
Time-Course of Transcriptomic Change in the Lungs of F344 Rats Repeatedly Exposed to a Multiwalled Carbon Nanotube in a 2-Year Test

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. MWCNT
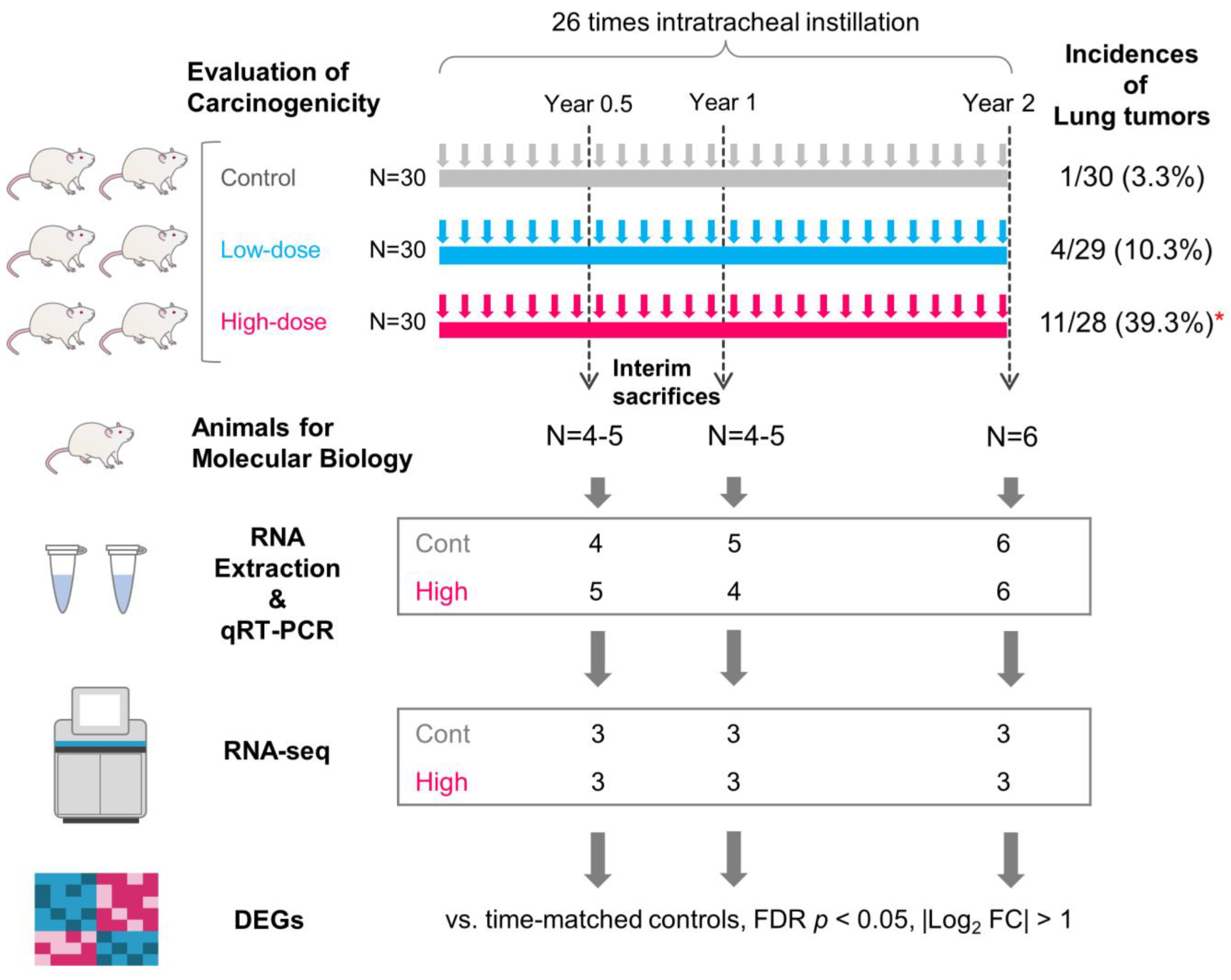
2.2. Animals and Experimental Design
2.3. RNA Preparation and Sequencing
2.4. Data Analysis
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Histopathology
2.7. Statistics
3. Results
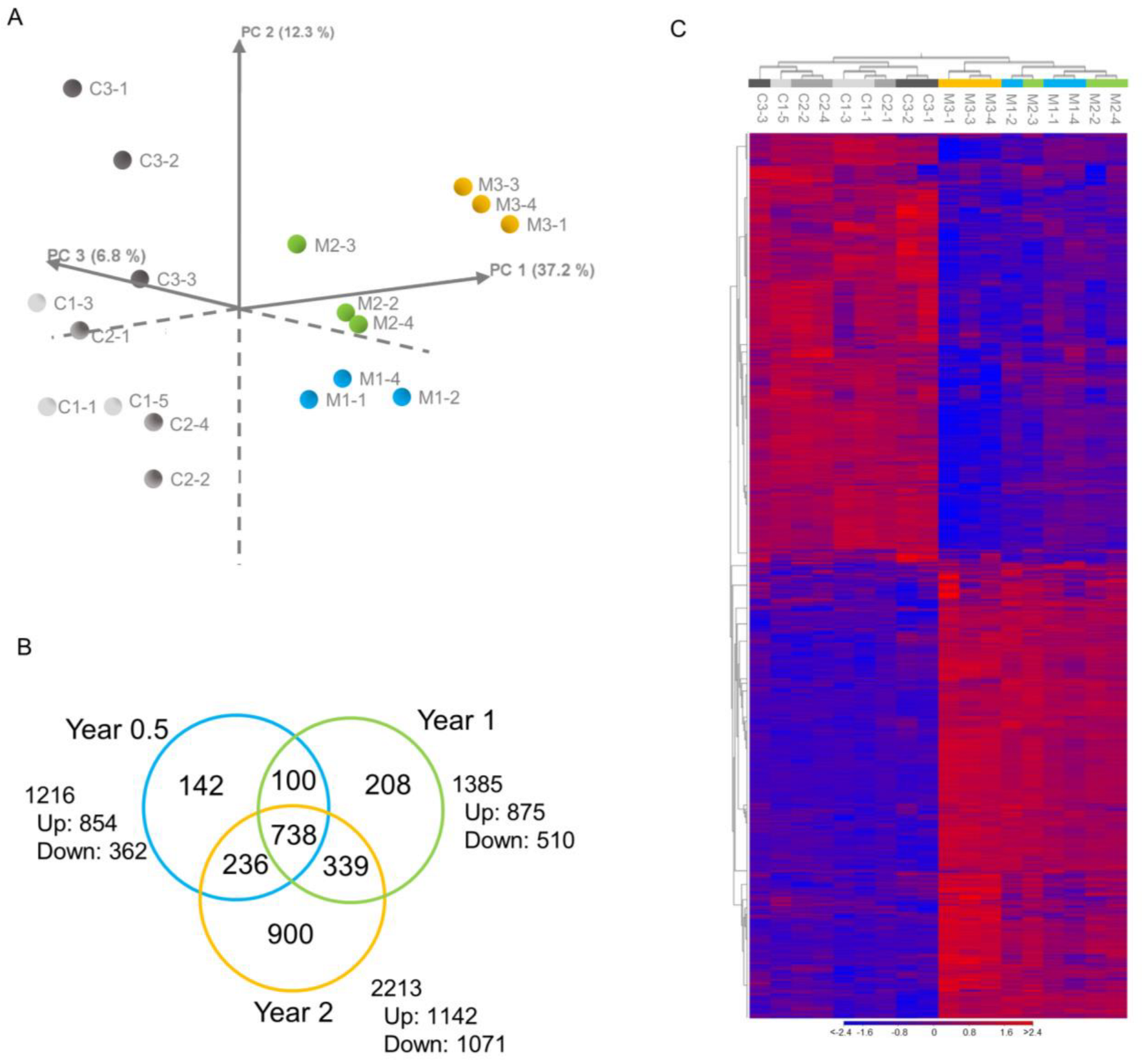
3.1. Identification of Significantly Differentially Expressed Genes (DEGs)
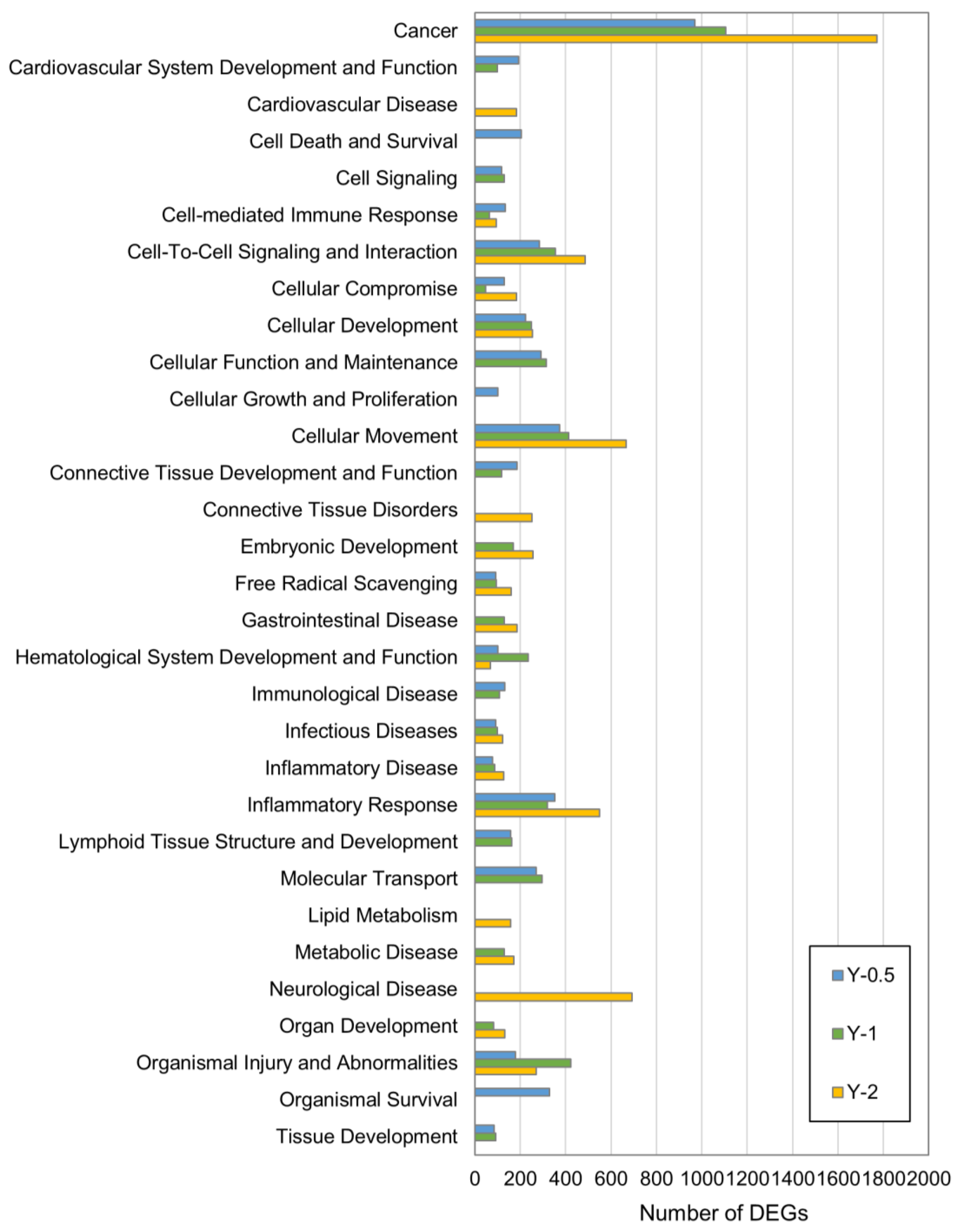
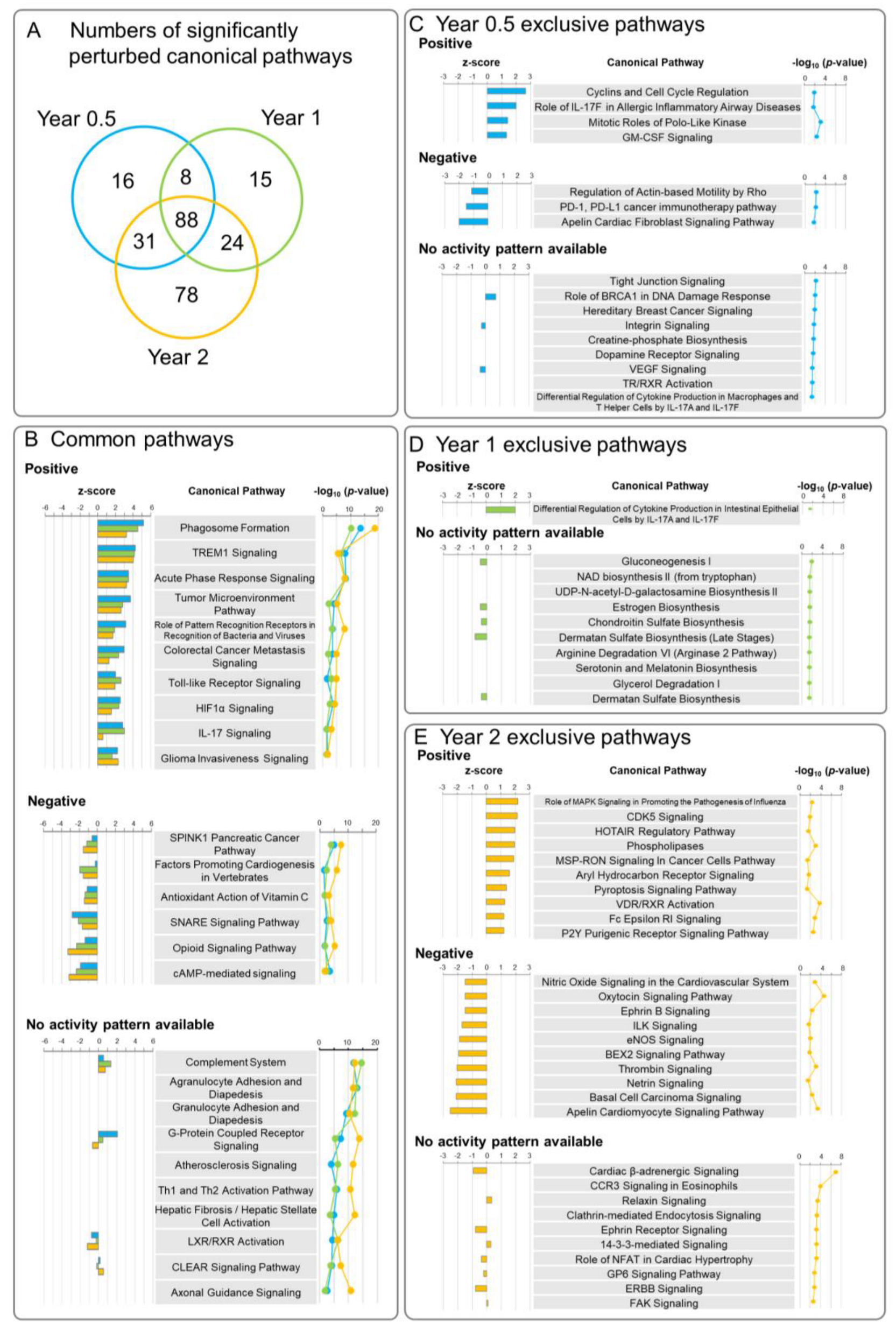
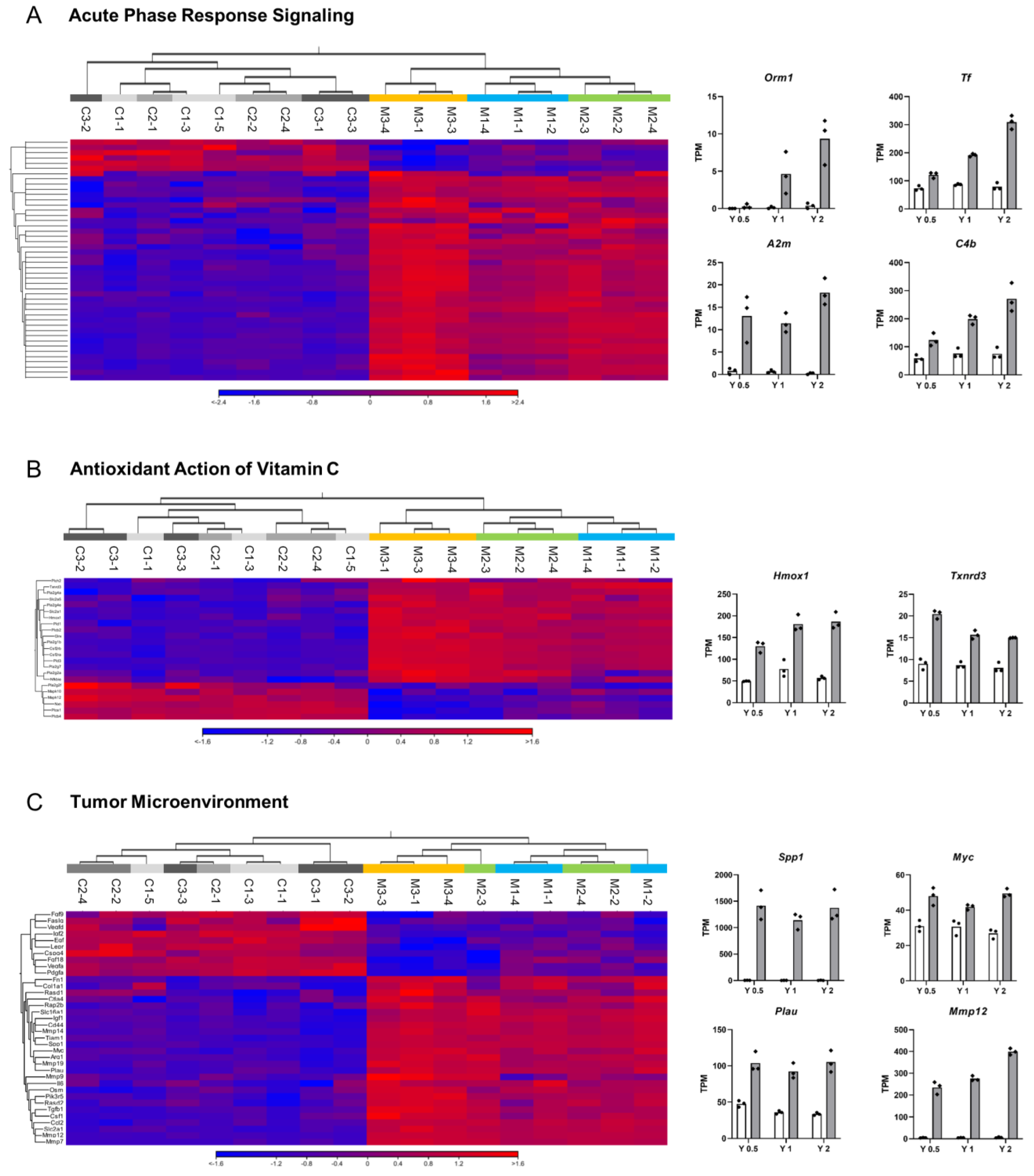
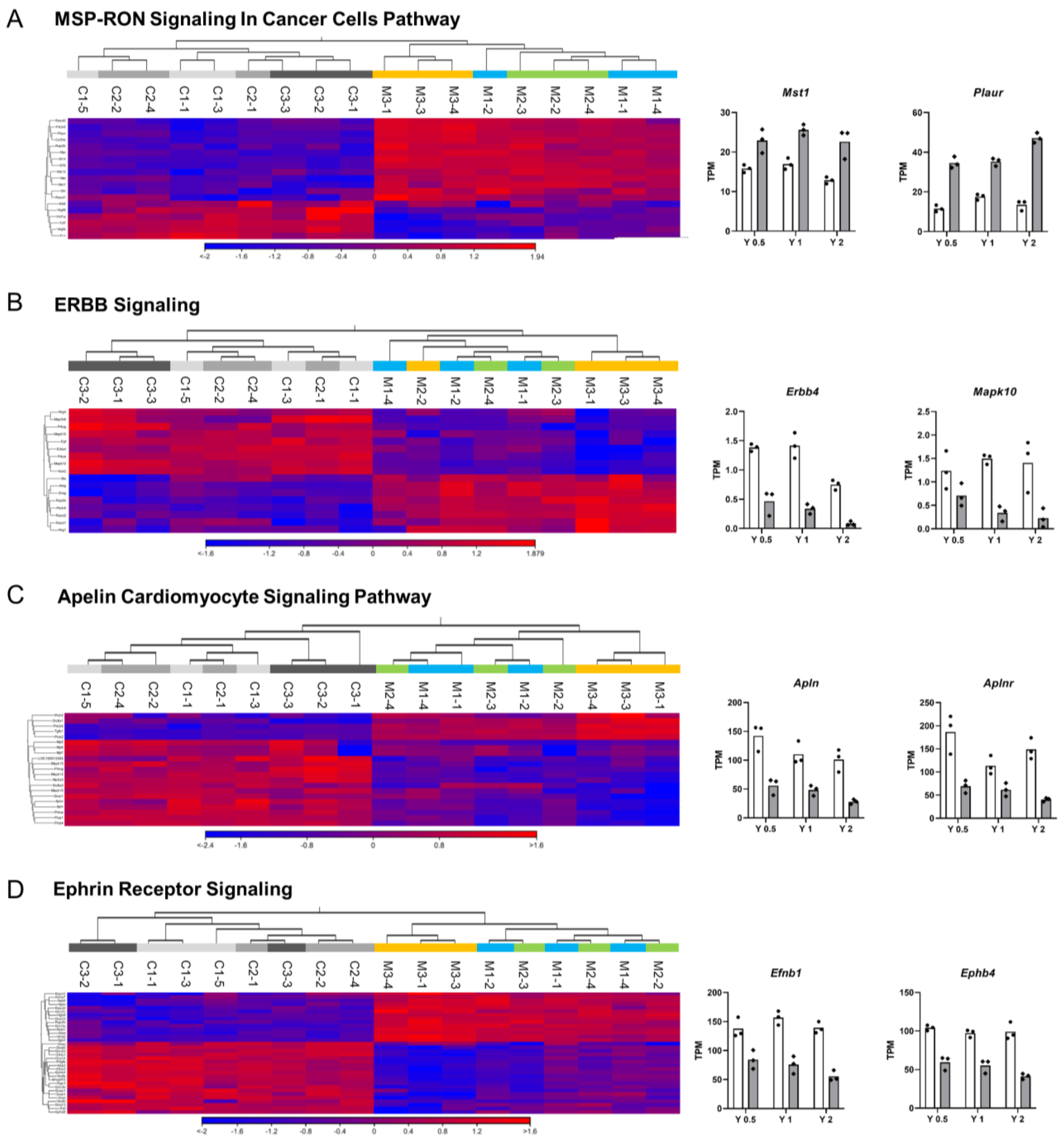
3.2. Ingenuity Pathway Analysis (IPA)
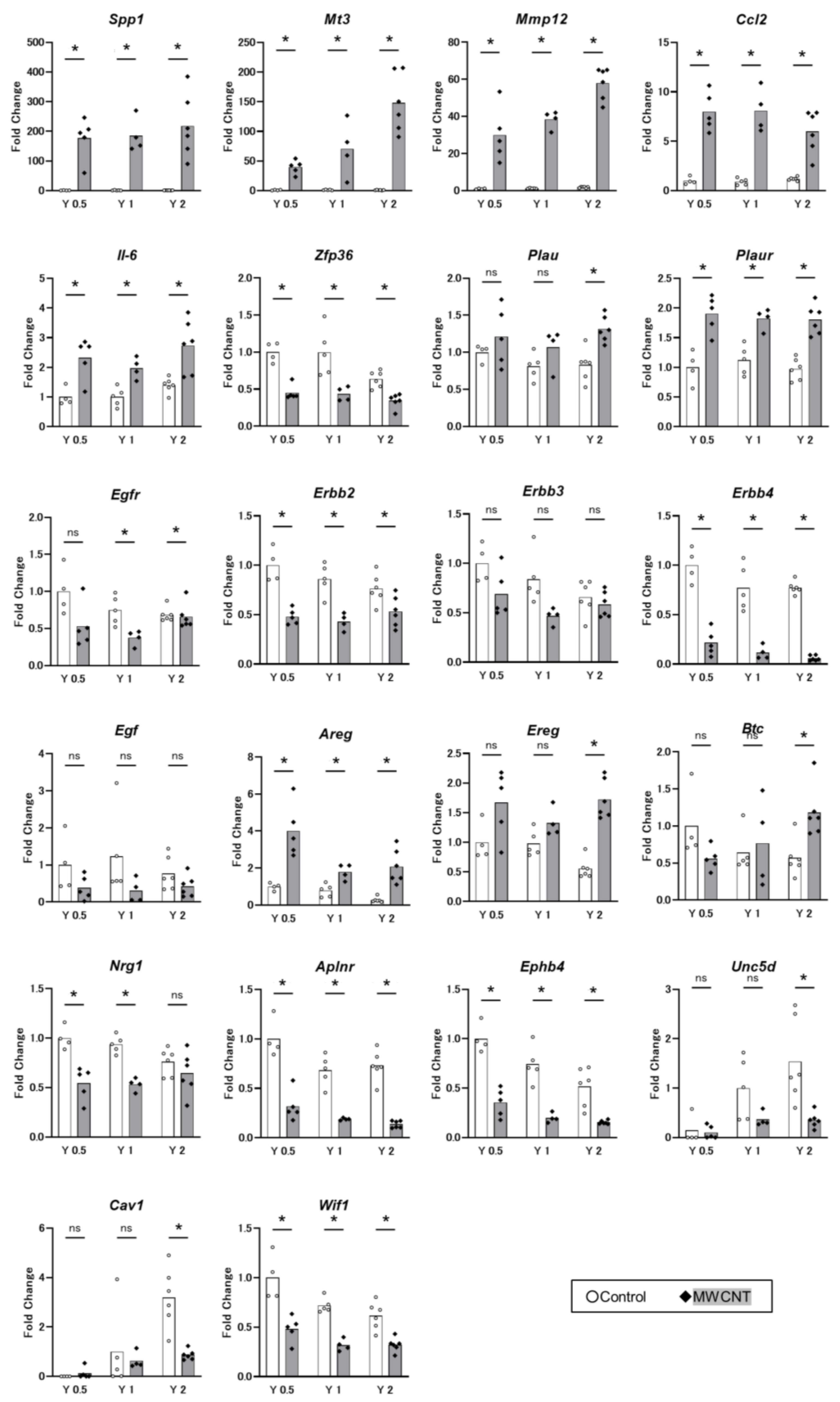
3.3. qRT-PCR
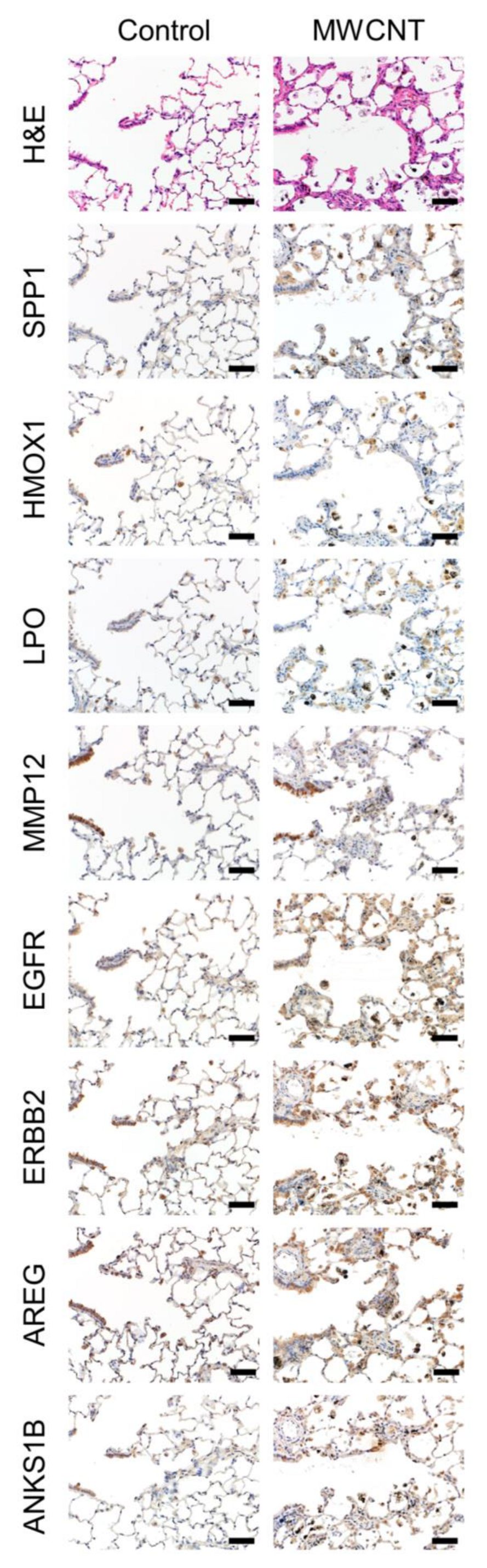
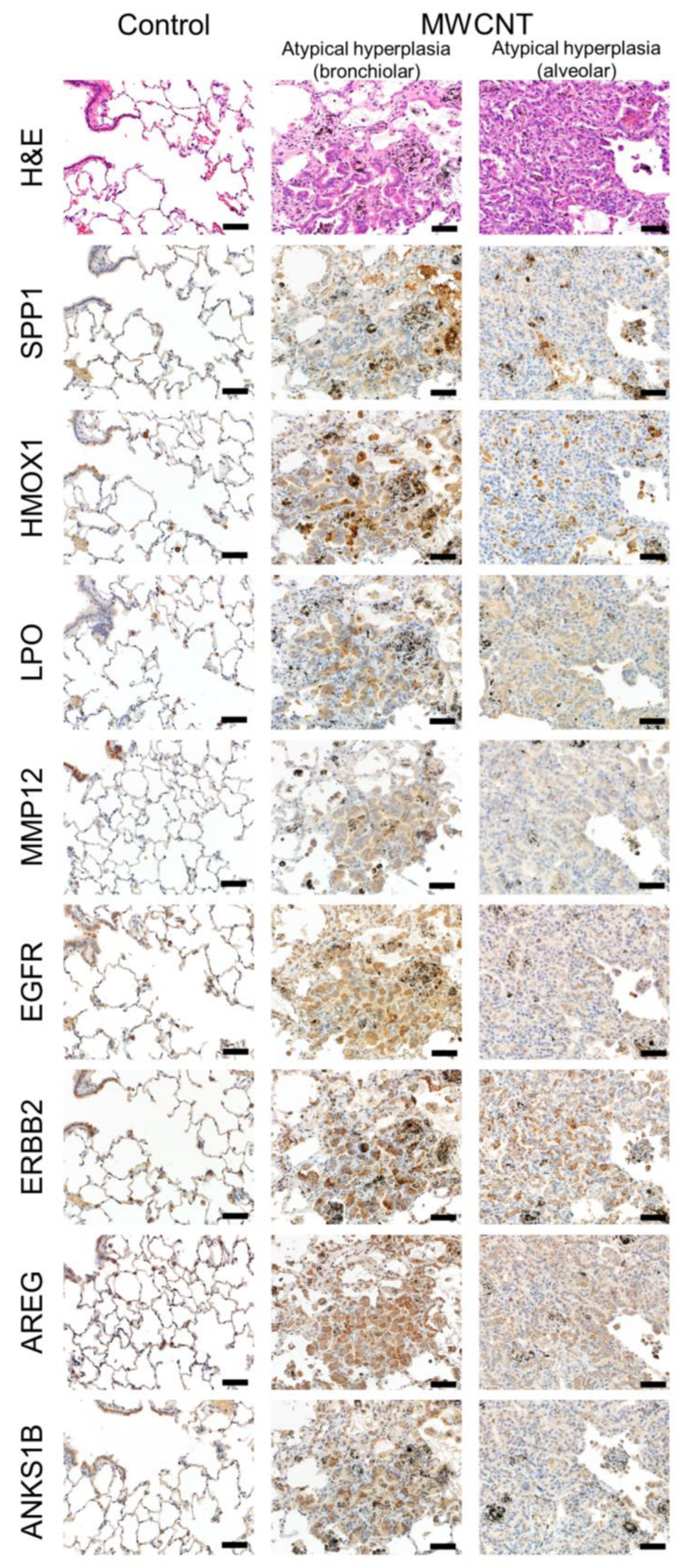
3.4. Immunohistochemistry
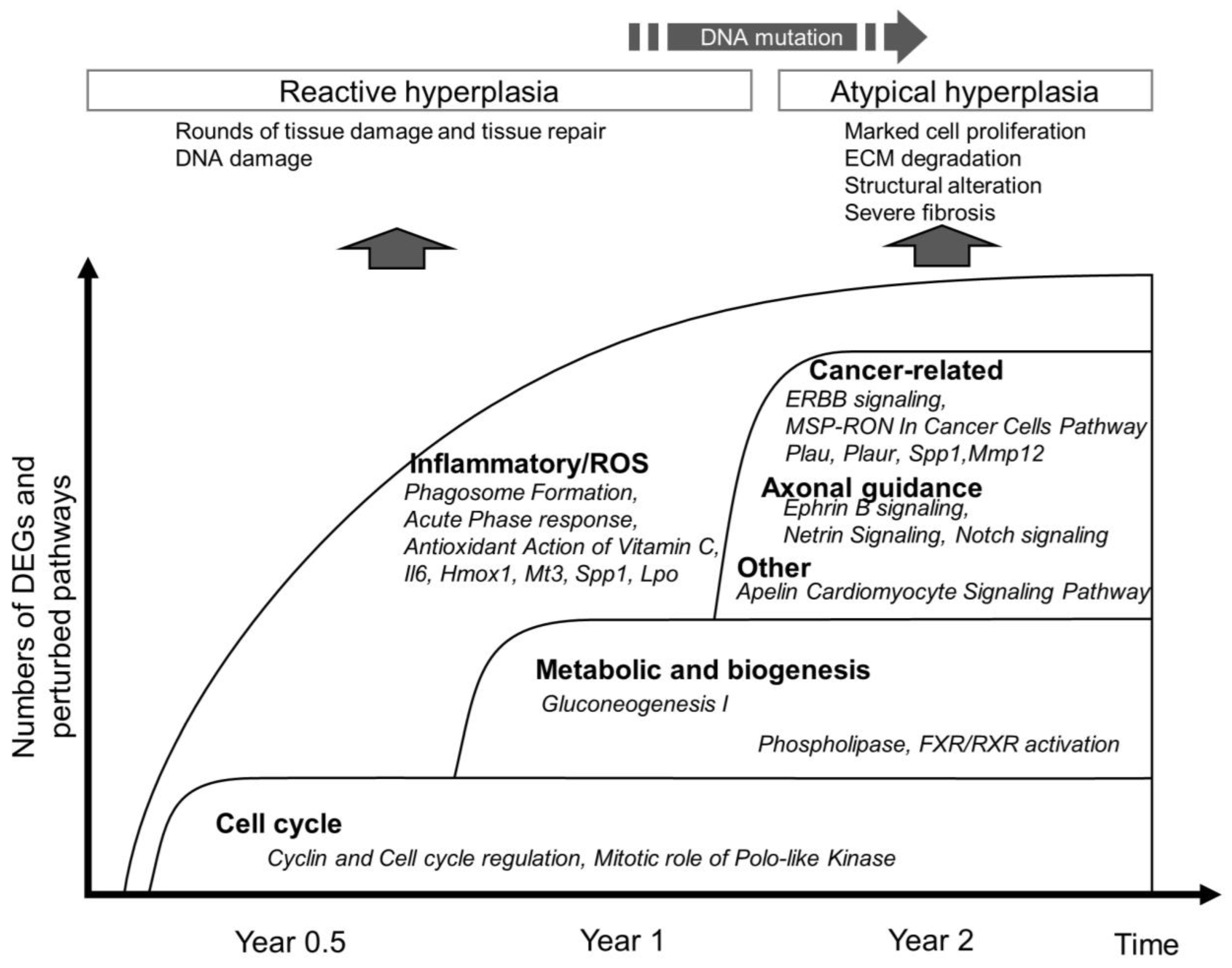
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kuempel, E.D.; Jaurand, M.C.; Moller, P.; Morimoto, Y.; Kobayashi, N.; Pinkerton, K.E.; Sargent, L.M.; Vermeulen, R.C.; Fubini, B.; Kane, A.B. Evaluating the mechanistic evidence and key data gaps in assessing the potential carcinogenicity of carbon nanotubes and nanofibers in humans. Crit. Rev. Toxicol. 2017, 47, 1–58. [Google Scholar] [CrossRef] [Green Version]
- Oberdörster, G.; Kuhlbusch, T.A.J. In vivo effects: Methodologies and biokinetics of inhaled nanomaterials. NanoImpact 2018, 10, 38–60. [Google Scholar] [CrossRef]
- Yuan, X.; Zhang, X.; Sun, L.; Wei, Y.; Wei, X. Cellular Toxicity and Immunological Effects of Carbon-based Nanomaterials. Part. Fibre Toxicol. 2019, 16, 18. [Google Scholar] [CrossRef]
- Moller, P.; Wils, R.S.; Di Ianni, E.; Gutierrez, C.A.T.; Roursgaard, M.; Jacobsen, N.R. Genotoxicity of multi-walled carbon nanotube reference materials in mammalian cells and animals. Mutat. Res. Rev. Mutat. Res. 2021, 788, 108393. [Google Scholar] [CrossRef] [PubMed]
- Witkowska, M.; Florek, E.; Mrowczynski, R. Assessment of Pristine Carbon Nanotubes Toxicity in Rodent Models. Int. J. Mol. Sci. 2022, 23, 15343. [Google Scholar] [CrossRef]
- Kane, A.B.; Hurt, R.H.; Gao, H. The asbestos-carbon nanotube analogy: An update. Toxicol. Appl. Pharmacol. 2018, 361, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, K.; Murphy, F.A.; Duffin, R.; Poland, C.A. Asbestos, carbon nanotubes and the pleural mesothelium: A review of the hypothesis regarding the role of long fibre retention in the parietal pleura, inflammation and mesothelioma. Part. Fibre. Toxicol. 2010, 7, 5. [Google Scholar] [CrossRef] [Green Version]
- Takagi, A.; Hirose, A.; Nishimura, T.; Fukumori, N.; Ogata, A.; Ohashi, N.; Kitajima, S.; Kanno, J. Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci. 2008, 33, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Nakae, D.; Fukumori, N.; Tayama, K.; Maekawa, A.; Imai, K.; Hirose, A.; Nishimura, T.; Ohashi, N.; Ogata, A. Induction of mesothelioma by a single intrascrotal administration of multi-wall carbon nanotube in intact male Fischer 344 rats. J. Toxicol. Sci. 2009, 34, 65–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagai, H.; Okazaki, Y.; Chew, S.H.; Misawa, N.; Yamashita, Y.; Akatsuka, S.; Ishihara, T.; Yamashita, K.; Yoshikawa, Y.; Yasui, H.; et al. Diameter and rigidity of multiwalled carbon nanotubes are critical factors in mesothelial injury and carcinogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1330–E1338. [Google Scholar] [CrossRef]
- Takagi, A.; Hirose, A.; Futakuchi, M.; Tsuda, H.; Kanno, J. Dose-dependent mesothelioma induction by intraperitoneal administration of multi-wall carbon nanotubes in p53 heterozygous mice. Cancer Sci. 2012, 103, 1440–1444. [Google Scholar] [CrossRef]
- Sargent, L.M.; Porter, D.W.; Staska, L.M.; Hubbs, A.F.; Lowry, D.T.; Battelli, L.; Siegrist, K.J.; Kashon, M.L.; Mercer, R.R.; Bauer, A.K.; et al. Promotion of lung adenocarcinoma following inhalation exposure to multi-walled carbon nanotubes. Part. Fibre Toxicol. 2014, 11, 3. [Google Scholar] [CrossRef] [Green Version]
- Grosse, Y.; Loomis, D.; Guyton, K.Z.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; et al. Carcinogenicity of fluoro-edenite, silicon carbide fibres and whiskers, and carbon nanotubes. Lancet Oncol. 2014, 15, 1427–1428. [Google Scholar] [CrossRef] [PubMed]
