Probing Subcellular Iron Availability with Genetically Encoded Nitric Oxide Biosensors

 , , ,
, , ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Experimental Buffers
2.3. Cellular Iron Loading Procedure
2.4. Imaging Experiments
2.5. Statistics
3. Results
3.1. Characterizing geNOps’ Ferrous Iron Dependency
3.2. Imaging Chelated Ferric Iron Bioavailability
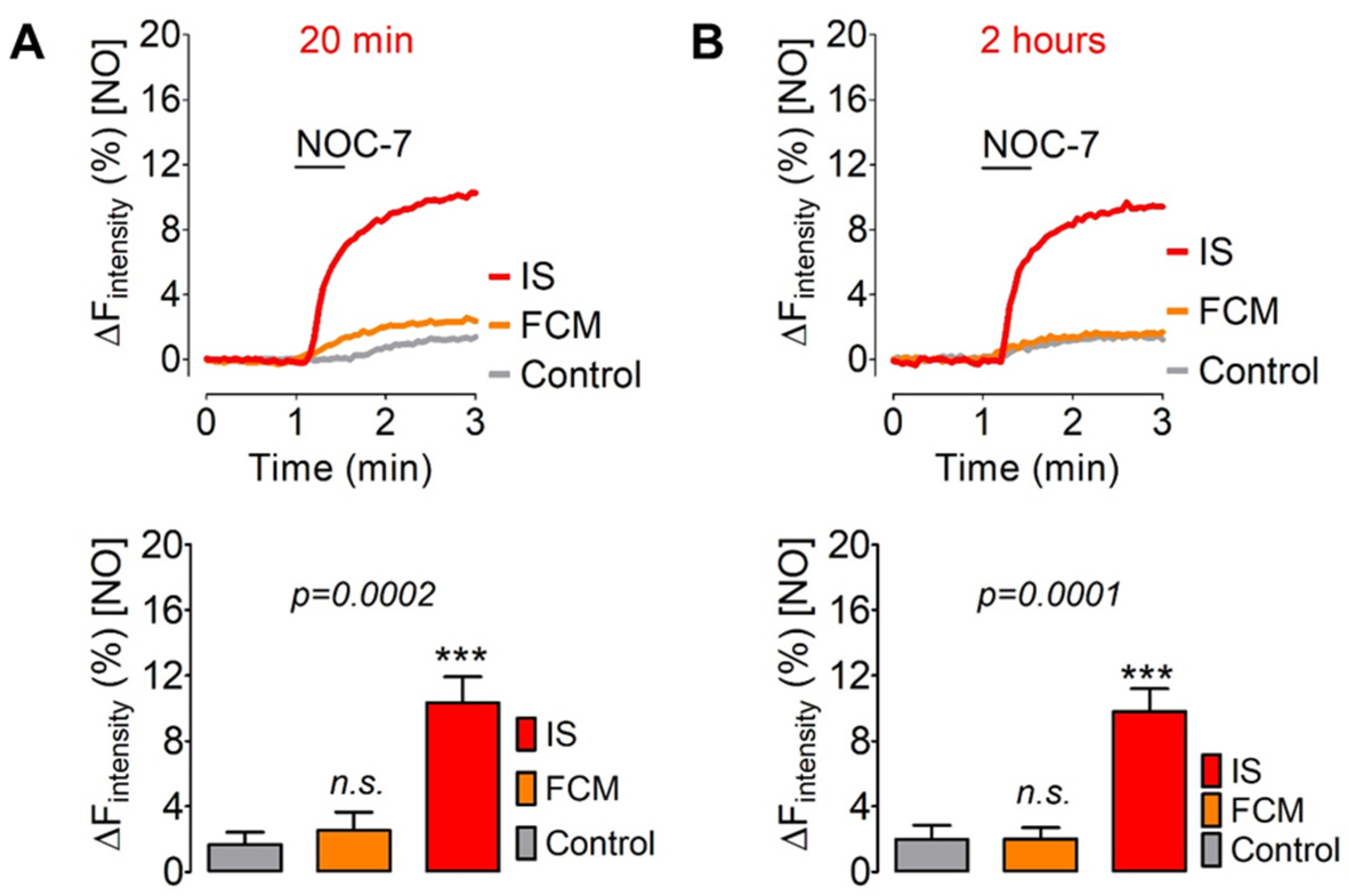
3.3. Comparing the Intracellular geNOps Activation with FCM and IS
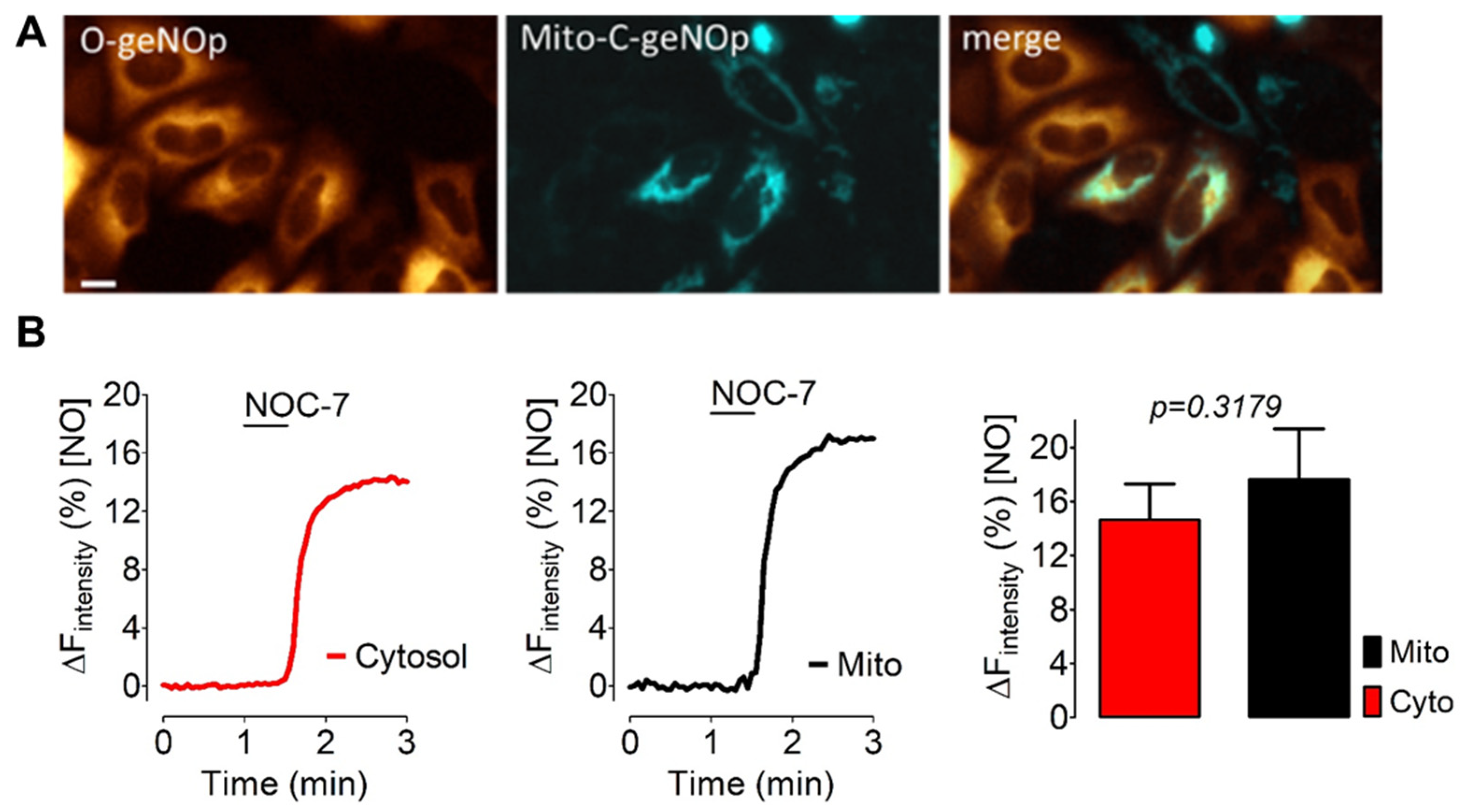
3.4. IS Supplies Ferrous Iron Also to Mitochondria of Cancer Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abbaspour, N.; Hurrell, R.; Kelishadi, R. Review on Iron and Its Importance for Human Health. J. Res. Med. Sci. 2014, 19, 164–174. [Google Scholar]
- Wang, J.; Pantopoulos, K. Regulation of Cellular Iron Metabolism. Biochem. J. 2011, 434, 365–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldvogel-Abramowski, S.; Waeber, G.; Gassner, C.; Buser, A.; Frey, B.M.; Favrat, B.; Tissot, J.-D. Physiology of Iron Metabolism. Transfus. Med. Hemother. 2014, 41, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garbowski, M.W.; Bansal, S.; Porter, J.B.; Mori, C.; Burckhardt, S.; Hider, R.C. Intravenous Iron Preparations Transiently Generate Non-Transferrin-Bound Iron from Two Proposed Pathways. Haematologica 2021, 106, 2885–2896. [Google Scholar] [CrossRef] [PubMed]
- Funk, F.; Flühmann, B.; Barton, A.E. Criticality of Surface Characteristics of Intravenous Iron–Carbohydrate Nanoparticle Complexes: Implications for Pharmacokinetics and Pharmacodynamics. Int. J. Mol. Sci. 2022, 23, 2140. [Google Scholar] [CrossRef]
- Gao, G.; Li, J.; Zhang, Y.; Chang, Y.-Z. Cellular Iron Metabolism and Regulation. In Brain Iron Metabolism and CNS Diseases; Chang, Y.-Z., Ed.; Advances in Experimental Medicine and Biology; Springer: Singapore, 2019; pp. 21–32. ISBN 9789811395895. [Google Scholar]
- Anderson, G.J.; Frazer, D.M. Current Understanding of Iron Homeostasis. Am. J. Clin. Nutr. 2017, 106, 1559S–1566S. [Google Scholar] [CrossRef] [Green Version]
