Colorimetric Systems for the Detection of Bacterial Contamination: Strategy and Applications


1
Center for Applied Life Science, Hanbat National University, Daejeon 34158, Korea
2
School of Integrative Engineering, Chung-Ang University, Seoul 06974, Korea
*
Author to whom correspondence should be addressed.
Biosensors 2022, 12(7), 532; https://doi.org/10.3390/bios12070532
Submission received: 9 June 2022
/
Revised: 10 July 2022
/
Accepted: 13 July 2022
/
Published: 16 July 2022
(This article belongs to the Special Issue Smart Materials for Chemical and Biosensing)
Abstract
:Bacterial contamination is a public health concern worldwide causing enormous social and economic losses. For early diagnosis and adequate management to prevent or treat pathogen-related illnesses, extensive effort has been put into the development of pathogenic bacterial detection systems. Colorimetric sensing systems have attracted increasing attention due to their simple and single-site operation, rapid signal readout with the naked eye, ability to operate without external instruments, portability, compact design, and low cost. In this article, recent trends and advances in colorimetric systems for the detection and monitoring of bacterial contamination are reviewed. This article focuses on pathogen detection strategies and technologies based on reaction factors that affect the color change for visual readout. Reactions used in each strategy are introduced by dividing them into the following five categories: external pH change-induced pH indicator reactions, intracellular enzyme-catalyzed chromogenic reactions, enzyme-like nanoparticle (NP)-catalyzed substrate reactions, NP aggregation-based reactions, and NP accumulation-based reactions. Some recently developed colorimetric systems are introduced, and their challenges and strategies to improve the sensing performance are discussed.
1. Introduction
Bacteria can be found in the air, soil, and water and play central roles in ecosystems. However, bacterial contamination poses a huge threat to global public health, affecting the food industry, hospital diagnostics, and society [1,2,3,4]. According to the Centers for Disease Control and Prevention (CDC), around 48 million people suffer from diseases caused by foodborne pathogens every year. Of this, 128,000 people are hospitalized and nearly 3000 people die, resulting in billions of dollars in medical expenses and productivity loss in the United States [5]. One of the major causes of waterborne diseases is bacterial infections associated with drinking contaminated water, especially in developing countries [6,7]. Foodborne and waterborne outbreaks have increased with various factors, such as changes in the emergence of antibiotic-resistant bacteria, the adaptation of bacteria to environmental changes, changes in food processing, production, and distribution, exposure to unsafe drinking water, inadequate sanitation control systems, inadequate public health services, and increased international travel [8,9]. Given this situation, bacterial contamination should be detected and monitored to ensure environmental and food safety and reduce the incidence of bacterial infection-related diseases. There is an urgent need for simple, rapid, on-site, and sensitive methods for early diagnosis and adequate management to prevent or treat pathogen-related illnesses.
Efforts to develop an effective system have led to the development of various biosensors using novel nanomaterials and nanostructures. Biosensors, which are analytical devices, convert recognizable biological signals into directly measurable signals using various sensing methods. Sensing methods such as electrical, electrochemical, mechanical, and optical methods have attracted increasing attention due to their high sensitivity, specificity, small assay volume, and shorter detection time and have been employed for sensitive and specific detection in clinical and environmental settings [10,11,12,13]. Notably, colorimetric sensing systems may be used for the detection of bacterial contamination due to their simple and on-site operation and rapid signal readout with the naked eye that requires no external instrument or detector. This approach has also various advantages such as rapidness, non-contact detection, compact design, cost-effectiveness, portability, and complex data processing [14,15,16,17,18]. These distinct advantages have led to the recent advances in colorimetric detection strategies and technologies, which have been actively applied to detect and monitor bacterial contamination in several fields and have played a key role in fulfilling industrial and academic needs for diagnostics [14,15,16,17,18]. Therefore, it is expected that colorimetric detection systems will have increasingly important roles in point-of-care testing and monitoring pathogenic bacteria in clinical and environmental settings and the food industry.
In this article, we review the recent developments in the field of colorimetric sensing of bacterial contamination for food, water, and environmental safety and their quality control. This article focuses on pathogen detection strategies and technologies based on reactions that affect the color change for signal readout, which can be largely divided into the following five categories: (1) color change generated by external pH change-induced pH-responsive molecules (pH indicator) reactions, (2) color change generated by intracellular enzyme-catalyzed chromogenic reactions, (3) color change generated by enzyme-like nanoparticle (NP)-catalyzed substrate reactions, (4) color change by aggregated NPs, and (5) color change by concentrated NPs.
Representative applications of colorimetric detection and monitoring, including those described in this paper and others that have been developed and employed for the detection of a wide range of bacteria and toxins, are shown in Table 1. Their challenges, together with strategies to overcome the limitations, are summarized in Table 2. This review also describes an approach that can allow the development of colorimetric detection systems to facilitate their practical use.
2. Colorimetric Detection and Monitoring Strategies of Bacterial Contamination
Currently, the color development stage of colorimetric systems has five potential routes for signal visualization to determine the bacteria contamination level. The first route is based on external pH change-induced pH indicator reactions [19,20,21,22,23]. In this route, visual signal readouts can yield a color change in pH indicators via external pH changes. pH indicators include litmus, phenol red, bromothymol blue (BTB), and phenolphthalein. External pH changes rely on the catalytic activity of various enzymes added to the solution. These enzymes include urease [24], glucose oxidase (GOx) [25,26], esterase [27], penicillinase [28], and organophosphate paraoxon [29]. Urease, one of the most widely used enzymes, especially for the detection of bacterial contamination, hydrolyzes urea into ammonium and carbon dioxide, which increases the pH value [19,30]. Its hydrolytic activity also increases the ionic strength of the solution and, thus, can serve as a signal amplifier. Urease has been widely used in colorimetric bacterial detection based on its catalytic activity [19,20,21]. In this route, the type of pH indicator is also important, which determines the sensitivity of the system. The concentration of the indicator is another factor that determines the sensitivity and the linear range that can be quantified.
The second route relies on intracellular enzyme-catalyzed chromogenic reactions [31,32,33,34,35,36]. This route is based on the colorization of colorless or faintly colored chromogens by chemical reaction. Chromogens exhibit a variety of colors when their colorless indole molecule are substituted with halogen atoms. The degree and type of color change depend on the electron withdrawing capability of the halogen and the position of the substituted halogen on the indole ring. Depending on what kind of intracellular enzyme is used, there are two different strategies catalyzing chromogenic reactions to detect bacteria. One is to use the enzymes involved in distinctive metabolic reactions and the other is to use enzymes in the universal reactions of bacteria. In the first strategy, unique metabolic reaction-associated enzymes can not only be used as markers to detect and identify bacterial species, but also allow colorimetric bacterial detection by interacting with their chromogens. The second strategy is based on the enzymatic reaction of the metabolic pathway inherent in bacteria. This method can achieve broad-spectrum bacterial detection but may require additional processes for the identification of bacterial species. In this process, the bacteria in a sample can directly reduce the substrate, which changes the color of the substrate [31,32,33,34]. Alternatively, the presence of bacteria in a sample can act as an inhibitor of the chemogenic reaction, which leads to no color change [35,36]. This method can also monitor the antibiotic-resistant level owing to the inhibition of cell viability or metabolic reaction associated with the mode of action of antibiotics. Such a system takes advantage of the intrinsic catalytic reaction of intracellular enzymes and, thus, is easy to operate and does not require any additives except a substrate corresponding to the enzyme. Furthermore, most systems do not require any receptor to capture bacterial cells, which makes the process simpler and more cost-effective. The mixing between the substrate and bacterial cells is important because of the sensing output produced from the binding of bacterial cells to the chromogen. The degree of color change is proportional to the cell concentration. Nevertheless, the interference of physiological cellular functions by redox molecules should be further investigated.
The third route is based on enzyme-like NP-catalyzed substrate reactions [37,38,39,40,41,42,43,44]. Enzyme reactions have been widely used for the visual detection of bacteria; however, they have limited advantages due to their low stability and the high cost of enzymes. To overcome their limitations, colorimetric detection systems based on reactions using functional NPs have been developed [45,46]. An strategy of using NPs for colorimetric detection is to harness their enzyme-like catalytic activity. In comparison with natural enzymes, functional NPs have several advantages, including high stability against harsh environmental conditions (pH and temperature), proper catalytic activity, low cost for synthesis and mass production, and facile functionalization [45,47]. Enzyme-like NPs include peroxidase mimics, oxidase mimics, catalase mimics, and hydrolase mimics. In such a system, these nanomaterials act as enzymes catalyzing chromogenic reactions, allowing the visual detection of pathogenic bacteria with a fast response and simple operation even under harsh conditions.
The fourth route is based on NP aggregation reactions [48,49,50,51,52,53,54,55,56,57]. The color change is based on the size of NPs and their degree of aggregation. Among the various NPs, gold NPs (AuNPs) have been most widely used for the detection of biomolecules because of their biocompatibility and low cytotoxicity. AuNPs have also been widely used in colorimetric systems because they can change color depending on the interparticle distance through transition between dispersion and aggregation states when exposed to diverse stimuli, such as salts, ions, pH changes, chemicals, and target molecules [48,49,50,51,52,53,54,55,56,57]. Depending on the type of stimulus, several strategies can be adopted. One strategy involves the salt-induced aggregation of AuNPs [48,49,50]. High salt levels can neutralize the stabilizing electrostatic forces on AuNPs, causing the van der Waals forces to drive the conversion of dispersion into aggregation [58]. Another strategy involves the ion-induced aggregation of AuNPs [51,55,59]. For example, Mn2+ can interact with carboxyl groups on the AuNP surface and lead to the aggregation of AuNPs, resulting in a change in color from red to purple. A recent study reported that the addition of Mn2+ and Cu2+ facilitates the cysteine-induced aggregation of AuNPs [59]. The thiol group of cysteine strongly binds to the AuNP surface, forming a zwitterionic structure with carboxyl and ammonium groups [60]. The zwitterionic attractions of the oppositely charged groups and hydrogen bonding induce AuNP aggregation [61]. AuNP aggregation, in turn, reduces the interparticle distance between NPs and induces interparticle surface plasmon coupling, resulting in a change in color from red to blue [62]. Mn2+ and Cu2+ can form a stable chelate complex with the zwitterionic groups of cysteine molecules attached to AuNPs, thereby enhancing the colorimetric response of AuNPs. The use of functionalized ligands can also lead to AuNP aggregation in the presence of a target molecule. For example, bacteriophages targeting specific strains have been engineered to bind to both bacterial cells and AuNPs and used to detect diverse species in a colorimetric manner [63,64]. AuNP aggregation can be achieved by the direct binding of the analyte to AuNPs [55].
The fifth route is based on NP accumulation reactions [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79]. This strategy has been generally used in flat substrate-based systems, such as the lateral flow assay (LFA), µ-paper-based analytical device (µ-PAD), and paper-based dipstick system [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79]. These systems rely on the capillary action of fluids and colored NPs. The liquid sample containing the target molecules flows along the test device and passes through various zones of strips on which molecules that can interact with the analyte are attracted. The colorimetric readout is attributed to the conjugation of concentrated NPs with ligands targeting target molecules. The concentrated NPs act as colorimetric labels or color generation factors. Colored NPs used for colorimetric detection include AuNPs, silver NPs (AgNPs), and colloidal carbon [65,66,67,68,69,70,71,72,73,74,75,76,77,78,79]. Red AuNPs are one of the widely used colorimetric labels because of their bright color, high stability, and high biocompatibility [80]. NPs with nanozyme or peroxidase-like amplification activities are also used in this strategy, resulting in a signal amplification effect [69,70,71]. These systems based on a concentrated NP-mediated signal readout exhibit rapid responses, portability, single-step operations, cost-effectiveness, and convenience, enabling instant on-site detection and monitoring. High sensitivity, accurate quantification, and multiplexed detection remain challenges for wide application in the real world.
The aforementioned signal readout routes can be combined with diverse signal amplification methods to enhance the sensitivity of the system. Several signal amplification methods, such as hybridization chain reaction (HCR), enzymatic strand displacement amplification (SDA), loop-mediated isothermal amplification (LAMP), recombinase polymerase amplification (RPA), helicase-dependent amplification (HAD), and catalytic hairpin assembly (CHA), have been used to detect diverse bacterial cells [57,65,69,81,82,83,84]. Enzyme-driven signal amplification reactions, such as SDA and RPA, exhibit high sensitivity and specificity; however, they require relatively complicated operations, specific reaction conditions, and specific assay times depending on the catalytic ability of the enzyme. Enzyme-free isothermal amplification reactions, such as HCR, LAMP, and CHA, are low cost, have rapid responses, and require simple operations [65,83,84]. The background noise caused by non-specific products without the target should be considered. However, lower sensitivity than enzyme-driven methods remains a challenge.
3. Current Colorimetric Sensing Systems for the Detection of Bacteria and Toxins
In this section, we highlight the factors that affect color change for visual signal readout. The challenges and strategies to improve the performance of colorimetric sensing systems, including the related parameters (linear range, detection limit, assay time, etc.), are summarized in Table 1.
3.1. Color Change of pH Indicators via External pH Change
Signal readout can be obtained via the color change in a pH indicator by adding a catalytic compound that causes external pH change. Urease and a pH indicator have been widely used for the colorimetric detection of bacteria [19,20,21]. Urease is one of the most used catalytic compounds for the detection of bacterial contamination in food and the environment. To select an optimal pH indicator for yielding high sensitivity in the sensing system, three different pH indicators, phenol red, BTB, and bromocresol purple (BCP), were compared [19] (Figure 1A). After different concentrations of urea were applied, BTB exhibited the largest slope and was unaffected by urea in terms of color change. Under an optimal indicator concentration, Listeria monocytogenes was detected by forming magnetic nanoparticle (MNP)–monoclonal antibody (mAb)–bacterial cell–polyclonal antibody (pAb)–AuNP–urease sandwich complexes using two different NPs: MNPs for cell separation to reduce background noise and AuNPs for capturing urease via electrostatic adsorption [85,86]. The sandwich complexes containing urease could hydrolyze urea, and the increased pH of the sample could lead to a color change in BTB. The system had a detection limit of 100 CFU/mL with a linear range of 1.1 × 102 CFU/mL to 1.1 × 106 CFU/mL.
Considering the catalytic reaction of urease, the competitive binding ability of urease and bacterial cells to metal ions has been employed for the detection of bacteria [20,21,87]. It is well-known that urease can bind to other metals such as Ag+, Hg2+, Cu2+, and Zn2+, and metal-bound urease loses its hydrolyzing activity. Among these ions, silver ions can penetrate bacterial cell walls and exert high bactericidal activity by inactivating enzymes, inducing lipid peroxidation, and damaging cell membranes to kill bacteria. Ag ions also bind to urease, inhibiting urease activity by blocking its active site through sulfhydryl linkages that mediate a color change reaction in the pH indicator. Based on the binding ability of Ag ions to two substances, a colorimetric assay for the detection of Salmonella was developed (Figure 1B) [21]. In the absence of bacteria, Ag ions bind to urease, and the inhibited enzyme does not digest urea into ammonia, resulting in no pH change and no color change in the phenol red indicator. In the presence of bacteria, there is a competitive equilibrium between the enzyme and the bacteria for Ag ions. Owing to the Ag ions being sequestrated by the bacteria, the amount of free urease increases. Free urease hydrolyzes urea, and the produced ammonia leads to a higher pH, at which the phenol red indicator turns pink-red.