- Kasai, T.; Umeda, Y.; Ohnishi, M.; Mine, T.; Kondo, H.; Takeuchi, T.; Matsumoto, M.; Fukushima, S. Lung carcinogenicity of inhaled multi-walled carbon nanotube in rats. Part. Fibre Toxicol. 2016, 13, 53. [Google Scholar] [CrossRef] [Green Version]
- Snyder-Talkington, B.N.; Dymacek, J.; Porter, D.W.; Wolfarth, M.G.; Mercer, R.R.; Pacurari, M.; Denvir, J.; Castranova, V.; Qian, Y.; Guo, N.L. System-based identification of toxicity pathways associated with multi-walled carbon nanotube-induced pathological responses. Toxicol. Appl. Pharmacol. 2013, 272, 476–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, S.S.; Saber, A.T.; Williams, A.; Andersen, O.; Kobler, C.; Atluri, R.; Pozzebon, M.E.; Mucelli, S.P.; Simion, M.; Rickerby, D.; et al. MWCNTs of different physicochemical properties cause similar inflammatory responses, but differences in transcriptional and histological markers of fibrosis in mouse lungs. Toxicol. Appl. Pharmacol. 2015, 284, 16–32. [Google Scholar] [CrossRef]
- Fujita, K.; Fukuda, M.; Fukui, H.; Horie, M.; Endoh, S.; Uchida, K.; Shichiri, M.; Morimoto, Y.; Ogami, A.; Iwahashi, H. Intratracheal instillation of single-wall carbon nanotubes in the rat lung induces time-dependent changes in gene expression. Nanotoxicology 2015, 9, 290–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaliullin, T.O.; Yanamala, N.; Newman, M.S.; Kisin, E.R.; Fatkhutdinova, L.M.; Shvedova, A.A. Comparative analysis of lung and blood transcriptomes in mice exposed to multi-walled carbon nanotubes. Toxicol. Appl. Pharmacol. 2020, 390, 114898. [Google Scholar] [CrossRef]
- Guo, N.L.; Wan, Y.W.; Denvir, J.; Porter, D.W.; Pacurari, M.; Wolfarth, M.G.; Castranova, V.; Qian, Y. Multiwalled carbon nanotube-induced gene signatures in the mouse lung: Potential predictive value for human lung cancer risk and prognosis. J. Toxicol. Environ. Health A 2012, 75, 1129–1153. [Google Scholar] [CrossRef] [Green Version]
- Snyder-Talkington, B.N.; Dong, C.; Singh, S.; Raese, R.; Qian, Y.; Porter, D.W.; Wolfarth, M.G.; Guo, N.L. Multi-Walled Carbon Nanotube-Induced Gene Expression Biomarkers for Medical and Occupational Surveillance. Int. J. Mol. Sci. 2019, 20, 2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shvedova, A.A.; Yanamala, N.; Kisin, E.R.; Khailullin, T.O.; Birch, M.E.; Fatkhutdinova, L.M. Integrated Analysis of Dysregulated ncRNA and mRNA Expression Profiles in Humans Exposed to Carbon Nanotubes. PLoS ONE 2016, 11, e0150628. [Google Scholar] [CrossRef] [PubMed]
- Solorio-Rodriguez, S.A.; Williams, A.; Poulsen, S.S.; Knudsen, K.B.; Jensen, K.A.; Clausen, P.A.; Danielsen, P.H.; Wallin, H.; Vogel, U.; Halappanavar, S. Single-Walled vs. Multi-Walled Carbon Nanotubes: Influence of Physico-Chemical Properties on Toxicogenomics Responses in Mouse Lungs. Nanomaterials 2023, 13, 1059. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Obara, S.; Maru, J.; Endoh, S. Cytotoxicity profiles of multi-walled carbon nanotubes with different physico-chemical properties. Toxicol. Mech. Methods 2020, 30, 477–489. [Google Scholar] [CrossRef] [PubMed]
- Seidel, C.; Zhernovkov, V.; Cassidy, H.; Kholodenko, B.; Matallanas, D.; Cosnier, F.; Gate, L. Inhaled multi-walled carbon nanotubes differently modulate global gene and protein expression in rat lungs. Nanotoxicology 2021, 15, 238–256. [Google Scholar] [CrossRef]
- Tsuda, H.; Alexander, D.B. Development of Intratracheal Intrapulmonary Spraying (TIPS) Administration as a Feasible Assay Method for Testing the Toxicity and Carcinogenic Potential of Multiwall Carbon Nanotubes. In In Vivo Inhalation Toxicity Screening Methods for Manufactured Nanomaterials; Takebayashi, T., Landsiedel, R., Gamo, M., Eds.; Springer: Cham, Switzerland, 2019; pp. 145–163. [Google Scholar]
- Suzui, M.; Futakuchi, M.; Fukamachi, K.; Numano, T.; Abdelgied, M.; Takahashi, S.; Ohnishi, M.; Omori, T.; Tsuruoka, S.; Hirose, A.; et al. Multiwalled carbon nanotubes intratracheally instilled into the rat lung induce development of pleural malignant mesothelioma and lung tumors. Cancer Sci. 2016, 107, 924–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, D.M.; Alexander, W.T.; Numano, T.; Ahmed, O.H.M.; Gunasekaran, S.; Alexander, D.B.; Abdelgied, M.; El-Gazzar, A.M.; Takase, H.; Xu, J.; et al. Comparative carcinogenicity study of a thick, straight-type and a thin, tangled-type multi-walled carbon nanotube administered by intra-tracheal instillation in the rat. Part. Fibre Toxicol. 2020, 17, 48. [Google Scholar] [CrossRef]
