
Exposure of Rats to Multi-Walled Carbon Nanotubes: Correlation of Inhalation Exposure to Lung Burden, Bronchoalveolar Lavage Fluid Findings, and Lung Morphology
Abstract
:1. Introduction
2. Materials and Methods
2.1. Test Materials
2.2. Animals and Breeding Environment
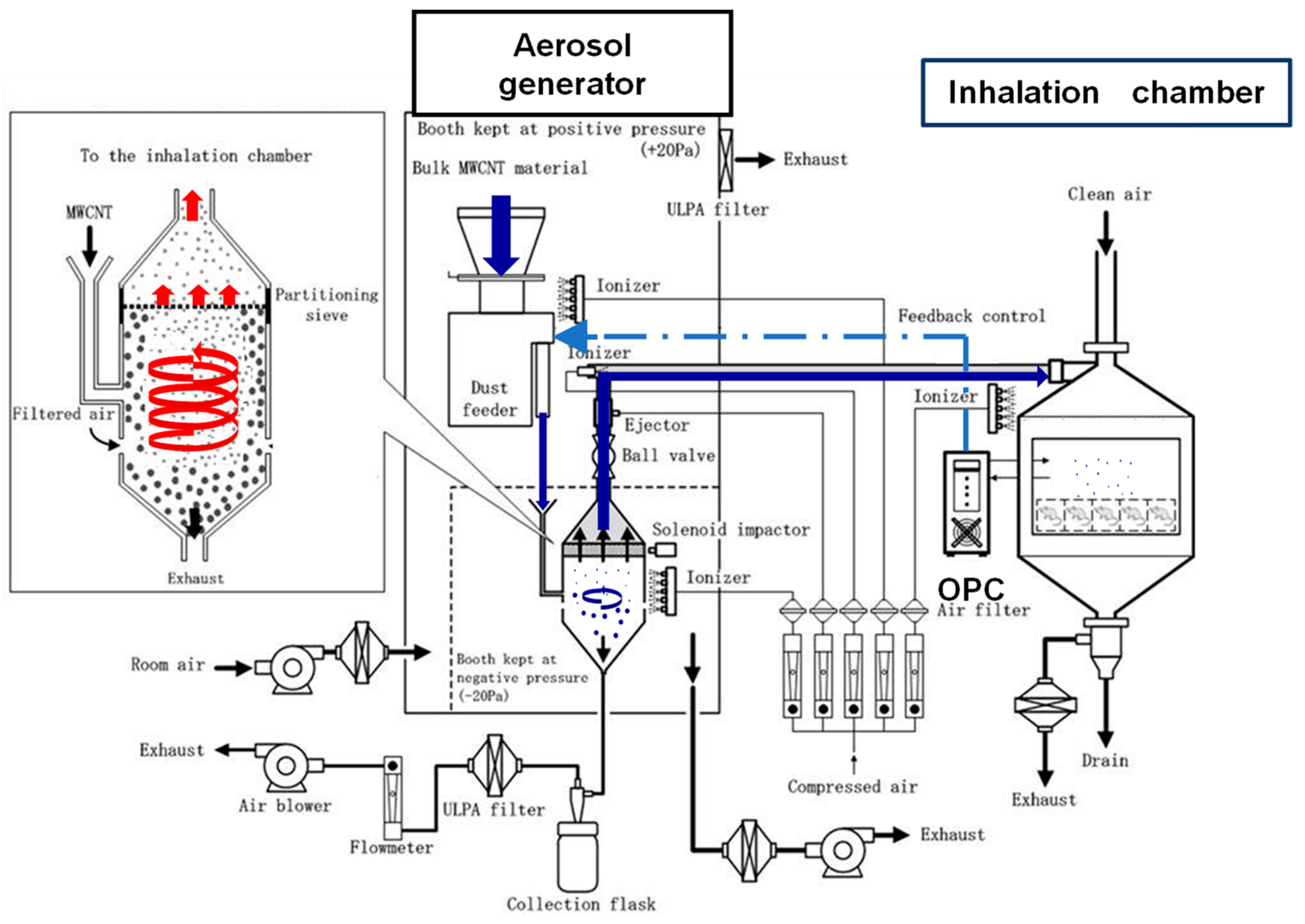
2.3. Development of the Generator Used for Inhalation Exposure to MWNT-7 Aerosols
2.4. Lung Burden Analysis
2.5. Bronchoalveolar Lavage Fluid (BALF) and Pathological Examinations
2.6. Summary of Studies Used to Complete the 2-Year Carcinogenicity Study
2.7. Statistics
3. Results
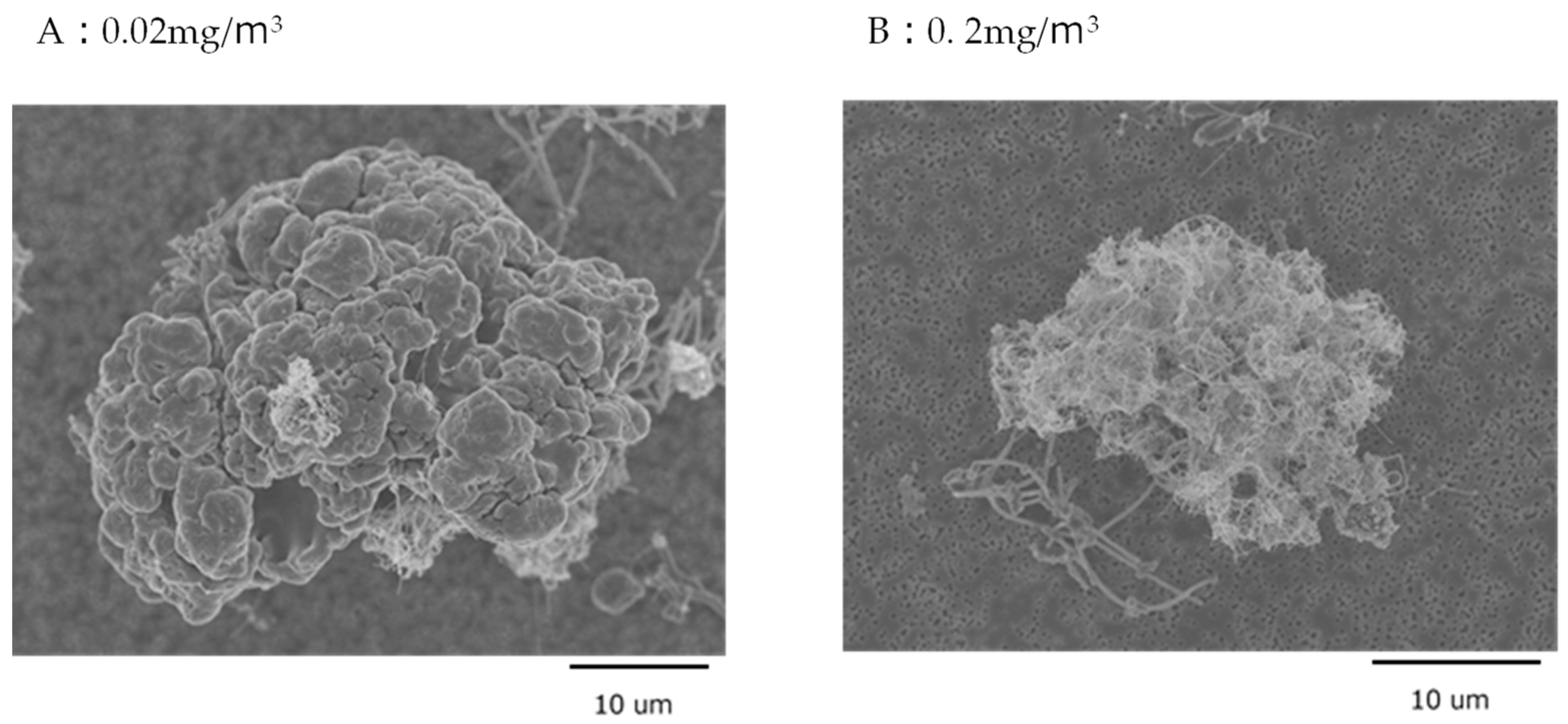
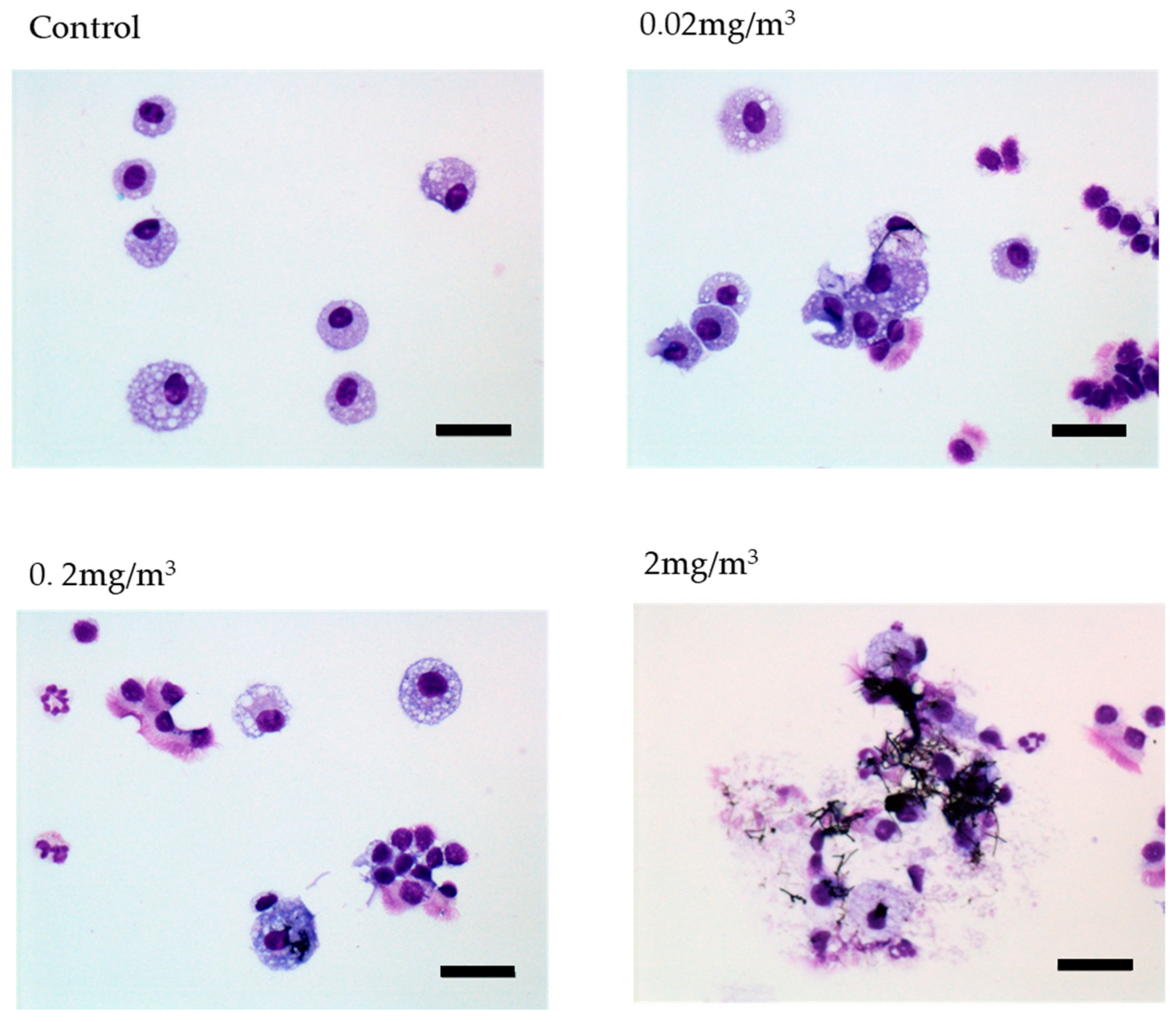
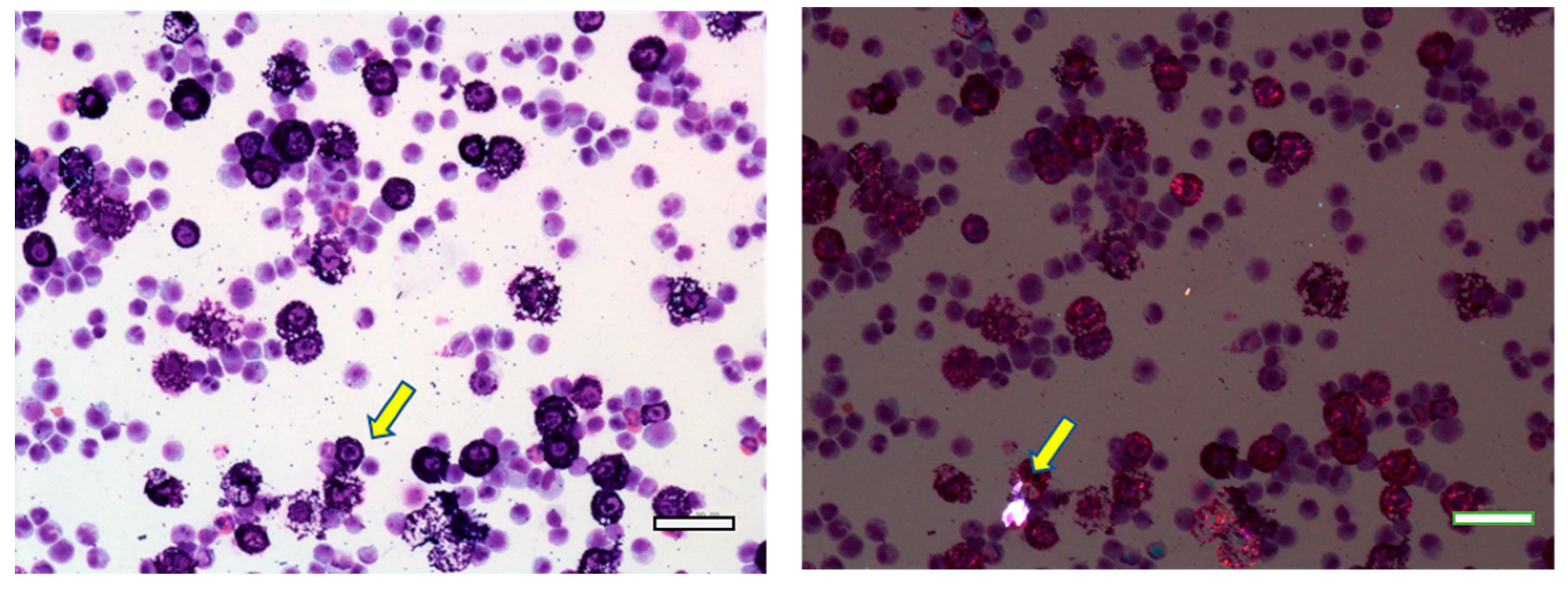
3.1. MWNT-7 Fibers in the Lung Tissue
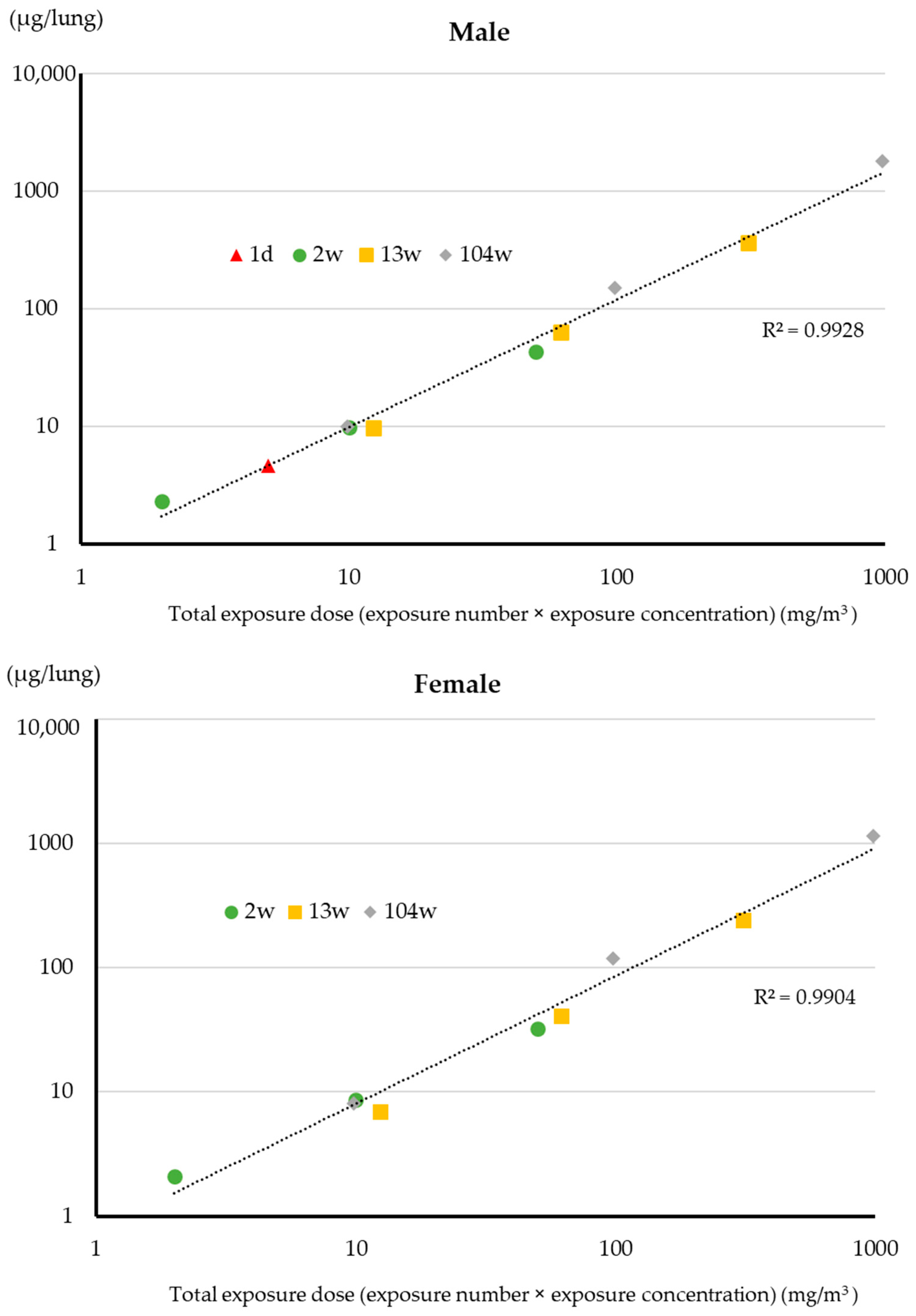
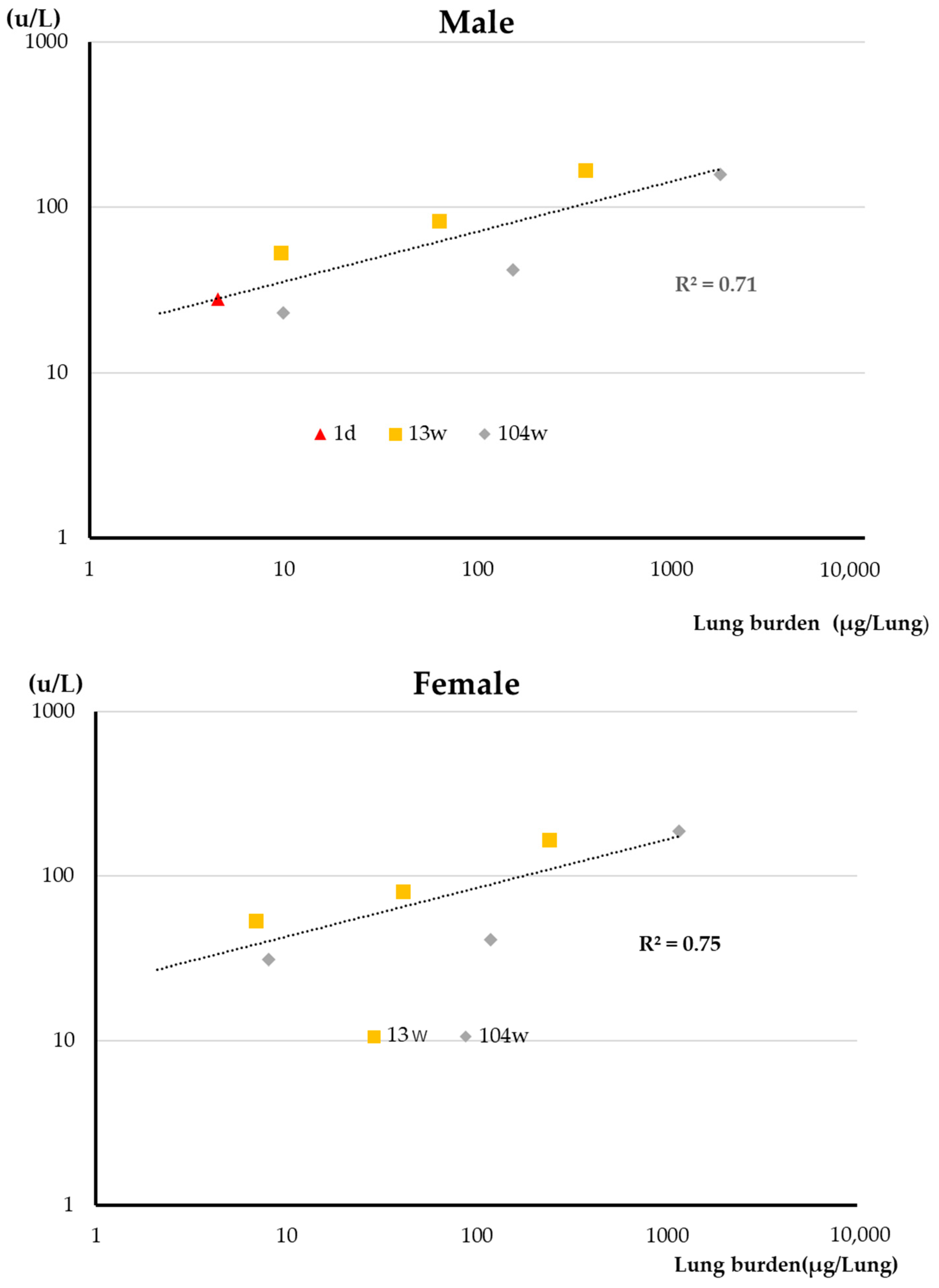
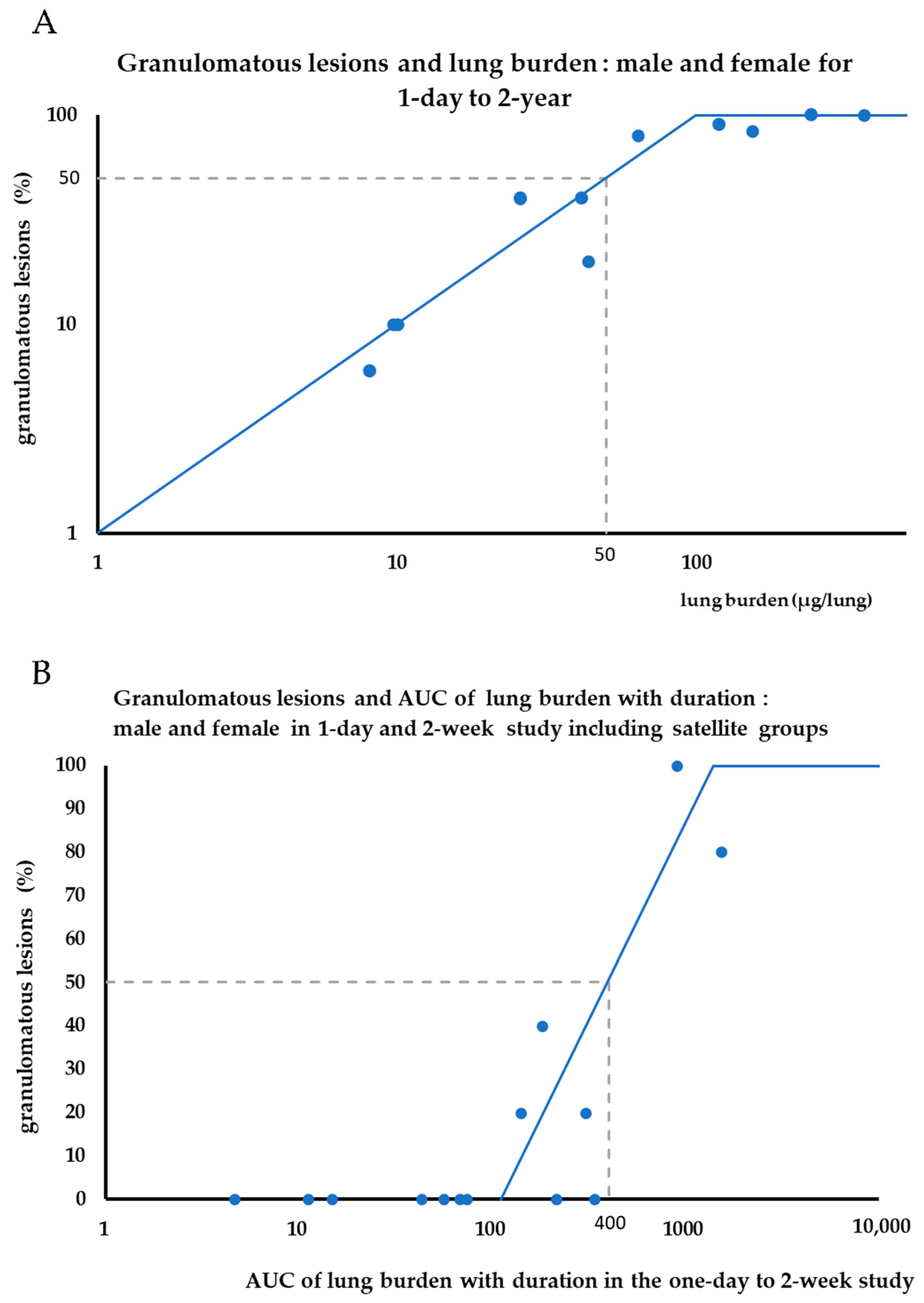
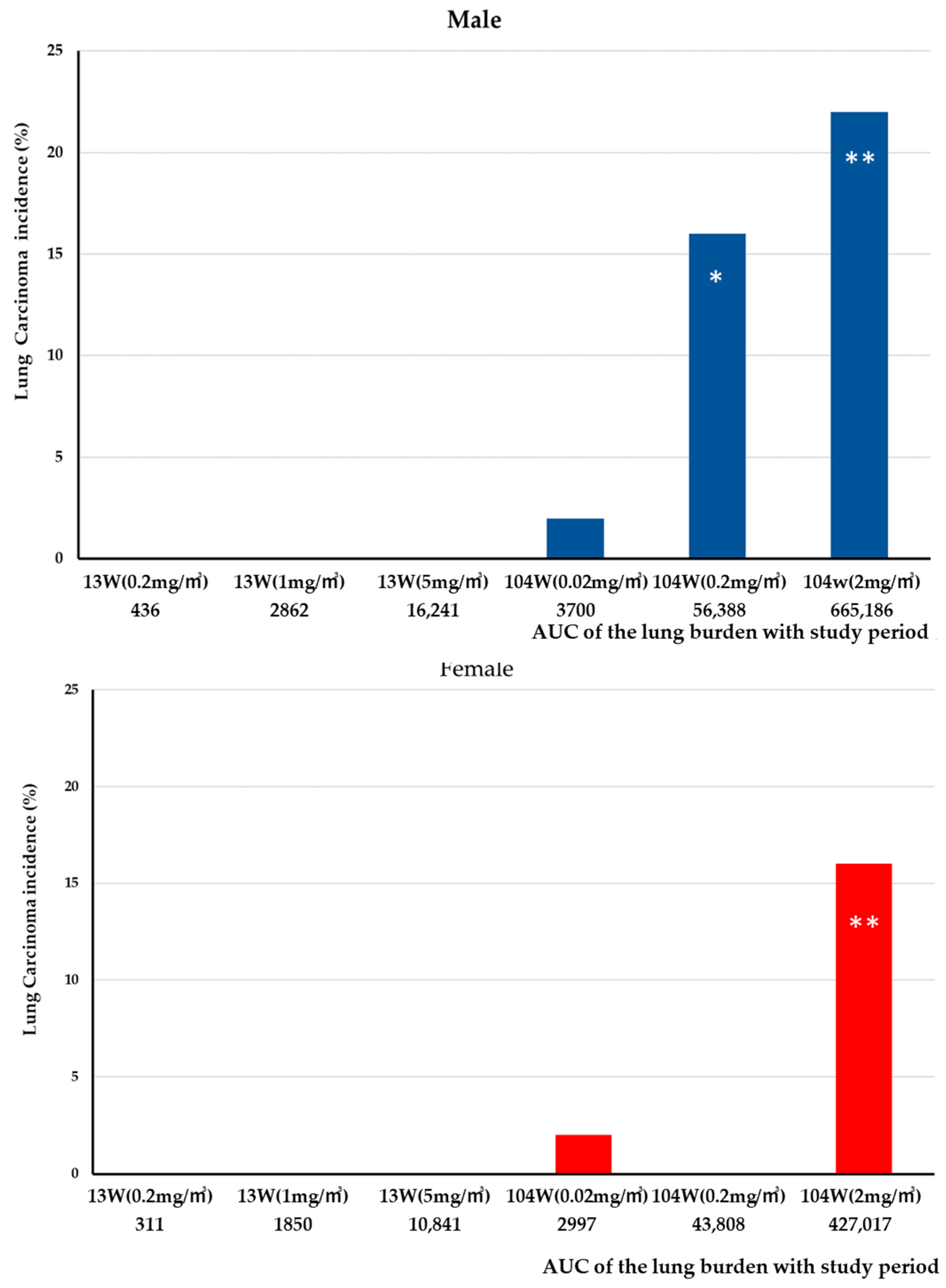
3.2. Lung Burden
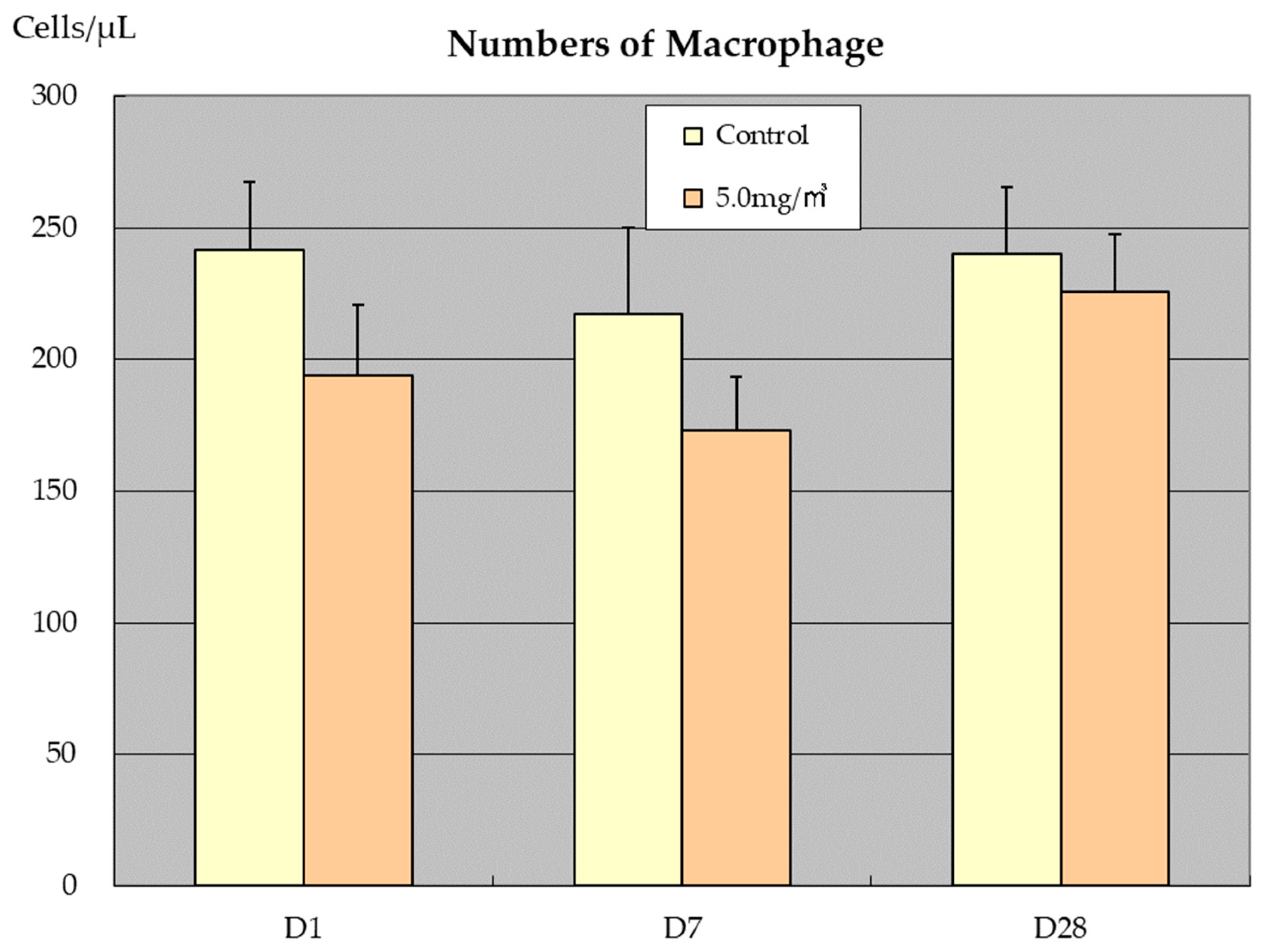
3.3. BALF and Abdominal and Thoracic Lavage Fluid Findings
3.4. Lung Morphology and Pathological Examinations
4. Discussion
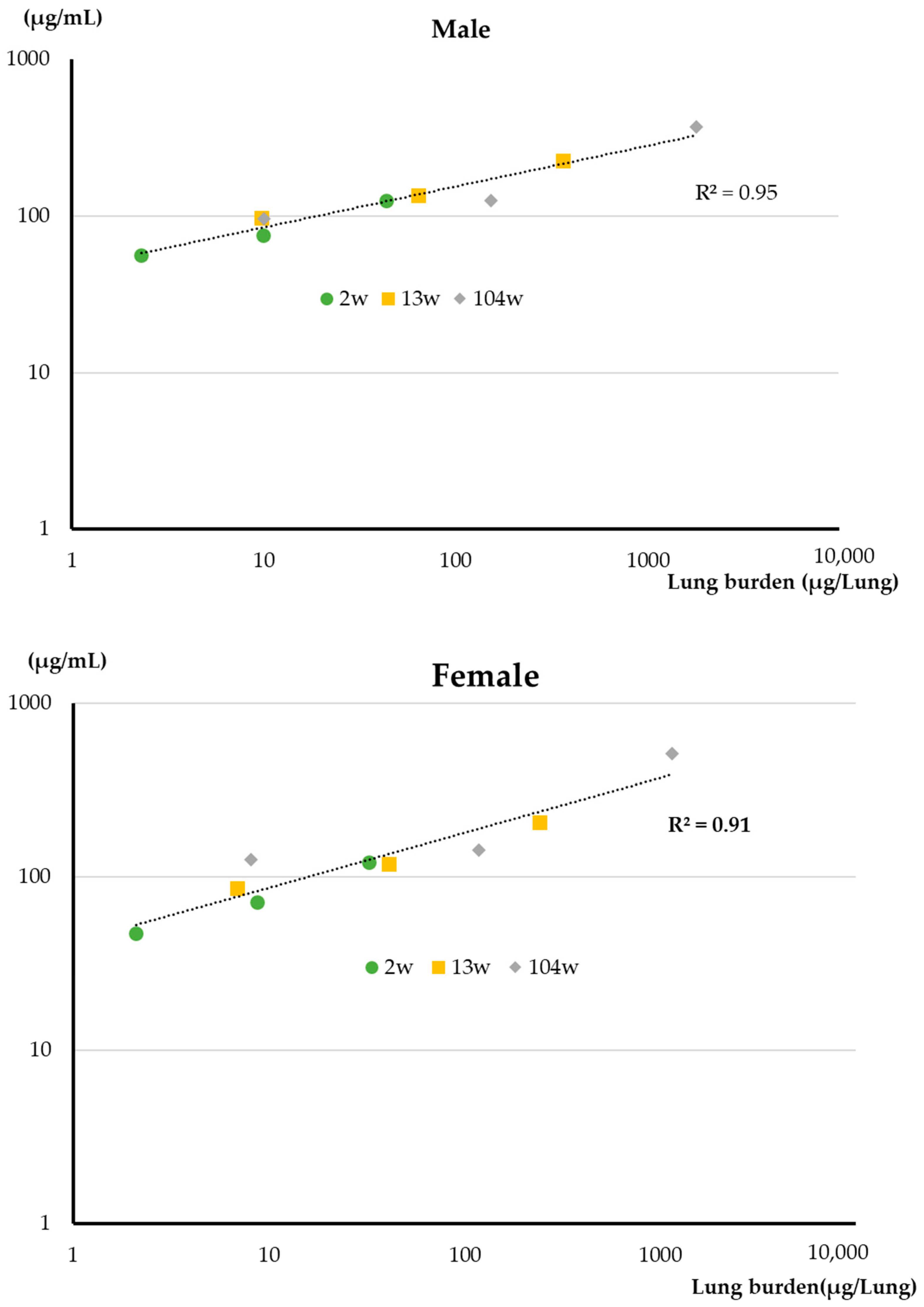
4.1. Lung Burden
4.2. Mechanisms of MWNT-7 Fiber Retention in the Lung
4.3. Pathogenicity of MWNT-7 Fibers
