
Interaction of Rhus typhina Tannin with Lipid Nanoparticles: Implication for the Formulation of a Tannin–Liposome Hybrid Biomaterial with Antibacterial Activity


 , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
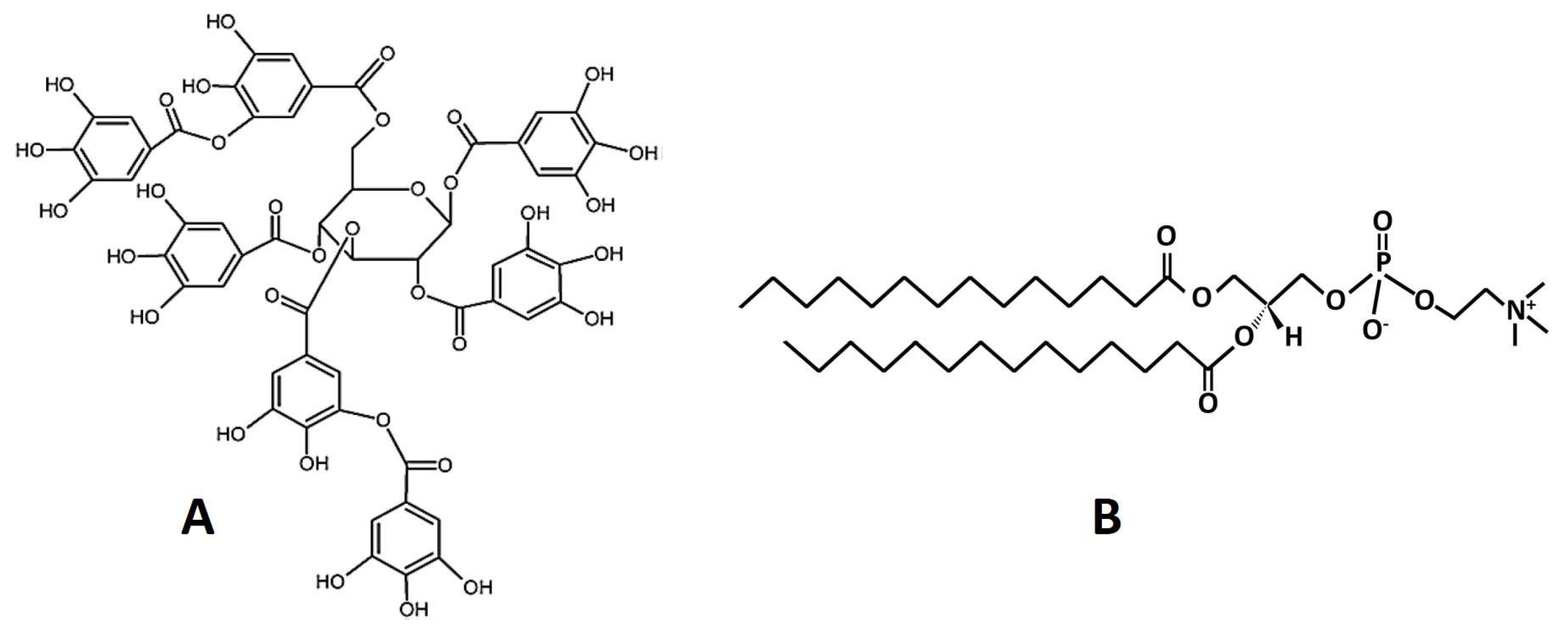
2.1. Materials
2.2. Liposome Preparation
2.3. Analysis of ζ-Potential and Particle Size—Light Scattering
2.4. Preparation of Lipid Bilayers
2.5. Differential Scanning Calorimetry (DSC)
2.6. RT–DMPC Interaction Studies: Fluorescence Analysis of Lipid Order, Nanodomain Formation, and Biophysical Parameters
