
Engineered Nanoparticles, Natural Nanoclay and Biochar, as Carriers of Plant-Growth Promoting Bacteria
 , , , , , , ,
, , , , , , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Synthesis of Chitosan-Coated Mesoporous Silica Nanoparticles (MSN) and Characterization of Nanoclay (NC) and MSN
2.3. Characterization of Biochars
2.4. Characterization of Soil Substrate and Fertilizer
2.5. Adsorption of Bacteria on Nanoparticles and Biochars, Determination of Loading Efficiency and Sample Characterization
2.6. Plant Growth Experimental Design
2.7. Determination of Soil pH after Treatment
2.8. Extraction and Characterization of Bacteria Populations
2.9. Determination of P and N Content in Plant Leaves
2.10. Physiological Endpoints
2.11. Statistical Analysis
3. Results and Discussion
3.1. Synthesis of Chitosan-Coated Mesoporous Silica Nanoparticles (MSN) and Characterization of Nanoclay (NC) and MSN
3.2. Characterization of Biochars
3.3. Adsorption of Bacteria
3.4. Content of P and N in Soil Substrate and Fertilizer
3.5. Content of P and N in Plants’ Leaves
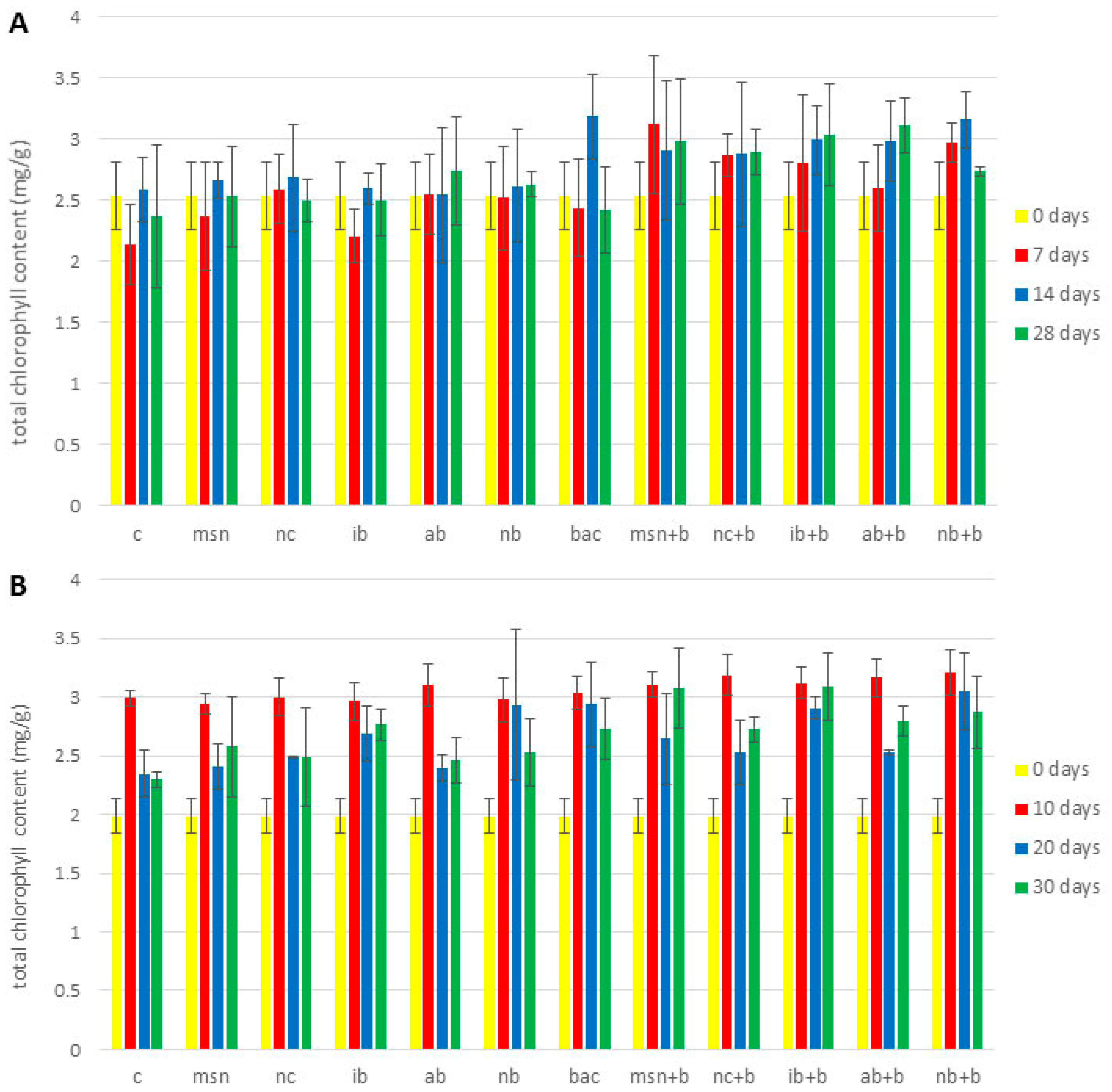
3.6. Chlorophyll Content
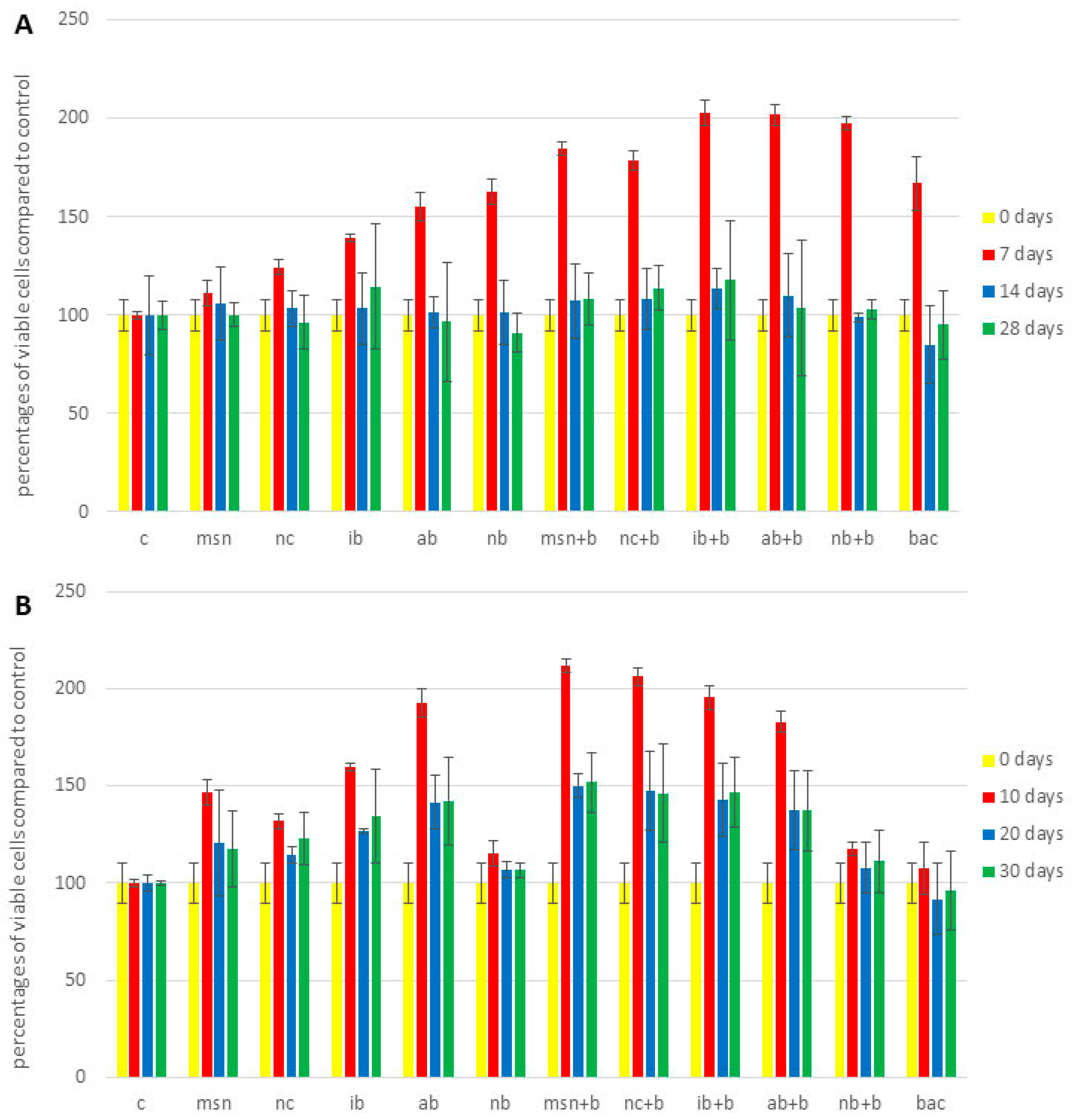
3.7. Plant Cell Viability
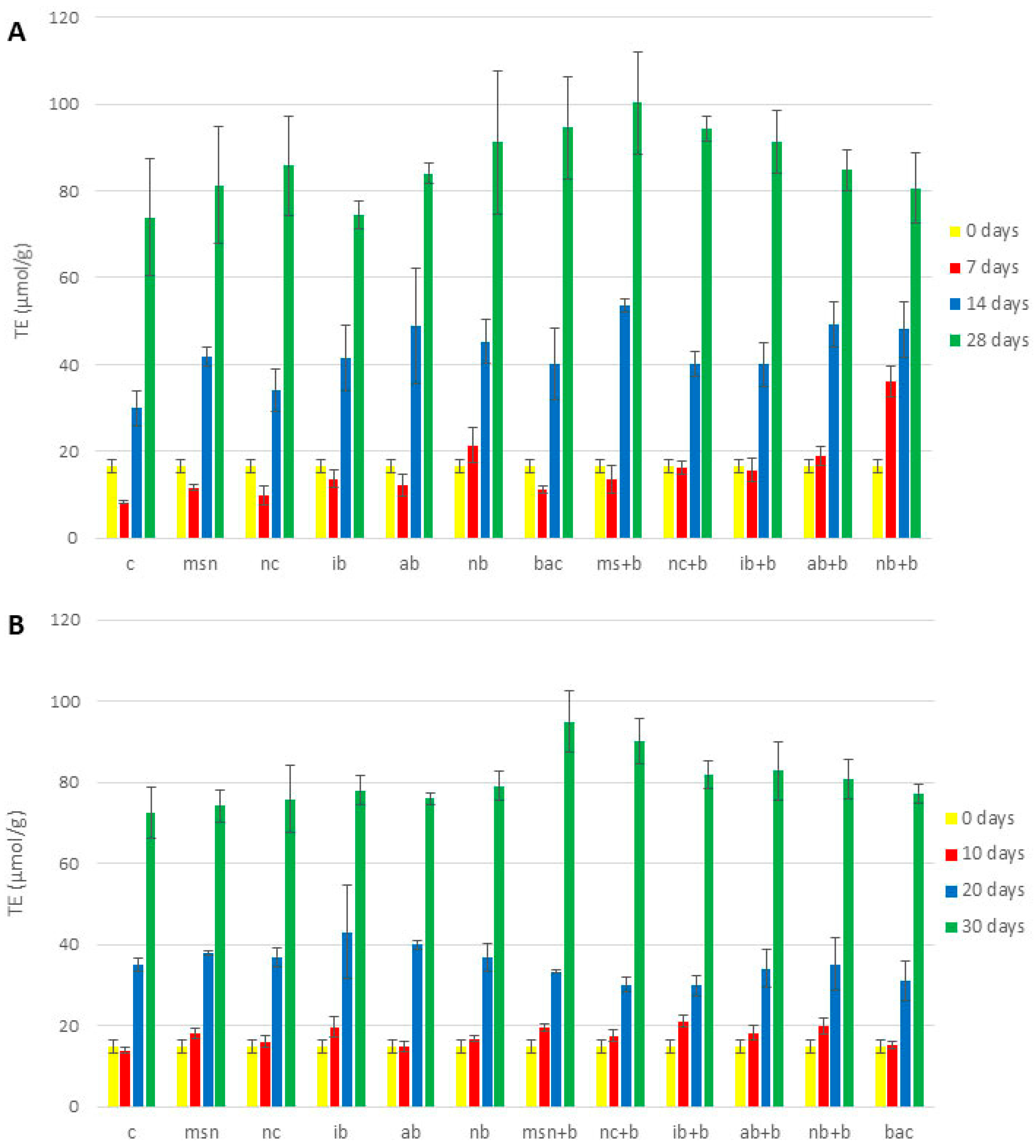
3.8. Antioxidative Properties
3.9. Changes in Soil pH
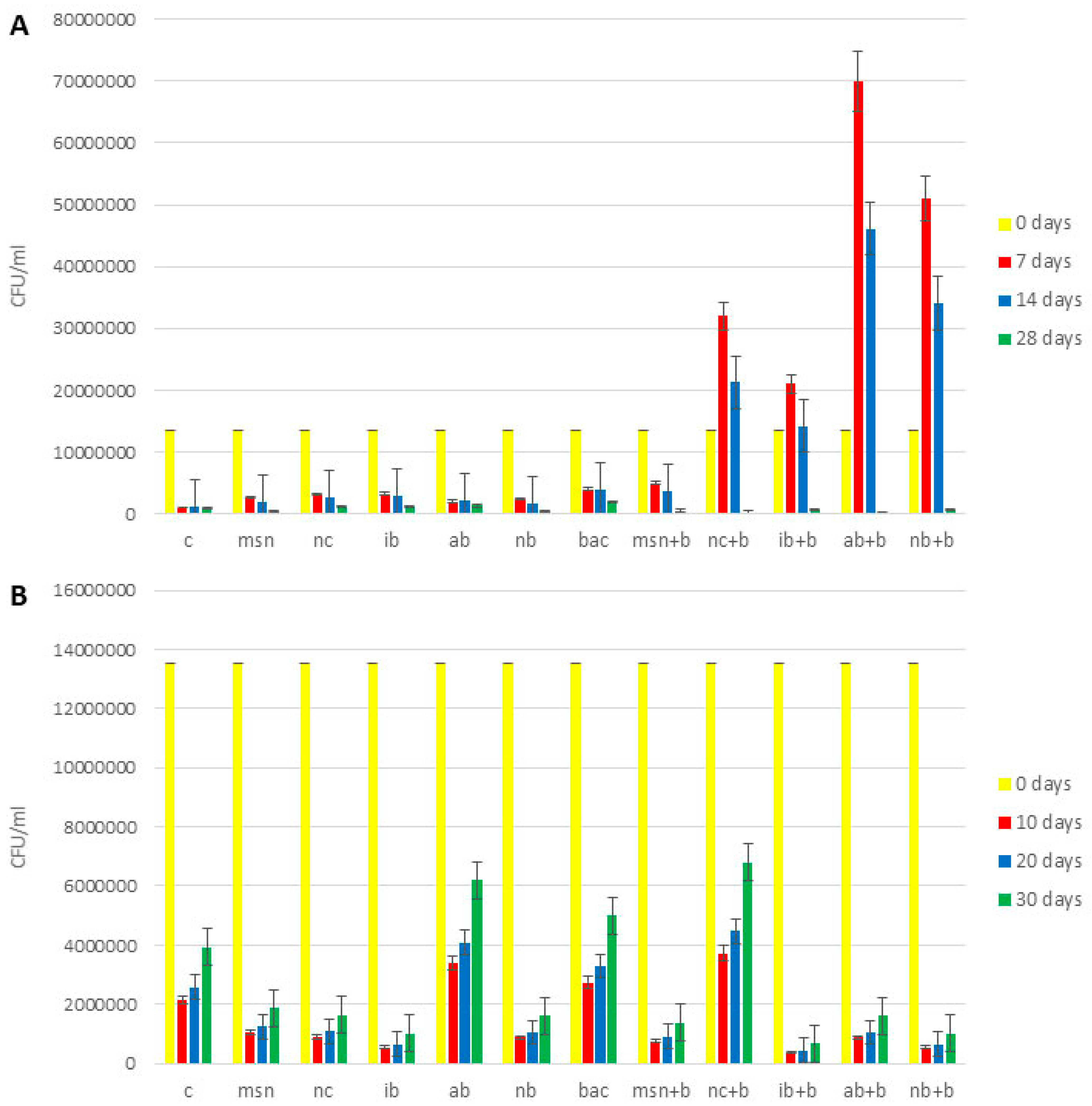
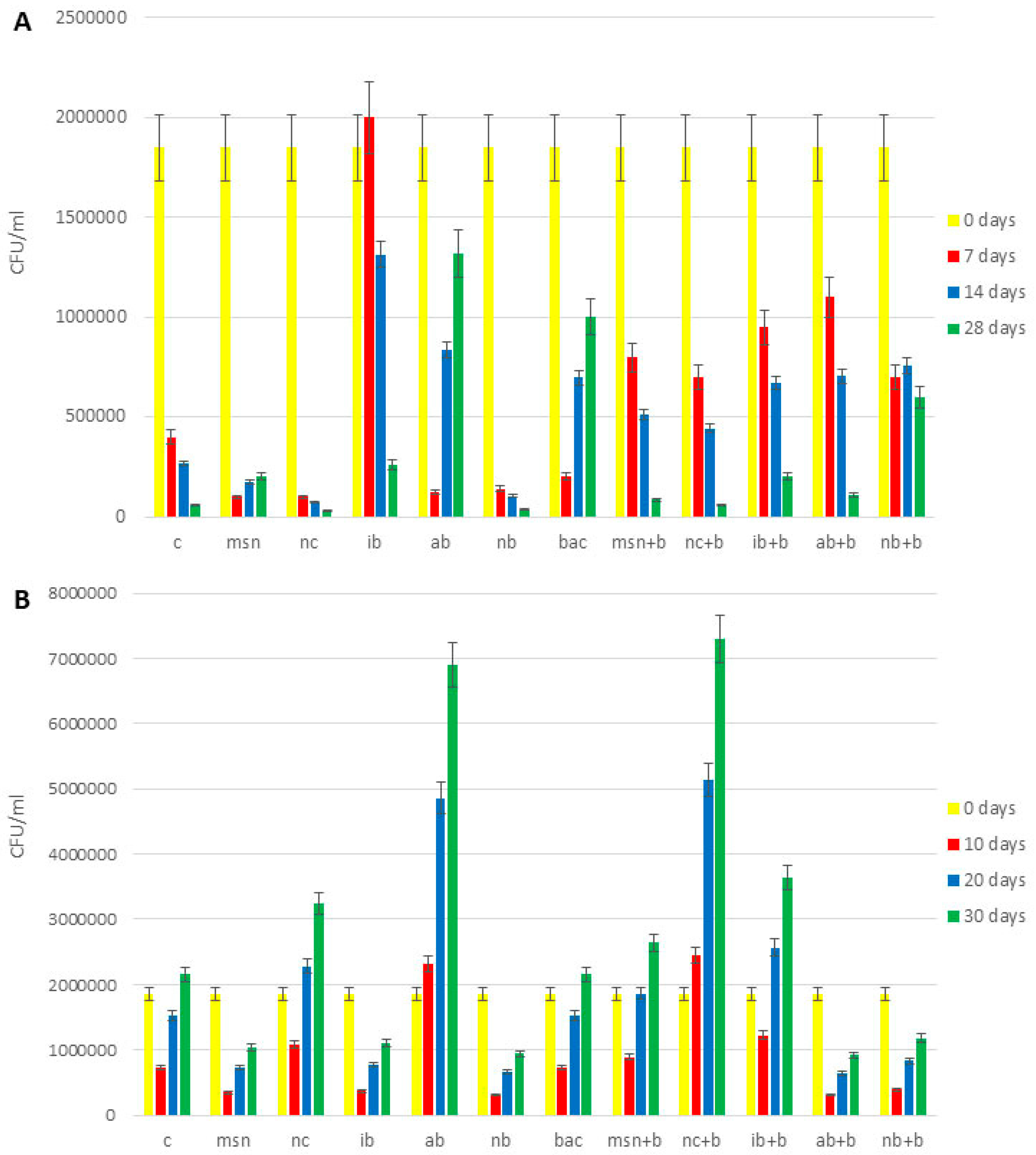
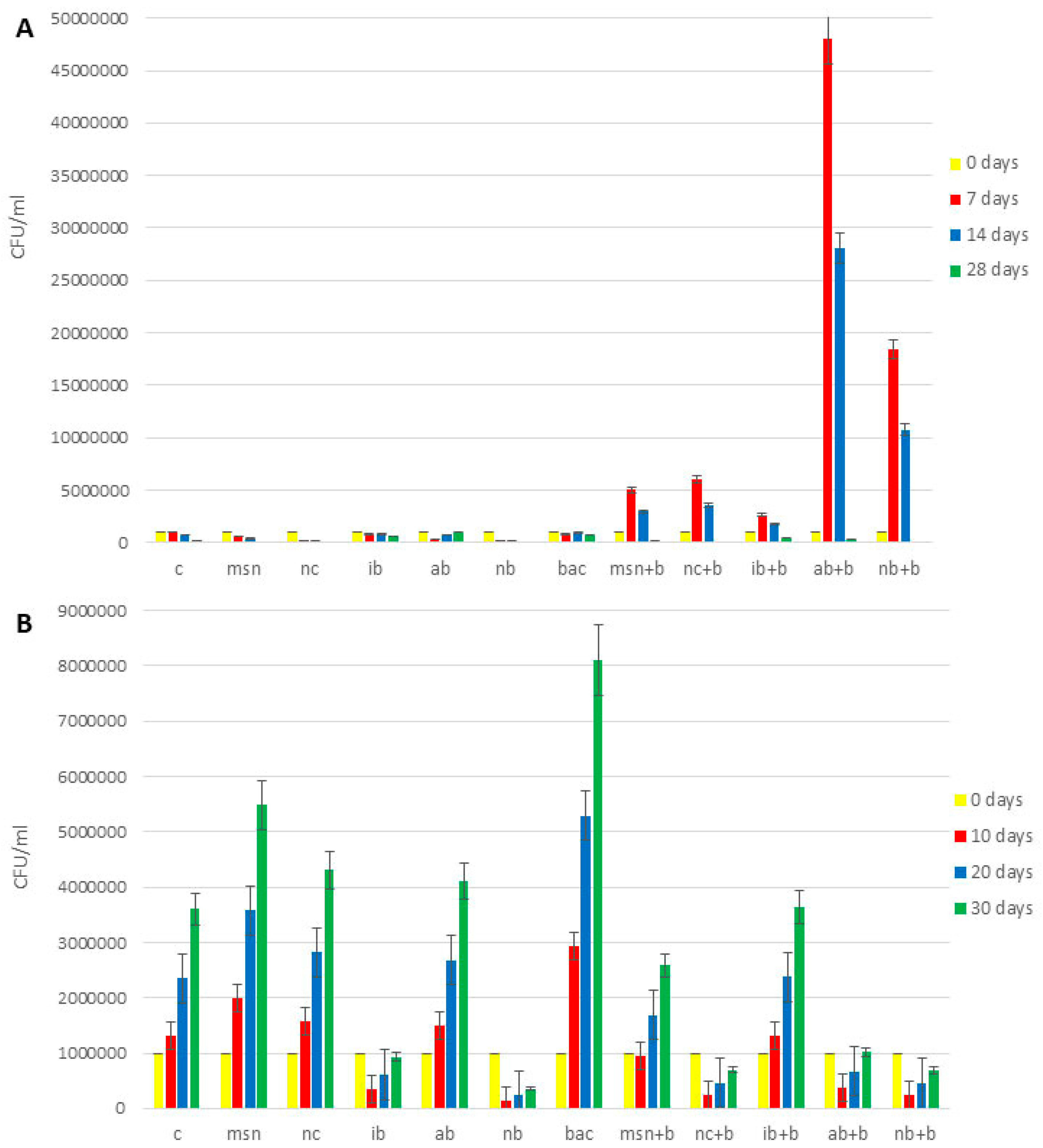
3.10. Microbiological Analysis of Soil
3.11. Plants Biomass
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damalas, C.A.; Koutroubas, S.D. Farmers’ exposure to pesticides: Toxicity types and ways of prevention. Toxics 2016, 4, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lo, C.C. Effect of pesticides on soil microbial community. J. Environ. Sci. Health—B 2020, 45, 348–359. [Google Scholar] [CrossRef]
- Qi, D.; Wu, Q.; Zhu, J. Nitrogen and phosphorus losses from paddy fields and the yield of rice with different water and nitrogen management practices. Sci. Rep. 2020, 10, 9734. [Google Scholar] [CrossRef] [PubMed]
- Savci, S. Investigation of effect of chemical fertilizers on environment. APCBEE Procedia 2012, 1, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Reid, K.; Schneider, K.; McConkey, B. Components of phosphorus loss from agricultural landscapes, and how to incorporate them into risk assessment tools. Front. Earth Sci. 2018, 6, 135. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Lin, M.; Zhou, H.; Wu, H.; Li, Z.; Lin, W. The effects of chemical and organic fertilizer usage on rhizosphere soil in tea orchards. PLoS ONE 2019, 14, e0217018. [Google Scholar] [CrossRef]
- Gil, J.D.B.; Reidsma, P.; Giller, K.; Todman, L.; Whitmore, A.; van Ittersum, M. Sustainable development goal 2: Improved targets and indicators for agriculture and food security. AMBIO 2019, 48, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Antón-Herrero, R.; Vega-Jara, L.; García-Delgado, C.; Mayans, B.; Camacho-Arévalo, R.; Moreno-Jiménez, E.; Plaza, C.; Eymar, E. Synergistic effects of biochar and biostimulants on nutrient and toxic element uptake by pepper in contaminated soils. J. Sci. Food Agric. 2021, 102, 167–174. [Google Scholar] [CrossRef]
- Pan, S.Y.; Dong, C.D.; Su, J.F.; Wang, P.Y.; Chen, C.W.; Chang, J.S.; Kim, H.; Huang, C.P.; Hung, C.M. The role of biochar in regulating the carbon, phosphorus, and nitrogen cycles exemplified by soil systems. Sustainability 2021, 13, 5612. [Google Scholar] [CrossRef]
- Chen, X.; Yang, S.; Ding, J.; Jiang, Z.; Sun, X. Effects of biochar addition on rice growth and yield under water-saving irrigation. Water 2021, 13, 209. [Google Scholar] [CrossRef]