4.4. Comparison with Intratracheal Instillation
5. Conclusions
6. Patents
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Donaldson, K.; Aitken, R.; Tran, L.; Stone, V.; Duffin, R.; Forrest, G.; Alexander, A. Carbon nanotubes: A review of their properties in relation to pulmonary toxicology and workplace safety. Toxicol. Sci. 2006, 92, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Takagi, A.; Hirose, A.; Nishimura, T.; Fukumori, N.; Ogata, A.; Ohashi, N.; Kitajima, S.; Kanno, J. Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci. 2008, 33, 105–116. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, K.; Stone, V.; Seaton, A.; Tran, L.; Aitken, R.; Poland, C. Re: Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci. 2008, 33, 385. [Google Scholar] [CrossRef] [PubMed]
- Ichihara, G.; Castranova, V.; Tanioka, A.; Miyazawa, K. Re: Induction of mesothelioma in p53+/− mouse by intraperitoneal application of multi-wall carbon nanotube. J. Toxicol. Sci. 2008, 33, 381–382. [Google Scholar] [CrossRef]
- Sakamoto, Y.; Nakae, D.; Fukumori, N.; Tayama, K.; Maekawa, A.; Imai, K.; Hirose, A.; Nishimura, T.; Ohashi, N.; Ogata, A. Induction of mesothelioma by a single intrascrotal administration of multi-wall carbon nanotube in intact male Fischer 344 rats. J. Toxicol. Sci. 2009, 34, 65–76. [Google Scholar] [CrossRef]
