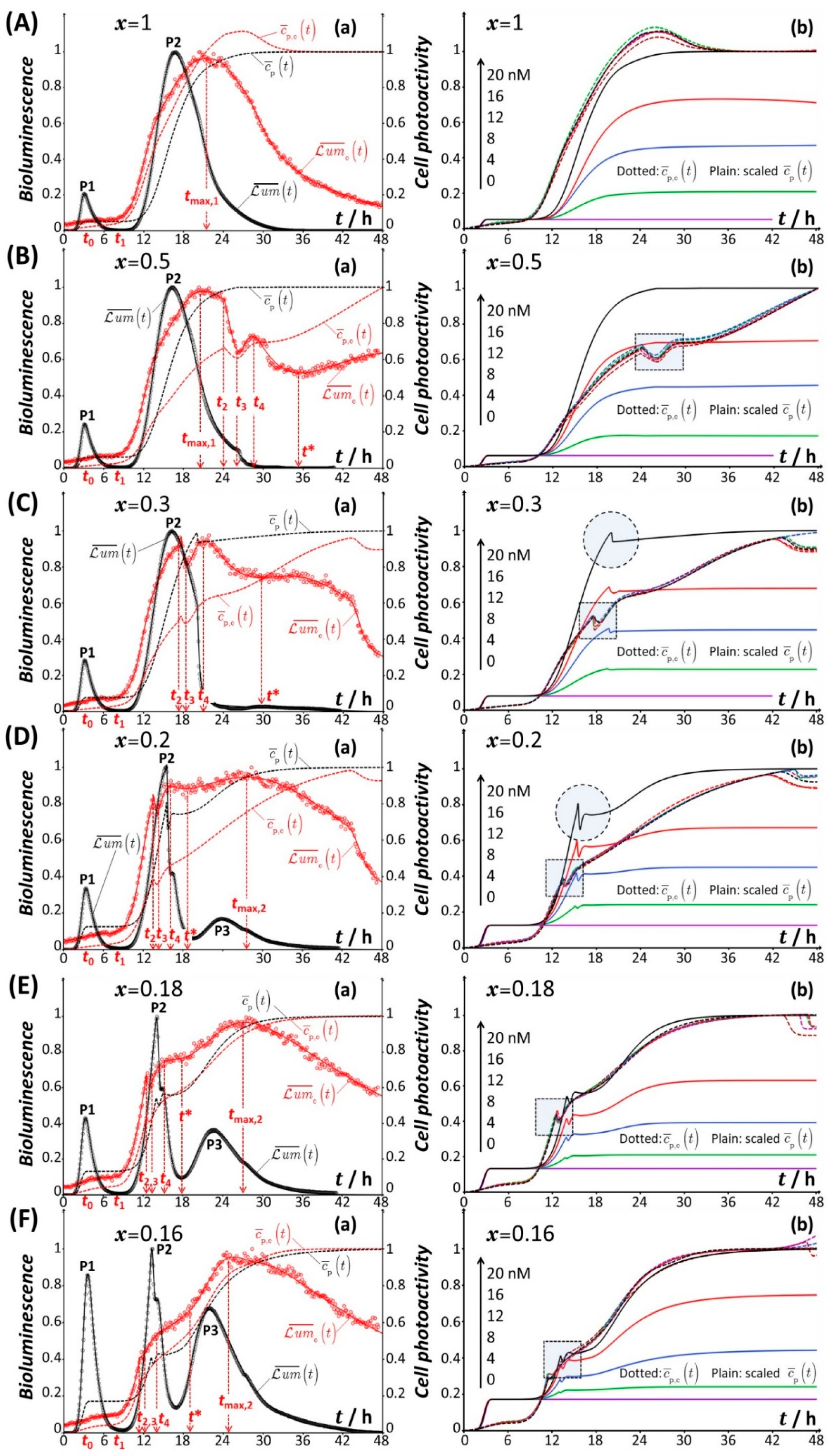
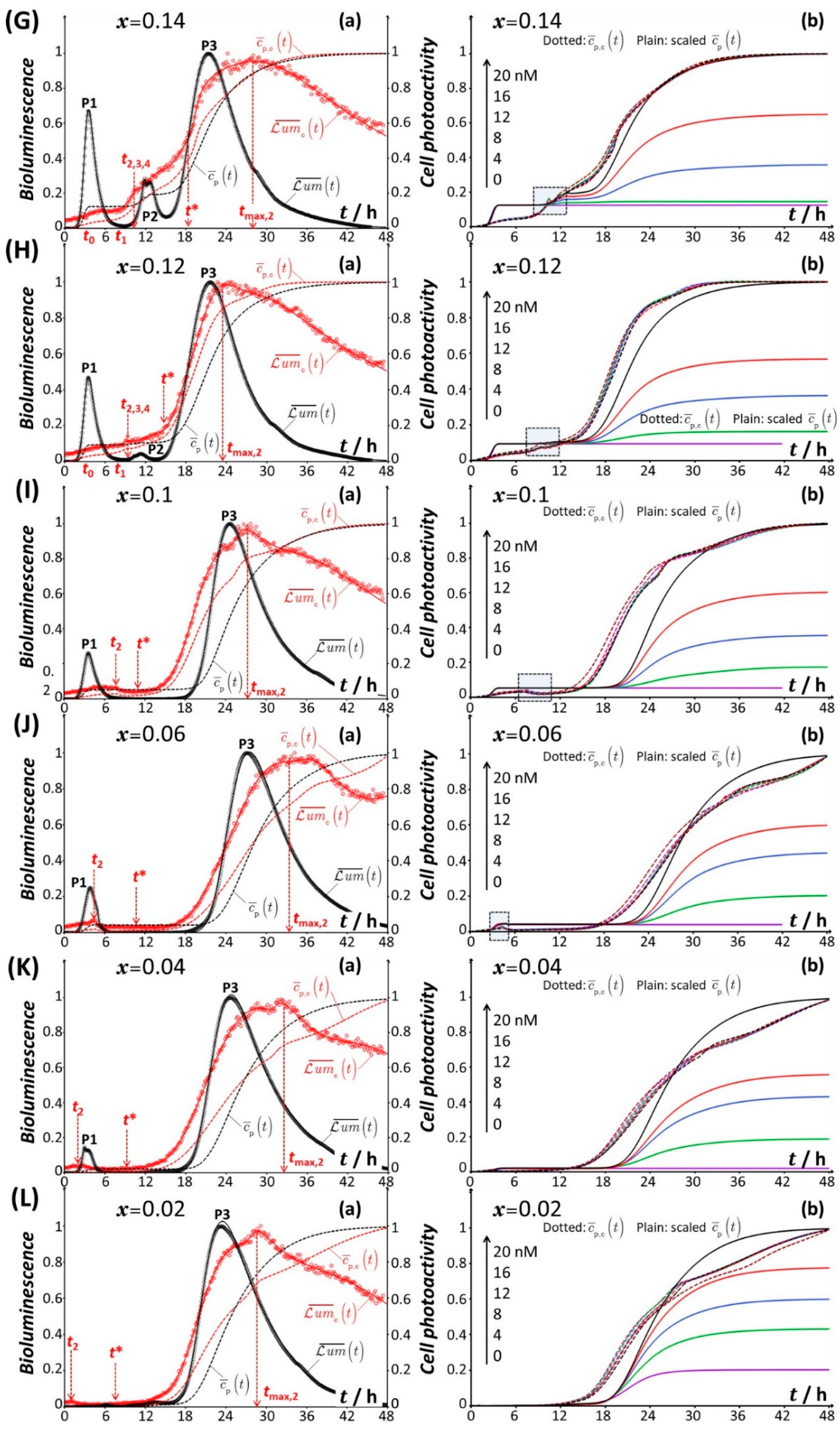
4.1. Description of the Time-Dependent Response of Non-Specific rrnB P1-luxCDABE E. coli Sensors versus Nutritional Medium Conditions



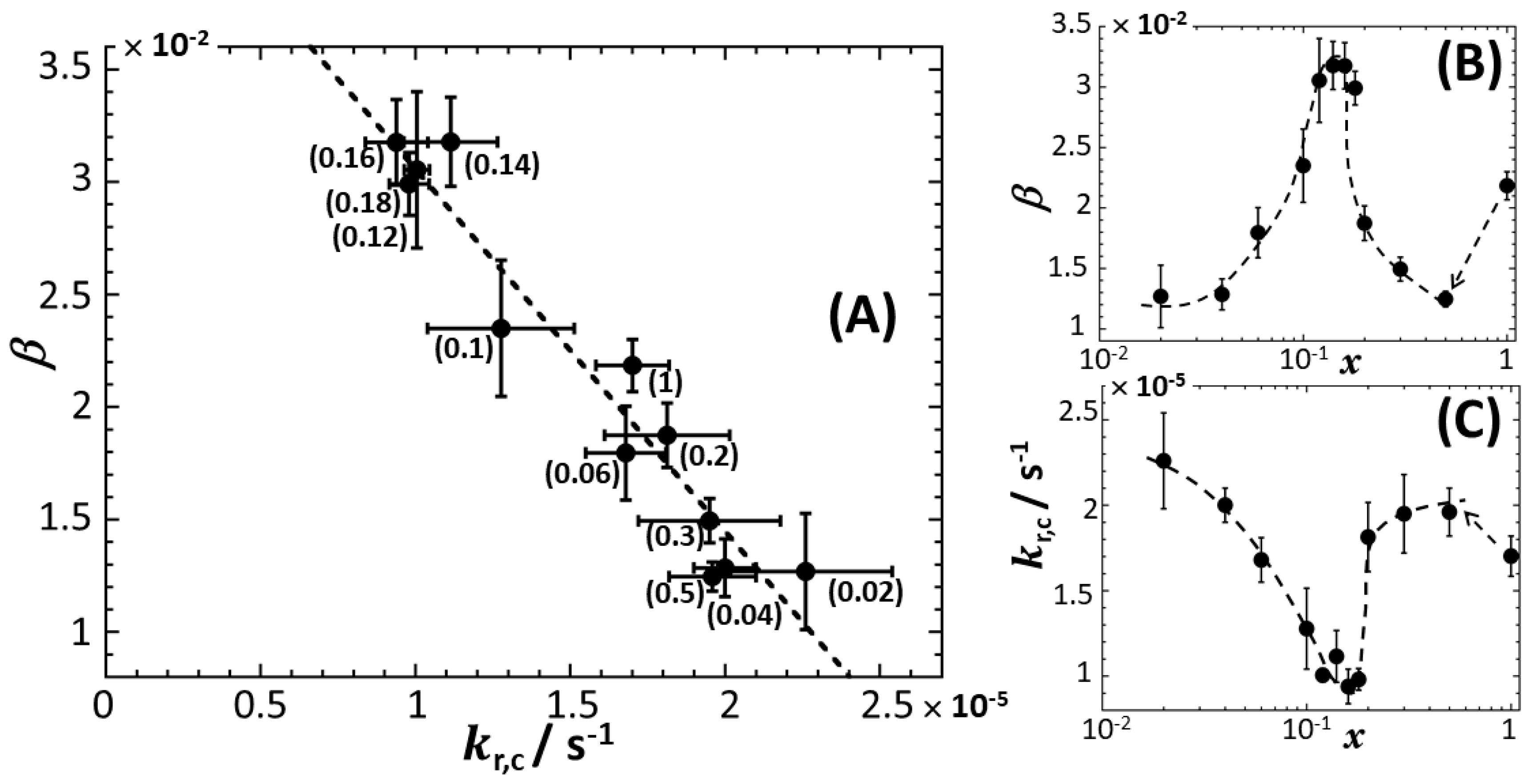
4.2. Rationale for the Dependence of Constitutive Cell Response on Time


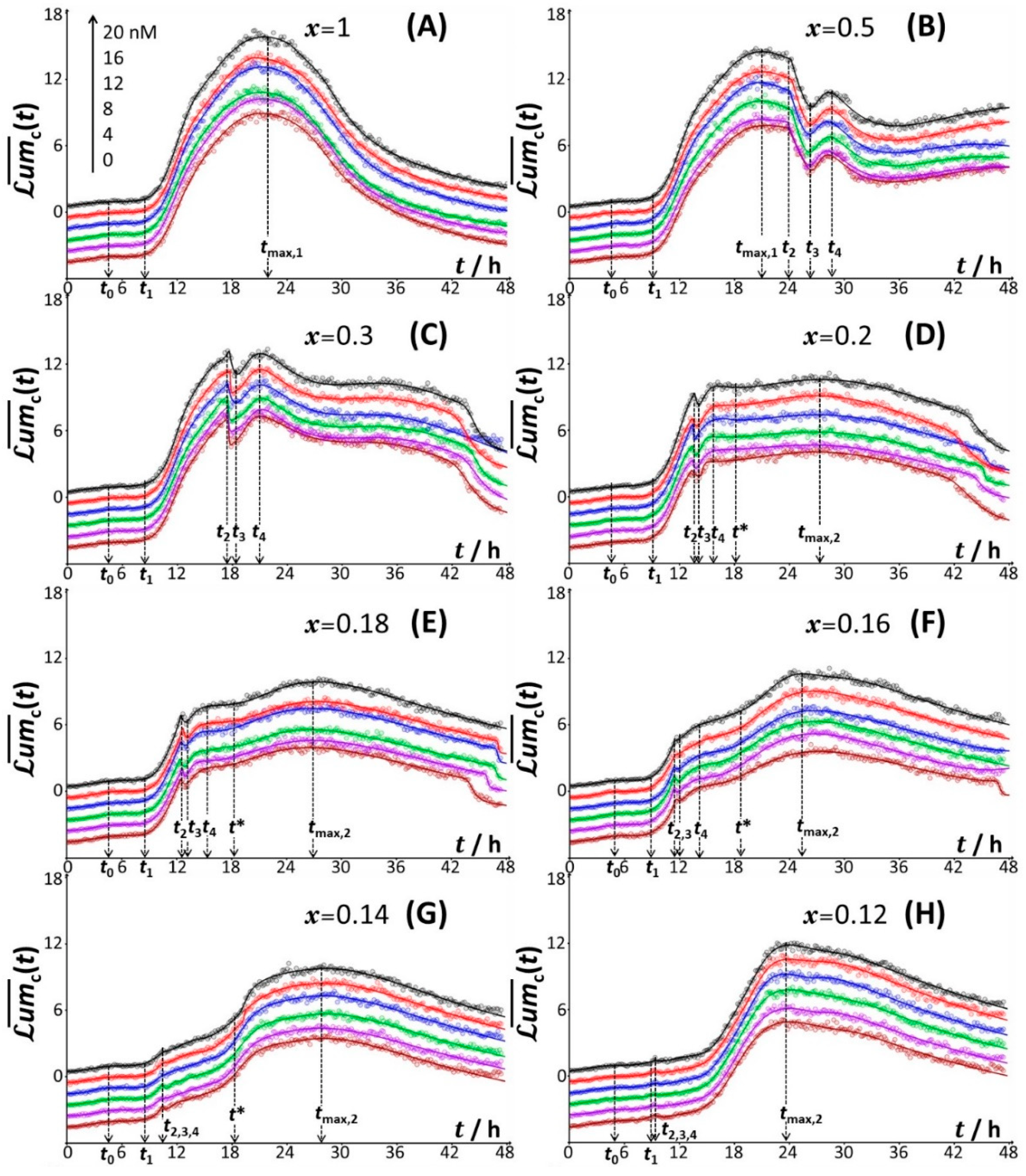
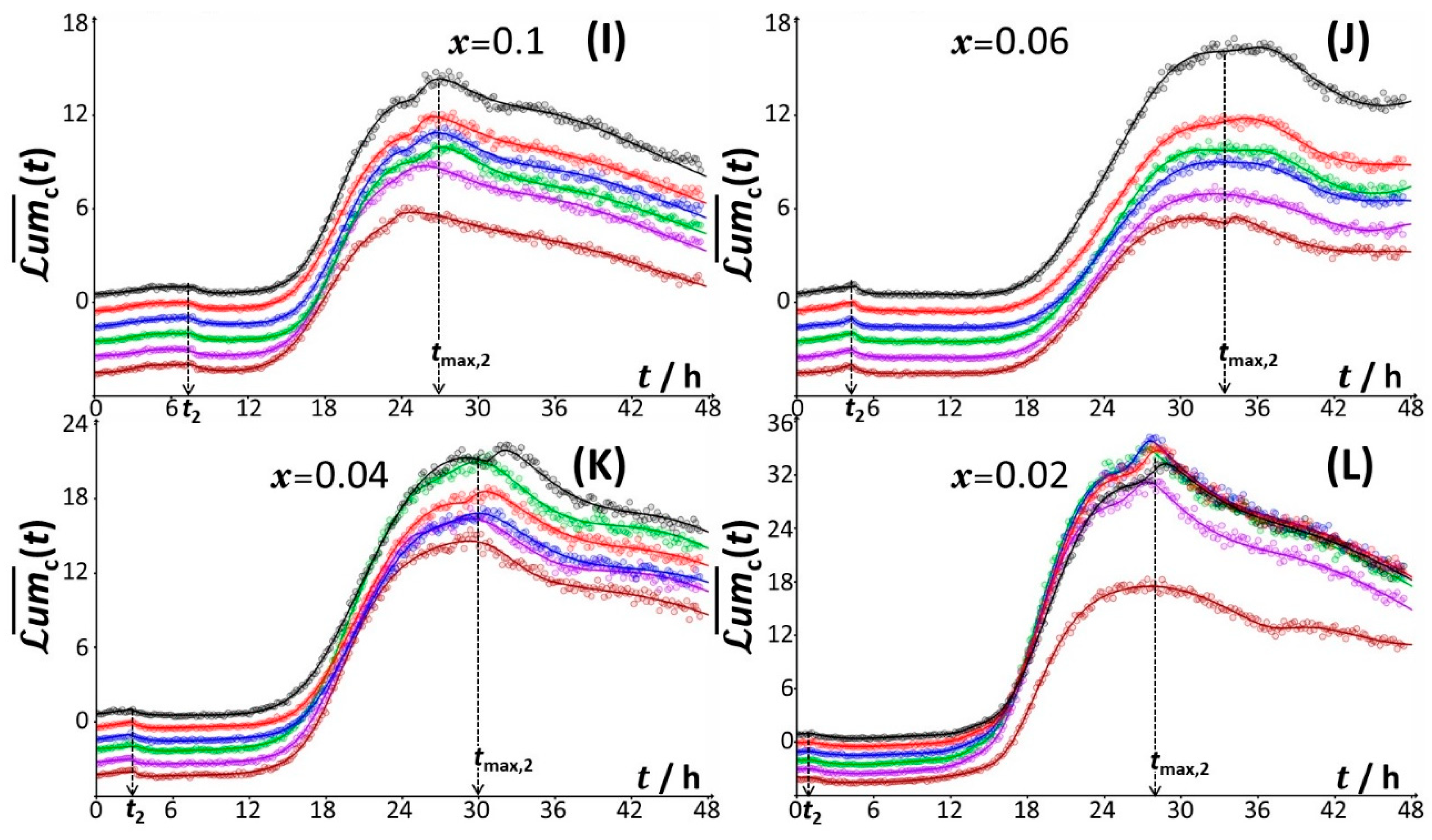
4.2.1. Mechanistic Correspondence between Bioluminescence Signals of Constitutive and Non-Constitutive Cells
4.2.2. Comparison between Time-Dependent Cell Photoactivity Profiles for Constitutive and Non-Constitutive Cells

5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hillger, J.M.; Schoop, J.; Boomsma, D.I.; Slagboom, P.E.; IJzerman, A.P.; Heitman, L.H. Whole-Cell Biosensor for Label-Free Detection of GPCR-Mediated Drug Responses in Personal Cell Lines. Biosens. Bioelectron. 2015, 74, 233–242. [Google Scholar] [CrossRef]
- Gui, Q.; Lawson, T.; Shan, S.; Yan, L.; Liu, Y. The Application of Whole Cell-Based Biosensors for Use in Environmental Analysis and in Medical Diagnostics. Sensors 2017, 17, E1623. [Google Scholar] [CrossRef]
- Kylilis, N.; Riangrungroj, P.; Lai, H.-E.; Salema, V.; Fernández, L.Á.; Stan, G.-B.V.; Freemont, P.S.; Polizzi, K.M. Whole-Cell Biosensor with Tunable Limit of Detection Enables Low-Cost Agglutination Assays for Medical Diagnostic Applications. ACS Sens. 2019, 4, 370–378. [Google Scholar] [CrossRef]
- Raut, N.; O’Connor, G.; Pasini, P.; Daunert, S. Engineered Cells as Biosensing Systems in Biomedical Analysis. Anal. Bioanal. Chem. 2012, 402, 3147–3159. [Google Scholar] [CrossRef]
- Visaggio, D.; Pirolo, M.; Frangipani, E.; Lucidi, M.; Sorrentino, R.; Mitidieri, E.; Ungaro, F.; Luraghi, A.; Peri, F.; Visca, P. A Highly Sensitive Luminescent Biosensor for the Microvolumetric Detection of the Pseudomonas Aeruginosa Siderophore Pyochelin. ACS Sens. 2021, 6, 3273–3283. [Google Scholar] [CrossRef]