- Yeboah, E.; Asamoah, G.; Ofori, P.; Amoah, B.; Agyeman, K.O.A. Method of biochar application affects growth, yield and nutrient uptake of cowpea. Open Agric. 2020, 5, 352–360. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, R.; Dong, G.; Xiang, M.; Hui, J.; Ou, J.; Qin, H. Biochar nanocomposite derived from watermelon peels for electrocatalytic hydrogen production. ACS Omega 2021, 6, 2066–2073. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Tian, F.; Zou, H.; Ye, Z.; Peng, C.; Huang, J.; Zheng, Y.; Zhang, Y.; Yang, Y.; Wei, X.; et al. ZnO/biochar nanocomposites via solvent free ball milling for enhanced adsorption and photocatalytic degradation of methylene blue. J. Hazard. Mater. 2021, 415, 125511. [Google Scholar]
- Mortazavian, S.; Murph, S.E.H.; Moon, J. Biochar nanocomposite as an inexpensive and highly efficient carbonaceous adsorbent for hexavalent chromium removal. Materials 2022, 15, 6055. [Google Scholar] [CrossRef] [PubMed]
- Ajeng, A.A.; Abdullah, R.; Ling, T.C.; Ismail, S.; Lau, B.F.; Ong, H.C.; Chew, K.W.; Show, P.L.; Chang, J.S. Bioformulation of biochar as a potential inoculant carrier for sustainable agriculture. Environ. Technol. Innov. 2020, 20, 101168. [Google Scholar] [CrossRef]
- Husna, N.; Budianta, D.; Munandar, M.; Napoleon, A. Evaluation of several biochar types as inoculant carrier for indigenous phosphate solubilizing microoorganism from acid sulphate soil. J. Ecol. Eng. 2019, 20, 1–8. [Google Scholar] [CrossRef]
- Kari, A.; Nagymáté, Z.; Romsics, C.; Vajna, B.; Tóth, E.; Lazanyi-Kovács, R.; Rizó, B.; Kutasi, J.; Bernhardt, B.; Farkas, É.; et al. Evaluating the combined effect of biochar and PGPR inoculants on the bacterial community in acidic sandy soil. Appl. Soil Ecol. 2021, 160, 103856. [Google Scholar] [CrossRef]
- Tu, C.; Wei, J.; Guan, F.; Liu, Y.; Sun, Y.; Luo, Y. Biochar and bacteria inoculated biochar enhanced Cd and Cu immobilization and enzymatic activity in a polluted soil. Environ. Int. 2020, 137, 105576. [Google Scholar] [CrossRef]
- Qi, Y.; Liu, H.; Wang, J.; Wang, Y. Effects of different straw biochar combined with microbial inoculants on soil environment in pot experiment. Sci. Rep. 2021, 11, 14685. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Ma, H.; Alimov, J.; Reckling, M.; Wirth, S.; Bellingrath-Kimura, S.D. Response of soybean to hydrochar-based Rhizobium inoculation in loamy sandy soil. Microorganisms 2020, 8, 1674. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Shurigin, V.; Alaylar, B.; Ma, H.; Müller, M.E.H.; Wirth, S.; Reckling, M.; Bellingrath-Kimura, S.D. The effect of biochars and endophytic bacteria on growth and root rot disease incidence of Fusarium infested narrow-leafed lupin (Lupinus angustifolius L.). Microorganisms 2020, 8, 496. [Google Scholar] [CrossRef]
- Hale, L.; Luth, M.; Crowley, D. Biochar characteristics relate to its utility as an alternative soil inoculum carrier to peat and vermiculite. Soil Biol. Biochem. 2015, 81, 228–235. [Google Scholar] [CrossRef]
- Hansen, V.; Müller-Stöver, D.; Imparato, V.; Krogh, P.H.; Jensen, L.S.; Dolmer, A.; Hauggaard-Nielsen, H. The effects of straw or straw-derived gasification biochar applications on soil quality and crop productivity: A farm case study. J. Environ. Manage. 2017, 186, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Tsibart, A.; Nam, H.; Hur, J.; El-Naggar, A.; Tack, F.M.G.; Wang, C.H.; Lee, Y.H.; Tsang, D.C.W.; Ok, Y.S. Effect of gasification biochar application on soil quality: Trace metal behavior, microbial community, and soil dissolved organic matter. J. Hazard. Mater. 2019, 365, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Liu, Y.; Liu, H.; Dumroese, R.K. Interaction of biochar type and rhizobia inoculation increases the growth and biological nitrogen fixation of Robinia pseudoacacia seedlings. Forests 2020, 11, 711. [Google Scholar] [CrossRef]