- Bou-Abdallah, F.; Paliakkara, J.J.; Melman, G.; Melman, A. Reductive Mobilization of Iron from Intact Ferritin: Mechanisms and Physiological Implication. Pharmaceuticals 2018, 11, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damasceno, F.C.; Condeles, A.L.; Lopes, A.K.B.; Facci, R.R.; Linares, E.; Truzzi, D.R.; Augusto, O.; Toledo, J.C. The Labile Iron Pool Attenuates Peroxynitrite-Dependent Damage and Can No Longer Be Considered Solely a pro-Oxidative Cellular Iron Source. J. Biol. Chem. 2018, 293, 8530–8542. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Ardehali, H.; Min, J.; Wang, F. The Molecular and Metabolic Landscape of Iron and Ferroptosis in Cardiovascular Disease. Nat. Rev. Cardiol 2022, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Role of Iron in Carcinogenesis: Cancer as a Ferrotoxic Disease. Cancer Sci. 2009, 100, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Wood, J.C. Use of Magnetic Resonance Imaging to Monitor Iron Overload. Hematol. Oncol. Clin. N. Am. 2014, 28, 747–764. [Google Scholar] [CrossRef] [PubMed]
- Rios, J.J.; Carrasco-Gil, S.; Abadía, A.; Abadía, J. Using Perls Staining to Trace the Iron Uptake Pathway in Leaves of a Prunus Rootstock Treated with Iron Foliar Fertilizers. Front. Plant Sci. 2016, 7, 893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.; Zhang, Q.; Dai, J.; Huang, X. Calcein as a Fluorescent Iron Chemosensor for the Determination of Low Molecular Weight Iron in Biological Fluids. Biometals 2003, 16, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Petrat, F.; Rauen, U.; de Groot, H. Determination of the Chelatable Iron Pool of Isolated Rat Hepatocytes by Digital Fluorescence Microscopy Using the Fluorescent Probe, Phen Green SK. Hepatology 1999, 29, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- Hirayama, T.; Okuda, K.; Nagasawa, H. A Highly Selective Turn-on Fluorescent Probe for Iron(II) to Visualize Labile Iron in Living Cells. Chem. Sci. 2013, 4, 1250–1256. [Google Scholar] [CrossRef]