2.7. Antibacterial Activity of RT–DMPC Hybrid Nanosystems
3. Results and Discussion
3.1. Physicochemical and Thermodynamic Characterization of Liposomes That Contain RT
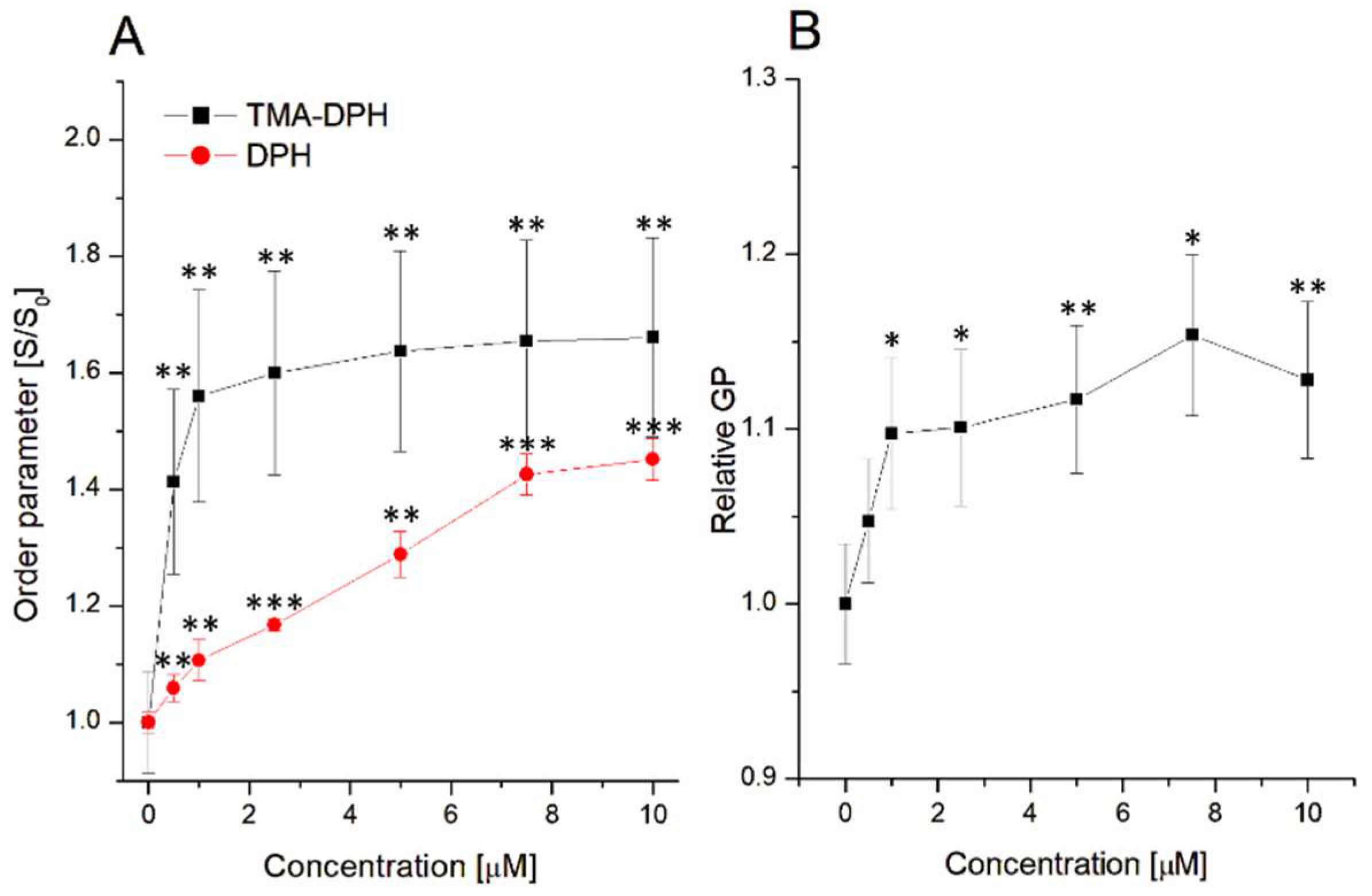
3.2. Influence on Lipid Order Parameter and Lipid Nanodomain Formation
3.3. Fluorescence Analysis of Sumac–Liposome Interactions
- F is the fluorescence in the presence of the quencher;
- KSV is the Stern–Volmer constant;
- [Q] is the quencher concentration.