- Graziano, S.; Caldara, M.; Gullì, M.; Bevivino, A.; Maestri, E.; Marmiroli, N.A. Metagenomic and gene expression analysis in wheat (T. durum) and maize (Z. mays) biofertilized with PGPM and biochar. Int. J. Mol. Sci. 2022, 23, 10376. [Google Scholar] [CrossRef] [PubMed]
- Abdelaziz, A.M.; Dacrory, S.; Hashem, A.H.; Attia, M.S.; Hasanin, M.; Fouda, H.M.; Kamel, S.; El Saied, H. Protective role of zinc oxide nanoparticles based hydrogel against wilt disease of pepper plant. Biocatal. Agric. Biotechnol. 2021, 35, 102083. [Google Scholar] [CrossRef]
- Elmer, W.; Ma, C.; White, J. Nanoparticles for plant disease management. Curr. Opin. Environ Sci Health 2018, 6, 66–70. [Google Scholar] [CrossRef]
- Guo, H.; White, J.C.; Wang, Z.; Xing, B. Nano-enabled fertilizers to control the release and use efficiency of nutrients. Curr. Opin. Environ. Sci. Health 2018, 6, 77–83. [Google Scholar] [CrossRef]
- Paramo, L.A.; Feregrino-Pérez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in agroindustry: Applications, toxicity, challenges, and trends. Nanomaterials 2020, 10, 1654. [Google Scholar] [CrossRef]
- Kah, M.; Kookana, R.S.; Gogos, A.; Bucheli, T.D. A critical evaluation of nanopesticides and nanofertilizers against their conventional analogues. Nat. Nanotechnol. 2018, 13, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; White, J.C.; Zhao, J.; Zhao, Q.; Xing, B. Uptake of engineered nanoparticles by food crops: Characterization, mechanisms, and implications. Annu. Rev. Food Sci. Technol. 2018, 9, 129–153. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Bhattacharyya, A.; Nguyen, Q.D. Nanotechnology in sustainable agriculture: Recent developments, challenges, and perspectives. Front. Microbiol. 2017, 8, 1014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Melendi, P.; Fernández-Pacheco, R.; Coronado, M.J.; Corredor, E.; Testillano, P.S.; Risueño, M.C.; Marquina, C.; Ibarra, M.R.; Rubiales, D.; Pérez-de-Luque, A. Nanoparticles as smart treatment-delivery systems in plants: Assessment of different techniques of microscopy for their visualization in plant tissues. Ann. Bot. 2008, 101, 187–195. [Google Scholar] [CrossRef]
- Sampathkumar, K.; Tan, K.X.; Loo, S.C.J. Developing nano-delivery systems for agriculture and food applications with nature-derived polymers. IScience 2020, 23, 101055. [Google Scholar] [CrossRef]
- Buchman, J.T.; Elmer, W.H.; Ma, C.; Landy, K.M.; White, J.C.; Haynes, C.L. Chitosan-coated mesoporous silica nanoparticle treatment of Citrullus lanatus (watermelon): Enhanced fungal disease suppression and modulated expression of stress-related genes. ACS Sustain. Chem. Eng. 2019, 7, 19649–19659. [Google Scholar] [CrossRef]
- Bolko Seljak, K.; Kocbek, P.; Gašperlin, M. Mesoporous silica nanoparticles as delivery carriers: An overview of drug loading techniques. J. Drug Deliv. Sci. Technol. 2020, 59, 101906. [Google Scholar] [CrossRef]
- Chang, F.P.; Kuang, L.Y.; Huang, C.A.; Jane, W.N.; Huang, Y.; Hsing, Y.C.; Mou, C.Y. A simple plant gene delivery system using mesoporous silica nanoparticles as carriers. J. Mater. Chem. B 2013, 1, 5279–5287. [Google Scholar] [CrossRef]
- Xu, Y.; Xu, C.; Huang, Q.; Cao, L.; Teng, F.; Zhao, P.; Jia, M. Size effect of mesoporous silica nanoparticles on pesticide loading, release, and delivery in cucumber plants. Appl. Sci. 2021, 11, 575. [Google Scholar] [CrossRef]
- Barbosa de Melo, R.; Araújo Ferreira, M.; Araújo Meirelles, L.M.; Zorato, N.; Nervo Raffin, F. Nanoclays in drug delivery systems. In Micro and Nano Technologies, Clay Nanoparticles; Cavallaro, G., Fakhrullin, R., Pasbakhsh., P., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 185–202. [Google Scholar]