- Zhang, G.; Hu, S.; Jia, X. Highly Sensitive Whole-Cell Biosensor for Cadmium Detection Based on a Negative Feedback Circuit. Front. Bioeng. Biotechnol. 2021, 9, 799781. [Google Scholar] [CrossRef]
- Roy, R.; Ray, S.; Chowdhury, A.; Anand, R. Tunable Multiplexed Whole-Cell Biosensors as Environmental Diagnostics for Ppb-Level Detection of Aromatic Pollutants. ACS Sens. 2021, 6, 1933–1939. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, C.-W.; Wang, D.; Wei, N. A Whole-Cell Biosensor for Point-of-Care Detection of Waterborne Bacterial Pathogens. ACS Synth. Biol. 2021, 10, 333–344. [Google Scholar] [CrossRef]
- Barger, N.; Oren, I.; Li, X.; Habib, M.; Daniel, R. A Whole-Cell Bacterial Biosensor for Blood Markers Detection in Urine. ACS Synth. Biol. 2021, 10, 1132–1142. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, B.; Zhu, Z.; Xin, X.; Wu, H.; Chen, B. Microfluidic Based Whole-Cell Biosensors for Simultaneously On-Site Monitoring of Multiple Environmental Contaminants. Front. Bioeng. Biotechnol. 2021, 9, 622108. [Google Scholar] [CrossRef]
- Branco, R.; Cristóvão, A.; Morais, P.V. Highly Sensitive, Highly Specific Whole-Cell Bioreporters for the Detection of Chromate in Environmental Samples. PLoS ONE 2013, 8, e54005. [Google Scholar] [CrossRef]
- Sørensen, S.J.; Burmølle, M.; Hansen, L.H. Making Bio-Sense of Toxicity: New Developments in Whole-Cell Biosensors. Curr. Opin. Biotechnol. 2006, 17, 11–16. [Google Scholar] [CrossRef]
- van der Meer, J.R.; Belkin, S. Where Microbiology Meets Microengineering: Design and Applications of Reporter Bacteria. Nat. Rev. Microbiol. 2010, 8, 511–522. [Google Scholar] [CrossRef]
- da Silva Nunes-Halldorson, V.; Duran, N.L. Bioluminescent Bacteria: Lux Genes as Environmental Biosensors. Braz. J. Microbiol. 2003, 34, 91–96. [Google Scholar] [CrossRef]
- Romero-Freire, A.; Joonas, E.; Muna, M.; Cossu-Leguille, C.; Vignati, D.A.L.; Giamberini, L. Assessment of the Toxic Effects of Mixtures of Three Lanthanides (Ce, Gd, Lu) to Aquatic Biota. Sci. Total Environ. 2019, 661, 276–284. [Google Scholar] [CrossRef]