- Kasai, T.; Gotoh, K.; Nishizawa, T.; Sasaki, T.; Katagiri, T.; Umeda, Y.; Toya, T.; Fukushima, S. Development of a new multi-walled carbon nanotube (MWCNT) aerosol generation and exposure system and confirmation of suitability for conducting a single-exposure inhalation study of MWCNT in rats. Nanotoxicology 2014, 8, 169–178. [Google Scholar] [CrossRef]
- Umeda, Y.; Kasai, T.; Saito, M.; Kondo, H.; Toya, T.; Aiso, S.; Okuda, H.; Nishizawa, T.; Fukushima, S. Two-week Toxicity of Multi-walled Carbon Nanotubes by Whole-body Inhalation Exposure in Rats. J. Toxicol. Pathol. 2013, 26, 131–140. [Google Scholar] [CrossRef]
- Kasai, T.; Umeda, Y.; Ohnishi, M.; Kondo, H.; Takeuchi, T.; Aiso, S.; Nishizawa, T.; Matsumoto, M.; Fukushima, S. Thirteen-week study of toxicity of fiber-like multi-walled carbon nanotubes with whole-body inhalation exposure in rats. Nanotoxicology 2015, 9, 413–422. [Google Scholar] [CrossRef]
- Sargent, L.M.; Porter, D.W.; Staska, L.M.; Hubbs, A.F.; Lowry, D.T.; Battelli, L.; Siegrist, K.J.; Kashon, M.L.; Mercer, R.R.; Bauer, A.K.; et al. Promotion of lung adenocarcinoma following inhalation exposure to multi-walled carbon nanotubes. Part. Fibre Toxicol. 2014, 11, 3. [Google Scholar] [CrossRef]