- Khatoon, N.; Chu, M.Q.; Zhou, C.H. Nanoclay-based drug delivery systems and their therapeutic potentials. J. Mater. Chem. B 2020, 8, 7335–7351. [Google Scholar] [CrossRef]
- Islam, W.; Tayyab, M.; Khalil, F.; Hua, Z.; Huang, Z.; Chen, H.Y.H. Silicon-mediated plant defense against pathogens and insect pests. Pestic. Biochem. Phys. 2020, 168, 104641. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Nikolic, M.; Bélanger, R.; Gong, H.; Song, A. Silicon in Agriculture; Springer Science: New York, NY, USA, 2015; pp. 181–197. [Google Scholar]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharyya, P.N.; Jha, D.K. Plant growth-promoting rhizobacteria (PGPR): Emergence in agriculture. World J. Microbiol. Biotechnol. 2012, 28, 1327–1350. [Google Scholar] [CrossRef] [PubMed]
- Noar, J.D.; Bruno-Bracena, J.M. Azotobacter vinelandii: The source of 100 years of discoveries and many more to come. Microbiology 2018, 164, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Plunkett, M.H.; Knutson, C.M.; Barney, B.M. Key factors afecting ammonium production by an Azotobacter vinelandii strain deregulated for biological nitrogen fIxation. Microb. Cell Fact. 2020, 19, 107. [Google Scholar] [CrossRef]
- Vary, P.S.; Biedendieck, R.; Fuerch, T.; Meinhardt, F.; Rohde, M.; Deckwer, W.D.; Jahn, D. Bacillus megaterium—From simple soil bacterium to industrial protein production host. Appl. Microbiol. Biotechnol. 2007, 76, 957–967. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, S. Identification and characterization of the phosphate-solubilizing bacterium Pantoea sp. S32 in reclamation soil in Shanxi, China. Front. Microbiol. 2019, 10, 2171. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.M.; Radhakrishnan, R.; You, Y.H.; Joo, G.J.; Lee, I.J.; Lee, K.E.; Kim, J.H. Phosphate solubilizing Bacillus megaterium mj1212 regulates endogenous plant carbohydrates and amino acids contents to promote mustard plant growth. Indian J. Microbiol. 2014, 54, 427–433. [Google Scholar] [CrossRef]
- Marmiroli, M.; Bonas, U.; Imperiale, D.; Lencioni, G.; Mussi, F.; Marmiroli, N.; Maestri, E. Structural and functional features of chars from different biomasses as potential plant amendments. Front. Plant Sci. 2018, 9, 1119. [Google Scholar] [CrossRef] [Green Version]
- Dume, B.; Berecha, G.; Tulu, S. Characterization of biochar produced at different temperature and its effect on acidic Nitosol of Jimma, southwest Ethiopia. Int. J. Soil Sci. 2015, 10, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Batista, E.M.C.C.; Shultz, J.; Matos, T.T.S.; Fornari, M.R.; Ferreira, T.M.; Szpoganicz, B.; de Freitas, R.A.; Mangrich, A.S. Effect of surface and porosity of biochar on water holding capacity aiming indirectly at preservation of the Amazon biome. Sci. Rep. 2018, 8, 10677. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.; Upadhyayula, V.K.K.; Smith, G.B.; Mitchell, M.C. Adsorption equilibrium and kinetics of microorganisms on single-wall carbon nanotubes. IEEE Sens. J. 2008, 8, 954–962. [Google Scholar] [CrossRef]
- Fox, A.; Kwapinski, W.; Griffiths, B.S.; Schmalenberger, A. The role of sulfur- and phosphorus-mobilizing bacteria in biochar-induced growth promotion of Lolium perenne. FEMS Microbiol. Ecol. 2014, 90, 78–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobereiner, J.; Urquiaga, S.; Boddey, R.M. Alternatives for nitrogen nutrition of crops in tropical agriculture. Fert. Res. 1995, 42, 339–346. [Google Scholar] [CrossRef]
- Li, Y.; Liu, C.; Zhang, J.; Yang, H.; Xu, L.; Wang, Q.; Sack, L.; Wu, X.; Hou, J.; He, N. Variation in leaf chlorophyll concentration from tropical to cold-temperate forests: Association with gross primary productivity. Ecol. Indic. 2018, 85, 383–389. [Google Scholar] [CrossRef]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Shoemaker, M.; Cohen, I.; Campbell, M. Reduction of MTT by aqueous herbal extracts in the absence of cells. J. Ethnopharmacol. 2004, 93, 381–384. [Google Scholar] [CrossRef]