- Gupta, N.; Renugopalakrishnan, V.; Liepmann, D.; Paulmurugan, R.; Malhotra, B.D. Cell-Based Biosensors: Recent Trends, Challenges and Future Perspectives. Biosens. Bioelectron. 2019, 141, 111435. [Google Scholar] [CrossRef]
- Kessenikh, A.G.; Novoyatlova, U.S.; Bazhenov, S.V.; Stepanova, E.A.; Khrulnova, S.A.; Gnuchikh, E.Y.; Kotova, V.Y.; Kudryavtseva, A.A.; Bermeshev, M.V.; Manukhov, I.V. Constructing of Bacillus Subtilis-Based Lux-Biosensors with the Use of Stress-Inducible Promoters. Int. J. Mol. Sci. 2021, 22, 9571. [Google Scholar] [CrossRef]
- Bereza-Malcolm, L.T.; Mann, G.; Franks, A.E. Environmental Sensing of Heavy Metals through Whole Cell Microbial Biosensors: A Synthetic Biology Approach. ACS Synth. Biol. 2015, 4, 535–546. [Google Scholar] [CrossRef]
- Jia, X.; Bu, R.; Zhao, T.; Wu, K. Sensitive and Specific Whole-Cell Biosensor for Arsenic Detection. Appl. Environ. Microbiol. 2019, 85, e00694-19. [Google Scholar] [CrossRef]
- Shah, N.; Naseby, D.C. Bioluminescence-Based Measurement of Viability of Pseudomonas Aeruginosa ATCC 9027 Harbouring Plasmid-Based Lux Genes under the Control of Constitutive Promoters. J. Appl. Microbiol. 2014, 117, 1373–1387. [Google Scholar] [CrossRef]
- Dalzell, D.J.B.; Alte, S.; Aspichueta, E.; de la Sota, A.; Etxebarria, J.; Gutierrez, M.; Hoffmann, C.C.; Sales, D.; Obst, U.; Christofi, N. A Comparison of Five Rapid Direct Toxicity Assessment Methods to Determine Toxicity of Pollutants to Activated Sludge. Chemosphere 2002, 47, 535–545. [Google Scholar] [CrossRef]
- Shin, Y.; Qayyum, M.; Pupov, D.; Esyunina, D.; Kulbachinskiy, A.; Murakami, K. Structural Basis of Ribosomal RNA Transcription Regulation. Nat. Commun. 2021, 12, 528. [Google Scholar] [CrossRef] [PubMed]
- Duval, J.F.L.; Pagnout, C. Bimodal Stringence-Mediated Response of Metal-Detecting Luminescent Whole Cell Bioreporters: Experimental Evidence and Quantitative Theory. Sens. Actuators B Chem. 2020, 309, 127751. [Google Scholar] [CrossRef]
