
Biological Activities of Glass Ionomer Cement Supplemented with Fortilin on Human Dental Pulp Stem Cells

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmid Construction and Expression of Recombinant Fortilins
2.2. Cultures of Human Dental Pulp Stem Cells (hDPSCs)
2.3. Cell Viability Testing
2.4. Histamine Releasing Assay
2.5. Specimens Preparation
2.6. Scanning Electron Microscopy (SEM)
2.7. Cytotoxicity of Modified GICs on hDPSCs
2.8. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
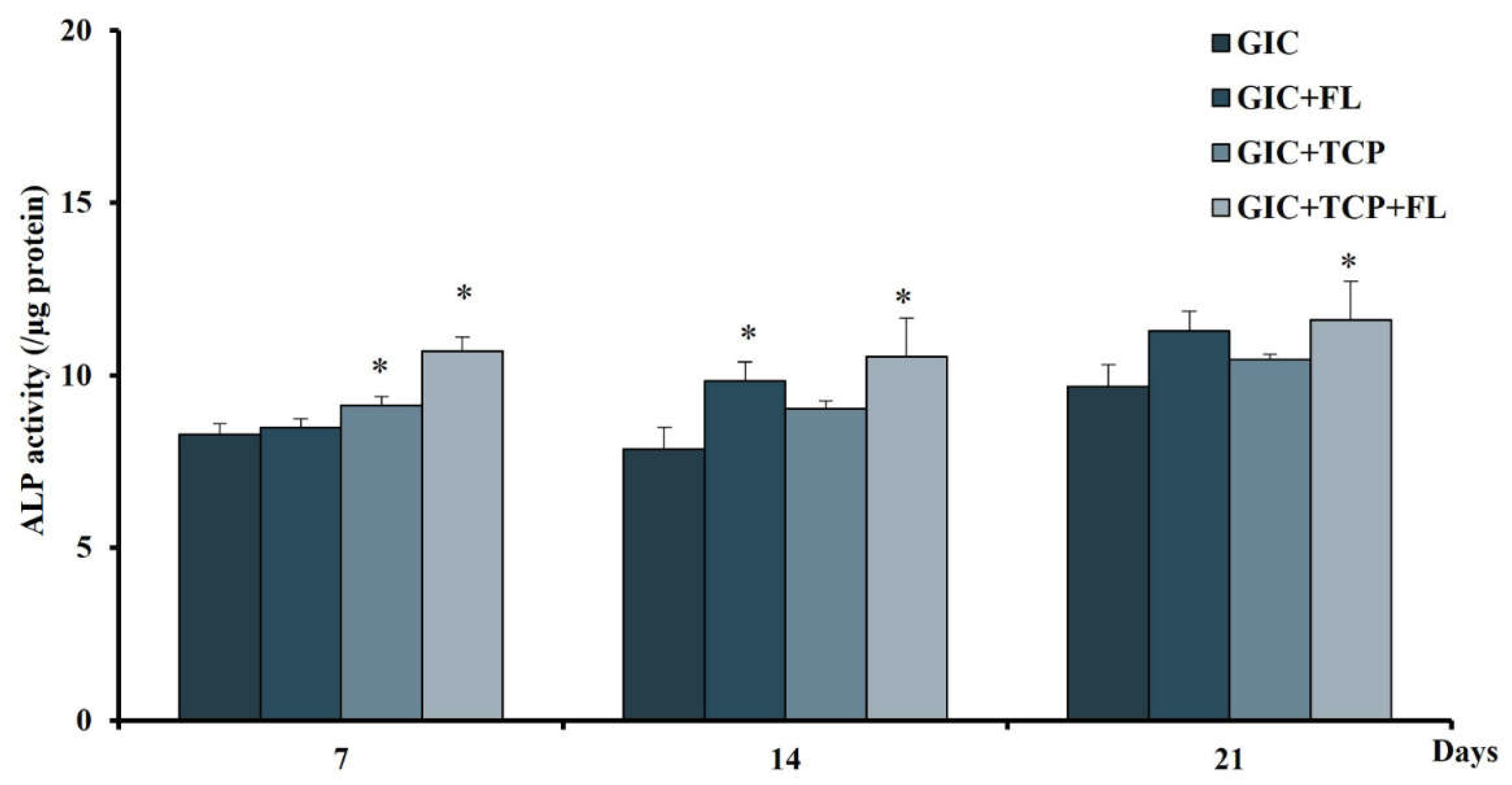
2.9. Alkaline Phosphatase (ALP) Assay
2.10. Alizarin Red S (ARS) Assay
2.11. Statistical Analysis
3. Results
3.1. Characterization of Mesenchymal Stem Cells
3.2. Effect of Full-Length and Fragments of Fortilin on hDPSCs
3.3. The Protective Effect of Fortilin on HEMA-Treated hDPSCs
3.4. Histamine Release
3.5. Morphology of GICs
3.6. Cytotoxicity of Modified GICs on hDPSCs
3.7. Expression of Odontogenic Differentiation Markers in hDPSCs after Cultured with Modified GICs
3.8. Effect of Modified GICs on Differentiation of hDPSCs
3.9. Effect of Modified GICs on Mineralization of hDPSCs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sidhu, S.K.; Nicholson, J.W. A review of glass-ionomer cements for clinical dentistry. J. Funct. Biomater. 2016, 7, 16. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, F.; Nair, L.; Laurencin, C. Inductive materials for regenerative engineering. J. Dent. Res. 2021, 100, 1011–1019. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Zhao, M.; Lash, B.; Martino, M.M.; Julier, Z. Growth factor engineering strategies for regenerative medicine applications. Front. Bioeng. Biotechnol. 2020, 7, 469. [Google Scholar] [CrossRef] [PubMed]
- Whitehouse, L.; Thomson, N.; Do, T.; Feichtinger, G. Bioactive molecules for regenerative pulp capping. Eur. Cell Mater. 2021, 42, 415–437. [Google Scholar] [CrossRef] [PubMed]
- Bommer, U.-A.; Telerman, A. Dysregulation of TCTP in biological processes and diseases. Cells 2020, 9, 1632. [Google Scholar] [CrossRef]
- Seo, E.-J.; Fischer, N.; Efferth, T. Role of TCTP for cellular differentiation and cancer therapy. In TCTP/tpt1-Remodeling Signaling from Stem Cell to Disease; Springer: Berlin/Heidelberg, Germany, 2017; pp. 263–281. [Google Scholar]
- Bommer, U.-A.; Thiele, B.-J. The translationally controlled tumour protein (TCTP). Int. J. Biochem. Cell Biol. 2004, 36, 379–385. [Google Scholar] [CrossRef]
- Bommer, U.-A. The translational controlled tumour protein TCTP: Biological functions and regulation. In TCTP/tpt1-Remodeling Signaling from Stem Cell to Disease; Springer: Berlin/Heidelberg, Germany, 2017; pp. 69–126. [Google Scholar]