- Nagai, H.; Okazaki, Y.; Chew, S.H.; Misawa, N.; Yamashita, Y.; Akatsuka, S.; Ishihara, T.; Yamashita, K.; Yoshikawa, Y.; Yasui, H.; et al. Diameter and rigidity of multiwalled carbon nanotubes are critical factors in mesothelial injury and carcinogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1330–E1338. [Google Scholar] [CrossRef]
- Takagi, A.; Hirose, A.; Futakuchi, M.; Tsuda, H.; Kanno, J. Dose-dependent mesothelioma induction by intraperitoneal administration of multi-wall carbon nanotubes in p53 heterozygous mice. Cancer Sci. 2012, 103, 1440–1444. [Google Scholar] [CrossRef] [PubMed]
- Grosse, Y.; Loomis, D.; Guyton, K.Z.; Lauby-Secretan, B.; El Ghissassi, F.; Bouvard, V.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; et al. Carcinogenicity of fluoro-edenite, silicon carbide fibres and whiskers, and carbon nanotubes. Lancet Oncol. 2014, 15, 1427–1428. [Google Scholar] [CrossRef] [PubMed]
- IARC. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. In Some Nanomaterials and Some Fibres; International Agency for Research on Cancer Lyon: Lyon, France, 2017; Volume 111. [Google Scholar]