- Till, P.; Toepel, J.; Bühler, B.; Mach, R.L.; Mach-Aigner, A.R. Regulatory Systems for Gene Expression Control in Cyanobacteria. Appl. Microbiol. Biotechnol. 2020, 104, 1977–1991. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, X.; Xu, M.; Fang, Y.; Wang, Y.; Sun, G.; Guo, J. Identification of Stress-Responsive Transcription Factors with Protein-Bound Escherichia Coli Genomic DNA Libraries. AMB Express 2020, 10, 199. [Google Scholar] [CrossRef]
- Kotova, V.Y.; Manukhov, I.V.; Zavilgelskii, G.B. Lux-Biosensors for Detection of SOS-Response, Heat Shock, and Oxidative Stress. Appl. Biochem. Microbiol. 2010, 46, 781–788. [Google Scholar] [CrossRef]
- Baumstark-Khan, C.; Cioara, K.; Rettberg, P.; Horneck, G. Determination of Geno- and Cytotoxicity of Groundwater and Sediments Using the Recombinant SWITCH Test. J. Environ. Sci. Health Part A 2005, 40, 245–263. [Google Scholar] [CrossRef]
- Wang, Y.-Z.; Christopher, J.K.; Yong, Y.-C.; Zhai, D.-D. Nutrient Detection with Whole-Cell Biosensors. In Handbook of Cell Biosensors; Thouand, G., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 747–766. ISBN 978-3-030-23217-7. [Google Scholar]
- Slaveykova, V.I.; Wilkinson, K.J.; Slaveykova, V.I.; Wilkinson, K.J. Predicting the Bioavailability of Metals and Metal Complexes: Critical Review of the Biotic Ligand Model. Environ. Chem. 2005, 2, 9–24. [Google Scholar] [CrossRef]
- van Leeuwen, H.P. Metal Speciation Dynamics and Bioavailability: Inert and Labile Complexes. Environ. Sci. Technol. 1999, 33, 3743–3748. [Google Scholar] [CrossRef]
- Duval, J.F.L.; Rotureau, E. Dynamics of Metal Uptake by Charged Soft Biointerphases: Impacts of Depletion, Internalisation, Adsorption and Excretion. Phys. Chem. Chem. Phys. 2014, 16, 7401–7416. [Google Scholar] [CrossRef]
- van Leeuwen, H.P.; Duval, J.F.L.; Pinheiro, J.P.; Blust, R.; Town, R.M. Chemodynamics and Bioavailability of Metal Ion Complexes with Nanoparticles in Aqueous Media. Environ. Sci. Nano 2017, 4, 2108–2133. [Google Scholar] [CrossRef]