- Saleh, D.M.; Luo, S.; Ahmed, O.H.M.; Alexander, D.B.; Alexander, W.T.; Gunasekaran, S.; El-Gazzar, A.M.; Abdelgied, M.; Numano, T.; Takase, H.; et al. Assessment of the toxicity and carcinogenicity of double-walled carbon nanotubes in the rat lung after intratracheal instillation: A two-year study. Part. Fibre Toxicol. 2022, 19, 30. [Google Scholar] [CrossRef]
- Hojo, M.; Maeno, A.; Sakamoto, Y.; Ohnuki, A.; Tada, Y.; Yamamoto, Y.; Ikushima, K.; Inaba, R.; Suzuki, J.; Taquahashi, Y.; et al. Two-year intermittent exposure of a multiwalled carbon nanotube by intratracheal instillation induces lung tumors and pleural mesotheliomas in F344 rats. Part. Fibre Toxicol. 2022, 19, 38. [Google Scholar] [CrossRef]
- Taquahashi, Y.; Ogawa, Y.; Takagi, A.; Tsuji, M.; Morita, K.; Kanno, J. Improved dispersion method of multi-wall carbon nanotube for inhalation toxicity studies of experimental animals. J. Toxicol. Sci. 2013, 38, 619–628. [Google Scholar] [CrossRef] [Green Version]
- Sakamoto, Y.; Hojo, M.; Kosugi, Y.; Watanabe, K.; Hirose, A.; Inomata, A.; Suzuki, T.; Nakae, D. Comparative study for carcinogenicity of 7 different multi-wall carbon nanotubes with different physicochemical characteristics by a single intraperitoneal injection in male Fischer 344 rats. J. Toxicol. Sci. 2018, 43, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Percie du Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. J. Cereb. Blood Flow Metab. 2020, 40, 1769–1777. [Google Scholar] [CrossRef] [PubMed]
- Huaux, F.; d’Ursel de Bousies, V.; Parent, M.A.; Orsi, M.; Uwambayinema, F.; Devosse, R.; Ibouraadaten, S.; Yakoub, Y.; Panin, N.; Palmai-Pallag, M.; et al. Mesothelioma response to carbon nanotubes is associated with an early and selective accumulation of immunosuppressive monocytic cells. Part. Fibre Toxicol. 2016, 13, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Numano, T.; Higuchi, H.; Alexander, D.B.; Alexander, W.T.; Abdelgied, M.; El-Gazzar, A.M.; Saleh, D.; Takase, H.; Hirose, A.; Naiki-Ito, A.; et al. MWCNT-7 administered to the lung by intratracheal instillation induces development of pleural mesothelioma in F344 rats. Cancer Sci. 2019, 110, 2485–2492. [Google Scholar] [CrossRef]
- Kato, T.; Totsuka, Y.; Ishino, K.; Matsumoto, Y.; Tada, Y.; Nakae, D.; Goto, S.; Masuda, S.; Ogo, S.; Kawanishi, M.; et al. Genotoxicity of multi-walled carbon nanotubes in both in vitro and in vivo assay systems. Nanotoxicology 2013, 7, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Tavares, A.M.; Louro, H.; Antunes, S.; Quarre, S.; Simar, S.; De Temmerman, P.J.; Verleysen, E.; Mast, J.; Jensen, K.A.; Norppa, H.; et al. Genotoxicity evaluation of nanosized titanium dioxide, synthetic amorphous silica and multi-walled carbon nanotubes in human lymphocytes. Toxicol In Vitro 2014, 28, 60–69. [Google Scholar] [CrossRef]
- Horibata, K.; Takasawa, H.; Hojo, M.; Taquahashi, Y.; Shigano, M.; Yokota, S.; Kobayashi, N.; Sugiyama, K.I.; Honma, M.; Hamada, S. In vivo genotoxicity assessment of a multiwalled carbon nanotube in a mouse ex vivo culture. Genes Environ. 2022, 44, 24. [Google Scholar] [CrossRef]
- Sager, T.M.; Umbright, C.M.; Mustafa, G.M.; Roberts, J.R.; Orandle, M.S.; Cumpston, J.L.; McKinney, W.G.; Boots, T.; Kashon, M.L.; Joseph, P. Pulmonary toxicity and gene expression changes in response to whole-body inhalation exposure to multi-walled carbon nanotubes in rats. Inhal. Toxicol. 2022, 34, 200–218. [Google Scholar] [CrossRef]
- Søs Poulsen, S.; Jacobsen, N.R.; Labib, S.; Wu, D.; Husain, M.; Williams, A.; Bøgelund, J.P.; Andersen, O.; Købler, C.; Mølhave, K.; et al. Transcriptomic analysis reveals novel mechanistic insight into murine biological responses to multi-walled carbon nanotubes in lungs and cultured lung epithelial cells. PLoS ONE 2013, 8, e80452. [Google Scholar] [CrossRef] [Green Version]