- Hirayama, T. Development of Chemical Tools for Imaging of Fe(II) Ions in Living Cells: A Review. Acta Histochem. Cytochem. 2018, 51, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Aron, A.T.; Heffern, M.C.; Lonergan, Z.R.; Vander Wal, M.N.; Blank, B.R.; Spangler, B.; Zhang, Y.; Park, H.M.; Stahl, A.; Renslo, A.R.; et al. In Vivo Bioluminescence Imaging of Labile Iron Accumulation in a Murine Model of Acinetobacter Baumannii Infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12669–12674. [Google Scholar] [CrossRef] [Green Version]
- Eroglu, E.; Gottschalk, B.; Charoensin, S.; Blass, S.; Bischof, H.; Rost, R.; Madreiter-Sokolowski, C.T.; Pelzmann, B.; Bernhart, E.; Sattler, W.; et al. Development of Novel FP-Based Probes for Live-Cell Imaging of Nitric Oxide Dynamics. Nat. Commun. 2016, 7, 10623. [Google Scholar] [CrossRef] [Green Version]
- Eroglu, E.; Bischof, H.; Charoensin, S.; Waldeck-Weiermaier, M.; Graier, W.F.; Malli, R. Real-Time Imaging of Nitric Oxide Signals in Individual Cells Using GeNOps. Methods Mol. Biol. 2018, 1747, 23–34. [Google Scholar] [CrossRef]
- Eroglu, E.; Rost, R.; Bischof, H.; Blass, S.; Schreilechner, A.; Gottschalk, B.; Depaoli, M.R.; Klec, C.; Charoensin, S.; Madreiter-Sokolowski, C.T.; et al. Application of Genetically Encoded Fluorescent Nitric Oxide (NO•) Probes, the GeNOps, for Real-Time Imaging of NO• Signals in Single Cells. J. Vis. Exp. 2017, 121, e55486. [Google Scholar] [CrossRef] [Green Version]
- Eroglu, E.; Hallström, S.; Bischof, H.; Opelt, M.; Schmidt, K.; Mayer, B.; Waldeck-Weiermair, M.; Graier, W.F.; Malli, R. Real-Time Visualization of Distinct Nitric Oxide Generation of Nitric Oxide Synthase Isoforms in Single Cells. Nitric Oxide 2017, 70, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, E.; Charoensin, S.; Bischof, H.; Ramadani, J.; Gottschalk, B.; Depaoli, M.R.; Waldeck-Weiermair, M.; Graier, W.F.; Malli, R. Genetic Biosensors for Imaging Nitric Oxide in Single Cells. Free. Radic. Biol. Med. 2018, 128, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, E.; Saravi, S.S.S.; Sorrentino, A.; Steinhorn, B.; Michel, T. Discordance between ENOS Phosphorylation and Activation Revealed by Multispectral Imaging