Similarly, the competitive binding ability of cationic AgNPs to two substances, bacterial cells and urease, has been applied to a colorimetric assay for the detection of Salmonella. Cationic AgNPs were fabricated by functionalizing them with polyethyleneimine, which renders the binding with the bacterial surface more attractive than that with urea because the bacterial cell surface is anionic due to an enriched negatively charged phosphate residue, and urease (pI = 5.0) is less negatively charged at a neutral pH [20]. The absence of bacteria in the sample makes AgNPs and urease bind to each other; thus, inactivated urease cannot hydrolyze urea, resulting in no pH change and a yellow color. The presence of bacteria can lead to the strong interaction of AgNPs and bacteria and can restore urease activity. Active urease produces ammonia with a color readout that allows bacterial quantification. This system is simple and cost-effective because it does not require tedious bioconjugation steps and additional receptors, and the limited identification of bacterial species can be overcome using species-specific receptors.
A recent study reported the development of pH-responsive NPs for the detection of bacterial contamination in food (Figure 1C) [22]. These pH-responsive NPs were fabricated using a phenolphthalein indicator (PP) and a thymolphthalein indicator (TP) in a self-assembly manner and functionalized by immobilizing the bacterial species-specific aptamer on each NP. The synthesized NPs were colorless at a neutral pH and red for PP-NPs and blue for TP-NPs at an alkaline pH. PP and TP also showed a rapid response to a change in the pH [88,89,90,91]. Another bacterial species-specific aptamer was modified with biotin and immobilized on a streptavidin-coated well plate. Two different aptamers captured bacterial cells present in the sample in a sandwich manner, indicating the co-existence of NPs in the sample. After NaOH was added, the increased pH caused the color change of the NPs. This process, combined with automatic equipment consisting of a pipetting head, a temperature controller, and an online control-transforming software, allowed multiplexing detection and shortened the time of the detection process to less than 1 h.
Another recent study reported the development of a paper-based colorimetric system for monitoring bacterial spoilage (Figure 1D) [23]. Bacterial spoilage changes the odor of food and increases the pH. This system relies on the pH change of food caused by volatile basic nitrogen generated from bacterial spoilage. Two different pH indicators, BCP and BTB, were coated on a piece of filter paper by soaking in a solution containing these dyes, resulting in the fabrication of a dual sensor. The dye-coated filter papers were placed onto meat and chicken fillets, and their color change was monitored for 8 days. Over time, the concentration of bacterial cells, pH value, and total basic nitrogen of the samples were increased, with the color of the paper changing from yellow to blue in the case of BTB and from yellow to purple in the case of BCP. The readout from the sensor was also analyzed using RGB software on a smartphone for the quantification of bacterial spoilage.
3.2. Color Change of Chemicals via Metabolic Activity of Intracellular Enzymes
Bacteria have distinctive metabolic reactions, producing substrates that are essential to the metabolism and survival of other organisms. In cellular respiration, electrons from carbon sources such as glucose move gradually through the electron transport chain toward oxygen, transitioning to lower energy states and releasing energy. The released energy is used to convert adenosine diphosphate into adenosine triphosphate, which can be used to power biological processes throughout the cell. Metabolic reactions in bacteria can trigger chromogenic reactions by directly reducing the substrate added to the solution, producing a visual readout. For example, non-O157 Escherichia coli has two metabolic pathways for cleaving 6-chloro-3-indoxyl-β-D-galactopyranoside (Sal-β-gal) and 5-bromo-4-chloro-3-indoxyl-β-D-glucuronic acid (X-β-gluc), producing chemicals that are purple and blue, respectively. In contrast, E. coli O157 only exhibits Sal-β-gal-degrading enzyme activity. Based on this discriminative feature, a system for detecting two different strains was developed with paper substrates [31,32]. The paper-based system consists of a multi-layered filter paper structure patterned with photoresists by ultraviolet light curing and functionalized with lysing, oxidizing, and chromogenic reagents in the different layers. By passing through successive layers of paper after injecting the sample, bacterial cells in the sample are lysed; the pH is adjusted for the optimal oxidation of the reactant in the oxidizing agent layer, and finally, the color is produced via the enzymatic reaction of the reactant in the chromogenic agent layer. The use of reagent-embedded paper allows the cost-effective and simple detection of bacterial species, even in resource-limited regions. This system could detect bacteria up to 10 CFU/mL with a linear range of 106 to 108 CFU/mL within 60 min for best visualization (Figure 2A; Table 1) [32]. With a similar system, four different bacterial species (E. coli, E. coli O157:H7, L. monocytogenes, and Vibrio vulnificus) were detected after expanding the number of chromogenic substrates to include X-β-gluc, 5-bromo-6-chloro-3-indoxyl-β-D-galactose, Aldol® 518 myoinositol-1-phosphate, and 5-bromo-4-chloro-3-indoxyl-β-D-glucose, each of which could react with β-glucuronidase, β-D-galactosidase, myoinositol mono-phosphates, and β-D-glucosidase, producing sky blue, burgundy, light pink, and strong blue colors, respectively [31]. This system also had a detection limit of 10 CFU/mL.
Oxidation–reduction reactions also occur during cellular respiration in living cells. Redox-active molecules can replace oxygen in respiratory enzymatic reactions and accept electrons for living cells. Intracellular oxidoreductase reduces redox-active molecules such as organic dyes, ferricyanide, dichlorophenolindophenol, and benzoquinone, resulting in color change or decolorization. The degree of color change in these molecules induced by intracellular enzymes can be used to assess cell metabolic activity and viability. These molecules also include 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) and 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, a monosodium salt (WST-8). For example, WST-8 is reduced by cellular dehydrogenase, generating an orange-yellow formazan product that is soluble in the culture medium. Through the reduction of molecules by intracellular enzymes, living or dead bacterial cells have been detected [33]. Dehydrogenase-based enzymatic reactions were observed to detect viable bacteria with a linear range of 2.6 × 102 to 1.16 × 109 CFU/ mL for E. coli and 9.75 × 102 to 6 × 109 CFU/mL for Staphylococcus aureus within 2 h. Although this system can easily detect bacteria in a broad range, it requires additional steps to identify the bacterial species.
Another redox-active molecule, benzoquinone, has been employed to develop a bacterial detection system (Figure 2B) [34]. Benzoquinone, which initially appeared as a yellowish-colored crystalline, was reduced to a white-colored hydroquinone by intracellular enzymes during cell respiration. Depending on the cell concentration, the color gradient could be monitored by the naked eye, and accurate quantification could be performed using RGB analysis software on a smartphone. The developed system had a linear range of 1.0 × 104 to 1.0 × 109 CFU/mL. It was also used to monitor the resistance level of E. coli cells against antibiotics with two different modes of action (trimethoprim and erythromycin). Trimethoprim exerts bacteriostatic activity and kills bacteria by preventing them from producing folic acid for purine and DNA synthesis. Erythromycin is a bacteriostatic antibiotic that inhibits protein synthesis by binding to 50S ribosomal subunits. Within 1 h of treatment with these two antibiotics, the colors of antibiotic-susceptible and low-antibiotic-resistant bacteria remained unchanged and faded, respectively. In contrast, the color of high-antibiotic-resistant bacteria became almost colorlessness.
Unlike chromogenic reactions triggered by cellular metabolic reactions, bacteria can inhibit extracellular chromogenic reactions, causing decolorization [35,36]. Accordingly, bacterial contamination has been detected based on the chromogenic reaction of H2O2 produced from GOx-mediated oxidation and glucose uptake by bacterial cells (Figure 2C) [35]. The substrate used was embedded with starch and iodide. After glucose was dropped on the starch–iodide doping paper in the absence of bacteria, GOx-catalyzed glucose oxidation produced H2O2, converting iodide to iodine. The produced iodine reacted with the starch, and the color of the paper changed to deep blue. When the sample was contaminated with bacteria, bacteria absorbed glucose as a carbon source for survival, inhibiting the subsequent chemical reaction and keeping the paper colorless. Using this system, E. coli and S. aureus could be detected at a concentration of 7.48 × 103 CFU/mL and 3.3 × 103 CFU/mL, respectively, within 20 min.
Another system based on the inhibition of chromogenic reactions by metabolic reactions has been developed using Cu2+ and o-phenylenediamine (OPD) [36]. Cu2+ can not only react with OPD, which produces oxidized OPD with a pale-yellow color, but can also bind to bacterial cells, which reduces Cu2+ to Cu+ through intracellular Cu2+ metabolic reaction-related enzymes [92,93]. The paper was prepared by dropping OPD solutions as a substrate. When Cu2+ was applied to OPD-embedded paper with no bacteria, the reaction between OPD and Cu2+ occurred without any inhibiting factors, changing the color of the paper to pale-yellow. However, the presence of bacteria in the sample impeded OPD oxidation by Cu2+, resulting in the OPD remaining colorless. Furthermore, the Cu2+-oxidized OPD is an orange-yellow fluorescent chemical; thus, its use could allow fluorescent and colorimetric dual-channel detection. In the dual-readout mode, this system had a detection limit of 44 CFU/mL with a linear range of 1 × 102 to 1 × 106 CFU/mL.
3.3. Color Change of Substrates via Enzyme-like Catalytic Reactions of NPs
Pathogenic bacteria can be detected based on the color change of chromogens. The change in the visible color is attributed to the catalytic reaction between enzymes and chromogenic substrates. 3,3′,5,5′-Tetramethylbenzidine (TMB) is one of the most widely used chromogens [94,95]. It is oxidized by peroxidase in the presence of H2O2 and undergoes color change from colorlessness to blue. Such a color change can occur due to enzymatic reactions involving horseradish peroxidase (HRP) and GOx. However, enzymes have some limitations, such as low stability under harsh conditions (pH and temperature), difficulty in labeling and functionalization, and high purification cost, especially in vitro. The use of enzyme mimics could be an alternative approach to overcome the limitations of natural enzymes [14,46]. Functional nanomaterials have been widely used in the colorimetric detection of bacterial contamination because of their unique properties. The catalytic activity of some nanomaterials is similar to that of enzymes, which triggers a catalytic reaction that induces a color change in the chromogen [38,40,96]. These enzyme-like materials include Fe3O4, metallic oxide NPs (Co3O4 and CeO2), noble metal NPs (AuNPs, Ag/PtNPs, and Au/PdNPs), and carbon-based nanomaterials (e.g., graphene oxide (GO)) [26,37,38,39,97,98,99,100,101]. A ZnFe2O4-modified reduced GO (rGO) nanostructure with HRP-like activity was used to detect S. typhimurium (Figure 3A) [37]. S. typhimurium was captured by two different aptamer-sandwich complexes, and the ZnFe2O4/rGO nanostructure could bind to the aptamer due to the specific affinity of GO for single-stranded DNA. The ZnFe2O4/rGO nanostructure was highly stable, even at pH 5.5 and 50 °C, demonstrating sensitive detection with a detection limit of 11 CFU/mL.
Peroxidase-like Cu2+-rGO was also used for the detection of S. typhimurium (Figure 3B) [38]. In this study, dsDNA amplified from bacterial cells acted as a competitor of Cu2+-rGO in the TMB interaction. If bacteria were present in the sample, dsDNA could be extracted from bacterial cells and amplified via polymerase chain reaction, and the dsDNA could bind to TMB. The competitive binding of TMB with dsDNA hindered the interaction of Cu2+-rGO and TMB, resulting in the solution color remaining the same. In a bacteria-free sample, Cu2+-rGO-based TMB oxidation occurred in H2O2, producing a blue color. This system had a detection limit of 0.51 CFU/mL with a linear range of 1.93 × 101 to 1.93 × 105 CFU/mL. Another peroxidase mimic, graphitic C3N4@Cu2O (g-C3N4@Cu2O), was used for the detection of S. typhimurium [39]. A S. typhimurium-specific aptamer was attached to C3N4@Cu2O, and the presence/absence of cells was monitored by detecting a color change. The g-C3N4@Cu2O nanostructure exhibits peroxidase-like activity and its functionalization with an aptamer lowers the peroxidase-like activity of the g-C3N4@Cu2O nanostructure. Therefore, the presence of bacteria in the sample strengthens the change in TMB color because the binding of the aptamer and cells induced the conformational change of the aptamer and weakened the interaction of the aptamer and C3N4@Cu2O. This system had a detection limit of 15 CFU/mL with a linear range of 1.5 × 101 to 1.5 × 105 CFU/mL. The paper-based system could detect bacterial cells within 6 min.
The bioconjugation of target-specific receptors can shield the surface of nanomaterials, reducing their catalytic activity. Based on this unique characteristic, a method of detecting S. aureus has been developed [40]. Bacterial cells were first captured by IgY-Fe3O4/Au nanocomposites and aptamer-functionalized AuNPs in a sandwich manner. Although Fe3O4 NPs are generally used for sample separation, they have limited active sites for conjugating bioreceptors directly and can aggregate, thereby losing their magnetic properties. To overcome this limitation, MNPs covered with an Au shell were used to enhance the modification yield and stability. AuNPs behave similarly to peroxidase; however, aptamer functionalization for capturing cells on the surface of the AuNPs causes them to lose their enzymatic activity. After cells were captured, the use of a magnet allowed the separation of cell-bound nanomaterial complexes from unbound aptamer-functionalized AuNPs. The H2O2 added to unbound AuNPs dissociated the aptamer and restored the catalytic activity of AuNPs, which catalyzed TMB, producing a yellow color. This system had a detection limit of 10 CFU/mL with a linear range of 101 to 106 CFU/mL.
The detectable sensitivity of peroxidase activity-based TMB signals is limited because H2O2 can produce only one colored TMB molecule with the aid of peroxidase. The selection and optimization of enzyme–chromogenic substrates are important for developing advanced colorimetric assays with high sensitivity. For example, the addition of Fe2+ and Cu2+ to reactions with H2O2 and TMB can produce two colored TMB molecules by forming Fe3+/Cu2+ and hydroxyl radicals, both of which directly oxidize TMB [102,103,104,105]. Through this signal amplification strategy, brevetoxin B (BTB), a potent neurotoxin naturally produced from Karenia brevis, was detected [41]. BTB was conjugated with magnetic beads and incubated with Ab-AuNP-GOx composites. If the sample contained BTB, a Mb-BTB-Ab-AuNP-GOx complex was formed and isolated after the magnetic separation process. Upon exposure to glucose and Fe2+, GOx oxidized glucose to gluconic acid and H2O2. H2O2 oxidized Fe2+ to Fe3+ and hydroxyl radicals, both of which changed the color of TMB to yellow. If BTB was absent in the sample, the TMB remained colorless TMB. This system could detect up to 0.076 ng/kg of BTB.