- Duval, J.F.L.; Présent, R.M.; Rotureau, E. Kinetic and Thermodynamic Determinants of Trace Metal Partitioning at Biointerphases: The Role of Intracellular Speciation Dynamics. Phys. Chem. Chem. Phys. 2016, 18, 30415–30435. [Google Scholar] [CrossRef] [PubMed]
- Présent, R.M.; Rotureau, E.; Billard, P.; Pagnout, C.; Sohm, B.; Flayac, J.; Gley, R.; Pinheiro, J.P.; Duval, J.F.L. Impact of Intracellular Metallothionein on Metal Biouptake and Partitioning Dynamics at Bacterial Interfaces. Phys. Chem. Chem. Phys. 2017, 19, 29114–29124. [Google Scholar] [CrossRef] [PubMed]
- Duval, J.F.L. Coupled Metal Partitioning Dynamics and Toxicodynamics at Biointerfaces: A Theory beyond the Biotic Ligand Model Framework. Phys. Chem. Chem. Phys. 2016, 18, 9453–9469. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, J.P.; van Leeuwen, H.P. Metal Speciation Dynamics and Bioavailability. 2. Radial Diffusion Effects in the Microorganism Range. Environ. Sci. Technol. 2001, 35, 894–900. [Google Scholar] [CrossRef]
- Wilkinson, K.J.; Buffle, J. Physicochemical Kinetics and Transport at Biointerfaces; Van Leeuwen Herman, P., Koster, W., Eds.; John Wiley & Sons: Chichester, UK, 2004; Chapter 10. [Google Scholar]
- Duval, J.F.L.; Town, R.M.; van Leeuwen, H.P. Lability of Nanoparticulate Metal Complexes at a Macroscopic Metal Responsive (Bio)Interface: Expression and Asymptotic Scaling Laws. J. Phys. Chem. C 2018, 122, 6052–6065. [Google Scholar] [CrossRef]
- Duval, J.F.L.; Pagnout, C. Decoding the Time-Dependent Response of Bioluminescent Metal-Detecting Whole-Cell Bacterial Sensors. ACS Sens. 2019, 4, 1373–1383. [Google Scholar] [CrossRef]
- Delatour, E.; Pagnout, C.; Zaffino, M.; Duval, J.F.L. Exploiting Catabolite Repression and Stringent Response to Control Delay and Multimodality of Bioluminescence Signal by Metal Whole-Cell Biosensors: Interplay between Metal Bioavailability and Nutritional Medium Conditions. Biosensors 2022, 12, 327. [Google Scholar] [CrossRef]