and Chemogenetic Methods. Proc. Natl. Acad. Sci. USA 2019, 116, 20210–20217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sevimli, G.; Smith, M.J.; Caglar, T.A.; Bilir, Ş.; Secilmis, M.; Altun, H.Y.; Yigit, E.N.; Yang, F.; Keeley, T.P.; Malli, R.; et al. Nitric Oxide Biosensor Uncovers Diminished Ferrous Iron-Dependency of Cultured Cells Adapted to Physiological Oxygen Levels. Redox Biol. 2022, 53, 102319. [Google Scholar] [CrossRef] [PubMed]
- D’Autréaux, B.; Tucker, N.P.; Dixon, R.; Spiro, S. A Non-Haem Iron Centre in the Transcription Factor NorR Senses Nitric Oxide. Nature 2005, 437, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Macdougall, I.C.; Comin-Colet, J.; Breymann, C.; Spahn, D.R.; Koutroubakis, I.E. Iron Sucrose: A Wealth of Experience in Treating Iron Deficiency. Adv. Ther. 2020, 37, 1960–2002. [Google Scholar] [CrossRef] [Green Version]
- Scott, L.J. Ferric Carboxymaltose: A Review in Iron Deficiency. Drugs 2018, 78, 479–493. [Google Scholar] [CrossRef]
- Secilmis, M.; Altun, H.Y.; Pilic, J.; Erdogan, Y.C.; Cokluk, Z.; Ata, B.N.; Sevimli, G.; Zaki, A.G.; Yigit, E.N.; Öztürk, G.; et al. A Co-Culture-Based Multiparametric Imaging Technique to Dissect Local H2O2 Signals with Targeted HyPer7. Biosensors 2021, 11, 338. [Google Scholar] [CrossRef]
- Charoensin, S.; Eroglu, E.; Opelt, M.; Bischof, H.; Madreiter-Sokolowski, C.T.; Kirsch, A.; Depaoli, M.R.; Frank, S.; Schrammel, A.; Mayer, B.; et al. Intact Mitochondrial Ca2+ Uniport Is Essential for Agonist-Induced Activation of Endothelial Nitric Oxide Synthase (ENOS). Free Radic. Biol. Med. 2017, 102, 248–259. [Google Scholar] [CrossRef]
- Opelt, M.; Eroglu, E.; Waldeck-Weiermair, M.; Russwurm, M.; Koesling, D.; Malli, R.; Graier, W.F.; Fassett, J.T.; Schrammel, A.; Mayer, B. Formation of Nitric Oxide by Aldehyde Dehydrogenase-2 Is Necessary and Sufficient for Vascular Bioactivation of Nitroglycerin *. J. Biol. Chem. 2016, 291, 24076–24084. [Google Scholar] [CrossRef] [Green Version]