- KSV is the Stern–ˆVolmer constant;
- τ0 is the average lifetime of fluorophore molecules (5 × 10−9 s).
- F0 is the fluorescence observed in absence of quencher;
- F is the fluorescence observed in presence of quencher;
- KSVa is the effective Stern–Volmer constant for the accessible fluorophores;
- fa is the fraction of accessible fluorophore.
- F0 is the fluorescence observed in absence of quencher molecules;
- F is the fluorescence observed in presence of quencher molecules;
- Ka is the binding constant;
- Q is the quencher concentration.
3.4. Antibacterial Activity of Sumac–Liposome Nanocomplexes
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Fraga-Corral, M.; Otero, P.; Cassani, L.; Echave, J.; Garcia-Oliveira, P.; Carpena, M.; Chamorro, F.; Lourenço-Lopes, C.; Prieto, M.A.; Simal-Gandara, J. Traditional applications of tannin rich extracts supported by scientific data: Chemical composition, bioavailability and bioaccessibility. Foods 2021, 10, 251. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Ito, H. Tannins of constant structure in medicinal and food plants—Hydrolysable tannins and polyphenols related to tannins. Molecules 2011, 16, 2191–2217. [Google Scholar] [CrossRef]
- Sieniawska, E. Activities of Tannins—From In Vitro Studies to Clinical Trials. Nat. Prod. Commun. 2015, 10, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Barreca, D.; Bellocco, E.; Trombetta, D. Proanthocyanidins and hydrolysable tannins: Occurrence, dietary intake and pharmacological effects. Br. J. Pharmacol. 2017, 174, 1244–1262. [Google Scholar] [CrossRef] [PubMed]
- Girard, M.; Bee, G. Invited review: Tannins as a potential alternative to antibiotics to prevent coliform diarrhea in weaned pigs. Animal 2020, 14, 95–107. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, F. Chemical composition and biological activity of staghorn sumac (Rhus typhina). Food Chem. 2017, 15, 431–443. [Google Scholar] [CrossRef]
- Olchowik-Grabarek, E.; Swiecicka, I.; Andreeva-Kovaleskaya, Z.; Solonin, A.; Bonarska-Kujawa, D.; Kleszczynska, H.; Mavlyanov, S.; Zamaraeva, M. Role of structural changes induced in biological membranes by hydrolysable tannins from sumac leaves (Rhus typhina L.) in their antihemolytic and antibacterial effects. J. Membr. Biol. 2014, 247, 533–540. [Google Scholar] [CrossRef]
- Olchowik-Grabarek, E.; Sekowski, S.; Bitiucki, M.; Dobrzynska, I.; Shlyonsky, V.; Ionov, M.; Burzynski, P.; Roszkowska, A.; Swiecicka, I.; Abdulladjanova, N.; et al. Inhibition of interaction between Staphylococcus aureus α-hemolysin and erythrocytes membrane by hydrolysable tannins: Structure-related activity study. Sci. Rep. 2020, 10, 11168. [Google Scholar] [CrossRef]
- Olchowik, E.; Lotkowski, K.; Mavlyanov, S.; Abdullajanova, N.; Ionov, M.; Bryszewska, M.; Zamaraeva, M. Stabilization of erythrocytes against oxidative and hypotonic stress by tannins isolated from sumac leaves (Rhus typhina L.) and grape seeds (Vitis vinifera L.). Cell Mol. Biol Lett. 2012, 17, 333–348. [Google Scholar] [CrossRef]