- Kinaret, P.; Ilves, M.; Fortino, V.; Rydman, E.; Karisola, P.; Lahde, A.; Koivisto, J.; Jokiniemi, J.; Wolff, H.; Savolainen, K.; et al. Inhalation and oropharyngeal aspiration exposure to rod-like carbon nanotubes induce similar airway inflammation and biological responses in mouse lungs. ACS Nano 2017, 11, 291–303. [Google Scholar] [CrossRef]
- Rahman, L.; Jacobsen, N.R.; Aziz, S.A.; Wu, D.; Williams, A.; Yauk, C.L.; White, P.; Wallin, H.; Vogel, U.; Halappanavar, S. Multi-walled carbon nanotube-induced genotoxic, inflammatory and pro-fibrotic responses in mice: Investigating the mechanisms of pulmonary carcinogenesis. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2017, 823, 28–44. [Google Scholar] [CrossRef] [Green Version]
- Barthel, H.; Sébillaud, S.; Lorcin, M.; Wolff, H.; Viton, S.; Cosnier, F.; Gaté, L.; Seidel, C. Needlelike, short and thin multi-walled carbon nanotubes: Comparison of effects on wild type and p53+/− rat lungs. Nanotoxicology 2023, 17, 270–288. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Wu, W.; Okuhira, K.; Miyazawa, K.; Hattori, T.; Sai, K.; Naito, M.; Suzuki, K.; Nishimura, T.; Sakamoto, Y.; et al. High-temperature calcined fullerene nanowhiskers as well as long needle-like multi-wall carbon nanotubes have abilities to induce NLRP3-mediated IL-1beta secretion. Biochem. Biophys. Res. Commun. 2014, 452, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Rabolli, V.; Lison, D.; Huaux, F. The complex cascade of cellular events governing inflammasome activation and IL-1beta processing in response to inhaled particles. Part. Fibre Toxicol. 2016, 13, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukai, E.; Sato, H.; Watanabe, M.; Nakae, D.; Totsuka, Y. Establishment of an in vivo simulating co-culture assay platform for genotoxicity of multi-walled carbon nanotubes. Cancer Sci. 2018, 109, 1024–1031. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, F.R.; Franklin, W.A.; Gazdar, A.F.; Bunn, P.A., Jr. Early detection of lung cancer: Clinical perspectives of recent advances in biology and radiology. Clin. Cancer Res. 2001, 7, 5–22. [Google Scholar]
- Brambilla, C.; Fievet, F.; Jeanmart, M.; de Fraipont, F.; Lantuejoul, S.; Frappat, V.; Ferretti, G.; Brichon, P.Y.; Moro-Sibilot, D. Early detection of lung cancer: Role of biomarkers. Eur. Respir. J. Suppl. 2003, 39, 36s–44s. [Google Scholar] [CrossRef] [Green Version]
- Blanco, D.; Vicent, S.; Fraga, M.F.; Fernandez-Garcia, I.; Freire, J.; Lujambio, A.; Esteller, M.; Ortiz-de-Solorzano, C.; Pio, R.; Lecanda, F.; et al. Molecular analysis of a multistep lung cancer model induced by chronic inflammation reveals epigenetic regulation of p16 and activation of the DNA damage response pathway. Neoplasia 2007, 9, 840–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishino, K.; Kato, T.; Kato, M.; Shibata, T.; Watanabe, M.; Wakabayashi, K.; Nakagama, H.; Totsuka, Y. Comprehensive DNA adduct analysis reveals pulmonary inflammatory response contributes to genotoxic action of magnetite nanoparticles. Int. J. Mol. Sci. 2015, 16, 3474–3492. [Google Scholar] [CrossRef] [Green Version]
- Totsuka, Y.; Watanabe, M.; Lin, Y. New horizons of DNA adductome for exploring environmental causes of cancer. Cancer Sci. 2021, 112, 7–15. [Google Scholar] [CrossRef]
- Iwashita, Y.; Ohnishi, I.; Matsushita, Y.; Ohtsuka, S.; Yamashita, T.; Inaba, K.; Fukazawa, A.; Ochiai, H.; Matsumoto, K.; Kurono, N.; et al. Geospatial Assessments of DNA Adducts in the Human Stomach: A Model of Field Cancerization. Cancers 2021, 13, 3728. [Google Scholar] [CrossRef]
- Kononenko, V.; Erman, A.; Petan, T.; Krizaj, I.; Kralj, S.; Makovec, D.; Drobne, D. Harmful at non-cytotoxic concentrations: SiO(2)-SPIONs affect surfactant metabolism and lamellar body biogenesis in A549 human alveolar epithelial cells. Nanotoxicology 2017, 11, 419–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, A.; Munari, F.; Sanchez-Rodriguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Jiang, Y.; Luo, Y.; Guo, H.; Huang, C.; Peng, J.; Cao, Y. Multi-walled carbon nanotubes (MWCNTs) transformed THP-1 macrophages into foam cells: Impact of pulmonary surfactant component dipalmitoylphosphatidylcholine. J. Hazard. Mater. 2020, 392, 122286. [Google Scholar] [CrossRef] [PubMed]