- Bommer, U.-A.; Kawakami, T. Role of TCTP in Cell Biological and Disease Processes. Cells 2021, 10, 2290. [Google Scholar] [CrossRef]
- Nagano-Ito, M.; Ichikawa, S. Biological effects of Mammalian translationally controlled tumor protein (TCTP) on cell death, proliferation, and tumorigenesis. Biochem. Res. Int. 2012, 2012, 204960. [Google Scholar] [CrossRef]
- Gnanasekar, M.; Dakshinamoorthy, G.; Ramaswamy, K. Translationally controlled tumor protein is a novel heat shock protein with chaperone-like activity. Biochem. Biophys. Res. Commun. 2009, 386, 333–337. [Google Scholar] [CrossRef]
- Wanachottrakul, N.; Chotigeat, W.; Kedjarune-Leggat, U. Translationally controlled tumor protein against apoptosis from 2-hydroxy-ethyl methacrylate in human dental pulp cells. J. Mater. Sci. Mater. Med. 2011, 22, 1479–1487. [Google Scholar] [CrossRef]
- Wanachottrakul, N.; Chotigeat, W.; Kedjarune-Leggat, U. Effect of novel chitosan-fluoroaluminosilicate resin modified glass ionomer cement supplemented with translationally controlled tumor protein on pulp cells. J. Mater. Sci. Mater. Med. 2014, 25, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Kedjarune-Leggat, U.; Saetan, U.; Khongsaengkaeo, A.; Suwannarat, S.; Deachamag, P.; Wonglapsuwan, M.; Pornprasit, R.; Thongkamwitoon, W.; Phumklai, P.; Chaichanan, J. Biological activities of a recombinant fortilin from Fenneropenaeus merguiensis. PLoS ONE 2020, 15, e0239672. [Google Scholar] [CrossRef] [PubMed]
- Pinkaew, D.; Fujise, K. Fortilin: A potential target for the prevention and treatment of human diseases. Adv. Clin. Chem. 2017, 82, 265–300. [Google Scholar] [PubMed]
- Sangsuwan, J.; Wanichpakorn, S.; Kedjarune-Leggat, U. Translationally controlled tumor protein supplemented chitosan modified glass ionomer cement promotes osteoblast proliferation and function. Mater. Sci. Eng. 2015, 54, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Galler, K.M.; Weber, M.; Korkmaz, Y.; Widbiller, M.; Feuerer, M. Inflammatory response mechanisms of the dentine–pulp complex and the periapical tissues. Inter. J. Mol. Sci. 2021, 22, 1480. [Google Scholar] [CrossRef]
- Ching, H.S.; Luddin, N.; Kannan, T.P.; Ab Rahman, I.; Abdul Ghani, N.R. Modification of glass ionomer cements on their physical-mechanical and antimicrobial properties. J. Esthet. Restor. Dent. 2018, 30, 557–571. [Google Scholar] [CrossRef]
- Najeeb, S.; Khurshid, Z.; Zafar, M.S.; Khan, A.S.; Zohaib, S.; Martí, J.M.N.; Sauro, S.; Matinlinna, J.P.; Rehman, I.U. Modifications in glass ionomer cements: Nano-sized fillers and bioactive nanoceramics. Inter. J. Mol. Sci. 2016, 17, 1134. [Google Scholar] [CrossRef]