- Kasai, T.; Umeda, Y.; Ohnishi, M.; Mine, T.; Kondo, H.; Takeuchi, T.; Matsumoto, M.; Fukushima, S. Lung carcinogenicity of inhaled multi-walled carbon nanotube in rats. Part. Fibre Toxicol. 2016, 13, 53. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, M.; Yajima, H.; Kasai, T.; Umeda, Y.; Yamamoto, M.; Yamamoto, S.; Okuda, H.; Suzuki, M.; Nishizawa, T.; Fukushima, S. Novel method using hybrid markers: Development of an approach for pulmonary measurement of multi-walled carbon nanotubes. J. Occup. Med. Toxicol. 2013, 8, 30. [Google Scholar] [CrossRef]
- Kohyama, N.; Suzuki, Y. Analysis of asbestos fibers in lung parenchyma, pleural plaques, and mesothelioma tissues of North American insulation workers. Ann. N. Y. Acad. Sci. 1991, 643, 27–52. [Google Scholar] [CrossRef]
- Stanton, M.F.; Wrench, C. Mechanisms of mesothelioma induction with asbestos and fibrous glass. J. Natl. Cancer Inst. 1972, 48, 797–821. [Google Scholar]
- Pott, F.; Huth, F.; Friedrichs, K.H. Tumorigenic effect of fibrous dusts in experimental animals. Environ. Health Perspect. 1974, 9, 313–315. [Google Scholar] [CrossRef]
- Harington, J.S. Fiber carcinogenesis: Epidemiologic observations and the Stanton hypothesis. J. Natl. Cancer Inst. 1981, 67, 977–989. [Google Scholar]