- Madison, M.C.; Landers, C.T.; Gu, B.H.; Chang, C.Y.; Tung, H.Y.; You, R.; Hong, M.J.; Baghaei, N.; Song, L.Z.; Porter, P.; et al. Electronic cigarettes disrupt lung lipid homeostasis and innate immunity independent of nicotine. J. Clin. Investig. 2019, 129, 4290–4304. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, F.R.; Varella-Garcia, M.; Bunn, P.A., Jr.; Di Maria, M.V.; Veve, R.; Bremmes, R.M.; Baron, A.E.; Zeng, C.; Franklin, W.A. Epidermal growth factor receptor in non-small-cell lung carcinomas: Correlation between gene copy number and protein expression and impact on prognosis. J. Clin. Oncol. 2003, 21, 3798–3807. [Google Scholar] [CrossRef]
- Hirsch, F.R.; Varella-Garcia, M.; Cappuzzo, F. Predictive value of EGFR and HER2 overexpression in advanced non-small-cell lung cancer. Oncogene 2009, 28 (Suppl. 1), S32–S37. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Research Network. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef]
- Piyathilake, C.J.; Frost, A.R.; Manne, U.; Weiss, H.; Bell, W.C.; Heimburger, D.C.; Grizzle, W.E. Differential expression of growth factors in squamous cell carcinoma and precancerous lesions of the lung. Clin. Cancer Res. 2002, 8, 734–744. [Google Scholar]
- Segers, V.F.M.; Dugaucquier, L.; Feyen, E.; Shakeri, H.; De Keulenaer, G.W. The role of ErbB4 in cancer. Cell. Oncol. 2020, 43, 335–352. [Google Scholar] [CrossRef]
- Huang, L.; Fang, X.; Shi, D.; Yao, S.; Wu, W.; Fang, Q.; Yao, H. MSP-RON Pathway: Potential Regulator of Inflammation and Innate Immunity. Front. Immunol. 2020, 11, 569082. [Google Scholar] [CrossRef]
- Van de Laar, E.; Clifford, M.; Hasenoeder, S.; Kim, B.R.; Wang, D.; Lee, S.; Paterson, J.; Vu, N.M.; Waddell, T.K.; Keshavjee, S.; et al. Cell surface marker profiling of human tracheal basal cells reveals distinct subpopulations, identifies MST1/MSP as a mitogenic signal, and identifies new biomarkers for lung squamous cell carcinomas. Respir. Res. 2014, 15, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camp, E.R.; Liu, W.; Fan, F.; Yang, A.; Somcio, R.; Ellis, L.M. RON, a tyrosine kinase receptor involved in tumor progression and metastasis. Ann. Surg. Oncol. 2005, 12, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Park, J.H.; Khoi, P.N.; Joo, Y.E.; Jung, Y.D. MSP-induced RON activation upregulates uPAR expression and cell invasiveness via MAPK, AP-1 and NF-kappaB signals in gastric cancer cells. Carcinogenesis 2011, 32, 175–181. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Malla, R. New Insight on the Role of Plasminogen Receptor in Cancer Progression. Cancer Growth Metastasis 2015, 8, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasarre, P.; Potiron, V.; Drabkin, H.; Roche, J. Guidance molecules in lung cancer. Cell Adh. Migr. 2010, 4, 130–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, M.; Jun, T.; Nie, Y.; Hao, J.; Fan, D. The Role of the Slit/Robo Signaling Pathway. J. Cancer 2019, 10, 2694–2705. [Google Scholar] [CrossRef] [Green Version]
- Xian, J.; Aitchison, A.; Bobrow, L.; Corbett, G.; Pannell, R.; Rabbitts, T.; Rabbitts, P. Targeted disruption of the 3p12 gene, Dutt1/Robo1, predisposes mice to lung adenocarcinomas and lymphomas with methylation of the gene promoter. Cancer Res. 2004, 64, 6432–6437. [Google Scholar] [CrossRef] [Green Version]
- Xian, J.; Clark, K.J.; Fordham, R.; Pannell, R.; Rabbitts, T.H.; Rabbitts, P.H. Inadequate lung development and bronchial hyperplasia in mice with a targeted deletion in the Dutt1/Robo1 gene. Proc. Natl. Acad. Sci. USA 2001, 98, 15062–15066. [Google Scholar] [CrossRef]
- Gottschling, B.C.; Maronpot, R.R.; Hailey, J.R.; Peddada, S.; Moomaw, C.R.; Klaunig, J.E.; Nyska, A. The role of oxidative stress in indium phosphide-induced lung carcinogenesis in rats. Toxicol. Sci. 2001, 64, 28–40. [Google Scholar] [CrossRef]
- Nagano, K.; Nishizawa, T.; Umeda, Y.; Kasai, T.; Noguchi, T.; Gotoh, K.; Ikawa, N.; Eitaki, Y.; Kawasumi, Y.; Yamauchi, T.; et al. Inhalation carcinogenicity and chronic toxicity of indium-tin oxide in rats and mice. J. Occup. Health 2011, 53, 175–187. [Google Scholar] [CrossRef]
- Huang, S.; Chen, L.; Lu, L.; Li, L. The apelin-APJ axis: A novel potential therapeutic target for organ fibrosis. Clin. Chim. Acta 2016, 456, 81–88. [Google Scholar] [CrossRef]