- Weng, Y.; Guo, X.; Gregory, R.; Xie, D. A novel antibacterial dental glass-ionomer cement. Eur. J. Oral Sci. 2010, 118, 531–534. [Google Scholar] [CrossRef]
- Fischer, N.G.; Münchow, E.A.; Tamerler, C.; Bottino, M.C.; Aparicio, C. Harnessing biomolecules for bioinspired dental biomaterials. J. Mater. Chem. 2020, 8, 8713–8747. [Google Scholar] [CrossRef]
- Graidist, P.; Yazawa, M.; Tonganunt, M.; Nakatomi, A.; Lin, C.C.-J.; Chang, J.-Y.; Phongdara, A.; Fujise, K. Fortilin binds Ca2+ and blocks Ca2+-dependent apoptosis in vivo. Biochem. J. 2007, 408, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Thébault, S.; Agez, M.; Chi, X.; Stojko, J.; Cura, V.; Telerman, S.B.; Maillet, L.; Gautier, F.; Billas-Massobrio, I.; Birck, C. TCTP contains a BH3-like domain, which instead of inhibiting, activates Bcl-xL. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gallorini, M.; Cataldi, A.; di Giacomo, V. HEMA-induced cytotoxicity: Oxidative stress, genotoxicity and apoptosis. Int. Endod. J. 2014, 47, 813–818. [Google Scholar] [CrossRef] [PubMed]
- Schweikl, H.; Petzel, C.; Bolay, C.; Hiller, K.A.; Buchalla, W.; Krifka, S. 2-Hydroxyethyl methacrylate-induced apoptosis through the ATM- and p53-dependent intrinsic mitochondrial pathway. Biomaterials 2014, 35, 2890–2904. [Google Scholar] [CrossRef] [PubMed]
- Limapornvanich, A.; Jitpukdeebodintra, S.; Hengtrakool, C.; Kedjarune-Leggat, U. Bovine serum albumin release from novel chitosan-fluoro-aluminosilicate glass ionomer cement: Stability and cytotoxicity studies. J. Dent. 2009, 37, 686–690. [Google Scholar] [CrossRef]
- Yang, X.; Li, L.; Xiao, L.; Zhang, D. Recycle the dental fairy’s package: Overview of dental pulp stem cells. Stem Cell Res. Ther. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Sabbagh, J.; Ghassibe-Sabbagh, M.; Fayyad-Kazan, M.; Al-Nemer, F.; Fahed, J.C.; Berberi, A.; Badran, B. Differences in osteogenic and odontogenic differentiation potential of DPSCs and SHED. J. Dent. 2020, 101, 103413. [Google Scholar] [CrossRef]
- Koh, B.; Sulaiman, N.; Ismadi, S.N.S.W.; Ramli, R.; Yunus, S.S.M.; Idrus, R.B.H.; Ariffin, S.H.Z.; Wahab, R.M.A.; Yazid, M.D. Mesenchymal stem cells: A comprehensive methods for odontoblastic induction. Biol. Proced. Online 2021, 23, 1–13. [Google Scholar] [CrossRef]
- Liu, M.; Li, W.; Xia, X.; Wang, F.; MacDougall, M.; Chen, S. Dentine sialophosphoprotein signal in dentineogenesis and dentine regeneration. Eur. Cell Mater. 2021, 42, 43. [Google Scholar] [CrossRef]