- Zemljič, L.F.; Plohl, O.; Vesel, A.; Luxbacher, T.; Potrč, S. Physicochemical characterization of packaging foils coated by chitosan and polyphenols colloidal formulations. Int. J. Mol. Sci. 2020, 21, 495. [Google Scholar] [CrossRef] [Green Version]
- García-Villén, F.; Faccendini, A.; Miele, D.; Ruggeri, M.; Sánchez-Espejo, R.; Borrego-Sánchez, A.; Cerezo, P.; Rossi, S.; Viseras, C.; Sandri, G. Wound healing activity of nanoclay/spring water hydrogels. Pharmaceutics 2020, 12, 467. [Google Scholar] [CrossRef]
- Nam, J.; Yang, S.; Yun, B.Y.; Kim, S. Evaluation of thermal/morphological performance of SSPCM based nanoclay: Influence of the interlayer microstructure of hydrophilic and hydrophobic. Sol. Energy Mater. Sol. Cells 2022, 235, 111479. [Google Scholar] [CrossRef]
- Singh, B.; Dolk, M.M.; Shen, Q.; Camps-Arbestain, M. Biochar pH, electrical conductivity and liming potential. In Biochar: A Guide to Analytical Methods; Singh, B., Camps-Arbestain, M., Lehmann, J., Eds.; CSIRO Publishing: Clayton, Australia, 2017; pp. 23–38. [Google Scholar]
- Fryda, L.; Visser, H. Biochar for soil improvement: Evaluation of biochar from gasification and slow pyrolysis. Agriculture 2015, 5, 1076–1115. [Google Scholar] [CrossRef] [Green Version]
- Tomczyk, A.; Sokołowska, Z.; Boguta, P. Biochar physicochemical properties: Pyrolysis temperature and feedstock kind effects. Rev. Environ. Sci. Biotechnol. 2020, 19, 191–215. [Google Scholar] [CrossRef] [Green Version]
- Sigmund, G.; Hüffer, T.; Hofmann, T.; Kah, M. Biochar total surface area and total pore volume determined by N2 and CO2 physisorption are strongly influenced by degassing temperature. Sci. Total Environ. 2017, 580, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Piash, M.I.; Iwabuchi, K.; Itoh, T.; Uemura, K. Release of essential plant nutrients from manure- and wood-based biochars. Geoderma 2021, 397, 115100. [Google Scholar] [CrossRef]
- Jastrzębska, A.M.; Karwowska, E.; Olszyna, A.R.; Kunicki, A. Influence of bacteria adsorption on zeta potential of Al2O3 and Al2O3/Ag nanoparticles in electrolyte and drinking water environment studied by means of zeta potential. Surf. Coat. Technol. 2015, 271, 225–233. [Google Scholar] [CrossRef]
- Darabdhara, G.; Boruah, P.K.; Hussain, N.; Borthakur, P.; Sharma, B.; Sengupta, P.; Das, M.R. Magnetic nanoparticles towards efficient adsorption of gram positive and gram negative bacteria: An investigation of adsorption parameters and interaction mechanism. Colloids Surf. A Physicochem. Eng. Asp. 2017, 516, 161–170. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Spilles, A.; Bonkowski, M. C: N: P stoichiometry and nutrient limitation of the soil microbial biomass in a grazed grassland site under experimental P limitation or excess. Ecol. Process. 2012, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Vitòria, L.; Otero, N.; Soler, A.; Canals, A. Fertilizer characterization: Isotopic data (N, S, O, C, and Sr). Environ. Sci. Technol. 2004, 38, 3254–3262. [Google Scholar] [CrossRef] [PubMed]
- Aasfar, A.; Bargaz, A.; Yaakoubi, K.; Hilali, A.; Bennis, I.; Zeroual, Y.; Meftah Kadmiri, I. Nitrogen fixing Azotobacter species as potential soil biological enhancers for crop nutrition and yield stability. Front. Microbiol. 2021, 12, 628379. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Mao, X.; Zhang, M.; Yang, W.; Di, H.J.; Ma, L.; Liu, W.; Li, B. The application of Bacillus megaterium alters soil microbial community composition, bioavailability of soil phosphorus and potassium, and cucumber growth in the plastic shed system of North China. Agric. Ecosyst. Environ. 2021, 307, 107236. [Google Scholar] [CrossRef]