Based on the chemical conversion of Fe2+ to Fe3+ by GOx-catalyzed H2O2, K3[Fe(CN)6] was employed as a color change indicator for visual readout [42]. K3[Fe(CN)6] is blue in the presence of a Fe2+-containing solution; however, the oxidization of Fe2+ to Fe3+ inhibits the production of blue color, making the solution colorless. This feature was exploited to determine the ochratoxin (OTA) contamination level [42]. OTA attached to MB was bound with OTA antibody-functionalized AuNPs, in which GOx was modified, forming the sensing platform (MB-OTA-OTA Ab-AuNP-GOx). Upon the addition of a sample, magnet-based separation was accomplished, and glucose, FeCl2, and K3[Fe(CN)6] were sequentially added to the precipitate. This assay is based on the competition of OTA attached to a sensing platform and OTA in a sample for binding with an anti-OTA antibody; thus, if the sample contained OTA, OTA antibody-AuNP-GOx in the sensing platform could bind with the OTA in the sample instead of that in the sensing platform. The amount of OTA antibody-AuNP-GOx in the sensing platform was decreased compared with that in the initial stage, which also led to a decreased amount of GOx, resulting in GOx-catalyzed H2O2 not being produced and, thus, K3[Fe(CN)6] not changing color. When OTA was absent from the sample, there was no competition, and the amount of OTA antibody-AuNP-GOx in the sensing platform remained the same, resulting in the production of GOx-catalyzed H2O2 and, thus, the color of K3[Fe(CN)6] changing from blue to colorless. Through this strategy, OTA could be detected up to 8.3 pg/mL with a linear range of 0.01 to 10 ng/mL.
Au nanoclusters (AuNCs) can also catalyze the oxidation of TMB by H2O2 and have been used for the detection of S. typhimurium (Figure 3C) [43]. AuNCs were functionalized as bacteria-specific aptamers and added to a sample containing TMB and H2O2. If the sample contained bacterial cells, bacteria could bind with not only aptamer@AuNCs but also TMB. These interactions might be possible due to the shorter distance between aptamer@AuNCs and the TMB substrate, resulting in enhanced peroxidase-like activity toward TMB. The enhanced activity may be related to the sensitivity and specificity of the system, which had a detection limit of 1 CFU/mL with a linear range of 101 to 106 CFU/mL.
Most peroxidase-like nanostructures are developed with a focus on catalytic activity in the presence of H2O2 as an oxidant. H2O2 is toxic to target analytes or cells and is unstable; thus, strict time control may be necessary. A MnO2-doped nanostructure was developed for the H2O2-free oxidization reaction of TMB into a blue-colored product [44,106]. MnO2-doped NPs have intrinsic peroxidase- and oxidase-like activities owing to the charge transfer complex of the iron oxide NPs, which could mediate the one-electron oxidation of TMB. MnO2-doped NPs could bind with bacteria via van der Waals interactions between the residual amino acids of the NPs and the anionic cell surface of the bacteria. These interactions could lead to the blocking of the active catalytic sites of NPs, causing them to lose their catalytic activity. Two different bacteria, S. aureus and Vibrio parahaemolyticus, were detected using MnO2-doped Fe3O4 NPs [44]. These NPs acted as multifunctional NPs, facilitating the recognition, absorption, and separation of bacterial cells as well as enzyme-like activity. This system could detect bacterial cells to 102 CFU/mL with a linear range of 101 to 106 CFU/mL.
3.4. Color Change via NP Aggregation-Based Reaction
Signal readout can be observed through a change in color resulting from the aggregation of NPs, such as AgNPs and AuNPs. The color change is attributed to the reduction of interparticle distance, which induces surface plasmon coupling. Au has been widely used for the colorimetric detection of bacteria through the aggregation of NPs induced by diverse stimuli. Among various NP aggregation factors, salt-indued NP aggregation was used to detect bacterial cells in drinking water [48]. AuNPs were initially functionalized with 4-mercaptophenylboronic acid (4-MPBA), which binds to glycoprotein on the surface of bacterial cells through reversible esterification [107]. In the absence of bacteria, NaCl treatment can change the color of AuNPs from red to purple as a result of aggregation. When the sample contains E. coli cells, surface colloidal AuNPs are covered with bacteria by covalent bonding between 4-MPBA and saccharides; this inhibits the aggregation of AuNPs and maintains their intrinsic color. As 4-MPBA has an affinity towards saccharides and glycosylated biomolecules, it can bind to both peptidoglycans on the gram-positive bacterial cell surface and lipopolysaccharides on the gram-negative bacterial cell surface, enabling broad-spectrum bacterial detection in practical applications.
The detection of specific strains can be achieved using species-specific receptors, such as aptamers, and attaching them to AuNPs [49]. As ssDNA aptamers have a flexible phosphate backbone, the bases can easily access AuNPs and get adsorbed onto the surface of negatively charged AuNPs through van der Waals interactions. The adsorbed aptamers can act as stabilizers of AuNPs to disperse uniformly without any aggregation in high-salt solutions, thereby maintaining their red color. When aptamers that bind to bacterial cells rather than NPs are used as receptors, the presence of bacteria in samples causes the detachment of aptamers from AuNPs, resulting in the aggregation of AuNPs and the change in color from red to purple. Using this method, as low as 80 CFU/mL of S. flexneri could be detected within 20 min, with the linear range being 1 × 102 to 1 × 106 CFU/mL [49].
Mn2+, which is another NP aggregation factor, can also change the interparticle distance of AuNPs and aggregate them, changing the color of the colloidal solution (Figure 4A) [51]. For the detection of V. parahaemolyticus, species-specific antibodies were attached to two different particles in a previous study: (1) MnO2 NPs for inducing AuNP aggregation and (2) MNPs for the easy separation of target analytes [51]. After V. parahaemolyticus was captured in a sandwich-type reaction of these particles, the added ascorbic acid reduced MnO2 NPs, producing MnO2 ions. The generated MnO2 ions could interact with carboxyl groups on the AuNP surface, inducing AuNP aggregation. As one bacterial cell can bind to many MnO2 NPs, the subsequent ascorbic acid-induced reduction of MnO2 NPs could facilitate Mn2+-induced AuNP aggregation, enhancing the sensitivity of the assay through signal amplification. Using this system, as low as 10 CFU/mL of V. parahaemolyticus could be detected, with the linear range being 1 × 101 to 1 × 106 CFU/mL.
A diverse pathogen-detecting system was developed using bacteriophages (Figure 4B) [52]. The bacteriophage acted as a ligand for binding to specific species and an NP aggregation factor. The ligand for binding to specific species was fabricated using M13 as a phage scaffold. The M13 phage is a non-lytic filamentous phage. The M13 scaffold was functionalized by displaying receptor-binding proteins (RBPs) fused to the minor coat protein pIII. Ligand specificity was achieved by engineering to displace RBPs from the M13 phage targeting E. coli (F+) into different RBPs from the following foreign phages targeting specific species: the CTXφ phage targeting V. cholerae, Pf1 phage targeting Pseudomonas aeruginosa, φLf and φXv phages targeting Xanthomonas campestris, and Ιφ1 phage targeting E. coli (I+). This was possible because each RBP could interact with a specific host and needed machinery for downstream infection and propagation, which was non-compatible with other hosts. The capsids of the scaffold were also chemically modified with thiol groups to bind to AuNPs, providing a signal amplification effect. When such an engineered M13 scaffold was incubated with bacterial cells present in the sample, it could capture cells and form cell–phage complexes. After centrifugation, the cell–phage complexes were separated and the added AuNPs were attached to the capsids of the M13 scaffold, inducing AuNP aggregation and a change in color to purple, as visible to the naked eye. This system had a detection limit of 102 CFU/mL within 1 h. Using this engineered phage scaffold, the antibiotic resistance and susceptibility of P. aeruginosa could be assessed [53]. A Pf1 protein-anchored thiolated phage was engineered for P. aeruginosa. When the sample contained antibiotic-resistant P. aeruginosa, live cells bound to the engineered phage and formed cell–phage complexes. Centrifugation separated the complexes, and the incubation of AuNPs induced a color change in the solution. Antibiotic-susceptible cells are dead or have low functionality, resulting in no interaction with or low affinity to the engineered phage. After centrifugation, no or few phages remained in the solution, resulting in no or slight color change.
Concanavalin A (ConA) lectin, which is another driving factor for AuNP aggregation, was also used to detect E. coli (Figure 4C) [54]. ConA is a carbohydrate-binding protein that specifically binds to glycans with α-mannose-containing cores, e.g., many N-glycans, and has a strong affinity to the glycosyl protein on the bacterial surface. ConA has a pH-responsive tetramer–dimer transition ability; it forms tetramers at pH levels higher than 6.0 and dissociates to dimers or monomers under pH 5.0. To detect E. coli, antibody-functionalized MNPs were used. Bacteria in the sample could be captured and separated by MNPs. The added ConA sequentially bound to the bacterial cell surface. Low pH (pH 5.0) caused the ConA tetramers to dissociate to dimers or monomers. After removing bacteria-captured MNPs using a magnet, the dissociated dimers were transformed into tetramers with an increase in pH. The recovered ConA tetramers could interact with dextran molecules coated on AuNPs, leading to AuNP aggregation and providing a colorimetric readout. The use of ConA resulted in a signal amplification effect because of the high abundance of ConA-binding sites in bacteria. The proposed system exhibited a detection limit of 41 CFU/mL within 95 min.
Direct binding of target molecules to functionalized AuNPs can aggregate AuNPs [55]. Using this approach, the fumonisin B1 (FB1) detection method was developed because the hydroxyl groups of FB1 bind to cysteamine of AuNPs through multiple hydrogen bonds and aggregate them [55]. FB1 is a type of mycotoxin secreted from Fusarium species, which are mainly found in corns [108,109,110]. To detect FB1, cysteamine-functionalized AuNPs (Cys-AuNPs) were used for stabilizing NPs through strong electrostatic repulsion between particles and for facilitating interaction with FB1. The large size of FB1 causes steric hindrance; therefore, FB1 was hydrolyzed to form a structure with a short chain, which has a high affinity towards cysteamine-attached AuNPs. On hydrolyzing FB1 with KOH, it bound to dispersed Cys-AuNPs, forming aggregated Cys-AuNPs and leading to a change in color from red to purple.
The signal readout induced by AuNP aggregation can be amplified by combining diverse methods [56,57]. For example, the enzyme-driven DNA walker system was combined with the AuNP-based colorimetric method for the detection of bacterial cells (Figure 4D) [56]. The DNA walker system consisted of AuNPs functionalized with two different thiolated ssDNAs (probe 1 and probe 2, dual walkers) and DNA polymers. DNA polymers contained bacterial species-specific aptamers and walking strands (DNA 1 and DNA 2) with a sequence complementary to probe 1 and probe 2. If target bacterial cells were present in the sample, the aptamers preferred to bind to bacterial cells, thereby releasing them from the DNA polymers. Walking strands were also dispersed freely in the solution and bound to probe 1 and probe 2 on AuNPs, forming duplexes. Upon exposure to exonuclease III that could specifically digest one strand of duplex DNA from the 3′-hydroxyl termini, only probe 1 and probe 2 were removed and the surviving walking strands bound to another probe 1 and probe 2, resulting in multiple cycling reactions of hybridization and digestion. Probe DNA-removed AuNPs were eventually aggregated, and their color changed from red to purple. The color change increased with an increase in the number of walkers on AuNPs and walking strands. Using this system, as low as 1 CFU/mL of S. aureus could be detected within 15 min.
CHA, another signal amplification method, was used for the detection of S. typhimurium (Figure 4E) [57]. The CHA method relies on self-assembly and disassembly reactions between DNAs with hairpin structures. This method is an enzyme-free isothermal amplification method with high catalytic efficiency, low background signals, and simple operations in comparison with the enzyme-driven signal amplification method, which requires complex operations, specific reaction conditions, and specific assay times depending on the catalytic abilities of the enzymes [83,84]. To amplify the signal readout, three different DNAs (Y1, Y2, and Y3) were used; they formed a hairpin structure because of their low stability in terms of kinetics [57]. If S. aureus was absent in the sample, these DNAs were absorbed on the surface of AuNPs, preventing AuNP aggregation. If the sample contained S. aureus, the bacterial cells were captured by two different aptamers, namely a biotinylated aptamer (Bapt) attached to streptavidin-coated MNPs and a free aptamer (Tapt), in a sandwich manner, forming aptamer-cell-aptamer-MNP complexes. Upon heating, Tapt was released from the complexes and joined the CHA reaction by sequentially binding to three different DNAs absorbed on AuNPs, eventually forming a Y-shaped structure. Thus, AuNPs were aggregated, inducing a change in color from red to purple.
3.5. Color Change via NP Accumulation-Based Reaction
The concentrated NPs themselves act as colorimetric labels and generated color in the test zone. The representative colored NPs widely used in this approach are colloidal AuNPs. In previous studies, AuNPs were generally applied to paper-based flow assay strips [74,75,76,111]. The concentrated NPs acted as colorimetric labels and generated color in the test zone. For example, to detect Brucella species, an LFA strip was fabricated in laminates consisting of a nitrocellulose (NC) membrane, conjugation pad, sample pad, and absorption pad. One anti-Brucella antibody was deposited on the control and test lines of the strip, and another anti-Brucella antibody conjugated with AuNP (Ab-AuNP) was deposited on the conjugate pad [74]. When a sample containing the target bacteria was deposited on the sample pad, the sample bacterial suspension bound to Ab-AuNPs during flow through the strip. Following this, the bacteria-antibody-AuNP complexes were captured by antibodies on the control and test lines in a sandwich manner, resulting in the accumulation of AuNPs on these lines and the appearance of intense red lines. The AuNP-based dual-antibody sandwich assay was modified in a vertical flow format. Unlike LFA, this format was designed to enable the fluid to flow vertically and pass through the pads [111]. This format consisted of two parts: the upper cover body contained staphylococcal protein A (SPA)-AuNPs and the detection cassette contained an IgG-coated NC membrane and absorbent paper. To detect Brucella antibodies, the sample was deposited on the detection cassette. Following this, the cover body was placed on the cassette. Thus, the analyte was first fixed in the test zone and then bound to SPA-AuNPs, thereby minimizing false negatives. When Brucella antibodies were present in the sample, SPA-AuNPs bound to the anti-Brucella antibodies and IgGs, generating a color within 3 min. Similar to AuNP-based LFIA, colloidal AgNPs were used as colored NPs for the detection of staphylococcal enterotoxin B (SEB) in LFIA (Figure 5A) [65]. Using this strip, as low as 0.5 ppm of SEB was visually detected within 15 min, with the linear range being 0 to 2 ppm. Colloidal carbon was used for the development of LIFA strips for detecting Mycoplasma bovis [79]. Colloidal carbon has some advantages, such as simple conjugation and stability. Importantly, its bright-colored readout (black line on the white strip) leads to a high signal–background contrast, enhancing sensitivity [76,79]. In addition, the sensitivity could be enhanced by combining Au or Ag growth-mediated signal amplification methods, in which the in situ growth of NPs increased their size, thereby strengthening the signal readout [66,70]. NH2OH·HCl was used as a signal enhancer; in this method, AuNPs acted as catalysts for the reduction reaction between HAuCl4 and NH2OH·HCl, generating more AuNPs. The generated AuNPs were deposited on the test zone of the strip or covered the surface of the initially added AuNPs to form larger-sized AuNPs, providing a signal amplification effect. This signal amplification strategy combined with LFA exhibited 100-fold-enhanced sensitivity and could detect 104 CFU/mL of S. enteritidis.