- Wang, H.; Cong, L.; Yin, X.; Zhang, N.; Zhu, M.; Sun, T.; Fan, J.; Xue, F.; Fan, X.; Gong, Y. The Apelin-APJ axis alleviates LPS-induced pulmonary fibrosis and endothelial mesenchymal transformation in mice by promoting Angiotensin-Converting Enzyme 2. Cell. Signal. 2022, 98, 110418. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Guo, H.; Wang, H.; Xing, D.; Lu, T.; Yang, J.; Wang, C. Apelin-13 alleviated cardiac fibrosis via inhibiting the PI3K/Akt pathway to attenuate oxidative stress in rats with myocardial infarction-induced heart failure. Biosci. Rep. 2020, 40, BSR20200040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarelli, A.V.; Masciale, V.; Aramini, B.; Colo, G.P.; Tonelli, R.; Marchioni, A.; Bruzzi, G.; Gozzi, F.; Andrisani, D.; Castaniere, I.; et al. Molecular Mechanisms and Cellular Contribution from Lung Fibrosis to Lung Cancer Development. Int. J. Mol. Sci. 2021, 22, 12179. [Google Scholar] [CrossRef] [PubMed]
- Lamort, A.S.; Giopanou, I.; Psallidas, I.; Stathopoulos, G.T. Osteopontin as a Link between Inflammation and Cancer: The Thorax in the Spotlight. Cells 2019, 8, 815. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Wang, X. Role of osteopontin in lung cancer evolution and heterogeneity. Semin. Cell Dev. Biol. 2017, 64, 40–47. [Google Scholar] [CrossRef]
- Giopanou, I.; Kanellakis, N.I.; Giannou, A.D.; Lilis, I.; Marazioti, A.; Spella, M.; Papaleonidopoulos, V.; Simoes, D.C.M.; Zazara, D.E.; Agalioti, T.; et al. Osteopontin drives KRAS-mutant lung adenocarcinoma. Carcinogenesis 2020, 41, 1134–1144. [Google Scholar] [CrossRef]
- Ogburn, D.; Bhalla, S.; Leffler, N.; Mohan, A.; Malur, A.; Malur, A.G.; McPeek, M.; Barna, B.P.; Thomassen, M.J. The M2a Macrophage Phenotype Accompanies Pulmonary Granuloma Resolution in Mmp12 Knock-Out Mice Instilled with Multiwall Carbon Nanotubes. Int. J. Mol. Sci. 2021, 22, 11019. [Google Scholar] [CrossRef]
- Tsunematsu, T.; Arakaki, R.; Sato, M.; Saito, M.; Otsuka, K.; Furukawa, Y.; Taquahashi, Y.; Kanno, J.; Ishimaru, N. Exposure to Multiwall Carbon Nanotubes Promotes Fibrous Proliferation by Production of Matrix Metalloproteinase-12 via NF-kappaB Activation in Chronic Peritonitis. Am. J. Pathol. 2022, 192, 1559–1572. [Google Scholar] [CrossRef]
- Werynska, B.; Pula, B.; Muszczynska-Bernhard, B.; Gomulkiewicz, A.; Jethon, A.; Podhorska-Okolow, M.; Jankowska, R.; Dziegiel, P. Expression of metallothionein-III in patients with non-small cell lung cancer. Anticancer. Res. 2013, 33, 965–974. [Google Scholar]
- Koh, J.Y.; Lee, S.J. Metallothionein-3 as a multifunctional player in the control of cellular processes and diseases. Mol. Brain 2020, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Sarr, D.; Toth, E.; Gingerich, A.; Rada, B. Antimicrobial actions of dual oxidases and lactoperoxidase. J. Microbiol. 2018, 56, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Lu, C.; Stewart, D.J.; Gu, J.; Huang, M.; Chang, D.W.; Lippman, S.M.; Wu, X. Systematic evaluation of apoptotic pathway gene polymorphisms and lung cancer risk. Carcinogenesis 2012, 33, 1699–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, K.; He, B.; Yang, B.B.; Xu, T.; Chen, X.; Xu, M.; Liu, X.; Sun, H.; Pan, Y.; Wang, S. The pro-metastasis effect of circANKS1B in breast cancer. Mol. Cancer 2018, 17, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerr, K.M.; Bibeau, F.; Thunnissen, E.; Botling, J.; Ryška, A.; Wolf, J.; Öhrling, K.; Burdon, P.; Malapelle, U.; Büttner, R. The evolving landscape of biomarker testing for non-small cell lung cancer in Europe. Lung Cancer 2021, 154, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Sholl, L.M. Biomarkers in lung adenocarcinoma: A decade of progress. Arch. Pathol. Lab. Med. 2015, 139, 469–480. [Google Scholar] [CrossRef]
- Sato, M.; Shames, D.S.; Gazdar, A.F.; Minna, J.D. A translational view of the molecular pathogenesis of lung cancer. J. Thorac. Oncol. 2007, 2, 327–343. [Google Scholar] [CrossRef] [Green Version]
- Tenzer, S.; Leidinger, P.; Backes, C.; Huwer, H.; Hildebrandt, A.; Lenhof, H.P.; Wesse, T.; Franke, A.; Meese, E.; Keller, A. Integrated quantitative proteomic and transcriptomic analysis of lung tumor and control tissue: A lung cancer showcase. Oncotarget 2016, 7, 14857. [Google Scholar] [CrossRef] [Green Version]