- Opelt, M.; Wölkart, G.; Eroglu, E.; Waldeck-Weiermair, M.; Malli, R.; Graier, W.F.; Kollau, A.; Fassett, J.T.; Schrammel, A.; Mayer, B.; et al. Sustained Formation of Nitroglycerin-Derived Nitric Oxide by Aldehyde Dehydrogenase-2 in Vascular Smooth Muscle without Added Reductants: Implications for the Development of Nitrate Tolerance. Mol. Pharmacol. 2018, 93, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, R.J.; Wiegand, J.; Brittenham, G.M. HBED: The Continuing Development of a Potential Alternative to Deferoxamine for Iron-Chelating Therapy. Blood 1999, 93, 370–375. [Google Scholar] [CrossRef] [PubMed]
- Möller, M.N.; Denicola, A. Diffusion of Nitric Oxide and Oxygen in Lipoproteins and Membranes Studied by Pyrene Fluorescence Quenching. Free Radic. Biol. Med. 2018, 128, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Kakuta, K.; Orino, K.; Yamamoto, S.; Watanabe, K. High Levels of Ferritin and Its Iron in Fetal Bovine Serum. Comp. Biochem. Physiol. A Physiol. 1997, 118, 165–169. [Google Scholar] [CrossRef]
- Spasojević, I. What If Cell Culture Media Do Not Mimic in Vivo Redox Settings? Redox Rep. 2016, 21, 127–129. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Qiu, Y.; Wang, M.; Zhang, C.; Zhang, T.; Zhou, H.; Zhao, W.; Zhao, W.; Xia, G.; Shao, R. Endocytosis and Organelle Targeting of Nanomedicines in Cancer Therapy. Int. J. Nanomed. 2020, 15, 9447–9467. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sevimli, G.; Alston, A.E.; Funk, F.; Flühmann, B.; Malli, R.; Graier, W.F.; Eroglu, E. Probing Subcellular Iron Availability with Genetically Encoded Nitric Oxide Biosensors. Biosensors 2022, 12, 903. https://doi.org/10.3390/bios12100903
Sevimli G, Alston AE, Funk F, Flühmann B, Malli R, Graier WF, Eroglu E. Probing Subcellular Iron Availability with Genetically Encoded Nitric Oxide Biosensors. Biosensors. 2022; 12(10):903. https://doi.org/10.3390/bios12100903
Chicago/Turabian StyleSevimli, Gulsah, Amy E. Alston, Felix Funk, Beat Flühmann, Roland Malli, Wolfgang F. Graier, and Emrah Eroglu. 2022. "Probing Subcellular Iron Availability with Genetically Encoded Nitric Oxide Biosensors" Biosensors 12, no. 10: 903. https://doi.org/10.3390/bios12100903