Concentrated NPs can also act as color generation factors because of these catalytic activities. NP-mediated chemical reactions can amplify visual signals [70,71,72]. NPs can interact with the substrate as signal amplifiers. Consequently, the substrate is converted into colorful chemicals by the catalytic activity of NPs. For example, a recent study used a combination of two methods for cascade signal amplification: the first method involved the use of NH2OH·HCl and hydroquinone for in situ AuNP growth and the second method involved the use of nanozyme-mediated catalytic deposition (Figure 5B) [70]. Similar to the use of NH2OH·HCl, the use of hydroquinone increased the size of the initially added AuNPs on the strip. The enlarged AuNPs had high catalytic activity and could oxidize the chromogen 3-amino-9-ethylcarbazole, resulting in the deposition of a colored product on the surface of AuNPs. This strip had a detection limit of 1.25 × 101 CFU/mL for E. coli O157:H7. Pd-Pt NPs were also used for signal amplification to detect E. coli O157:H7 [71]. Pd-Pt NPs with peroxidase-like activity were conjugated with anti-E. coli O157:H7 antibodies and deposited on the conjugation pad of the LFIA strip. After injecting a sample containing bacterial cells on the strip, TMB was added to another antibody-deposited test zone. The concentrated Pd-Pt NPs at the test zone could oxidize TMB in the presence of H2O2, producing a colored product and visual signal. Similarly, Pt-Au NPs with peroxidase-like activity were used to amplify the signal readout and to detect anti-E. coli O157:H7, exhibiting a detection limit of 1 × 102 CFU/mL within 1 min. The catalytic activity of NPs resulted in a rapid response depending on the activity and enhanced the sensitivity of the assay. This system would, however, require an additional process, such as the addition of substrates and enzymatic components.
4. Conclusions
This review summarizes the recent advances in colorimetric sensing systems for the detection of bacterial contamination in food, water, and environmental sources. Emphasis was placed on the different factors affecting the signal readout and color changes of substrates. This review confirms that colorimetric sensing systems are a rapidly developing technology with advantages such as simple detection and operation, low cost, and ability to operate without an instrument or detector.
Despite these advantages, there is still room to improve the sensing performance of systems, and to achieve enhanced capability (Table 2). Ongoing efforts have been made in the development of methods with enhanced sensing performance. Sensitivity can be enhanced by using a dual-readout system and chemicals with various sensing properties [34,36]. The use of optimal chemicals and concentrations can improve the detection limit of systems [19]. A sensitive system could also be obtained using a combination of signal amplification methods, such as HCR, RPA, HAD, RCA, and CHA [56,57,65,69,81,82,83,84]. Moreover, the use of a 3D paper structure embedded with a chemical reaction-related reagent can make systems simple to use [31,32]. An automation device can be combined with sensing systems to allow the detection of multiple bacteria in a simple manner [22]. Recently, much effort has also been given to the fabrication of enzyme-like functional NPs and their application in the detection of bacterial contamination [45]. Unlike traditional NP aggregation-based methods, which are mostly used for signal visualization, the chemical reaction-catalyzing NP-based strategy is minimally affected by the interference of the complex components of actual samples, providing relatively correct signals [45]. This sensing system can quantitatively detect the levels of bacteria and toxins in a broad range using digital cameras, smartphones, or mobile devices to obtain the color intensity and conduct data processing [22,23,34,36,112,113].
With ongoing advances, we believe that colorimetric sensing systems will be considered a promising option for monitoring and detecting bacterial contamination in food, water, and environmental sources during production or manufacturing processes.
Author Contributions
Conceptualization, D.-M.K. and S.-M.Y.; writing, D.-M.K. and S.-M.Y.; supervision, S.-M.Y.; funding acquisition, S.-M.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by an NRF grant, which was funded by the Ministry of Science and ICT (NRF-2022R1A2C2004292).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created in this study. Data sharing is thus not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Chen, J.; Andler, S.M.; Goddard, J.M.; Nugen, S.R.; Rotello, V.M. Integrating recognition elements with nanomaterials for bacteria sensing. Chem. Soc. Rev. 2016, 46, 1272–1283. [Google Scholar] [CrossRef] [PubMed]
- Florentin, A.; Lizon, J.; Asensio, E.; Forin, J.; Rivier, A. Water and surface microbiologic quality of point-of-use water filters: A comparative study. Am. J. Infect. Control 2016, 44, 1061–1062. [Google Scholar] [CrossRef]
- Lemarchand, K.; Lebaron, P. Occurrence of Salmonella spp. and Cryptosporidium spp. in a French coastal watershed: Relationship with fecal indicators. FEMS Microbiol. Lett. 2003, 218, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrell, M.L.; Joyce, A.; Duane, S.; Fitzhenry, K.; Hooban, B.; Burke, L.P.; Morris, D. Evaluating the potential for exposure to organisms of public health concern in naturally occurring bathing waters in Europe: A scoping review. Water Res. 2021, 206, 117711. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control. Outbreak of E. coli Infections Linked to Romaine Lettuce, E. coli Infections, E. coli, CDC. November 2019. Available online: https://www.cdc.gov/ecoli/2019/o157h7–11-19/ (accessed on 1 June 2022).
- Nguendo-Yongsi, H.B. Microbiological evaluation of drinking water in a sub-saharan urban community (Yaunde). Am. J. Biochem. Mol. Biol. 2011, 1, 61–81. [Google Scholar]
- Delaire, C.; Peletz, R.; Kumpel, E.; Kisiangani, J.; Bain, R.; Khus, R. How much will it cost to monitor microbial drinking water quality in sub-Saharan Africa? Environ. Sci. Technol. 2017, 51, 5869–5878. [Google Scholar] [CrossRef] [Green Version]
- Schirone, M.; Visciano, P.; Tofalo, R.; Suzzi, G. Editorial: Foodborne pathogens: Hygiene and safety. Front. Microbiol. 2019, 10, 1974. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.; Alzua, M.L.; Osbert, N.; Pickering, A. Community-level sanitation coverage more strongly associated with child growth and household drinking water quality than access to a private toilet in rural Mali. Environ. Sci. Technol. 2017, 51, 7219–7227. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Wang, X.; Yang, Q.; Wu, W. Quantum dot: Lightning invisible foodborne pathogens. Trends Food Sci. Technol. 2021, 110, 1–12. [Google Scholar] [CrossRef]
- Li, T.; Ou, G.; Chen, X.; Li, Z.; Hu, R.; Li, Y.; Yang, Y.; Liu, M. Naked-eye based point-of-care detection of E. coli O157: H7 by a signal-amplified microfluidic aptasensor. Anal. Chim. Acta. 2020, 1130, 20–28. [Google Scholar] [CrossRef]
- Castle, L.M.; Schuh, D.A.; Reynolds, E.E.; Furst, A.L. Electrochemical sensors to detect bacterial foodborne pathogens. ACS Sens. 2021, 6, 1717–1730. [Google Scholar] [CrossRef] [PubMed]
- Luo, K.; Kim, H.-Y.; Oh, M.-H.; Kim, Y.-R. Paper-based lateral flow strip assay for the detection of foodborne pathogens: Principles, applications, technological challenges and opportunities. Crit. Rev. Food Sci. Nutr. 2020, 60, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Verma, M.S.; Rogowski, J.L.; Jones, L.; Gu, F.X. Colorimetric biosensing of pathogens using gold nanoparticles. Biotechnol. Adv. 2015, 33 Pt 1, 666–680. [Google Scholar] [CrossRef]
- Choi, Y.; Hwang, J.H.; Lee, S.Y. Recent trends in nanomaterials-based colorimetric detection of pathogenic bacteria and viruses. Small Methods 2018, 2, 1700351. [Google Scholar] [CrossRef]
- Filik, H.; Avan, A.A. Nanotechnology-based colorimetric approaches for pathogenic virus sensing: A review. Curr. Med. Chem. 2022, 29, 2691–2718. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, Q.H.; Kim, M.I. Nanomaterial-mediated paper-based biosensors for colorimetric pathogen detection. Trends Analyt. Chem. 2020, 132, 116038. [Google Scholar] [CrossRef]
- Liu, B.; Zhuang, J.; Wei, G. Recent advances in the design of colorimetric sensors for environmental monitoring. Environ. Sci. Nano 2020, 7, 2195–2213. [Google Scholar] [CrossRef]
- Chen, Q.; Huang, F.; Cai, G.; Wang, M.; Lin, J. An optical biosensor using immunomagnetic separation, urease catalysis and pH indication for rapid and sensitive detection of Listeria monocytogenes. Sens. Actuators B-Chem. 2018, 258, 447–453. [Google Scholar] [CrossRef]
- Singh, P.; Kakkar, S.; Bharti; Kumar, R.; Bhalla, V. Rapid and sensitive colorimetric detection of pathogens based on silver-urease interactions. Chem. Commun. 2019, 55, 4765–4768. [Google Scholar] [CrossRef]
- Kumar, V.; Chopra, A.; Bisht, B.; Bhalla, V. Colorimetric and electrochemical detection of pathogens in water using silver ions as a unique probe. Sci. Rep. 2020, 10, 11986. [Google Scholar] [CrossRef]
- Yan, C.; Sun, Y.; Yao, M.; Jin, X.; Yang, Q.; Wu, W. pH-responsive nanoparticles and automated detection apparatus for dual detection of pathogenic bacteria. Sens. Acturator B-Chem. 2022, 354, 131117. [Google Scholar] [CrossRef]
- Abo Dena, A.S.; Khalid, S.A.; Ghanem, A.F.; Shehata, A.I.; El-Sherbiny, I.M. User-friendly lab-on-paper optical sensor for the rapid detection of bacterial spoilage in packaged meat products. RSC Adv. 2021, 11, 35165–35173. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ran, X.; Lin, Y.; Ren, J.; Qu, X. Enzyme-regulated the changes of pH values for assembling a colorimetric and multistage interconnection logic network with multiple readouts. Anal. Chim. Acta 2015, 870, 92–98. [Google Scholar] [CrossRef]
- Jankowska, D.A.; Bannwarth, M.B.; Schulenburg, C.; Faccio, G.; Maniura-Weber, K.; Rossi, R.M.; Scherer, L.; Richter, M.; Boesel, L.F. Simultaneous detection of pH value and glucose concentrations for wound monitoring applications. Biosens. Bioelectron. 2017, 87, 312–319. [Google Scholar] [CrossRef]
- Song, Y.; Qu, K.; Zhao, C.; Ren, J.; Qu, X. Graphene oxide: Intrinsic peroxidase catalytic activity and its application to glucose detection. Adv. Mater. 2010, 22, 2206–2210. [Google Scholar] [CrossRef]
- Contin, A.; Frasca, S.; Vivekananthan, J.; Leimkühler, S.; Wollenberger, U.; Plumeré, N.; Schuhmann, W. A pH responsive redox hydrogel for electrochemical detection of redox silent biocatalytic processes. Control of Hydrogel Solvation. Electroanalysis 2015, 27, 938–944. [Google Scholar] [CrossRef]
- Bidmanova, S.; Hlavacek, A.; Damborsky, J.; Prokop, Z. Conjugation of 5(6)-carboxyfluorescein and 5(6)-carboxynaphthofluorescein with bovine serum albumin and their immobilization for optical pH sensing. Sens. Actuators B-Chem. 2012, 161, 93–99. [Google Scholar] [CrossRef]
- Chen, C.H.; Yang, K.L. A liquid crystal biosensor for detecting organophosphates through the localized pH changes induced by their hydrolytic products. Sens. Actuators B-Chem. 2013, 181, 368–374. [Google Scholar] [CrossRef]
- Lei, C.; Dai, H.; Fu, Y.; Ying, Y.; Li, Y. Colorimetric sensor array for thiols discrimination based on urease-metal ion pairs. Anal. Chem. 2016, 88, 8542–8547. [Google Scholar] [CrossRef]
- Kim, H.J.; Kwon, C.; Lee, B.S.; Noh, H. One-step sensing of foodborne pathogenic bacteria using a 3D paper-based device. Analyst 2019, 144, 2248–2255. [Google Scholar] [CrossRef]
- Kim, H.J.; Kwon, C.; Noh, H. Paper-based diagnostic system facilitating Escherichia coli assessments by duplex coloration. ACS Sens. 2019, 4, 2435–2441. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhong, Y.; Wang, D.; Lu, Z. A simple colorimetric method for viable bacteria detection based on cell counting Kit-8. Anal. Methods 2021, 13, 5211–5215. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Warden, A.R.; Huang, J.; Wang, W.; Ding, X. Colorimetric and electrochemical detection of Escherichia coli and antibiotic resistance based on a p-benzoquinone-mediated bioassay. Anal. Chem. 2019, 91, 7524–7530. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Huang, J.; Li, Y.; Lv, J.; Ding, X. A simple and rapid colorimetric bacteria detection method based on bacterial inhibition of glucose oxidase-catalyzed reaction. Talanta 2019, 197, 304–309. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gao, X.; Wang, S.; Liu, Y. A smartphone-integrated paper sensing system for fluorescent and colorimetric dual-channel detection of foodborne pathogenic bacteria. Anal. Bioanal. Chem. 2020, 412, 611–620. [Google Scholar] [CrossRef]
- Wu, S.; Duan, N.; Qiu, Y.; Li, J.; Wang, Z. Colorimetric aptasensor for the detection of Salmonella enterica serovar typhimurium using ZnFe2O4-reduced graphene oxide nanostructures as an effective peroxidase mimetics. Int. J. Food Microbiol. 2017, 261, 42–48. [Google Scholar] [CrossRef]
- Wang, L.; Liao, T.; Zhou, H.; Huang, Y.V.; Chen, P.; Yang, X.; Chen, X. Colorimetric method for Salmonella spp. detection based on peroxidase-like activity of Cu(II)-rGO nanoparticles and PCR. Anal. Biochem. 2021, 615, 114068. [Google Scholar] [CrossRef]
- Tarokh, A.; Pebdeni, A.B.; Othman, H.O.; Salehnia, F.; Hosseini, M. Sensitive colorimetric aptasensor based on g-C3N4@Cu2O composites for detection of Salmonella typhimurium in food and water. Mikrochim. Acta. 2021, 188, 87. [Google Scholar] [CrossRef]
- Yao, S.; Li, J.; Pang, B.; Wang, X.; Shi, Y.; Song, X.; Xu, K.; Wang, J.; Zhao, C. Colorimetric immunoassay for rapid detection of Staphylococcus aureus based on etching-enhanced peroxidase-like catalytic activity of gold nanoparticles. Mikrochim. Acta. 2020, 187, 504. [Google Scholar] [CrossRef]