- Stanton, M.F. Editorial: Fiber carcinogenesis: Is asbestos the only hazard? J. Natl. Cancer Inst. 1974, 52, 633–634. [Google Scholar] [CrossRef]
- Stanton, M.F.; Layard, M.; Tegeris, A.; Miller, E.; May, M.; Morgan, E.; Smith, A. Relation of particle dimension to carcinogenicity in amphibole asbestoses and other fibrous minerals. J. Natl. Cancer Inst. 1981, 67, 965–975. [Google Scholar]
- Pott, F.; Dolgner, R.; Friedrichs, K.H.; Huth, F. The oncogenic effect of fibrous dust. Animal experiments and their relationship with human carcinogenesis. Ann. Anat. Pathol. 1976, 21, 237–246. [Google Scholar]
- Pott, F.; Friedrichs, K.H.; Huth, F. Results of animal experiments concerning the carcinogenic effect of fibrous dusts and their interpretation with regard to the carcinogenesis in humans (author’s transl). Zentralblatt Fur. Bakteriol. Parasitenkd. Infekt. Und. Hygiene. Erste Abt. Originale. Reihe B Hyg. Prav. Med. 1976, 162, 467–505. [Google Scholar]
- Pott, F.; Huth, F.; Friedrichs, K.H. Results of animal carcinogenesis studies after application of fibrous glass and their implications regarding human exposure. Occup. Expo. Fibrous Glass 1976, 183–191. Available online: https://ntrl.ntis.gov/NTRL/dashboard/searchResults/titleDetail/PB258869.xhtml (accessed on 30 July 2023).