- Jeong, J.; Kim, J.H.; Shim, J.H.; Hwang, N.S.; Heo, C.Y. Bioactive calcium phosphate materials and applications in bone regeneration. Biomater. Res. 2019, 23, 1–11. [Google Scholar] [CrossRef]
- Khan, A.S.; Syed, M.R. A review of bioceramics-based dental restorative materials. Dent. Mater. J. 2019, 38, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Viale-Bouroncle, S.; Bey, B.; Reichert, T.E.; Schmalz, G.; Morsczeck, C. β-tricalcium-phosphate stimulates the differentiation of dental follicle cells. J. Mater. Sci. Mater. Med. 2011, 22, 1719–1724. [Google Scholar] [CrossRef] [PubMed]
- Eslaminejad, M.B.; Bordbar, S.; Nazarian, H. Odontogenic differentiation of dental pulp-derived stem cells on tricalcium phosphate scaffolds. J. Dent. Sci. 2013, 8, 306–313. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Powder Components |
|---|---|
| GIC | 100% calcium fluoroaluminosilicate glass and 5% copolymer acid |
| GIC + TCP | 99.95% calcium fluoroaluminosilicate glass, 5% copolymer acid, and 0.05% tricalcium phosphate |
| GIC + FL | 100% calcium fluoroaluminosilicate glass, 5% copolymer acid, and 1 µg fortilin |
| GIC + TCP + FL | 99.95% calcium fluoroaluminosilicate glass, 5% copolymer acid, 0.05% tricalcium phosphate, and 1 µg fortilin |
| Gene | Primer (5′–3′) | GenBank Accession No. |
|---|---|---|
| ALP | F: CCACAAGCCCGTGACAGA | NM_001127501 |
| R: GCGGCAGACTTTGGTTTC | ||
| OPN | F: ACACATATTGATGGCCGAAGGTGA | NM_00582.2 |
| R: TGTGAGGTGATGTCCTCGTCTGT | ||
| DMP-1 | F: GCAGAGTGATGACCCAGAG | NM_004407.3 |
| R: GCTCGCTTCTGTCATCTTCC | ||
| DSPP | F: GGGATGTTGGCGATGCA | NM_014208.3 |
| R: CCAGCTACTTGAGGTCCATCTTC | ||
| GAPDH | F: GCACCGTCAAGGCTGAGAAC | NM_001289745.1 |
| R: ATGGTGGTGAAGACGCCAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangsuwan, P.; Tannukit, S.; Chotigeat, W.; Kedjarune-Leggat, U. Biological Activities of Glass Ionomer Cement Supplemented with Fortilin on Human Dental Pulp Stem Cells. J. Funct. Biomater. 2022, 13, 132. https://doi.org/10.3390/jfb13030132
Sangsuwan P, Tannukit S, Chotigeat W, Kedjarune-Leggat U. Biological Activities of Glass Ionomer Cement Supplemented with Fortilin on Human Dental Pulp Stem Cells. Journal of Functional Biomaterials. 2022; 13(3):132. https://doi.org/10.3390/jfb13030132
Chicago/Turabian StyleSangsuwan, Prawichaya, Sissada Tannukit, Wilaiwan Chotigeat, and Ureporn Kedjarune-Leggat. 2022. "Biological Activities of Glass Ionomer Cement Supplemented with Fortilin on Human Dental Pulp Stem Cells" Journal of Functional Biomaterials 13, no. 3: 132. https://doi.org/10.3390/jfb13030132