- Terry, N.; Ulrich, A. Effects of phosphorus deficiency on the photosynthesis and respiration of leaves of sugar beet. Plant Physiol. 1973, 51, 43–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciompi, S.; Gentili, E.; Guidi, L.; Soldatini, G.F. The effect of nitrogen deficiency on leaf gas exchange and chlorophyll fluorescence parameters in sunflower. Plant Sci. 1996, 118, 177–184. [Google Scholar] [CrossRef]
- Paul, M.J.; Stitt, M. Effects of nitrogen and phosphorus deficiencies on levels of carbohydrates, respiratory enzymes and metabolites in seedlings of tobacco and their response to exogenous sucrose. Plant Cell Environ. 1993, 16, 1047–1057. [Google Scholar] [CrossRef]
- Hou, Z.; Tang, Y.; Li, C.; Lim, K.J.; Wang, Z. The additive effect of biochar amendment and simulated nitrogen deposition stimulates the plant height, photosynthesis and accumulation of NPK in pecan (Carya illinoinensis) seedlings. AoB PLANTS 2020, 12, plaa035. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.R.; Wei, X.Q.; Song, X.; Hao, L.Y.; Cai, X.X.; Zhang, Z.R.; Peng, Q.; Lin, Y.F. Independent effect of polymeric nanoparticle zeta potential/surface charge, on their cytotoxicity and affinity to cells. Cell Prolif. 2015, 48, 465–474. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Teng, X.; Chen, D.; Tang, F.; He, J. The effect of the shape of mesoporous silica nanoparticles on cellular uptake and cell function. Biomaterials 2010, 31, 438–448. [Google Scholar] [CrossRef]
- Cui, Y.; Mao, F.; Zhang, J.; He, Y.; Tong, Y.W.; Peng, Y. Biochar enhanced high-solid mesophilic anaerobic digestion of food waste: Cell viability and methanogenic pathways. Chemosphere 2021, 272, 129863. [Google Scholar] [CrossRef]
- Litan, A.; Langhans, S.A. Cancer as a channelopathy: Ion channels and pumps in tumor development and progression. Front. Cell Neurosci. 2015, 9, 86. [Google Scholar] [CrossRef] [Green Version]
- Wołosiak, R.; Drużyńska, B.; Derewiaka, D.; Piecyk, M.; Majewska, E.; Ciecierska, M.; Worobiej, E.; Pakosz, P. Verification of the conditions for determination of antioxidant activity by ABTS and DPPH assays-A practical approach. Molecules 2021, 27, 50. [Google Scholar] [CrossRef]
- Sadeghnia, H.R.; Zoljalali, N.; Hanafi-Bojd, M.Y.; Nikoofal-Sahlabadi, S.; Malaekeh-Nikouei, B. Effect of mesoporous silica nanoparticles on cell viability and markers of oxidative stress. Toxicol Mech Methods. 2015, 25, 433–439. [Google Scholar] [PubMed]
- Lee, S.; Yun, H.-S.; Kim, S.-H. The comparative effects of mesoporous silica nanoparticles and colloidal silica on inflammation and apoptosis. Biomaterials 2011, 32, 9434–9443. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Raihan, M.R.H.; Khojah, E.; Samra, B.N.; Fujita, M.; Nahar, K. Biochar and chitosan regulate antioxidant defense and methylglyoxal detoxification systems and enhance salt tolerance in Jute (Corchorus olitorius L.). Antioxidants 2021, 10, 2017. [Google Scholar] [CrossRef] [PubMed]
- Hinsinger, P.; Plassard, C.; Tang, C.; Jaillard, B. Origins of root-mediated pH changes in the rhizosphere and their responses to environmental constraints: A review. Plant Soil 2003, 248, 43–59. [Google Scholar] [CrossRef]
- Zhang, M.; Riaz, M.; Zhang, L.; El-desouki, Z.; Jiang, C. Biochar induces changes to basic soil properties and bacterial communities of different soils to varying degrees at 25 mm rainfall: More effective on acidic soils. Front. Microbiol. 2019, 10, 1321. [Google Scholar] [CrossRef] [Green Version]