- Martín-Betancor, K.; Rodea-Palomares, I.; Muñoz-Martín, M.A.; Leganés, F.; Fernández-Piñas, F. Construction of a Self-Luminescent Cyanobacterial Bioreporter That Detects a Broad Range of Bioavailable Heavy Metals in Aquatic Environments. Front. Microbiol. 2015, 6, 186. [Google Scholar] [CrossRef]
- Blanco-Ameijeiras, S.; Cabanes, D.J.E.; Hassler, C.S. Towards the Development of a New Generation of Whole-Cell Bioreporters to Sense Iron Bioavailability in Oceanic Systems-Learning from the Case of Synechococcus Sp. PCC7002 Iron Bioreporter. J. Appl. Microbiol. 2019, 127, 1291–1304. [Google Scholar] [CrossRef] [Green Version]
- Käkinen, A.; Bondarenko, O.; Ivask, A.; Kahru, A. The Effect of Composition of Different Ecotoxicological Test Media on Free and Bioavailable Copper from CuSO4 and CuO Nanoparticles: Comparative Evidence from a Cu-Selective Electrode and a Cu-Biosensor. Sensors 2011, 11, 10502–10521. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, X.; Tefsen, B.; Wells, M. From Speciation to Toxicity: Using a “Two-in-One” Whole-Cell Bioreporter Approach to Assess Harmful Effects of Cd and Pb. Water Res. 2022, 217, 118384. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, B.; Deng, J.; Qin, B.; Wells, M.; Tefsen, B. Advances in Freshwater Risk Assessment: Improved Accuracy of Dissolved Organic Matter-Metal Speciation Prediction and Rapid Biological Validation. Ecotoxicol. Environ. Saf. 2020, 202, 110848. [Google Scholar] [CrossRef] [PubMed]
- Magrisso, S.; Erel, Y.; Belkin, S. Microbial Reporters of Metal Bioavailability: Microbial Reporters of Metal Bioavailability. Microb. Biotechnol. 2008, 1, 320–330. [Google Scholar] [CrossRef]
- Pinheiro, J.P.; Galceran, J.; van Leeuwen, H.P. Metal Speciation Dynamics and Bioavailability: Bulk Depletion Effects. Environ. Sci. Technol. 2004, 38, 2397–2405. [Google Scholar] [CrossRef]