- Lai, W.; Wei, Q.; Zhuang, J.; Lu, M.; Tang, D. Fenton reaction-based colorimetric immunoassay for sensitive detection of brevetoxin B. Biosens. Bioelectron. 2016, 80, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Lai, W.; Guo, J.; Wu, Q.; Chen, Y.; Cai, Q.; Wu, L.; Wang, S.; Song, J.; Tang, D. A novel colorimetric immunoassay based on enzyme-regulated instant generation of Turnbull’s blue for the sensitive determination of ochratoxin A. Analyst 2020, 145, 2420–2424. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gao, R.; Jia, L. Enhancement of the peroxidase-like activity of aptamers modified gold nanoclusters by bacteria for colorimetric detection of Salmonella typhimurium. Talanta 2021, 221, 121476. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, C.; Zhao, W.; Zhang, H.; Yao, S.; Shi, Y.; Li, J.; Wang, J. Multi-functional MnO2-doped Fe3O4 nanoparticles as an artificial enzyme for the colorimetric detection of bacteria. Anal. Bioanal. Chem. 2020, 412, 3135–3140. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Wang, X.; Wang, Q.; Lou, Z.; Li, S.; Zhu, Y.; Qin, L.; Wei, H. Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes (II). Chem. Soc. Rev. 2019, 48, 1004–1076. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Bhardwaj, S.K.; Bhatt, D.; Lim, D.K.; Kim, K.H.; Deep, A. Optical detection of waterborne pathogens using nanomaterials. TrAC Trends Anal. Chem. 2019, 113, 280–300. [Google Scholar] [CrossRef]
- Wei, H.; Wang, E. Nanomaterials with enzyme-like characteristics (nanozymes): Next-generation artificial enzymes. Chem. Soc. Rev. 2013, 42, 6060–6093. [Google Scholar] [CrossRef]
- Huang, J.; Sun, J.; Warden, A.R.; Ding, X. Colorimetric and photographic detection of bacteria in drinking water by using 4-mercaptophenylboronic acid functionalized AuNPs. Food Control 2020, 108, 106885. [Google Scholar] [CrossRef]
- Feng, J.; Shen, Q.; Wu, J.; Dai, Z.; Wang, Y. Naked-eyes detection of Shigella flexneri in food samples based on a novel gold nanoparticle-based colorimetric aptasensor. Food Control 2019, 98, 333–341. [Google Scholar] [CrossRef]
- Jalalian, S.H.; Lavaee, P.; Ramezani, M.; Danesh, N.M.; Alibolandi, M.; Abnous, K.; Taghdisi, S.M. An optical aptasensor for aflatoxin M1 detection based on target-induced protection of gold nanoparticles against salt-induced aggregation and silica nanoparticles. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2021, 246, 119062. [Google Scholar] [CrossRef]
- Fu, K.; Zheng, Y.; Li, J.; Liu, Y.; Pang, B.; Song, X.; Xu, K.; Wang, J.; Zhao, C. Colorimetric immunoassay for rapid detection of Vibrio parahemolyticus based on Mn2+ mediates the assembly of gold nanoparticles. J. Agric. Food Chem. 2018, 66, 9516–9521. [Google Scholar] [CrossRef]
- Peng, H.; Chen, I.A. Rapid colorimetric detection of bacterial species through the capture of gold nanoparticles by chimeric phages. ACS Nano 2019, 13, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Borg, R.E.; Nguyen, A.B.N.; Chen, I.A. Chimeric phage nanoparticles for rapid characterization of bacterial pathogens, detection in complex biological samples and determination of antibiotic sensitivity. ACS Sens. 2020, 5, 1491–1499. [Google Scholar] [CrossRef]
- Xu, X.; Yuan, Y.; Hu, G.; Wang, X.; Qi, P.; Wang, Z.; Wang, Q.; Wang, X.; Fu, Y.; Li, Y.; et al. Exploiting pH-regulated dimer-tetramer transformation of concanavalin A to develop colorimetric biosensing of bacteria. Sci. Rep. 2017, 7, 1452. [Google Scholar] [CrossRef] [PubMed]
- Chotchuang, T.; Cheewasedtham, W.; Jayeoye, T.J.; Rujiralai, T. Colorimetric determination of fumonisin B1 based on the aggregation of cysteamine-functionalized gold nanoparticles induced by a product of its hydrolysis. Mikrochim. Acta 2019, 186, 655. [Google Scholar] [CrossRef]
- Yang, H.; Xiao, M.; Lai, W.; Wan, Y.; Li, L.; Pei, H. Stochastic DNA dual-walkers for ultrafast colorimetric bacteria detection. Anal. Chem. 2020, 92, 4990–4995. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zong, X.; Zheng, J.; Zhang, J.; Zhou, M.; Chen, Q.; Man, C.; Jiang, Y. A colorimetric strategy based on aptamer-catalyzed hairpin assembly for the on-site detection of Salmonella typhimurium in milk. Foods 2021, 10, 2539. [Google Scholar] [CrossRef] [PubMed]
- Albanese, A.; Chan, W.C. Effect of gold nanoparticle aggregation on cell uptake and toxicity. ACS Nano 2011, 5, 5478–5489. [Google Scholar] [CrossRef] [PubMed]
- Thuy Nguyen, T.T.; Han, O.A.; Lim, E.B.; Haam, S.; Park, J.S.; Lee, S.W. The effect of pH and transition metal ions on cysteine-assisted gold aggregation for a distinct colorimetric response. RSC Adv. 2021, 11, 9664–9674. [Google Scholar] [CrossRef] [PubMed]
- Dodero, G.; De Michieli, L.; Cavalleri, O.; Rolandi, R.; Oliveri, L.; Daccà, A.; Parodi, R. L-Cysteine chemisorption on gold: An XPS and STM study. Colloids Surf. A 2000, 175, 121–128. [Google Scholar] [CrossRef]
- Aryal, S.; Remant, B.K.C.; Dharmaraj, N.; Bhattarai, N.; Kim, C.H.; Kim, H.Y. Spectroscopic identification of S-Au interaction in cysteine capped gold nanoparticles. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2006, 63, 160–163. [Google Scholar] [CrossRef]
- Kreibig, U.; Genzel, L. Optical absorption of small metallic particles. Surf. Sci. 1985, 156, 678–700. [Google Scholar] [CrossRef]
- Brovko, L.Y.; Anany, H.; Griffiths, M.W. Bacteriophages for detection and control of bacterial pathogens in food and food-processing environment. Adv. Food Nutr. Res. 2012, 67, 241–288. [Google Scholar] [PubMed]
- Yang, X.; Wisuthiphaet, N.; Young, G.M.; Nitin, N. Rapid detection of Escherichia coli using bacteriophage-induced lysis and image analysis. PLoS ONE 2020, 15, e0233853. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.H.; Huang, W.C.; Shyu, R.H.; Chang, S.C. Silver nanoparticle-base lateral flow immunoassay for rapid detection of Staphylococcal enterotoxin B in milk and honey. J. Inorg. Biochem. 2020, 210, 111163. [Google Scholar] [CrossRef]
- Bu, T.; Huang, Q.; Yan, L.; Huang, L.; Zhang, M.; Yang, Q.; Yang, B.; Wang, J.; Zhang, D. Ultra technically-simple and sensitive detection for Salmonella enteritidis by immunochromatographic assay based on gold growth. Food Control 2018, 84, 536–543. [Google Scholar] [CrossRef]
- Rodriguez-Quijada, C.; Lyons, C.; Santamaria, C.; Quinn, S.; Tlusty, M.F.; Shiaris, M.; Hamad-Schifferli, K. Optimization of paper-based nanoparticle immunoassays for direct detection of the bacterial pathogen V. parahaemolyticus in oyster hemolymph. Anal. Methods 2020, 12, 3056–3063. [Google Scholar] [CrossRef]
- Lu, C.; Gao, X.; Chen, Y.; Ren, J.; Liu, C. Aptamer-based lateral flow test strip for the simultaneous detection of Salmonella typhimurium, Escherichia coli O157:H7 and Staphylococcus aureus. Anal. Lett. 2020, 53, 646–659. [Google Scholar] [CrossRef]
- Ying, N.; Wang, Y.; Song, X.; Yang, L.; Qin, B.; Wu, Y.; Fang, W. Lateral flow colorimetric biosensor for detection of Vibrio parahaemolyticus based on hybridization chain reaction and aptamer. Mikrochim. Acta 2021, 188, 381. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Zhou, Y.; Huang, X.; Zhang, W.; Wu, Y.; Fang, H.; Zhang, C.; Xiong, Y. Dramatically enhanced immunochromatographic assay using cascade signal amplification for ultrasensitive detection of Escherichia coli O157:H7 in milk. J. Agric. Food Chem. 2020, 68, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Zhang, L.; Hu, L.; Xing, K.; Lu, X.; Huang, Y.; Zhang, J.; Lai, W.; Chen, T. Nanozyme-based lateral flow assay for the sensitive detection of Escherichia coli O157:H7 in milk. J. Dairy Sci. 2018, 101, 5770–5779. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Song, Y.; Wei, T.; Li, H.; Du, D.; Zhu, M.J.; Lin, Y. Sensitive detection of Escherichia coli O157: H7 using Pt-Au bimetal nanoparticles with peroxidase-like amplification. Biosens. Bioelectron. 2016, 77, 687–694. [Google Scholar] [CrossRef]
- Park, J.; Shin, J.H.; Park, J.-K. Pressed paper-based dipstick for detection of foodborne pathogens with multistep reactions. Anal. Chem. 2016, 88, 3781–3788. [Google Scholar] [CrossRef] [PubMed]
- Prakash, C.; Kumar, B.; Singh, R.P.; Singh, P.; Shrinet, G.; Das, A.; Ashmi, M.; Abhishek Singh, K.P.; Singh, M.K.; Gupta, V.K. Development and evaluation of a gold nanoparticle based lateral flow assay (LFA) strip test for detection of Brucella spp. J. Microbiol. Methods 2021, 184, 106185. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q. Simultaneous quantitative analysis of Listeria monocytogenes and Staphylococcus aureus based on antibiotic-introduced lateral flow immunoassay. Anal. Methods 2021, 13, 5866–5874. [Google Scholar] [CrossRef]
- Porras, J.C.; Bernuz, M.; Marfa, J.; Pallares-Rusiñol, A.; Martí, M.; Pividori, M.I. Comparative study of gold and carbon nanoparticles in nucleic acid lateral flow assay. Nanomaterials 2021, 11, 741. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.H.; Huang, W.C.; Chang, S.C.; Shyu, R.H. Colloidal silver-based lateral flow immunoassay for detection of profenofos pesticide residue in vegetables. RSC Adv. 2022, 12, 13035–13044. [Google Scholar] [CrossRef]
- Liang, P.; Guo, Q.; Zhao, T.; Wen, C.Y.; Tian, Z.; Shang, Y.; Xing, J.; Jiang, Y.; Zeng, J. Ag nanoparticles with ultrathin Au shell-based lateral flow immunoassay for colorimetric and SERS dual-mode detection of SARS-CoV-2 IgG. Anal. Chem. 2022, 94, 8466–8473. [Google Scholar] [CrossRef]
- Shi, F.; Zhao, Y.; Sun, Y.; Chen, C. Development and application of a colloidal carbon test strip for the detection of antibodies against Mycoplasma bovis. World J. Microbiol. Biotechnol. 2020, 36, 157. [Google Scholar] [CrossRef]
- Li, C.M.; Li, Y.F.; Wang, J.; Huang, C.Z. Optical investigations on ATP-induced aggregation of positive-charged gold nanoparticles. Talanta 2010, 81, 1339–1345. [Google Scholar] [CrossRef]
- Du, X.J.; Zhou, T.J.; Li, P.; Wang, S. A rapid Salmonella detection method involving thermophilic helicase-dependent amplification and a lateral flow assay. Mol. Cell. Probes 2017, 34, 37–44. [Google Scholar] [CrossRef]
- Özay, B.; McCalla, E.S. A review of reaction enhancement strategies for isothermal nucleic acid amplification reactions. Sens. Actuator Rep. 2021, 3, 100033. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, Y.; Xie, H.; Zhao, L.; Zheng, L.; Ye, H. Applications of catalytic hairpin assembly reaction in biosensing. Small 2019, 15, 1902989. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fu, C.; Shi, W.; Chen, J. Recent advances in catalytic hairpin assembly signal amplification-based sensing strategies for microRNA detection. Talanta 2021, 235, 122735. [Google Scholar] [CrossRef]
- Chen, Q.; Lin, J.; Gan, C.; Wang, Y.; Wang, D.; Xiong, Y.; Lai, W.; Li, Y.; Wang, M. A sensitive impedance biosensor based on immunomagnetic separation and urease catalysis for rapid detection of Listeria monocytogenes using an immobilization-free interdigitated array microelectrode. Biosens. Bioelectron. 2015, 74, 504–511. [Google Scholar] [CrossRef]
- Chen, Q.; Wang, D.; Cai, G.; Xiong, Y.; Li, Y.; Wang, M.; Huo, H.; Lin, J. Fast and sensitive detection of foodborne pathogen using electrochemical impedance analysis, urease catalysis and microfluidics. Biosens. Bioelectron. 2016, 86, 770–776. [Google Scholar] [CrossRef] [PubMed]
- Du, N.; Chen, M.; Liu, Z.; Sheng, L.; Xu, H.; Chen, S. Kinetics and mechanism of jack bean urease inhibition by Hg2+. Chem. Cent. J. 2012, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Ren, R.; Cai, G.; Yu, Z.; Zeng, Y.; Tang, D. Metal-polydopamine framework: An innovative signal-generation tag for colorimetric immunoassay. Anal. Chem. 2018, 90, 11099–11105. [Google Scholar] [CrossRef]
- Jiao, L.; Yan, H.; Xu, W.; Wu, Y.; Gu, W.; Li, H.; Du, D.; Lin, Y.; Zhu, C. Self-assembly of all-inclusive allochroic nanoparticles for the improved ELISA. Anal. Chem. 2019, 91, 8461–8465. [Google Scholar] [CrossRef]
- Courbat, J.; Briand, D.; Damon-Lacoste, J.; Wöllenstein, J.; de Rooij, N.F. Evaluation of pH indicator-based colorimetric films for ammonia detection using optical waveguides. Sens. Actuators B Chem. 2009, 143, 62–70. [Google Scholar] [CrossRef]
- Xu, W.; Jiao, L.; Ye, H.; Guo, Z.; Wu, Y.; Yan, H.; Gu, W.; Du, D.; Lin, Y.; Zhu, C. pH-responsive allochroic nanoparticles for the multicolor detection of breast cancer biomarkers. Biosens. Bioelectron. 2020, 148, 111780. [Google Scholar] [CrossRef]
- Hodgkinson, V.; Petris, M.J. Copper homeostasis at the host-pathogen interface. J. Biol. Chem. 2012, 287, 13549–13555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rensing, C.; Grass, G. Escherichia coli mechanisms of copper homeostasis in a changing environment. FEMS Microbiol. Rev. 2003, 27, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, M.; Qin, Y.; Chu, Z.; Zhao, S. A Convenient label free colorimetric assay for pyrophosphatase activity based on a pyrophosphate-inhibited Cu2+-ABTS-H2O2 reaction. Analyst 2014, 139, 6298–6303. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Huang, Y.; Liu, W.; Zuo, W.; Ye, F.; Zhao, S. Colorimetric detection of salicylic acid in aspirin using MIL-53(Fe) nanozyme. Front. Chem. 2020, 8, 671. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, Y.; Guo, X.; Liu, W.; Lv, C.; Zhang, C.; Jin, Y.; Li, B. Fe(III) bipyridyl or phenanthroline complexes with oxidase-like activity for sensitive colorimetric detection of glutathione. Luminescence 2020, 35, 1350–1359. [Google Scholar] [CrossRef]
- Gao, L.; Zhuang, J.; Nie, L.; Zhang, J.; Zhang, Y.; Gu, N.; Wang, T.; Feng, J.; Yang, D.; Perrett, S.; et al. Intrinsic peroxidase-like activity of ferromagnetic nanoparticles. Nat. Nanotechnol. 2007, 2, 577–583. [Google Scholar] [CrossRef]