- Olchowik-Grabarek, E.; Mavlyanov, S.; Abdullajanova, N.; Gieniusz, R.; Zamaraeva, M. Specificity of hydrolysable tannins from Rhus typhina L. to oxidants in cell and cell-free models. Appl. Biochem. Biotechnol. 2017, 181, 495–510. [Google Scholar] [CrossRef]
- Olchowik-Grabarek, E.; Makarova, K.; Mavlyanov, S.; Abdullajanova, N.; Zamaraeva, M. Comparative analysis of BPA and HQ toxic impact on human erythrocytes, protective effect mechanism of tannins (Rhus typhina). Environ. Sci. Pollut. Res. Int. 2018, 25, 1200–1209. [Google Scholar] [CrossRef] [PubMed]
- Sekowski, S.; Ionov, M.; Abdulladjanova, N.; Makhmudov, R.; Mavlyanov, S.; Milowska, K.; Bryszewska, M.; Zamaraeva, M. Interaction of α-synuclein with Rhus typhina tannin—Implication for Parkinson’s disease. Colloids Surf. B Biointerfaces 2017, 155, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Sekowski, S.; Olchowik-Grabarek, E.; Dubis, A.T.; Sharan, L.; Kumar, A.; Abdulladjanova, N.; Markiewicz, P.; Zamaraeva, M. Inhibition of AGEs formation, antioxidative, and cytoprotective activity of Sumac (Rhus typhina L.) tannin under hyperglycemia: Molecular and cellular study. Mol. Cell. Biochem. 2023, 478, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xu, M.L.; Xin, L.; Ma, C.; Yu, G.; Saravanakumar, K.; Myeong-Hyeon, W. Oxidative stress induced apoptosis mediated anticancer activity of Rhus typhina fruits extract in human colon cancer. Med. Chem. Res. 2019, 28, 917–925. [Google Scholar] [CrossRef]
- Arora, A.; Byrem, T.M.; Nair, M.G.; Strasburg, G.M. Modulation of liposomal membrane fluidity by flavonoids and isoflavonoids. Arch. Biochem. Biophys. 2000, 373, 102–109. [Google Scholar] [CrossRef]
- Caturla, N.; Vera-Samper, E.; Villalain, J.; Mateo, C.R.; Micol, V. The relationship between the antioxidant and the antibacterial properties of galloylated catechins and the structure of phospholipids model membranes. Free Radic. Biol. Med. 2003, 34, 648–662. [Google Scholar] [CrossRef]
- Soto-Arriaza, M.A.; Sotomayor, C.P.; Lissi, E.A. Relationship between lipid peroxidation and rigidity in L-α-phosphatidylcholine-DPPC vesicles. J. Colloid Interface Sci. 2008, 323, 70–74. [Google Scholar] [CrossRef]
- Margina, D.; Gradinaru, D.; Manda, G.; Neagoe, I.; Ilie, M. Membranar effects exerted in vitro by polyphenols—Quercetin, epigallocatechin gallate and curcumin—On HUVEC and Jurkat cells, relevant for diabetes mellitus. Food Chem. Toxicol. 2013, 61, 86–93. [Google Scholar] [CrossRef]
- Sekowski, S.; Terebka, M.; Veiko, A.; Lapshina, E.; Sulkowska, U.; Zavodnik, I.B.; Abdulladjanova, N.; Mavlyanov, S.; Roszkowska, A.; Zamaraeva, M. Epigallocatechingallate (EGCG) activity against UV light-induced photo damages in erythrocytes and serum albumin—Theoretical and experimental studies. J. Photochem. Photobiol. A Chem. 2018, 356, 379–388. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, R.-F.; Khalifa, I.; Li, C.-M. Understanding toward the biophysicali of polymeric proanthocyanidins (persimmon condensed tannins) with biomembranes: Relevance for biological effects. J. Agric. Food Chem. 2019, 67, 11044–11052. [Google Scholar] [CrossRef]