- Pagnout, C.; Présent, R.M.; Billard, P.; Rotureau, E.; Duval, J.F.L. What Do Luminescent Bacterial Metal-Sensors Probe? Insights from Confrontation between Experiments and Flux-Based Theory. Sens. Actuators B Chem. 2018, 270, 482–491. [Google Scholar] [CrossRef]
- Kelly, C.J.; Hsiung, C.-J.; Lajoie, C.A. Kinetic Analysis of Bacterial Bioluminescence. Biotechnol. Bioeng. 2003, 81, 370–378. [Google Scholar] [CrossRef]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia Coli K-12 in-Frame, Single-Gene Knockout Mutants: The Keio Collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef]
- Pupov, D.; Petushkov, I.; Esyunina, D.; Murakami, K.S.; Kulbachinskiy, A. Region 3.2 of the σ Factor Controls the Stability of RRNA Promoter Complexes and Potentiates Their Repression by DksA. Nucleic Acids Res. 2018, 46, 11477–11487. [Google Scholar] [CrossRef]
- Ang, J.; Harris, E.; Hussey, B.J.; Kil, R.; McMillen, D.R. Tuning Response Curves for Synthetic Biology. ACS Synth. Biol. 2013, 2, 547–567. [Google Scholar] [CrossRef]
- De Paepe, B.; Maertens, J.; Vanholme, B.; De Mey, M. Modularization and Response Curve Engineering of a Naringenin-Responsive Transcriptional Biosensor. ACS Synth. Biol. 2018, 7, 1303–1314. [Google Scholar] [CrossRef] [PubMed]
- Santillán, M. On the Use of the Hill Functions in Mathematical Models of Gene Regulatory Networks. Math. Model. Nat. Phenom. 2008, 3, 85–97. [Google Scholar] [CrossRef]
- Delle Side, D.; Nassisi, V.; Pennetta, C.; Alifano, P.; Di Salvo, M.; Talà, A.; Chechkin, A.; Seno, F.; Trovato, A. Bacterial Bioluminescence Onset and Quenching: A Dynamical Model for a Quorum Sensing-Mediated Property. R. Soc. Open Sci. 2017, 4, 171586. [Google Scholar] [CrossRef] [PubMed]
- Deeva, A.A.; Temlyakova, E.A.; Sorokin, A.A.; Nemtseva, E.V.; Kratasyuk, V.A. Structural Distinctions of Fast and Slow Bacterial Luciferases Revealed by Phylogenetic Analysis. Bioinforma. Oxf. Engl. 2016, 32, 3053–3057. [Google Scholar] [CrossRef]