- Martinsen, V.; Alling, V.; Nurida, N.L.; Mulder, J.; Hale, S.E.; Ritz, C.; Rutherford, D.W.; Heikens, A.; Breedveld, G.D.; Cornelissen, G. pH effects of the addition of three biochars to acidic Indonesian mineral soils. Soil Sci. Plant Nut. 2015, 61, 821–834. [Google Scholar] [CrossRef] [Green Version]
- Alan, Ö.; Sen, F.; Düzyaman, E. The effectiveness of growth cycles on improving fruit quality for grafted watermelon combinations. Food Sci. Techol. 2017, 38, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Shamshiri, R.R.; Jones, J.W.; Thorp, K.R.; Ahmad, D.; Man, H.C.; Taheri, S. Review of optimum temperature, humidity, and vapour pressure deficit for microclimate evaluation and control in greenhouse cultivation of tomato: A review. Int. Agrophys. 2018, 32, 287–302. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (C) | ||
|---|---|---|
| Treatment with Bacteria (BAC) | ||
| chitosan-coated mesoporous silica | treatment with chitosan-coated mesoporous silica (MSN) | treatment with chitosan- coated mesoporous silica with loaded bacteria (MSN + B) |
| nanoclay | treatment with nanoclay (NC) | treatment with nanoclay with loaded bacteria (NC + B) |
| “Italian” biochar | treatment with “Italian” biochar (IB) | treatment with “Italian” biochar with loaded bacteria (IB + B) |
| Aries biochar | treatment with Aries green biochar (AB) | treatment with Aries Green biochar with loaded bacteria (AB + B) |
| Naked biochar | treatment with Naked biochar (NB), | treatment with Naked biochar with loaded bacteria (NB + B). |
| Sample | Tomato Experiment P Content (mg/g) | Watermelon Experiment P Content (mg/g) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 Days | 7 Days | 14 Days | 28 Days | 0 Days | 10 Days | 20 Days | 30 Days | |
| C | 9.3 ± 0.5 a | 3.6 ± 0.4 a | 3.5 ± 0.2 a | 3.0 ± 0.4 a | 4.3 ± 0.6 a | 4.7 ± 0.5 a | 1.9 ± 0.3 a | 1.6 ± 0.2 a |
| MSN | 9.3 ± 0.5 a | 3.7 ± 0.3 a | 3.9 ± 0.1 b | 3.3 ± 0.3 a | 4.3 ± 0.6 a | 3.2 ± 0.4 b | 1.9 ± 0.1 a | 1.8 ± 0.3 a |
| NC | 9.3 ± 0.5 a | 3.6 ± 0.3 a | 4.7 ± 0.2 c | 3.3 ± 0.5 a | 4.3 ± 0.6 a | 2.9 ± 0.5 b | 2.4 ± 0.5 b | 1.6 ± 0.2 a |
| IB | 9.3 ± 0.5 a | 3.7 ± 0.2 a | 4.3 ± 0.4 b,c | 3.4 ± 0.4 a | 4.3 ± 0.6 a | 3.7 ± 0.4 c | 2.9 ± 0.6 b | 2.1 ± 0.3 b |
| AB | 9.3 ± 0.5 a | 3.4 ± 0.4 a | 5.0 ± 0.5 c | 3.5 ± 0.3 a,b | 4.3 ± 0.6 a | 3.5 ± 0.b,c | 2.6 ± 0.5 b | 2.0 ± 0.2 b |
| NB | 9.3 ± 0.5 a | 3.2 ± 0.6 a | 4.7 ± 0.3 c | 3.9 ± 0.4 b | 4.3 ± 0.6 a | 3.6 ± 0.5 b,c | 2.4 ± 0.1 b | 1.8 ± 0.3 a |
| BAC | 9.3 ± 0.5 a | 5.0 ± 0.4 b | 6.5 ± 0.7 d | 4.3 ± 0.4 b,c | 4.3 ± 0.6 a | 3.8 ± 0.4 c | 2.8 ± 0.4 b | 2.3 ± 0.4 b |
| MSN + B | 9.3 ± 0.5 a | 4.6 ± 0.3 a,b | 4.3 ± 0.2 b,c | 3.7 ± 0.2 b | 4.3 ± 0.6 a | 2.9 ± 0.5 b | 2.0 ± 0.6 a | 1.9 ± 0.1 b |
| NC + B | 9.3 ± 0.5 a | 4.8 ± 0.3 a,b | 4.5 ± 0.5 b,c | 3.6 ± 0.5 a,b | 4.3 ± 0.6 a | 3.2 ± 0.2 b | 2.4 ± 0.4 b | 1.9 ± 0.3 a,b |
| IB + B | 9.3 ± 0.5 a | 4.0 ± 0.4 a | 5.2 ± 0.8 c | 3.9 ± 0.3 b | 4.3 ± 0.6 a | 3.6 ± 0.8 b,c | 2.5 ± 0.6 b | 1.9 ± 0.1 b |
| AB + B | 9.3 ± 0.5 a | 4.3 ± 0.4 a | 4.7 ± 0.6 b | 3.8 ± 0.4 a,b | 4.3 ± 0.6 a | 3.9 ± 0.8 c | 2.1 ± 0.3 a | 2.0 ± 0.1 b |
| NB + B | 9.3 ± 0.5 a | 3.8 ± 0.5 a | 5.0 ± 0.6 b,c | 4.8 ± 0.7 b,c | 4.3 ± 0.6 a | 3.6 ± 0.2 b,c | 2.8 ± 0.5 b | 2.1 ± 0.4 b |
| Sample | Tomato Experiment N Content (mg/g) | Watermelon Experiment N Content (mg/g) | ||||||
|---|---|---|---|---|---|---|---|---|
| 0 Days | 7 Days | 14 Days | 28 Days | 0 Days | 10 Days | 20 Days | 30 Days | |
| C | 57.1 ± 1.2 a | 54.8 ± 2.0 a | 41.4 ± 2.3 a | 22.0 ± 3.2 a | 29.9 ± 3.1 a | 25.4 ± 1.4 a | 18.5± 0.3 a | 17.7 ± 1.5 a |
| MSN | 57.1 ± 1.2 a | 44.7 ± 1.7 b | 39.1 ± 1.3 a | 22.9 ± 1.8 a | 29.9 ± 3.1 a | 28.2 ± 4.6 a | 19.8 ± 1.1 a | 20.0 ± 2.4 a |
| NC | 57.1 ± 1.2 a | 42.6 ± 0.8 c | 38.8 ± 2.8 a | 23.4 ± 2.6 a | 29.9 ± 3.1 a | 24.9 ± 2.5 a | 20.2 ± 1.6 a | 21.9 ± 2.6 a |