- Lim, S.B.; Tan, S.J.; Lim, W.T.; Lim, C.T. A merged lung cancer transcriptome dataset for clinical predictive modeling. Sci. Data 2018, 5, 180136. [Google Scholar] [CrossRef] [Green Version]
- Pandiri, A.R.; Sills, R.C.; Ziglioli, V.; Ton, T.V.T.; Hong, H.H.L.; Lahousse, S.A.; Gerrish, K.E.; Auerbach, S.S.; Shockley, K.R.; Bushel, P.R.; et al. Differential transcriptomic analysis of spontaneous lung tumors in b6c3f1 mice: Comparison to human non–small cell lung cancer. Toxicol. Pathol. 2012, 40, 1141–1159. [Google Scholar] [CrossRef] [Green Version]
- Pacurari, M.; Qian, Y.; Porter, D.W.; Wolfarth, M.; Wan, Y.; Luo, D.; Ding, M.; Castranova, V.; Guo, N.L. Multi-walled carbon nanotube-induced gene expression in the mouse lung: Association with lung pathology. Toxicol. Appl. Pharmacol. 2011, 255, 18–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belanger, M.M.; Gaudreau, M.; Roussel, E.; Couet, J. Role of caveolin-1 in etoposide resistance development in A549 lung cancer cells. Cancer Biol. Ther. 2004, 3, 954–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, V.A.; Tapia, J.C.; Rodriguez, D.A.; Parraga, M.; Lisboa, P.; Montoya, M.; Leyton, L.; Quest, A.F. Caveolin-1 controls cell proliferation and cell death by suppressing expression of the inhibitor of apoptosis protein survivin. J. Cell Sci. 2006, 119, 1812–1823. [Google Scholar] [CrossRef] [Green Version]
- Mazieres, J.; He, B.; You, L.; Xu, Z.; Jablons, D.M. Wnt signaling in lung cancer. Cancer Lett. 2005, 222, 1–10. [Google Scholar] [CrossRef]
- Stewart, D.J. Wnt signaling pathway in non-small cell lung cancer. J. Natl. Cancer Inst. 2014, 106, djt356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, P.L.; Salmen, F.; Vickovic, S.; Lundmark, A.; Navarro, J.F.; Magnusson, J.; Giacomello, S.; Asp, M.; Westholm, J.O.; Huss, M.; et al. Visualization and analysis of gene expression in tissue sections by spatial transcriptomics. Science 2016, 353, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, M.; Oki, S.; Kimura, R.; Harada, A.; Maehara, K.; Tanaka, K.; Meno, C.; Ohkawa, Y. High-depth spatial transcriptome analysis by photo-isolation chemistry. Nat. Commun. 2021, 12, 4416. [Google Scholar] [CrossRef] [PubMed]
- Orsi, M.; Palmai-Pallag, M.; Yakoub, Y.; Ibouraadaten, S.; De Beukelaer, M.; Bouzin, C.; Bearzatto, B.; Ambroise, J.; Gala, J.L.; Brusa, D.; et al. Monocytic Ontogeny of Regenerated Macrophages Characterizes the Mesotheliomagenic Responses to Carbon Nanotubes. Front. Immunol. 2021, 12, 666107. [Google Scholar] [CrossRef]
- Morrow, P.E. Possible mechanisms to explain dust overloading of the lungs. Fundam. Appl. Toxicol. 1988, 10, 369–384. [Google Scholar] [CrossRef]
- Rodelsperger, K. Extrapolation of the carcinogenic potency of fibers from rats to humans. Inhal. Toxicol. 2004, 16, 801–807. [Google Scholar] [CrossRef]
- Rodelsperger, K.; Woitowitz, H.J. Airborne fibre concentrations and lung burden compared to the tumour response in rats and humans exposed to asbestos. Ann. Occup. Hyg. 1995, 39, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Institute, I.R.S. The relevance of the rat lung response to particle overload for human risk assessment: A workshop consensus report. Inhal. Toxicol. 2000, 12, 1–17. [Google Scholar] [CrossRef]
- Jarabek, A.M.; Asgharian, B.; Miller, F.J. Dosimetric adjustments for interspecies extrapolation of inhaled poorly soluble particles (PSP). Inhal. Toxicol. 2005, 17, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Kasai, T.; Umeda, Y.; Sasaki, T.; Fukushima, S. Thinking on occupational exposure assessment of multi-walled carbon nanotube carcinogenicity. J. Occup. Health 2019, 61, 208–210. [Google Scholar] [CrossRef]
- Fukushima, S.; Kasai, T.; Umeda, Y.; Ohnishi, M.; Sasaki, T.; Matsumoto, M. Carcinogenicity of multi-walled carbon nanotubes: Challenging issue on hazard assessment. J. Occup. Health 2018, 60, 10–30. [Google Scholar] [CrossRef] [Green Version]
- Guseva Canu, I.; Batsungnoen, K.; Maynard, A.; Hopf, N.B. State of knowledge on the occupational exposure to carbon nanotubes. Int. J. Hyg. Environ. Health 2020, 225, 113472. [Google Scholar] [CrossRef]