- Kajiya, K.; Kumazawa, S.; Nakayama, T. Steric effects on interaction of tea catechins with lipid bilayers. Biosci. Biotechnol. Biochem. 2001, 65, 2638–2643. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, T.; Kajiya, K.; Kumazawa, S. Interaction of plant polyphenols with liposomes. Adv. Planar Lipid Bilayers Liposomes 2006, 4, 107–133. [Google Scholar]
- Veiko, A.G.; Olchowik-Grabarek, E.; Sekowski, S.; Roszkowska, A.; Lapshina, E.A.; Dobrzynska, I.; Zamaraeva, M.; Zavodnik, I.B. Antimicrobial activity of quercetin, naringenin and catechin: Flavonoids inhibit Staphylococcus aureus-induced hemolysis and modify membranes of bacteria and erythrocytes. Molecules 2023, 28, 1252. [Google Scholar] [CrossRef] [PubMed]
- Pignatello, R.; Musumeci, T.; Basile, L.; Carbone, C.; Puglisi, G. Biomembrane models and drug-biomembrane interaction studies: Involvement in drug design and development. J. Pharm. Bioallied Sci. 2011, 3, 4–14. [Google Scholar] [CrossRef] [PubMed]
- Jurczak, P.; Szutkowski, K.; Lach, S.; Jurga, S.; Czaplewska, P.; Szymanska, A.; Zhukov, I. DMPC Phospholipid Bilayer as a Potential Interface for Human Cystatin C Oligomerization: Analysis of Protein-Liposome Interactions Using NMR Spectroscopy. Membranes 2021, 11, 13. [Google Scholar] [CrossRef] [PubMed]
- Sekowski, S.; Veiko, A.; Olchowik-Grabarek, E.; Dubis, A.; Wilczewska, A.; Markiewicz, K.; Zavodnik, I.; Lapshina, E.; Dobrzynska, I.; Abdulladjanova, N.; et al. Hydrolysable tannins change physicochemical parameters of lipid nano-vesicles and reduce DPPH radical—Experimental studies and quantum chemical analysis. Biochim. Biophys. Acta Biomembr. 2022, 1864, 183778. [Google Scholar] [CrossRef]
- Pippa, N.; Deli, E.; Mentzali, E.; Pispas, S.; Demetzos, C. PEO-b-PCL grafted DPPC liposomes: Physicochemical characterization and stability studies of novel bio-inspired advanced Drug Delivery nano Systems (aDDnSs). J. Nanosci. Nanotechnol. 2014, 14, 5676–5681. [Google Scholar] [CrossRef]
- Czajkowska-Szczykowska, D.; Olchowik-Grabarek, E.; Sekowski, S.; Zarkowski, J.; Morzycki, J.W. Concise synthesis of E/F ring spiroethers from tigogenin. Carbaanalogs of steroidal sapogenins and their biological activity. J. Steroid Biochem. Mol. Biol. 2022, 224, 106174. [Google Scholar] [CrossRef]
- Sekowski, S.; Ionov, M.; Kaszuba, M.; Mavlyanov, S.; Bryszewska, M.; Zamaraeva, M. Biophysical studies of interaction between hydrolysable tannins isolated from Oenothera gigas and Geranium sanguineum with human serum albumin. Colloids Surf. B Biointerfaces 2014, 123, 623–628. [Google Scholar] [CrossRef]