- Jia, H.; Yang, D.; Han, X.; Cai, J.; Liu, H.; He, W. Peroxidase-like activity of the Co3O4 nanoparticles used for biodetection and evaluation of antioxidant behavior. Nanoscale 2016, 8, 5938–5945. [Google Scholar] [CrossRef]
- Chen, W.; Fang, X.; Li, H.; Cao, H.; Kong, J. DNA-mediated inhibition of peroxidase-like activities on platinum nanoparticles for simple and rapid colorimetric detection of nucleic acids. Biosens. Bioelectron. 2017, 94, 169–175. [Google Scholar] [CrossRef]
- Li, J.; Liu, W.; Wu, X.; Gao, X. Mechanism of pH-switchable peroxidase and catalase-like activities of gold, silver, platinum and palladium. Biomaterials 2015, 48, 37–44. [Google Scholar] [CrossRef]
- Wang, S.; Cazelles, R.; Liao, W.C.; Vázquez-González, M.; Zoabi, A.; Abu-Reziq, R.; Willner, I. Mimicking horseradish peroxidase and NADH peroxidase by heterogeneous Cu2+-modified graphene oxide nanoparticles. Nano Lett. 2017, 17, 2043–2048. [Google Scholar] [CrossRef]
- Lai, W.; Tang, D.; Zhuang, J.; Chen, G.; Yang, H. Magnetic bead-based enzyme-chromogenic substrate system for ultrasensitive colorimetric immunoassay accompanying cascade reaction for enzymatic formation of squaric acid-iron(III) chelate. Anal. Chem. 2014, 86, 5061–5068. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Du, J. Selective sensing of submicromolar iron(III) with 3,3′,5,5′-tetramethylbenzidine as a chromogenic probe. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2016, 158, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Zheng, A.; Zhang, X.; Gao, J.; Liu, X.; Liu, J. Peroxidase-like catalytic activity of copper ions and its application for highly sensitive detection of glypican-3. Anal. Chim. Acta 2016, 941, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zou, Y.; Huang, M.; Dong, S.; Wu, X.; Liang, G.; Han, Z.; Zhang, Z. A sensitive, colorimetric immunosensor based on Cu-MOFs and HRP for detection of dibutyl phthalate in environmental and food samples. Talanta 2018, 186, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Pang, H.; Dong, Y.; Chi, Y.; Fu, F. Colorimetric determination of glutathione by using a nanohybrid composed of manganese dioxide and carbon dots. Mikrochim. Acta 2018, 185, 291. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; He, J.; Niu, Y.; Chen, J.; Zhao, Y.; Zhang, Y.; Yu, C. Sandwich-type biosensor for the detection of alpha2, 3-sialylated glycans based on fullerene-palladium-platinum alloy and 4-mercaptophenylboronic acid nanoparticle hybrids coupled with Au-methylene blue-MAL signal amplification. Biosens. Bioelectron. 2018, 102, 321–327. [Google Scholar] [CrossRef]
- Bryła, M.; Roszko, M.; Szymczyk, K.; Jędrzejczak, R.; Obiedziński, M.W. Fumonisins and their masked forms in maize products. Food Control 2016, 59, 619–627. [Google Scholar] [CrossRef]
- Voss, K.A.; Smith, G.W.; Haschek, W.M. Fumonisins: Toxicokinetics, mechanism of action and toxicity. Anim. Feed Sci. Technol. 2007, 137, 299–325. [Google Scholar] [CrossRef]
- Li, Y.S.; Zhou, Y.; Lu, S.Y.; Guo, D.J.; Ren, H.L.; Meng, X.M.; Zhi, B.H.; Lin, C.; Wang, Z.; Li, X.B.; et al. Development of a one-step test strip for rapid screening of fumonisins B1, B2 and B3 in maize. Food Control 2012, 24, 72–77. [Google Scholar] [CrossRef]
- Shi, F.; Sun, Y.; Wu, Y.; Zhu, M.; Feng, D.; Zhang, R.; Peng, L.; Chen, C. A novel, rapid and simple method for detecting brucellosis based on rapid vertical flow technology. J. Appl. Microbiol. 2020, 128, 794–802. [Google Scholar] [CrossRef]
- Morbioli, G.G.; Mazzu-Nascimento, T.; Stockton, A.M.; Carrilho, E. Technical aspects and challenges of colorimetric detection with microfluidic paper-based analytical devices (μPADs)—A review. Anal. Chim. Acta 2017, 970, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Kanchi, S.; Sabela, M.I.; Mdluli, P.S.; Inamuddin; Bisetty, K. Smartphone based bioanalytical and diagnosis applications: A review. Biosens. Bioelectron. 2018, 102, 136–149. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Colorimetric sensing strategy based on external pH change-induced pH indicator reactions. (A) Detection of Listeria monocytogenes using an antibody (Ab), urease-modified magnetic nanobeads, and gold nanoparticles (AuNPs). Reprinted with permission from [19]. Copyright 2017, Elsevier. (B) Detection of Salmonella typhimurium using silver ions and urease. This strategy is based on the Ag-induced inhibition of urease activity and Ag ion utilization. Reprinted with permission from [21]. Copyright 2020, Springer Nature. (C) Detection of Escherichia coli and S. typhimurium using two different pH-responsive NPs, which were made using PP or TP. Reprinted with permission from [22]. Copyright 2022, Elsevier. (D) Detection of bacterial spoilage using a paper-based pH indicator consisting of BTB and BCG. Quantification can be performed using RGB analysis software on a smartphone. Reprinted with permission from [23]. Copyright 2021, Royal Society of Chemistry. PEI, polyethyleneimine; BSA, bovine serum albumin; RGB, red-green-blue.
Figure 1.
Colorimetric sensing strategy based on external pH change-induced pH indicator reactions. (A) Detection of Listeria monocytogenes using an antibody (Ab), urease-modified magnetic nanobeads, and gold nanoparticles (AuNPs). Reprinted with permission from [19]. Copyright 2017, Elsevier. (B) Detection of Salmonella typhimurium using silver ions and urease. This strategy is based on the Ag-induced inhibition of urease activity and Ag ion utilization. Reprinted with permission from [21]. Copyright 2020, Springer Nature. (C) Detection of Escherichia coli and S. typhimurium using two different pH-responsive NPs, which were made using PP or TP. Reprinted with permission from [22]. Copyright 2022, Elsevier. (D) Detection of bacterial spoilage using a paper-based pH indicator consisting of BTB and BCG. Quantification can be performed using RGB analysis software on a smartphone. Reprinted with permission from [23]. Copyright 2021, Royal Society of Chemistry. PEI, polyethyleneimine; BSA, bovine serum albumin; RGB, red-green-blue.

Figure 2.
Colorimetric sensing strategy based on intracellular enzyme-catalyzed chromogenic reactions. (A) Discrimination of Escherichia coli and E. coli O157:H7 based on different cellular metabolic reactions (β-glucuronidase- and β-galactosidase-based enzymatic reactions, respectively). Reprinted with permission from [32]. Copyright 2019, American Chemical Society. (B) Detection of E. coli and Staphylococcus aureus based on the reduction reaction of p-benzoquinone by intracellular enzymes. Reprinted with permission from [34]. Copyright 2019, American Chemical Society. (C) Detection of E. coli, S. aureus, Enterococcus faecalis, Streptococcus mutans, and Salmonella pullorum based on the conversion from iodide to iodine on the starch–iodide doping paper. This conversion is catalyzed by H2O2 produced from glucose oxidase-mediated oxidation and glucose uptake of bacterial cells. Reprinted with permission from [35]. Copyright 2019, Elsevier. WST-8, 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt; RGB, red-green-blue; FIB, fecal indicator bacteria.
Figure 2.
Colorimetric sensing strategy based on intracellular enzyme-catalyzed chromogenic reactions. (A) Discrimination of Escherichia coli and E. coli O157:H7 based on different cellular metabolic reactions (β-glucuronidase- and β-galactosidase-based enzymatic reactions, respectively). Reprinted with permission from [32]. Copyright 2019, American Chemical Society. (B) Detection of E. coli and Staphylococcus aureus based on the reduction reaction of p-benzoquinone by intracellular enzymes. Reprinted with permission from [34]. Copyright 2019, American Chemical Society. (C) Detection of E. coli, S. aureus, Enterococcus faecalis, Streptococcus mutans, and Salmonella pullorum based on the conversion from iodide to iodine on the starch–iodide doping paper. This conversion is catalyzed by H2O2 produced from glucose oxidase-mediated oxidation and glucose uptake of bacterial cells. Reprinted with permission from [35]. Copyright 2019, Elsevier. WST-8, 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt; RGB, red-green-blue; FIB, fecal indicator bacteria.

Figure 3.
Colorimetric sensing strategy based on enzyme-like NP-catalyzed substrate reactions. (A) Detection of Salmonella typhimurium based on the peroxidase-like catalytic reaction of a ZnFe2O4/rGO nanostructure. Reprinted with permission from [37]. Copyright 2017, Elsevier. (B) Detection of Salmonella spp. based on the peroxidase-like catalytic reaction of rGO. Reprinted with permission from [38]. Copyright 2020, Elsevier. (C) Detection of S. typhimurium using AuNCs. Reprinted with permission from [43]. Copyright 2020, Elsevier. TMB, 5,5′-tetramethylbenzidine; oxTMB, oxidized TMB; AuNC, gold nanocrystal; rGO, reduced graphene oxide; PCR, polymerase chain reaction; apt, aptamer.
Figure 3.
Colorimetric sensing strategy based on enzyme-like NP-catalyzed substrate reactions. (A) Detection of Salmonella typhimurium based on the peroxidase-like catalytic reaction of a ZnFe2O4/rGO nanostructure. Reprinted with permission from [37]. Copyright 2017, Elsevier. (B) Detection of Salmonella spp. based on the peroxidase-like catalytic reaction of rGO. Reprinted with permission from [38]. Copyright 2020, Elsevier. (C) Detection of S. typhimurium using AuNCs. Reprinted with permission from [43]. Copyright 2020, Elsevier. TMB, 5,5′-tetramethylbenzidine; oxTMB, oxidized TMB; AuNC, gold nanocrystal; rGO, reduced graphene oxide; PCR, polymerase chain reaction; apt, aptamer.

Figure 4.
Colorimetric sensing strategy based on the aggregation of NPs. (A) Detection of Vibrio parahemolyticus based on Mn2+-mediated aggregation of AuNPs. Reprinted with permission from [51]. Copyright 2018, American Chemical Society. (B) Detection of Escherichia coli, Vibrio cholerae, Pseudomonas aeruginosa, and Xanthomonas campestris based on engineered bacteriophage-mediated aggregation of AuNPs. Reprinted with permission from [52]. Copyright 2018, American Chemical Society. (C) Detection of E. coli O157:H7 through pH-responsive concanavalin A-mediated aggregation of AuNPs. Reprinted with permission from [54]. Copyright 2017, Springer Nature. (D) Detection of Staphylococcus aureus using DNA dual-walker signal amplification combined with colorimetric systems. Reprinted with permission from [56]. Copyright 2020, American Chemical Society. (E) Detection of Salmonella typhimurium using colorimetric systems in combination with the aptamer-catalytic hairpin assembly (CHA) method. Reprinted with permission from [57]. Copyright 2021, MDPI. GNP, gold nanoparticle; AA, ascorbic acid; RBP, receptor-binding protein; EDC, 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide; ConA, concanavalin A; MNP, magnetic nanoparticle; ExoIII, exonuclease III; MRSA, methicillin-resistant S. aureus; SMB, streptavidin-coated MNP; Bapt, biotinylated aptamer; Tapt, non-labelled aptamer; Y-CHA, Y-shaped catalytic hairpin assembly.
Figure 4.
Colorimetric sensing strategy based on the aggregation of NPs. (A) Detection of Vibrio parahemolyticus based on Mn2+-mediated aggregation of AuNPs. Reprinted with permission from [51]. Copyright 2018, American Chemical Society. (B) Detection of Escherichia coli, Vibrio cholerae, Pseudomonas aeruginosa, and Xanthomonas campestris based on engineered bacteriophage-mediated aggregation of AuNPs. Reprinted with permission from [52]. Copyright 2018, American Chemical Society. (C) Detection of E. coli O157:H7 through pH-responsive concanavalin A-mediated aggregation of AuNPs. Reprinted with permission from [54]. Copyright 2017, Springer Nature. (D) Detection of Staphylococcus aureus using DNA dual-walker signal amplification combined with colorimetric systems. Reprinted with permission from [56]. Copyright 2020, American Chemical Society. (E) Detection of Salmonella typhimurium using colorimetric systems in combination with the aptamer-catalytic hairpin assembly (CHA) method. Reprinted with permission from [57]. Copyright 2021, MDPI. GNP, gold nanoparticle; AA, ascorbic acid; RBP, receptor-binding protein; EDC, 1-ethyl-3-(3-dimethylaminopropyl)-carbodiimide; ConA, concanavalin A; MNP, magnetic nanoparticle; ExoIII, exonuclease III; MRSA, methicillin-resistant S. aureus; SMB, streptavidin-coated MNP; Bapt, biotinylated aptamer; Tapt, non-labelled aptamer; Y-CHA, Y-shaped catalytic hairpin assembly.

Figure 5.
Colorimetric sensing strategy based on the accumulation of NPs. (A) Detection of staphylococcal enterotoxin B based on color readout by concentrated AgNPs on the lateral flow immunoassay (LFIA) strip. Reprinted with permission from [65]. Copyright 2020, Elsevier. (B) Detection of Escherichia coli O157:H7 based on color readout by concentrated AuNPs on the LFIA strip. This system consisted of a combination of two methods for cascade signal amplification. Reprinted with permission from [70]. Copyright 2020, American Chemical Society. AEC, 3-amino-9-ethylcarbazole; ICA, immunochromatography assay; SEB, staphylococcal enterotoxin B; mAB, monoclonal antibody.
Figure 5.
Colorimetric sensing strategy based on the accumulation of NPs. (A) Detection of staphylococcal enterotoxin B based on color readout by concentrated AgNPs on the lateral flow immunoassay (LFIA) strip. Reprinted with permission from [65]. Copyright 2020, Elsevier. (B) Detection of Escherichia coli O157:H7 based on color readout by concentrated AuNPs on the LFIA strip. This system consisted of a combination of two methods for cascade signal amplification. Reprinted with permission from [70]. Copyright 2020, American Chemical Society. AEC, 3-amino-9-ethylcarbazole; ICA, immunochromatography assay; SEB, staphylococcal enterotoxin B; mAB, monoclonal antibody.