| IB | 57.1 ± 1.2 a | 46.0 ± 4.3 b | 39.7 ± 2.5 a | 22.2 ± 3.1 a | 29.9 ± 3.1 a | 27.2 ± 2.1 a | 20.8 ± 0.4 a | 19.0 ± 2.4 a |
| AB | 57.1 ± 1.2 a | 51.7 ± 0.9 d | 36.5 ± 2.2 b | 22.3 ± 3.6 a | 29.9 ± 3.1 a | 24.8 ± 3.0 a | 18.8 ± 0.3 a | 18.7 ± 1.6 a |
| NB | 57.1 ± 1.2 a | 55.8 ± 4.6 a | 34.9 ± 3.1 b | 28.8 ± 3.3 b | 29.9 ± 3.1 a | 28.5 ± 0.7 a | 20.2 ± 0.4 a | 19.3 ± 2.2 a |
| BAC | 57.1 ± 1.2 a | 53.5 ± 4.1 a | 44.5 ± 2.4 a | 25.2 ± 2.4 c | 29.9 ± 3.1 a | 25.6 ± 2.9 a | 25.4 ± 2.9 b | 21.6 ± 0.8 c |
| MSN + B | 57.1 ± 1.2 a | 47.4 ± 3.1 b | 38.4 ± 2.3 a | 24.1 ± 2.2 c | 29.9 ± 3.1 a | 27.5 ± 3.4 a | 20.7 ± 3.4 a | 22.2 ± 2.4 b,c |
| NC + B | 57.1 ± 1.2 a | 48.3 ± 1.2 d | 38.9 ± 2.2 a | 22.2 ± 1.1 a | 29.9 ± 3.1 a | 23.2 ± 3.0 a | 18.3 ± 3.0 a | 21.6 ± 2.6 b |
| IB + B | 57.1 ± 1.2 a | 44.4 ± 4.1 b,c | 39.0± 1.5 a | 28.3 ± 1.0 c,d | 29.9 ± 3.1 a | 27.8 ± 2.2 a | 19.9 ± 2.2 a | 20.9 ± 1.8 b,c |
| AB + B | 57.1 ± 1.2 a | 51.7 ± 2.4 d | 38.9 ± 3.2 a | 32.2 ± 3.7 d,e | 29.9 ± 3.1 a | 29.9 ± 3.9 a | 19.5 ± 3.9 a | 21.6 ± 2.8 b |
| NB + B | 57.1 ± 1.2 a | 53.2 ± 4.7 a | 41.5 ± 2.0 a | 33.7 ± 4.3 d,e | 29.9 ± 3.1 a | 28.7 ± 3.3 a | 20.7 ± 3.3 a | 24.1 ± 0.6 c |
| Sample | Tomato Experiment | Watermelon Experiment | ||||
|---|---|---|---|---|---|---|
| Total Mass (g) | Root Mass (g) | Stem Length (cm) | Total Mass (g) | Root Mass (g) | Stem Length (cm) | |
| C | 18.35 ± 2.09 a | 5.97 ±1.14 a | 23.66 ± 2.31 a | 10.69 ± 2.16 a | 1.25 ± 0.29 a | 37.97 ± 3.61 a |
| MSN | 19.15 ± 2.74 a | 6.48 ± 1.25 a | 26.75 ± 1.26 b | 10.78 ± 0.63 a | 1.09 ± 0.22 a | 43.65 ± 4.97 b |
| NC | 18.37 ± 2.41 a | 6.51 ± 0.84 a | 26.32 ± 2.77 b | 11.36 ± 2.11 a | 0.97 ± 0.11 a | 53.16 ± 5.43 c |
| IB | 18.69 ± 2.63 a | 5.99 ± 0.68 a | 25.01 ± 2.49 a | 10.82 ± 0.95 a | 1.04 ± 0.14 a | 45.46 ± 3.48 b |
| AB | 19.05 ± 2.51 a | 6.08 ± 0.76 a | 25.61 ± 2.19 a | 11.22 ± 0.91 a | 0.95 ± 0.12 a | 52.30 ± 6.14 c |
| NB | 18.46 ± 1.95 a | 6.35 ± 1.16 a | 24.84 ± 3.10 a | 11.86 ± 1.66 a | 1.03 ± 0.20 a | 47.30 ± 4.05 b |
| BAC | 18.19 ± 1.76 a | 5.91 ± 0.56 a | 22.86 ± 2.36 a | 11.02 ± 1.26 a | 0.75 ± 0.16 a | 48.35 ± 4.41 d |
| MSN + B | 21.99 ± 1.51 b | 5.99 ± 1.13 a | 27.54 ± 2.31 c | 11.89 ± 0.59 b | 0.92 ± 0.19 a | 45.22 ± 4.26 b |
| NC + B | 19.65 ± 1.72 a | 5.62 ± 0.70 a | 27.50 ± 2.24 c | 12.65 ± 2.02 b | 0.89 ± 0.17 a | 55.45 ± 3.02 c |
| IB + B | 20.52 ± 2.55 b | 6.53 ± 0.66 a | 25.58 ± 1.49 b | 12.11 ± 1.09 b | 0.74 ± 0.11 a | 50.48 ± 4.74 d |
| AB + B | 19.87 ± 1.07 a | 5.81 ± 0.67 a | 26.90 ± 3.19 b | 13.68 ± 1.61 b | 0.94 ± 0.18 a | 56.64 ± 4.01 c |
| NB + B | 18.93 ± 1.84 a | 5.93 ± 1.21 a | 23.81 ± 1.98 a | 12.28 ± 1.89 b | 0.72 ± 0.08 a | 50.18 ± 4.23 d |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlicevic, M.; Abdelraheem, W.; Zuverza-Mena, N.; O’Keefe, T.; Mukhtar, S.; Ridge, G.; Ranciato, J.; Haynes, C.; Elmer, W.; Pignatello, J.; et al. Engineered Nanoparticles, Natural Nanoclay and Biochar, as Carriers of Plant-Growth Promoting Bacteria. Nanomaterials 2022, 12, 4474. https://doi.org/10.3390/nano12244474
Pavlicevic M, Abdelraheem W, Zuverza-Mena N, O’Keefe T, Mukhtar S, Ridge G, Ranciato J, Haynes C, Elmer W, Pignatello J, et al. Engineered Nanoparticles, Natural Nanoclay and Biochar, as Carriers of Plant-Growth Promoting Bacteria. Nanomaterials. 2022; 12(24):4474. https://doi.org/10.3390/nano12244474
Chicago/Turabian StylePavlicevic, Milica, Wael Abdelraheem, Nubia Zuverza-Mena, Tana O’Keefe, Salma Mukhtar, Gale Ridge, John Ranciato, Christy Haynes, Wade Elmer, Joseph Pignatello, and et al. 2022. "Engineered Nanoparticles, Natural Nanoclay and Biochar, as Carriers of Plant-Growth Promoting Bacteria" Nanomaterials 12, no. 24: 4474. https://doi.org/10.3390/nano12244474