- Tjørve, K.M.C.; Tjørve, E. The Use of Gompertz Models in Growth Analyses, and New Gompertz-Model Approach: An Addition to the Unified-Richards Family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef]
- Salinitro, M.; Mattarello, G.; Guardigli, G.; Odajiu, M.; Tassoni, A. Induction of Hormesis in Plants by Urban Trace Metal Pollution. Sci. Rep. 2021, 11, 20329. [Google Scholar] [CrossRef]
- Lendenmann, U.; Snozzi, M.; Egli, T. Kinetics of the Simultaneous Utilization of Sugar Mixtures by Escherichia Coli in Continuous Culture. Appl. Environ. Microbiol. 1996, 62, 1493–1499. [Google Scholar] [CrossRef]
- Okano, H.; Hermsen, R.; Kochanowski, K.; Hwa, T. Regulation Underlying Hierarchical and Simultaneous Utilization of Carbon Substrates by Flux Sensors in Escherichia Coli. Nat. Microbiol. 2020, 5, 206–215. [Google Scholar] [CrossRef]
- Iqbal, M.; Doherty, N.; Page, A.M.L.; Qazi, S.N.A.; Ajmera, I.; Lund, P.A.; Kypraios, T.; Scott, D.J.; Hill, P.J.; Stekel, D.J. Reconstructing Promoter Activity from Lux Bioluminescent Reporters. PLoS Comput. Biol. 2017, 13, e1005731. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delatour, E.; Pagnout, C.; Zaffino, M.L.; Duval, J.F.L. Comparative Analysis of Cell Metabolic Activity Sensing by Escherichia coli rrnB P1-lux and Cd Responsive-Lux Biosensors: Time-Resolved Experiments and Mechanistic Modelling. Biosensors 2022, 12, 763. https://doi.org/10.3390/bios12090763
Delatour E, Pagnout C, Zaffino ML, Duval JFL. Comparative Analysis of Cell Metabolic Activity Sensing by Escherichia coli rrnB P1-lux and Cd Responsive-Lux Biosensors: Time-Resolved Experiments and Mechanistic Modelling. Biosensors. 2022; 12(9):763. https://doi.org/10.3390/bios12090763
Chicago/Turabian StyleDelatour, Eva, Christophe Pagnout, Marie L. Zaffino, and Jérôme F. L. Duval. 2022. "Comparative Analysis of Cell Metabolic Activity Sensing by Escherichia coli rrnB P1-lux and Cd Responsive-Lux Biosensors: Time-Resolved Experiments and Mechanistic Modelling" Biosensors 12, no. 9: 763. https://doi.org/10.3390/bios12090763