- Pott, F. Some aspects on the dosimetry of the carcinogenic potency of asbestos and other fibrous dusts. Staub-Reinhalt Luft 1978, 38, 486–490. [Google Scholar]
- Pott, F. Animal experiments on biological effects of mineral fibres. IARC Sci. Publ. 1980, 30, 261–272. [Google Scholar]
- Schneider, T.; Skotte, J.; Nissen, P. Man-made mineral fiber size fractions and their interrelation. Scand. J. Work Environ. Health 1985, 11, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Adachi, S.; Kawamura, K.; Takemoto, K. A trial on the quantitative risk assessment of man-made mineral fibers by the rat intraperitoneal administration assay using the JFM standard fibrous samples. Ind. Health 2001, 39, 168–174. [Google Scholar] [CrossRef] [PubMed]
- McDonald, J.C.; Armstrong, B.; Case, B.; Doell, D.; McCaughey, W.T.E.; McDonald, A.D.; Sébastien, P. Mesothelioma and asbestos fiber type. Evidence from lung tissue analyses. Cancer 1989, 63, 1544–1547. [Google Scholar] [CrossRef]
- Davis, J.M.; Addison, J.; Bolton, R.E.; Donaldson, K.; Jones, A.D.; Smith, T. The pathogenicity of long versus short fibre samples of amosite asbestos administered to rats by inhalation and intraperitoneal injection. Br. J. Exp. Pathol. 1986, 67, 415–430. [Google Scholar]
- Donaldson, K.; Murphy, F.A.; Duffin, R.; Poland, C.A. Asbestos, carbon nanotubes and the pleural mesothelium: A review of the hypothesis regarding the role of long fibre retention in the parietal pleura, inflammation and mesothelioma. Part. Fibre Toxicol. 2010, 7, 5. [Google Scholar] [CrossRef]
- Donaldson, K.; Murphy, F.; Schinwald, A.; Duffin, R.; Poland, C.A. Identifying the pulmonary hazard of high aspect ratio nanoparticles to enable their safety-by-design. Nanomedicine 2011, 6, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Morrow, P.E. Possible mechanisms to explain dust overloading of the lungs. Fundam. Appl. Toxicol. 1988, 10, 369–384. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, K.E. Review of Lung Particle Overload, Rat Lung Cancer, and the Conclusions of the Edinburgh Expert Panel-It’s Time to Revisit Cancer Hazard Classifications for Titanium Dioxide and Carbon Black. Front. Public Health 2022, 10, 907318. [Google Scholar] [CrossRef] [PubMed]
- IARC. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. In Man-Made Vitreous Fibres; International Agency for Research on Cancer, World Health Organization: Geneva, Switzerland, 2002. [Google Scholar]
- Pauluhn, J. Poorly soluble particulates: Searching for a unifying denominator of nanoparticles and fine particles for DNEL estimation. Toxicology 2011, 279, 176–188. [Google Scholar] [CrossRef]
- Pauluhn, J. Subchronic 13-week inhalation exposure of rats to multiwalled carbon nanotubes: Toxic effects are determined by density of agglomerate structures, not fibrillar structures. Toxicol. Sci. 2010, 113, 226–242. [Google Scholar] [CrossRef]
- Neupane, A.S.; Willson, M.; Chojnacki, A.K.; Castanheira, F.V.E.S.; Morehouse, C.; Carestia, A.; Keller, A.E.; Peiseler, M.; DiGiandomenico, A.; Kelly, M.M.; et al. Patrolling Alveolar Macrophages Conceal Bacteria from the Immune System to Maintain Homeostasis. Cell 2020, 183, 110.e11–125.e11. [Google Scholar] [CrossRef]
- Francis, A.P.; Ganapathy, S.; Palla, V.R.; Murthy, P.B.; Ramaprabhu, S.; Devasena, T. One time nose-only inhalation of MWCNTs: Exploring the mechanism of toxicity by intermittent sacrifice in Wistar rats. Toxicol. Rep. 2015, 2, 111–120. [Google Scholar] [CrossRef]
- Boulanger, G.; Andujar, P.; Pairon, J.-C.; Billon-Galland, M.-A.; Dion, C.; Dumortier, P.; Brochard, P.; Sobaszek, A.; Bartsch, P.; Paris, C.; et al. Quantification of short and long asbestos fibers to assess asbestos exposure: A review of fiber size toxicity. Environ. Health 2014, 13, 59. [Google Scholar] [CrossRef]
- Fukushima, S.; Kasai, T.; Umeda, Y.; Ohnishi, M.; Sasaki, T.; Matsumoto, M. Carcinogenicity of multi-walled carbon nanotubes: Challenging issue on hazard assessment. J. Occup. Health 2018, 60, 10–30. [Google Scholar] [CrossRef]
- Asakura, M.; Sasaki, T.; Sugiyama, T.; Takaya, M.; Koda, S.; Nagano, K.; Arito, H.; Fukushima, S. Genotoxicity and cytotoxicity of multi-wall carbon nanotubes in cultured Chinese hamster lung cells in comparison with chrysotile A fibers. J. Occup. Health 2010, 52, 155–166. [Google Scholar] [CrossRef]
- Fukushima, S.; Kasai, T.; Senoh, H.; Umeda, Y.; Mine, T.; Sasaki, T.; Kondo, H.; Matsumoto, M.; Aiso, S. Contribution of toxicological pathology to occupational health: Lung carcinogenicity of fibrous and particulate substances in rats. J. Toxicol. Pathol. 2023, 36, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Totsuka, Y.; Ishino, K.; Matsumoto, Y.; Tada, Y.; Nakae, D.; Goto, S.; Masuda, S.; Ogo, S.; Kawanishi, M.; et al. Genotoxicity of multi-walled carbon nanotubes in both in vitro and in vivo assay systems. Nanotoxicology 2013, 7, 452–461. [Google Scholar] [CrossRef] [PubMed]
- Møller, P.; Vest Christophersen, D.; Jensen, D.M.; Kermanizadeh, A.; Roursgaard, M.; Jacobsen, N.R.; Hemmingsen, J.G.; Danielsen, P.H.; Cao, Y.; Jantzen, K.; et al. Role of oxidative stress in carbon nanotube-generated health effects. Arch. Toxicol. 2014, 88, 1939–1964. [Google Scholar] [CrossRef]
- Møller, P.; Jacobsen, N.R. Weight of evidence analysis for assessing the genotoxic potential of carbon nanotubes. Crit. Rev. Toxicol. 2017, 47, 867–884. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, K.J.; Reynolds, S.H.; Porter, D.W.; Mercer, R.R.; Bauer, A.K.; Lowry, D.; Cena, L.; Stueckle, T.A.; Kashon, M.L.; Wiley, J.; et al. Mitsui-7, heat-treated, and nitrogen-doped multi-walled carbon nanotubes elicit genotoxicity in human lung epithelial cells. Part. Fibre Toxicol. 2019, 16, 36. [Google Scholar] [CrossRef]
- Yasui, M.; Kamoshita, N.; Nishimura, T.; Honma, M. Mechanism of induction of binucleated cells by multiwalled carbon nanotubes as revealed by live-cell imaging analysis. Genes Environ. 2015, 37, 6. [Google Scholar] [CrossRef]
- Funahashi, S.; Okazaki, Y.; Ito, D.; Asakawa, A.; Nagai, H.; Tajima, M.; Toyokuni, S. Asbestos and multi-walled carbon nanotubes generate distinct oxidative responses in inflammatory cells. J. Clin. Biochem. Nutr. 2015, 56, 111–117. [Google Scholar] [CrossRef]