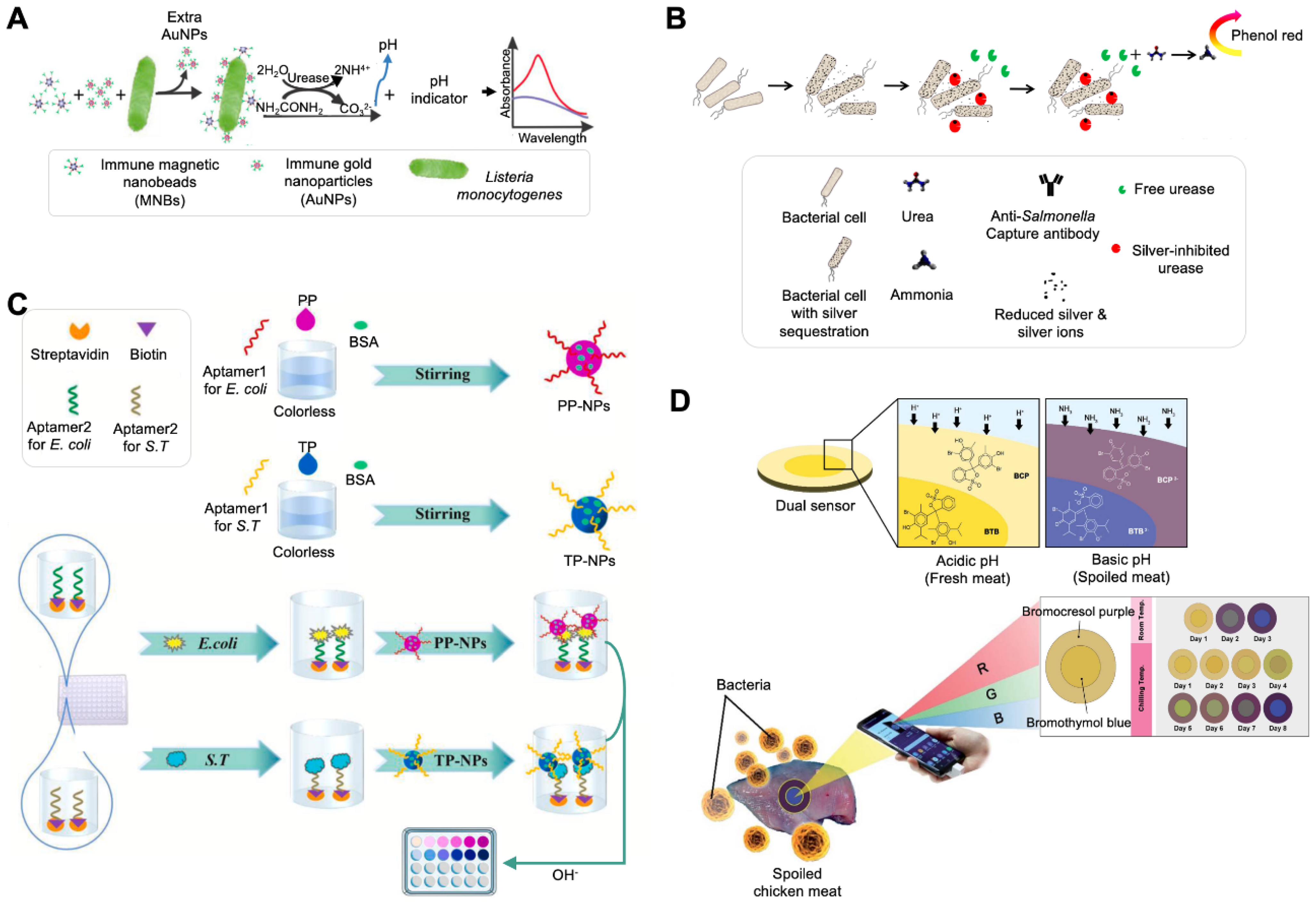{kind=link}
{kind=link}
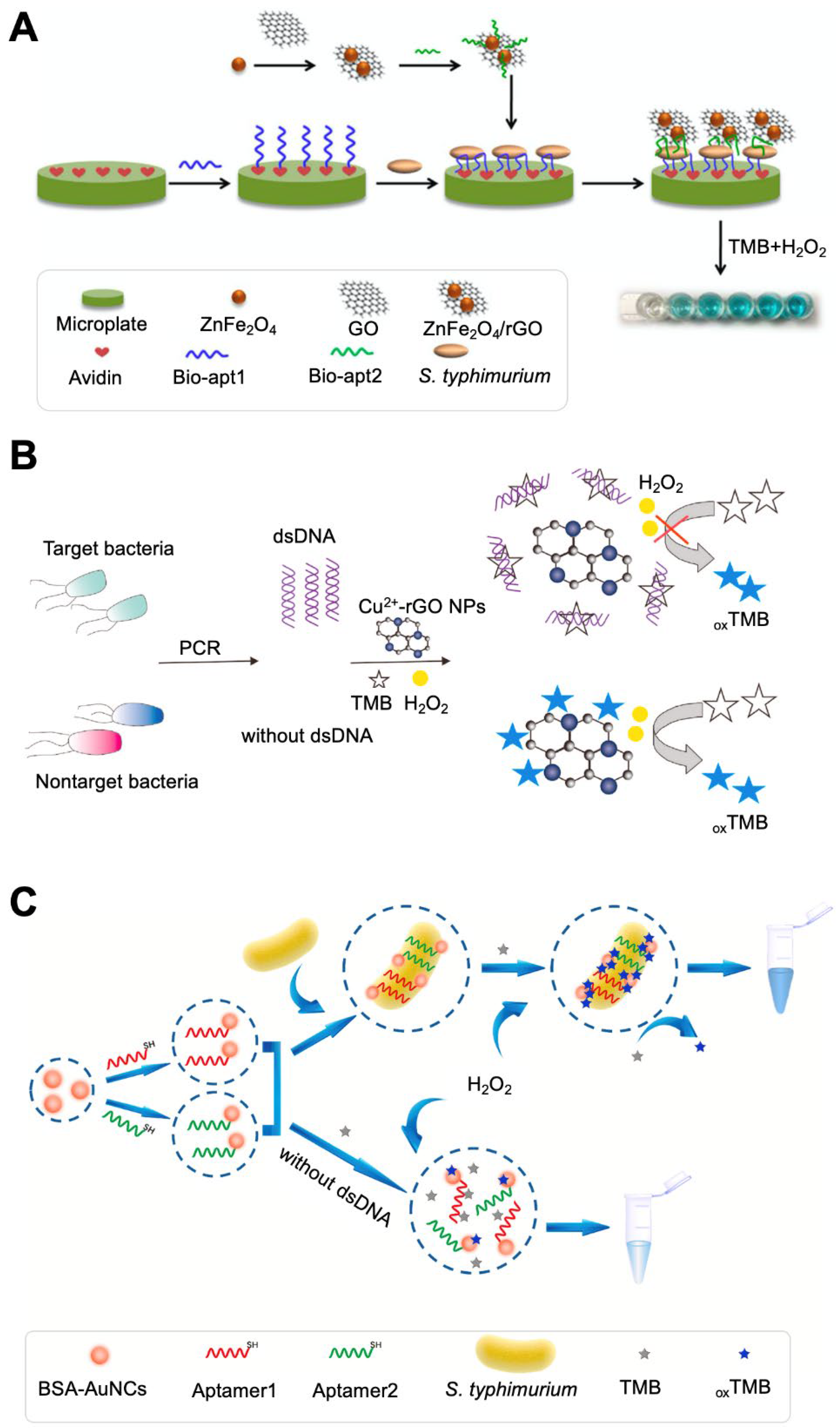{kind=link}
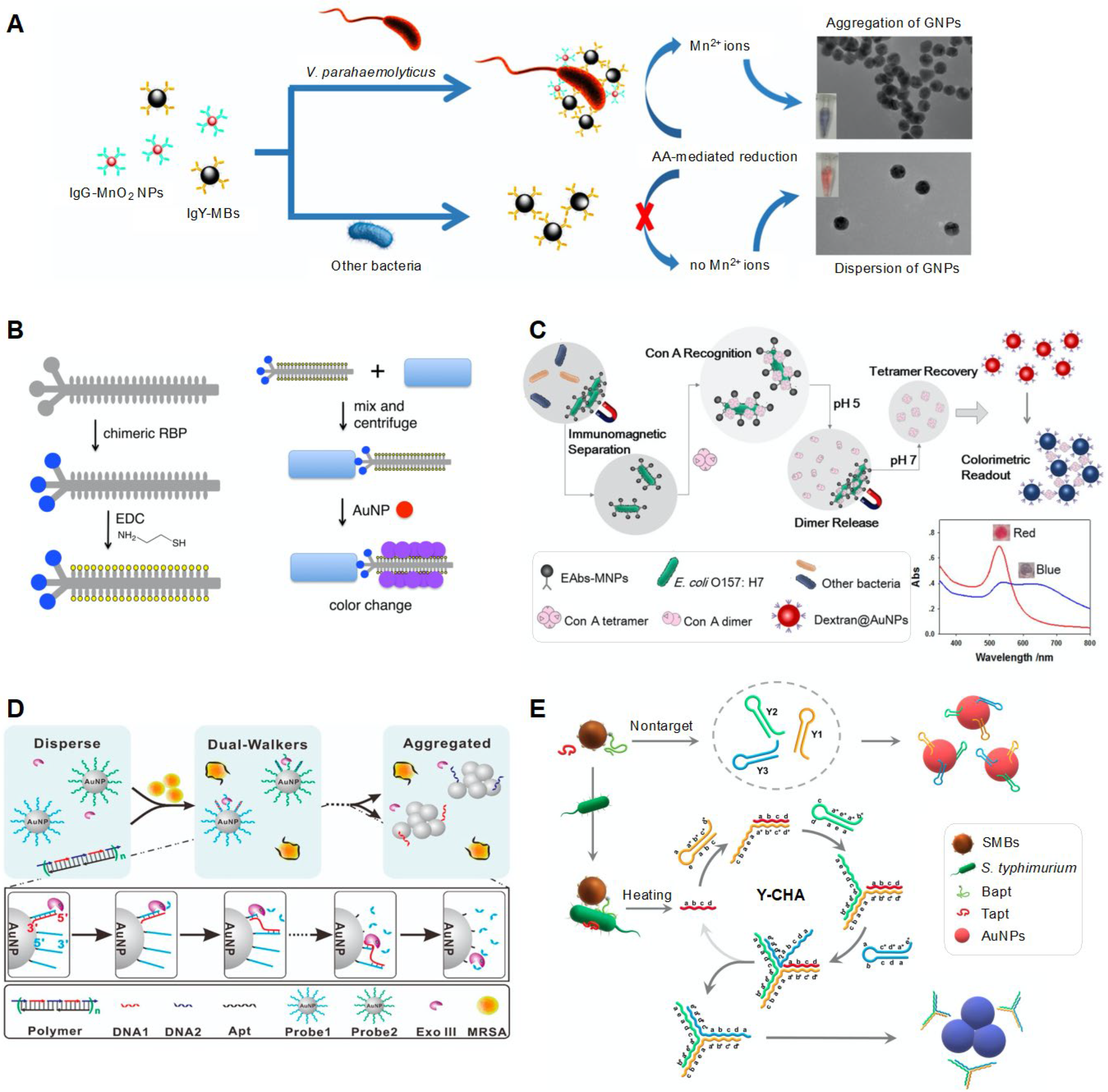{kind=link}
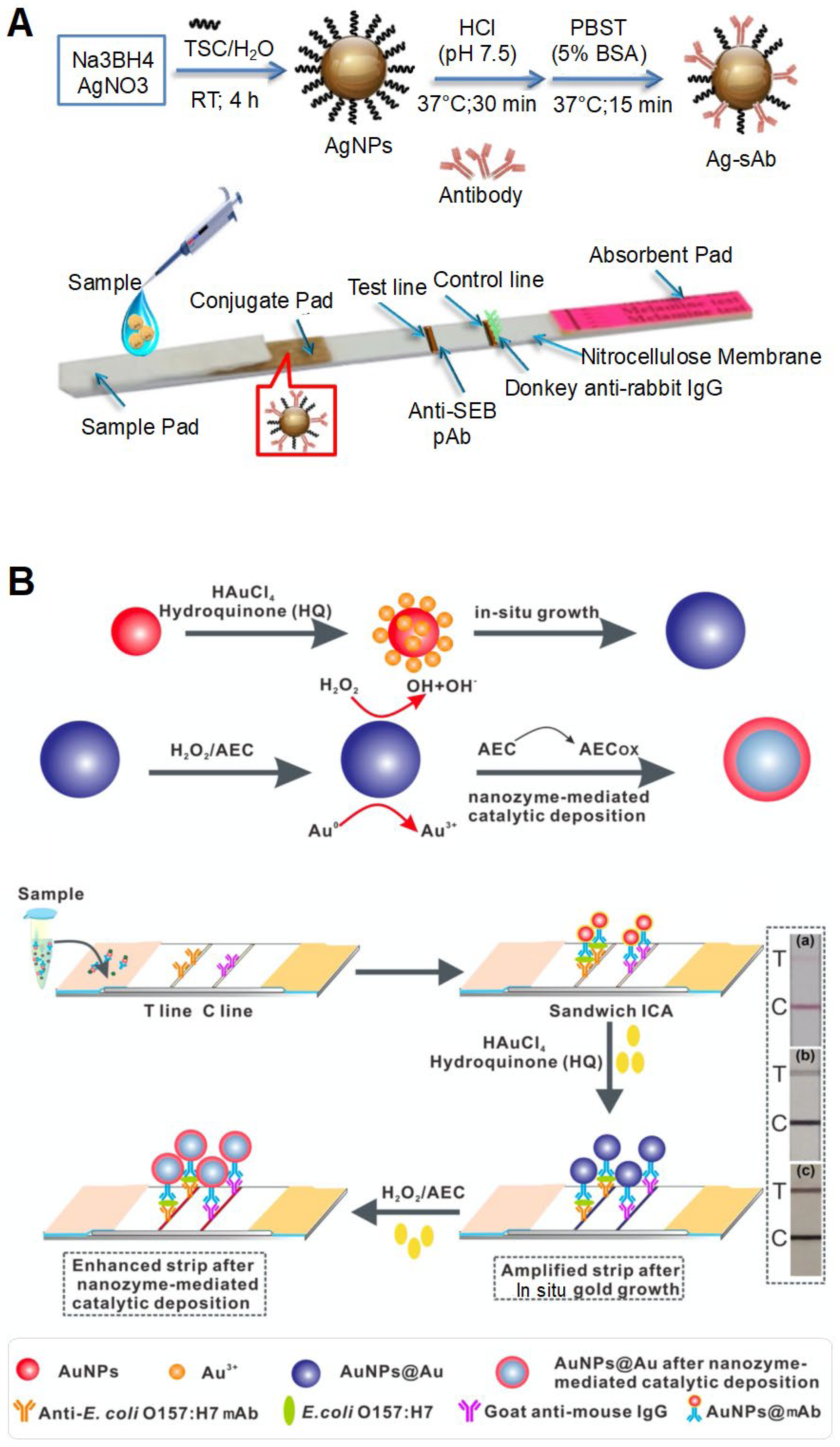{kind=link}
Table 1.