- Kuijpers, E.; Bekker, C.; Fransman, W.; Brouwer, D.; Tromp, P.; Vlaanderen, J.; Godderis, L.; Hoet, P.; Lan, Q.; Silverman, D.; et al. Occupational Exposure to Multi-Walled Carbon Nanotubes During Commercial Production Synthesis and Handling. Ann. Occup. Hyg. 2016, 60, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Erdely, A.; Dahm, M.; Chen, B.T.; Zeidler-Erdely, P.C.; Fernback, J.E.; Birch, M.E.; Evans, D.E.; Kashon, M.L.; Deddens, J.A.; Hulderman, T.; et al. Carbon nanotube dosimetry: From workplace exposure assessment to inhalation toxicology. Part. Fibre Toxicol. 2013, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Mostovenko, E.; Dahm, M.M.; Schubauer-Berigan, M.K.; Eye, T.; Erdely, A.; Young, T.L.; Campen, M.J.; Ottens, A.K. Serum peptidome: Diagnostic window into pathogenic processes following occupational exposure to carbon nanomaterials. Part. Fibre Toxicol. 2021, 18, 39. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year 0.5 | Year 1 | Year 2 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Upregulated | Downregulated | Upregulated | Downregulated | Upregulated | Downregulated | ||||||
| Gene Symbol | Log2FC | Gene Symbol | Log2FC | Gene Symbol | Log2FC | Gene Symbol | Log2FC | Gene Symbol | Log2FC | Gene Symbol | Log2FC |
| RragB | 9.32 | Faahl | −8.32 | Mmp7 | 9.57 | Rps27a_2 | −12.12 | Nlrp6_1 | 9.39 | Rps27a_2 | −12.18 |
| Spp1 | 8.98 | RT1-DMa | −8.13 | Spp1 | 9.40 | Snip1 | −8.22 | Spp1 | 9.20 | Rpl39 | −11.37 |
| Zfp84_2 | 8.65 | Snip1 | −7.69 | Mt3 | 8.90 | Ndst2 | −6.19 | Ankrd34c | 8.80 | Tsen34 | −8.39 |
| Pla2g4e | 8.36 | Ccdc89 | −5.95 | Lpo | 7.90 | Yme1l1 | −6.18 | Mt3 | 8.72 | Omd | −7.73 |
| Ankrd34c | 8.06 | Sbk1 | −5.28 | Adamts18 | 7.80 | Rin1 | −4.99 | Anks1b | 8.57 | Myh9 | −7.68 |
| Adamts18 | 7.92 | Dlk1 | −5.14 | Nlrp6_1 | 7.78 | Myl2 | −3.95 | Mc5r | 8.51 | Ceacam4 | −7.44 |
| Impad1_1 | 7.79 | Myl2 | −3.98 | Defb3 | 6.49 | Pnpla2 | −3.94 | Mmp7 | 8.26 | Dusp11 | −7.35 |
| Tnf | 7.08 | Dhrs7c | −3.08 | Retnla | 6.46 | Dlk1 | −3.69 | Lpo | 8.22 | Chid1 | −5.53 |
| Mmp7 | 6.95 | Alpg | −2.86 | AABR07035791.1 | 6.18 | Cdh19 | −3.41 | Faim_1 | 8.07 | Slc6a21 | −4.70 |
| Aldh18a1 | 6.91 | Rbm12_1 | −2.78 | Mmp12 | 5.99 | Upk1a | −3.25 | Csap1 | 8.06 | Cfc1 | −4.49 |
| Retnla | 6.44 | Kcna6 | −2.72 | Psca | 5.62 | Slc6a15 | −2.98 | Tpsab1 | 7.86 | Acoxl | −4.48 |
| Srd5a2 | 6.34 | Serpinb10 | −2.66 | Sctr | 5.61 | Cct8l1_2 | −2.96 | RGD1564571 | 7.47 | Dlk1 | −4.35 |
| Marco | 6.24 | Galnt14 | −2.62 | Slc26a4 | 5.57 | Hif3a | −2.93 | Ccl1 | 7.26 | Prrt4 | −4.24 |
| Mt3 | 6.05 | Slc6a15 | −2.55 | Reg3b | 5.51 | LOC685767 | −2.90 | Srd5a2 | 7.16 | Myl2 | −4.09 |
| Yme1l1 | 5.84 | Nhlrc4 | −2.49 | Srd5a2 | 5.45 | Kcne5 | −2.89 | Mrgprx2 | 6.74 | Krtap17-1 | −3.96 |
| Mmp12 | 5.71 | Gp1bb | −2.37 | Ocm2 | 5.44 | Gpr50 | −2.88 | Sctr | 6.74 | Serpinb10 | −3.90 |
| Slc26a4 | 5.62 | Lmod2 | −2.30 | Smtnl1 | 5.44 | Clcn2 | −2.88 | Sult1c2 | 6.67 | Aldh18a1 | −3.87 |
| Lpo | 5.47 | Sh2d6 | −2.29 | Tmem72 | 5.37 | Tenm2 | −2.77 | A2m | 6.54 | Unc5d | −3.83 |
| AABR07035791.1 | 5.41 | Vit | −2.23 | Ankrd34c | 5.31 | Rorb | −2.77 | Aox4 | 6.29 | Clcn2 | −3.60 |
| Prlhr | 5.40 | Slc6a21 | −2.22 | Orm1 | 5.26 | Fbxo40 | −2.73 | Mmp12 | 6.22 | Slc7a10 | −3.52 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hojo, M.; Maeno, A.; Sakamoto, Y.; Yamamoto, Y.; Taquahashi, Y.; Hirose, A.; Suzuki, J.; Inomata, A.; Nakae, D. Time-Course of Transcriptomic Change in the Lungs of F344 Rats Repeatedly Exposed to a Multiwalled Carbon Nanotube in a 2-Year Test. Nanomaterials 2023, 13, 2105. https://doi.org/10.3390/nano13142105
Hojo M, Maeno A, Sakamoto Y, Yamamoto Y, Taquahashi Y, Hirose A, Suzuki J, Inomata A, Nakae D. Time-Course of Transcriptomic Change in the Lungs of F344 Rats Repeatedly Exposed to a Multiwalled Carbon Nanotube in a 2-Year Test. Nanomaterials. 2023; 13(14):2105. https://doi.org/10.3390/nano13142105
Chicago/Turabian StyleHojo, Motoki, Ai Maeno, Yoshimitsu Sakamoto, Yukio Yamamoto, Yuhji Taquahashi, Akihiko Hirose, Jin Suzuki, Akiko Inomata, and Dai Nakae. 2023. "Time-Course of Transcriptomic Change in the Lungs of F344 Rats Repeatedly Exposed to a Multiwalled Carbon Nanotube in a 2-Year Test" Nanomaterials 13, no. 14: 2105. https://doi.org/10.3390/nano13142105