- Gualtieri, A.F. Journey to the centre of the lung. The perspective of a mineralogist on the carcinogenic effects of mineral fibres in the lungs. J. Hazard. Mater. 2023, 442, 130077. [Google Scholar] [CrossRef]
- Hardy, J.A.; Aust, A.E. Iron in asbestos chemistry and carcinogenicity. Chem. Rev. 1995, 95, 97–118. [Google Scholar] [CrossRef]
- Hojo, M.; Maeno, A.; Sakamoto, Y.; Ohnuki, A.; Tada, Y.; Yamamoto, Y.; Ikushima, K.; Inaba, R.; Suzuki, J.; Taquahashi, Y.; et al. Two-year intermittent exposure of a multiwalled carbon nanotube by intratracheal instillation induces lung tumors and pleural mesotheliomas in F344 rats. Part. Fibre Toxicol. 2022, 19, 38. [Google Scholar] [CrossRef]
- Hojo, M.; Maeno, A.; Sakamoto, Y.; Yamamoto, Y.; Taquahashi, Y.; Hirose, A.; Suzuki, J.; Inomata, A.; Nakae, D. Time-course of transcriptomic change in the lungs of F344 rats repeatedly exposed to a multiwalled carbon nanotube in a 2-year test. Nanomaterials 2023, 13, 2105. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Okazaki, Y.; Chew, S.H.; Misawa, N.; Miyata, Y.; Shinohara, H.; Toyokuni, S. Intraperitoneal administration of tangled multiwalled carbon nanotubes of 15 nm in diameter does not induce mesothelial carcinogenesis in rats. Pathol. Int. 2013, 63, 457–462. [Google Scholar] [CrossRef]
- Xue, J.; Patergnani, S.; Giorgi, C.; Suarez, J.; Goto, K.; Bononi, A.; Tanji, M.; Novelli, F.; Pastorino, S.; Xu, R.; et al. Asbestos induces mesothelial cell transformation via HMGB1-driven autophagy. Proc. Natl. Acad. Sci. USA 2020, 117, 25543–25552. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, M.E.; Crippa, M.P.; Manfredi, A.A.; Mezzapelle, R.; Querini, P.R.; Venereau, E. High-mobility group box 1 protein orchestrates responses to tissue damage via inflammation, innate and adaptive immunity, and tissue repair. Immunol. Rev. 2017, 280, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Suarez, J.S.; Minaai, M.; Li, S.; Gaudino, G.; Pass, H.I.; Carbone, M.; Yang, H. HMGB1 as a therapeutic target in disease. J. Cell. Physiol. 2021, 236, 3406–3419. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, Z.; Yan, J.; Ying, S. HMGB1 as a Potential Biomarker and Therapeutic Target for Malignant Mesothelioma. Dis. Markers 2019, 2019, 4183157. [Google Scholar] [CrossRef]
- Fukai, E.; Sato, H.; Watanabe, M.; Nakae, D.; Totsuka, Y. Establishment of an in vivo simulating co-culture assay platform for genotoxicity of multi-walled carbon nanotubes. Cancer Sci. 2018, 109, 1024–1031. [Google Scholar] [CrossRef]
- Bonham, C.A.; Strek, M.E.; Patterson, K.C. From granuloma to fibrosis: Sarcoidosis associated pulmonary fibrosis. Curr. Opin. Pulm. Med. 2016, 22, 484–491. [Google Scholar] [CrossRef]
- Mornex, J.; Leroux, C.; Greenland, T.; Ecochard, D. From granuloma to fibrosis in interstitial lung diseases: Molecular and cellular interactions. Eur. Respir. J. 1994, 7, 779–785. [Google Scholar] [CrossRef]
- Abdelgied, M.; El-Gazzar, A.M.; Alexander, W.T.; Numano, T.; Iigou, M.; Naiki-Ito, A.; Takase, H.; Hirose, A.; Taquahashi, Y.; Kanno, J.; et al. Carcinogenic effect of potassium octatitanate (POT) fibers in the lung and pleura of male Fischer 344 rats after intrapulmonary administration. Part. Fibre Toxicol. 2019, 16, 34. [Google Scholar] [CrossRef]
- Numano, T.; Higuchi, H.; Alexander, D.B.; Alexander, W.T.; Abdelgied, M.; El-Gazzar, A.M.; Saleh, D.; Takase, H.; Hirose, A.; Naiki-Ito, A.; et al. MWCNT-7 administered to the lung by intratracheal instillation induces development of pleural mesothelioma in F344 rats. Cancer Sci. 2019, 110, 2485–2492. [Google Scholar] [CrossRef] [PubMed]
- Suzui, M.; Futakuchi, M.; Fukamachi, K.; Numano, T.; Abdelgied, M.; Takahashi, S.; Ohnishi, M.; Omori, T.; Tsuruoka, S.; Hirose, A.; et al. Multiwalled carbon nanotubes intratracheally instilled into the rat lung induce development of pleural malignant mesothelioma and lung tumors. Cancer Sci. 2016, 107, 924–935. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Male | ||||
| Study period | Exposure Conc. (mg/m3) | Exposure Times (Number of days) | Concentration × number of exposures | Lung burden (μg)/Whole Lung (mean ± SD) |
| One-day | 5 | 1 | 5 | 4.6 ± 0.8 |
| 0.2 | 10 | 2 | 2.3 ± 0.4 | |
| 2-week | 1 | 10 | 10 | 9.9 ± 1.2 |
| 5 | 10 | 50 | 43.4 ± 9.3 | |
| 0.2 | 62 | 12.4 | 9.7 ± 0.8 | |
| 13-week | 1 | 62 | 62 | 63.6 ± 3.9 |
| 5 | 62 | 310 | 360.9 ± 51.9 | |
| 104-week (2-year) | 0.02 | 492 | 9.84 | 10.0 ± 2.6 |
| 0.2 | 492 | 98.4 | 152.4 ± 19.4 | |
| 2 | 492 | 984 | 1797.8 ± 146.0 | |
| Female | ||||
| Study period | Exposure Conc. (mg/m3) | Exposure Times (Number of days) | Concentration × number of exposures | Lung burden (μg)/Whole Lung (mean ± SD) |
| 0.2 | 10 | 2 | 2.2 ± 0.4 | |
| 2-week | 1 | 10 | 10 | 8.7 ± 2.3 |
| 5 | 10 | 50 | 32.5 ± 9.7 | |
| 0.2 | 62 | 12.4 | 6.9 ± 0.7 | |
| 13-week | 1 | 62 | 62 | 41.1 ± 6.6 |
| 5 | 62 | 310 | 240.9 ± 41.1 | |
| 104-week (2-year) | 0.02 | 492 | 9.84 | 8.1 ± 1.8 |
| 0.2 | 492 | 98.4 | 118.4 ± 13.4 | |
| 2 | 492 | 984 | 1154.1 ± 77.6 |
| Males | |||
| Lung Burden | Total Protein | LDH | |
| Total Exposure | 0.99 | 0.97 | 0.93 |
| Lung Burden | 0.95 | 0.71 | |
| ALP | 0.87 | ||
| Females | |||
| Lung Burden | Total Protein | LDH | |
| Total Exposure | 0.99 | 0.99 | 0.81 |
| Lung Burden | 0.91 | 0.75 | |
| ALP | 0.95 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kasai, T.; Fukushima, S. Exposure of Rats to Multi-Walled Carbon Nanotubes: Correlation of Inhalation Exposure to Lung Burden, Bronchoalveolar Lavage Fluid Findings, and Lung Morphology. Nanomaterials 2023, 13, 2598. https://doi.org/10.3390/nano13182598
Kasai T, Fukushima S. Exposure of Rats to Multi-Walled Carbon Nanotubes: Correlation of Inhalation Exposure to Lung Burden, Bronchoalveolar Lavage Fluid Findings, and Lung Morphology. Nanomaterials. 2023; 13(18):2598. https://doi.org/10.3390/nano13182598
Chicago/Turabian StyleKasai, Tatsuya, and Shoji Fukushima. 2023. "Exposure of Rats to Multi-Walled Carbon Nanotubes: Correlation of Inhalation Exposure to Lung Burden, Bronchoalveolar Lavage Fluid Findings, and Lung Morphology" Nanomaterials 13, no. 18: 2598. https://doi.org/10.3390/nano13182598