- Phan, H.T.T.; Yoda, T.; Chahal, B.; Morita, M.; Takagi, M.; Vestergaard, M.C. Structure-dependent interactions of polyphenols with a biomimetic membrane system. Biochim. Biophys. Acta Biomembr. 2014, 1838, 2670–2677. [Google Scholar] [CrossRef]
- Reis, A.; Soares, S.; Sousa, C.F.; Dias, R.; Gameiro, P.; Soares, S.; Freitas, V. Interaction of polyphenols with model membranes: Putative implications to mouthfeel perception. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183133. [Google Scholar] [CrossRef]
- Veiko, A.G.; Sekowski, S.; Lapshina, E.A.; Wilczewska, A.Z.; Markiewicz, K.H.; Zamaraeva, M.; Zhao, H.C.; Zavodnik, I.B. Flavonoids modulate liposomal membrane structure, regulate mitochondrial membrane permeability and prevent erythrocyte oxidative damage. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183442. [Google Scholar] [CrossRef] [PubMed]
- Danaei, M.; Dehghankhold, M.; Ataei, S.; Hasanzadeh Davarani, F.; Javanmard, R.; Dokhani, A.; Khorasani, S.; Mozafari, M.R. Impact of Particle Size and Polydispersity Index on the Clinical Applications of Lipidic Nanocarrier Systems. Pharmaceutics 2018, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidi, A.; Chountoulesi, M.; Naziris, N.; Sartori, B.; Amenitsch, H.; Mali, G.; Cendak, T.; Plakantonaki, M.; Triantafyllakou, I.; Tselios, T.; et al. The boundary lipid around DMPC-spanning influenza A M2 transmembrane domain channels: Its structure and potential for drug accommodation. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183156. [Google Scholar] [CrossRef] [PubMed]
- Konstantinidi, A.; Naziris, N.; Chountoulesi, M.; Kiriakidi, S.; Sartori, B.; Kolokouris, D.; Amentisch, H.; Mali, G.; Ntountaniotis, D.; Demetzos, C.; et al. Comparative Perturbation Effects Exerted by the Influenza M2 Protein Inhibitors Amantadine and the Spiro[pyrrolidine-2,2-adamantane] Variant AK13 to Membrane Bilayers Studied Using Biophysical Experiments and Molecular Dynamics Simulations. J. Phys. Chem. B 2018, 122, 9877–9895. [Google Scholar] [CrossRef] [PubMed]
- Stampolaki, M.; Malwal, S.R.; Alvarez-Cabrera, N.; Gao, Z.; Moniruzzaman, M.; Babii, S.O.; Naziris, N.; Rey-Cibati, A.; Valladares-Delgado, M.; Turcu, A.L.; et al. Synthesis and Testing of Analogs of the Tuberculosis Drug Candidate SQ109 against Bacteria and Protozoa: Identification of Lead Compounds against Mycobacterium abscessus and Malaria Parasites. ACS Infect. Dis. 2023, 9, 342–364. [Google Scholar] [CrossRef] [PubMed]
- Chountoulesi, M.; Naziris, N.; Mavromoustakos, T.; Demetzos, C. A differential scanning calorimetry (DSC) experimental protocol for evaluating the modified thermotropic behavior thermotropic behavior of liposomes with incorporated guest molecules. In Supramolecules in Drug Discovery and Drug Delivery Book; Series Title: Methods Molecular Biology; Mavromoustakos, T., Tzakos, A.G., Durdagi, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; Volume 2207, pp. 299–312. [Google Scholar]