Colorimetric sensing systems for the detection of pathogenic bacteria and toxins a.
| Material | Receptor | Target | Linear Range | LoD | Assay Time | Real Sample to Be Tested | Driving Force of Color Change | Feature | References |
|---|---|---|---|---|---|---|---|---|---|
| AuNPs, magnetic beads | Monoclonal antibody, polyclonal antibody | Listeria monocytogenes | 1.1 × 102 CFU/mL–1.1 × 106 CFU/mL | 100 CFU/mL | 30 min | Lettuce samples | Induction of pH change | Use of magnetic nanobeads modified with urease and monoclonal antibodies. | [19] |
| Use of AuNPs modified with urease and polyclonal antibodies. | |||||||||
| Use of BCP. | |||||||||
| AgNPs | Monoclonal antibody | Salmonella typhimurium | 1 × 108 CFU/mL–1 × 101 CFU/mL | 100 CFU/mL | Apple juice, lake water sample | Induction of pH change | Based on the competitive binding ability of urease and bacterial cells to PEI-functionalized AgNPs. | [20] | |
| Ag ion | None | S. typhimurium | 1 × 107 CFU/mL–1 × 101 CFU/mL | 100 CFU/mL | Tap water | Induction of pH change | Based on the Ag-induced inhibition of urease activity and Ag ion utilization. | [21] | |
| Combined with electrochemical sensing. | |||||||||
| NPs | Aptamer | Escherichia coli., S. typhimurium | 1 × 105 CFU/mL–1 × 101 CFU/mL | 1 CFU/mL | <1 h | Milk | Induction of pH change | Use of pH-responsive NPs made of phenolphthalein (PP) and thymolphthalein (TP) indicators. | [22] |
| Combined with automated equipment. | |||||||||
| Allows multiplexing detection. | |||||||||
| Filter paper | None | Bacteria | 11.2 × 103–1.12 × 106 CFU/g (using BTB), 38.0 × 103–1.12 × 106 CFU/g (using BCP) | 11.64 × 103 CFU/g | Chicken and meat samples | Induction of pH change | Monitoring of bacterial contamination level using paper-based pH indicators, BTB, and BCP. | [23] | |
| Sensing of external pH change caused by volatile basic nitrogen generated from bacterial spoilage. | |||||||||
| Use of RGB analysis software on a smartphone. | |||||||||
| Filter paper | None | E. coli, E. coli O157:H7, L. monocytogenes, Vibrio vulnificus | 1 × 106–1 × 108 CFU/mL | 10 CFU/mL | 1 h | Milk | Chemical reaction between intracellular enzymes and their chromogenic substrates | One-step-based 3D paper sensor functionalized with lysing and oxidizing agents. | [31] |
| Filter paper | None | E. coli, E. coli O157:H7 | 1 × 106–1 × 109 CFU/mL | 10 CFU/mL | <4 h | Milk | Chemical reaction between intracellular enzymes and their chromogenic substrates | Use of a multi-layered paper structure. | [32] |
| Use of β-glucuronidase and β-galactosidase-based enzymatic reactions. | |||||||||
| None | None | Staphylococcus aureus, E. coli | 2.6 × 102–1.16 × 109 CFU/mL (for E. coli), 9.75 × 102–6 × 109 CFU/mL (for S. aureus) | ND | 2 h | Drinking water, milk | Redox reaction between the cell counting kit-8 (CCK-8) solution and dehydrogenase | Measurement of formazan generated from the reduction reaction between dehydrogenase and CCK-8 (containing WST-8 and 1-methoxy-5-methylphenazinium methyl sulfate). | [33] |
| None | None | E. coli | 1 × 104–1 × 109 CFU/mL | 1 × 104 CFU/mL | 1 h | Unfiltered tap water | Reduction reaction of p-benzoquinone by intracellular enzymes | Use of RGB analysis software on a smartphone for quantification. | [34] |
| Filter paper | None | E. coli, S. aureus, Enterococcus faecalis, Streptococcus mutans, Salmonella pullorum | 1 × 104–1 × 108 CFU/mL | 7.48 × 103 CFU/mL (for E. coli) and 3.3 × 103 CFU/mL (for S. aureus) | 20 min | Inhibition of GOx activity by glucose uptake of bacterial cells | Use of starch–iodide doping paper as a substrate. | [35] | |
| Based on the conversion from iodide to iodine by H2O2 involving GOx-mediated glucose oxidation (causing color change of starch–iodine) and glucose uptake of bacterial cells (causing inhibition of color change of starch–iodine). | |||||||||
| Filter paper | None | E. coli | 1 × 102–1 × 106 CFU/mL | 44 CFU/mL | Tap water, degrease milk | Inhibition of color change of OPD via Cu2+ reduction by intracellular enzymes | Use of paper as a substrate. | [36] | |
| Based on the competitive reaction between the oxidation of OPD by Cu2+ (causing color change of OPD) and the reduction of Cu2+ by bacteria (causing inhibition of color change of OPD). | |||||||||
| Use of RGB analysis software on a smartphone for quantification. | |||||||||
| Allows dual-readout assay (colorimetry and fluorescence). | |||||||||
| ZnFe2O4/rGO | Aptamer | S. typhimurium | 11–1.10 × 105 CFU/mL | 11 CFU/mL | None | Peroxidase-like catalytic reaction of the ZnFe2O4/rGO nanostructure | Shows highly stable catalytic activity at low pH (over 5.5) and high temperature (over 50 °C). | [37] | |
| Cu2-rGO NPs | None | Salmonella spp. | 1.93 × 101–1.93 × 105 CFU/mL | 0.51 CFU/mL | Milk | Peroxidase-like catalytic reaction of GO | Use of dsDNA amplified via PCR from cells. Based on the competitive binding of bacterial dsDNA and Cu2-rGO NPs to TMB. | [38] | |
| Graphitic-C3N4@Cu2O | Aptamer | S. typhimurium | 1.5 × 101–1.5 × 105 CFU/mL | 15 CFU/mL | 6 min | Milk | Peroxidase-like catalytic reaction of the g-C3N4@Cu2O nanostructure | Based on the competitive binding of the aptamer and g-C3N4@Cu2O to TMB. | [39] |
| Fe3O4/Au magnetic nanocomposite | Antibody, aptamer | S. aureus | 1 × 101–1 × 106 CFU/mL | 10 CFU/mL | Pork, milk | Peroxidase-like catalytic reaction of AuNPs by H2O2 etching | Use of a magnetic nanocomposite consisting of a Fe3O4 core and an Au shell as a capture probe. | [40] | |
| Use of Apt–AuNPs as a signal amplifier. | |||||||||
| AuNPs, magnetic beads | Antibody | Brevotoxin B | 0.1–150 ng/kg | 0.076 ng/kg | Seafood sample | Peroxidase-based TMB oxidation reaction | Addition of Fe2+ for color signal amplification. | [41] | |
| Magnetic beads | Antibody | Ochratoxin A | 0.01–10 ng/mL | 8.3 pg/mL | 30 min (for color development) | Red wine sample | Enzyme-controlled Turnbull’s blue generation | Based on the formation or inhibition of Prussian blue from K3[Fe(CN)6] via GOx-catalyzed H2O2 production. | [42] |
| Aptamer@ BSA- AuNCs | Aptamer | S. typhimurium | 1 × 101–1 × 106 CFU/mL | 1 CFU/mL | Eggshell, Egg white | Peroxidase-like catalytic reaction of AuNCs | Based on the enhanced catalytic activity of a cell-bound nanostructure (cell-aptamer@BSA-AuNC composite). | [43] | |
| MnO2-doped Fe3O4 NPs | None | S. aureus, Vibrio parahaemolyticus | 1 × 101–1 × 106 CFU/mL | 1 × 102 CFU/mL | Lake water sample | Peroxidase-like catalytic reaction | Use of multifunctional NPs for recognition, absorption, and separation of the analyte. | [44] | |
| Exhibits the catalytic activity of TMB in the presence of oxygen in a solution without H2O2. | |||||||||
| AuNPs | 4-MPBA | E. coli | 1 × 104–1 × 107 CFU/mL | 1.02 × 103 CFU/mL | 20 min | Drinking water | Salt-induced aggregation | Use of AuNPs functionalised with 4-MPBA, which binds to LPS and peptidoglycan existing on the surface of gram-negative and gram-positive bacterial cells, respectively. | [48] |
| Use of RGB analysis software on a smartphone for quantification. | |||||||||
| AuNPs | Aptamer | Shigella flexneri | 1 × 102–1 × 106 CFU/mL | 80 CFU/mL | 20 min | Salmon | Salt-induced aggregation | Use of aptamers that can bind to bacterial cells rather than AuNPs. | [49] |
| AuNPs, silica nanoparticles (SNPs) | Aptamer | Aflatoxin M1 | 300–75,000 ng/L | 30 ng/L | Milk | Salt-induced AuNP aggregation | Salt-induced aggregation by releasing complementary strands from aptamer-modified SNPs in the presence of the target. | [50] | |
| AuNPs, magnetic nanoparticles (MNPs) | Antibody | V. parahaemolyticus | 1 × 101–1 × 106 CFU/mL | 10 CFU/mL | Oyster | Mn2+-induced AuNP aggregation | Combination with the signal amplification method based on ascorbic acid-mediated Mn2+ reduction and a sandwich assay using IgG-MnO2 NPs and IgY-MNPs. | [51] | |
| AuNPs | Chimeric phage | E. coli, V. cholerae, Pseudomonas aeruginosa, Xanthomonas campestris | 1 × 102 CFU/mL | <1 h | Sea water, tap water | AuNP aggregation | Use of thiolated chimeric phages that can bind to both bacterial cells and AuNPs | [52] | |
| AuNPs | Chimeric phage | P. aeruginosa | 1 × 101–1 × 106 CFU/mL | 1 × 102 CFU/mL | ~30 min | Drinking water, non-fat bovine milk | AuNP aggregation | Detection of antibiotic resistance/susceptibility of bacterial cells | [53] |
| Dextran-coated AuNPs, MNPs | Antibody | E. coli | 1 × 103–1 × 106 CFU/mL | 41 CFU/mL | 95 min | Milk | ConA-driven aggregation of dextran-coated AuNPs | Use of ConA with pH-regulated transformation ability of dimers/tetramers | [54] |
| AuNPs | Fumonisin B1 (FB1) | 2–8 mg/kg | 0.9 mg/kg | Corn | Hydrolyzed FB1-induced AgNP aggregation | Use of cysteamine-functionalised AuNPs (Cys-AuNPs). Need for NaOH treatment to obtain hydrolyzed FB1 with a high affinity towards Cys-AuNPs. | [55] | ||
| AuNPs | DNA | S. aureus | 1–1 × 105 CFU/mL | 1 CFU/mL | 15 min | CSF, urine, spit, serum | Enzyme-driven DNA walker-induced AgNP aggregation | Use of an exonuclease III-driven DNA walker system for signal amplification. | [56] |
| AuNPs, MNPs | Aptamer | S. typhimurium | 1 × 102–1 × 106 CFU/mL | 2.4 × 102 CFU/mL | Milk | Catalytic hairpin assembly (CHA)-driven AuNP aggregation | Use of Y-shaped CHA for signal amplification. | [57] | |
| AgNPs | Antibody | Staphylococcal enterotoxin B | 0–2 ppm | 0.5 ppm | 15 min | Milk, honey | AgNP accumulation | Use of AgNP-based sandwich-type lateral flow immunoassay (LFIA). | [65] |
| AuNPs | Antibody | S. enteritidis | 1 × 105–1 × 108 CFU/mL | 1 × 104 CFU/mL | 20 min | Milk | AuNP accumulation | Use of LFIA. | [66] |
| Use of a signal enhancer, HAuCl4 and NH2OH·HCl for in situ AuNP growth. | |||||||||
| AuNPs | Antibody | V. parahaemolyticus | 4.66 × 105 CFU/mL | 2 h | Oyster hemolymph | AuNP accumulation | Use of a dipstick. | [67] | |
| AuNPs | Aptamer | S. typhimurium, E. coli O157:H7, S. aureus | 1 × 103 CFU/mL for S. typhimurium and 1 × 104 CFU/mL for E. coli O157:H7 and S. aureus | 10 min | Milk, chicken, food | AuNP accumulation | Use of LFA. | [68] | |
| AuNPs, MNPs | Aptamer | V. parahaemolyticus | 1 × 103–1 × 108 CFU/mL | 2.6 × 103 CFU/mL | 67 min | Shrimp | AuNP accumulation | Combination of HCR-mediated signal amplification methods. | [69] |
| AuNPs | Antibody | E. coli O157:H7 | 1.25 × 101–1.25 × 105 CFU/mL | 1.25 × 101 CFU/mL | Milk | AuNP accumulation | Use of LFA. | [70]; | |
| Combination of two signal amplification strategies; use of a signal enhancer (hydroquinone) for in situ AuNP growth and nanozyme-mediated catalytic deposition. | |||||||||
| Pd-Pt NPs | Antibody | E. coli O157:H7 | 1 × 102–1 × 106 CFU/mL | 0.87 × 102 CFU/mL | 10 min | Milk | Pd-Pt NP accumulation-driven catalytic reaction | Use of LFA. | [71] |
| Signal readout by oxidised TMB through Pd-Pt NP-mediated catalytic reactions. | |||||||||
| Pt-Au NPs | Antibody | E. coli O157:H7 | 1 × 102–1 × 108 CFU/mL | 1 × 102 CFU/mL | 1 min | Pt-Au NP accumulation-driven catalytic reaction | Use of LFA. | [72] | |
| Use of Pt-Au-mediated signal amplification. |
a Abbreviations: LoD, limit of detection; AuNPs, gold nanoparticles; CFU, colony-forming unit; TMB, 3,3′,5,5′-tetramethylbenzidine; BTB, bromothymol blue; BCG, bromocresol green; BCP, bromocresol purple; RGB, red-green-blue; PCR, polymerase chain reaction; PEI, polyethylenimine; WST-8, 2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodium salt; OPD, o-phenylenediamine; dsDNA, double-stranded DNA; GO, graphene oxide; rGO, reduced GO; BSA, bovine serum albumin; GOx, glucose oxidase; AuNCs, gold nanoclusters; 4-MPBA, 4-mercaptophenylboronic acid; LPS, lipopolysaccharide; CSF, cerebral spinal fluid; ConA, concanavalin A; HCR, hybridization chain reaction; ND, not determined.
Table 2.
Challenges and strategies to improve the performance of colorimetric systems for the detection of bacterial contamination.
Table 2.
Challenges and strategies to improve the performance of colorimetric systems for the detection of bacterial contamination.
| Challenge | Performance Improvement Strategy | Reference(s) |
|---|---|---|
| Sensitivity |
| [22] [36] [34] [19] [56,57,69,81,82,83,84] [70,71,72] |
| Simple operation |
| [32] [21] [22] |
| Correct signal in complex real samples |
| [37,38,39,40,41,42,43,44] |
| Multiplexing capability |
| [22] [31] |
| Quantification and expansion of analyzed spectra |
| [22,23,34,36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, D.-M.; Yoo, S.-M. Colorimetric Systems for the Detection of Bacterial Contamination: Strategy and Applications. Biosensors 2022, 12, 532. https://doi.org/10.3390/bios12070532
AMA Style
Kim D-M, Yoo S-M. Colorimetric Systems for the Detection of Bacterial Contamination: Strategy and Applications. Biosensors. 2022; 12(7):532. https://doi.org/10.3390/bios12070532
Chicago/Turabian StyleKim, Dong-Min, and Seung-Min Yoo. 2022. "Colorimetric Systems for the Detection of Bacterial Contamination: Strategy and Applications" Biosensors 12, no. 7: 532. https://doi.org/10.3390/bios12070532
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.