- Pippa, N.; Chountoulesi, M.; Kyrili, A.; Meristoudi, A.; Pispas, S.; Demetzos, C. Calorimetric study on pH-responsive block copolymer grafted lipid bilayers: Rational design and development of liposomes. J. Liposome Res. 2016, 26, 211–220. [Google Scholar] [CrossRef]
- Naziris, S.; Sekowski, S.; Olchowik-Grabarek, E.; Buczkowski, A.; Balcerzak, Ł.; Chrysostomou, V.; Pispas, S.; Małecka, M.; Bryszewska, M.; Ionov, M. Biophysical interactions of mixed lipid-polymer nanoparticles incorporating curcumin: Potential as antibacterial agent. Biomater. Adv. 2023, 144, 213200. [Google Scholar] [CrossRef]
- Parasassi, T.; Krasnowska, E.K.; Bagatolli, L.; Gratton, E. Laurdan and prodan as polarity-sensitive fluorescence membrane probes. J. Fluoresc. 1998, 8, 365–373. [Google Scholar] [CrossRef]
- Lin, W.; Klein, J. Control of surface forces through hydrated boundary layers. Curr. Opin. Colloid Interface Sci. 2019, 44, 94–106. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer US: New York, NY, USA, 2006; ISBN 1-978-0-387-31278-1. [Google Scholar]
- Nasri, R.; Bidel, L.P.R.; Rugani, N.; Perrier, V.; Ceriere, F.; Dubreucq, E.; Jay-Allemand, C. Inhibition of CpLIP2 Lipase Hydrolytic Activity by Four Flavonols (Galangin, Kaempferol, Quercetin, Myricetin) Compared to Orlistat and Their Binding Mechanisms Studied by Quenching of Fluorescence. Molecules 2019, 24, 2888. [Google Scholar] [CrossRef] [PubMed]
- Maciążek-Jurczyk, M.; Równicka-Zubik, J.; Dyja, R.; Sułkowska, A. Comparative Analysis of KP-HSA Complex by Spectroscopic Methods. Acta Phys. Pol. A 2013, 123, 673–680. [Google Scholar] [CrossRef]
- Liu, P.; Chen, G.; Zhang, J. A Review of Liposomes as a Drug Delivery System: Current Status of Approved Products, Regulatory Environments, and Future Perspectives. Molecules 2022, 27, 1372. [Google Scholar] [CrossRef] [PubMed]
- Olusanya, T.O.B.; Ahmad, R.R.H.; Ibegbu, D.M.; Smith, J.R.; Elkordy, A.A. Liposomal drug delivery system and anticancer drugs. Molecules 2018, 23, 907. [Google Scholar] [CrossRef]
- Enaru, B.; Socaci, S.; Farcas, A.; Socaciu, C.; Danciu, C.; Stanila, A.; Diaconeasa, Z. Novel Delivery Systems of Polyphenols and Their Potential Health Benefits. Pharmaceuticals 2021, 14, 946. [Google Scholar] [CrossRef]
- Tziveleka, L.-A.; Pippa, N.; Ioannou, E.; Demetzos, C.; Roussis, V. Development of Ulvan-Containing Liposomes as Antibacterial Drug Delivery Platforms. J. Funct. Biomater. 2022, 13, 186. [Google Scholar] [CrossRef]
- Gupta, P.D.; Birdi, T.J. Development of botanicals to combat antibiotic resistance. J. Ayurveda Integr. Med. 2017, 8, 266–275. [Google Scholar] [CrossRef]
- Olchowik-Grabarek, E.; Sekowski, S.; Kwiatek, A.; Płaczkiewicz, J.; Abdulladjanova, N.; Shlyonsky, V.; Swiecicka, I.; Zamaraeva, M. The Structural Changes in the Membranes of Staphylococcus aureus Caused by Hydrolysable Tannins Witness Their Antibacterial Activity. Membranes 2022, 12, 1124. [Google Scholar] [CrossRef]
- Risaliti, L.; Kehagia, A.; Daoultzi, E.; Lazari, D.; Bergonzi, M.C.; Vergkizi-Nikolakaki, S.; Hadjipavlou-Litina, D.; Bilia, A.R. Liposomes loaded with Salvia triloba and Rosmarinus officinalis essential oils: In vitro assessment of antioxidant, antiinflammatory and antibacterial activities. J. Drug Deliv. Sci. Technol. 2019, 51, 493–498. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| t (Days) | DMPC | DMPC:RT1 | DMPC:RT5 | DMPC:RT10 | ||||
|---|---|---|---|---|---|---|---|---|
| Dh (nm) | PDI | Dh (nm) | PDI | Dh (nm) | PDI | Dh (nm) | PDI | |
| 0 | 113.5 ± 0.5 | 0.453 ± 0.009 | 113.6 ± 1.0 | 0.441 ± 0.006 | 97.5 ± 2.7 | 0.492 ± 0.005 | 107.2 ± 1.6 | 0.466 ± 0.009 |
| 1 | 131.5 ± 2.3 | 0.568 ± 0.012 | 132.6 ± 1.9 | 0.512 ± 0.006 | 389.9 ± 10.0 | 0.742 ± 0.233 | 215.9 ± 4.1 | 0.703 ± 0.096 |
| 5 | 146.9 ± 2.0 | 0.646 ± 0.012 | 143.0 ± 2.4 | 0.515 ± 0.018 | 203.2 ± 3.1 | 0.623 ± 0.008 | 237.5 ± 2.2 | 0.680 ± 0.120 |
| Sample | RT [µM] | Tonset,m (°C) | Tm (°C) | ΔT1/2,m (°C) | ΔHm (kJ/mol) | Tonset,s (°C) | Ts (°C) | ΔT1/2,s (°C) | ΔHs (kJ/mol) |
|---|---|---|---|---|---|---|---|---|---|
| DMPC | - | 22.94 | 23.37 | 0.77 | 29.47 | 10.84 | 12.32 | 1.62 | 1.33 |
| DMPC:RT1 | 1 | 21.00 | 22.61 | 1.50 | 29.41 | - | - | - | - |
| DMPC:RT5 | 5 | 19.26 | 21.75 | 2.41 | 29.54 | - | - | - | - |
| DMPC:RT10 | 10 | 18.60 | 21.16 | 2.35 | 29.29 | - | - | - | - |
| S. aureus ATCC 700699 | S. aureus 8325-4 | S. epidermitis ATCC 14990 | B. cereus ATCC 13061 | E. coli ATCC 35218 | P. aeruginosa ATCC BAA-1744 | |
|---|---|---|---|---|---|---|
| MIC [μM] | ||||||
| RT | 62.5 | 15.625 | 62.5 | 62.5 | 250 | 250 |
| RT-DMPC nanoparticles | 15.625 | 1.95 | 7.81 | 31.25 | >500 | >500 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sekowski, S.; Naziris, N.; Chountoulesi, M.; Olchowik-Grabarek, E.; Czerkas, K.; Veiko, A.; Abdulladjanova, N.; Demetzos, C.; Zamaraeva, M. Interaction of Rhus typhina Tannin with Lipid Nanoparticles: Implication for the Formulation of a Tannin–Liposome Hybrid Biomaterial with Antibacterial Activity. J. Funct. Biomater. 2023, 14, 296. https://doi.org/10.3390/jfb14060296
Sekowski S, Naziris N, Chountoulesi M, Olchowik-Grabarek E, Czerkas K, Veiko A, Abdulladjanova N, Demetzos C, Zamaraeva M. Interaction of Rhus typhina Tannin with Lipid Nanoparticles: Implication for the Formulation of a Tannin–Liposome Hybrid Biomaterial with Antibacterial Activity. Journal of Functional Biomaterials. 2023; 14(6):296. https://doi.org/10.3390/jfb14060296
Chicago/Turabian StyleSekowski, Szymon, Nikolaos Naziris, Maria Chountoulesi, Ewa Olchowik-Grabarek, Krzysztof Czerkas, Artem Veiko, Nodira Abdulladjanova, Costas Demetzos, and Maria Zamaraeva. 2023. "Interaction of Rhus typhina Tannin with Lipid Nanoparticles: Implication for the Formulation of a Tannin–Liposome Hybrid Biomaterial with Antibacterial Activity" Journal of Functional Biomaterials 14, no. 6: 296. https://doi.org/10.3390/jfb14060296