Alpha ERS-ERD Pattern during Divergent and Convergent Thinking Depends on Individual Differences on Metacontrol


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Definition of Metacontrol
1.2. Association between Metacontrol and Creative Thinking
1.3. Aims of the Study
2. Methods
2.1. Participants
2.2. Procedure
2.3. Task and Experimental Materials
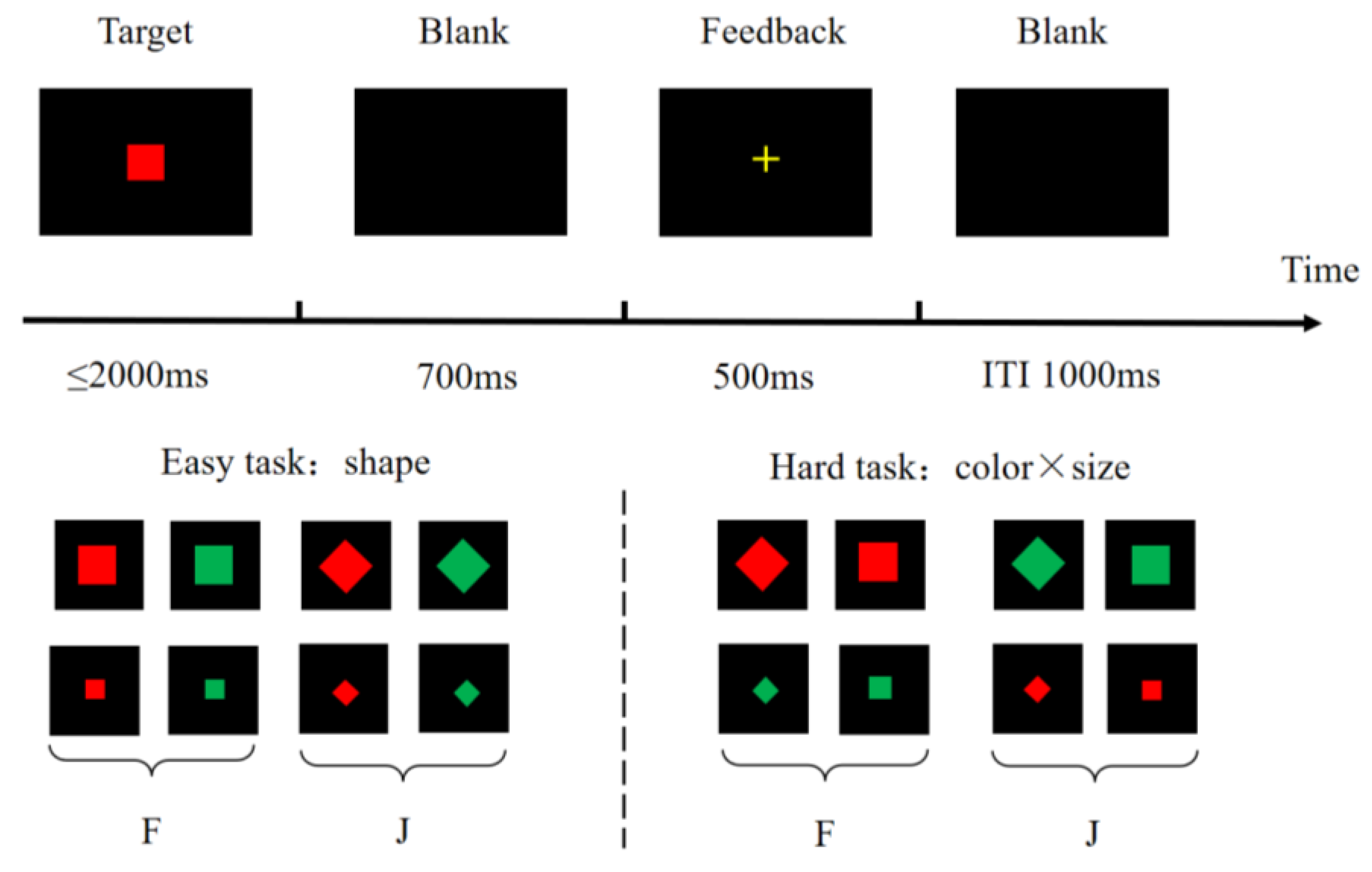
2.3.1. Metacontrol Task
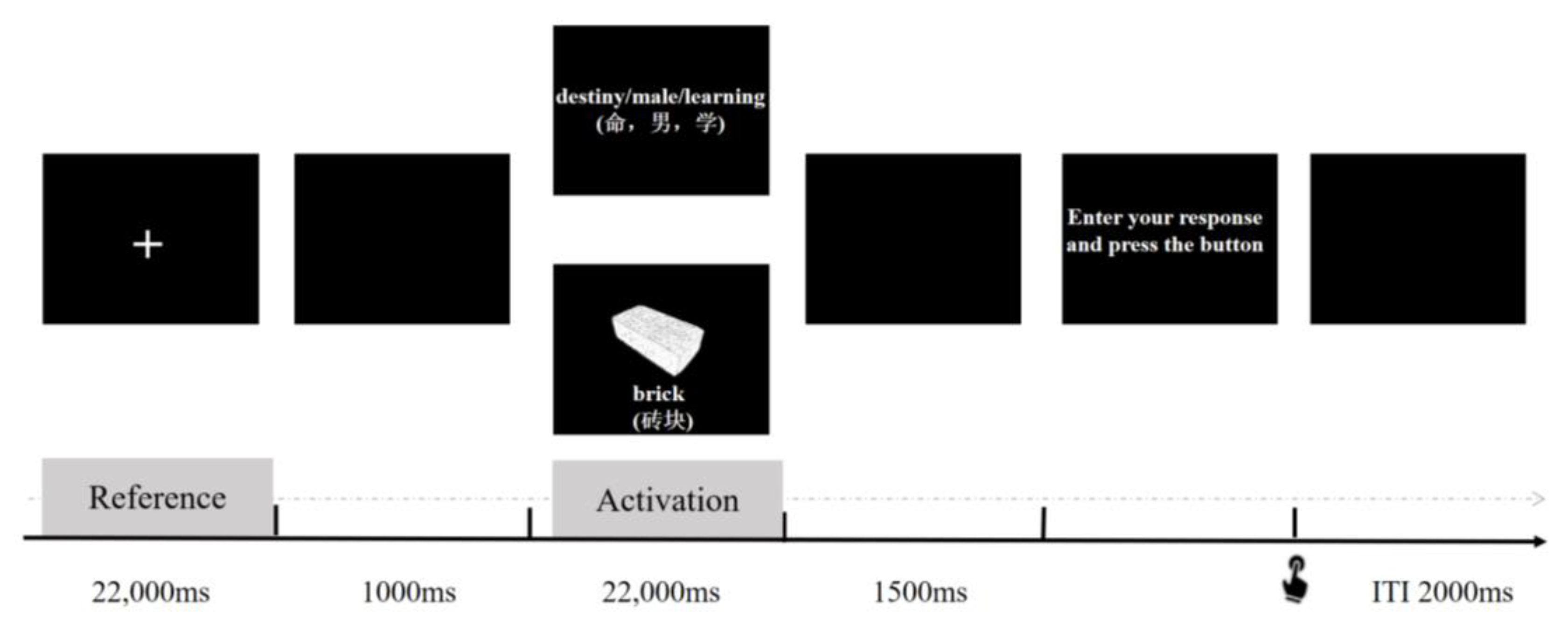
2.3.2. Creative Task
- (1)
- The Alternative Uses Task (AUT)
- (2)
- The Remote Associates Test (RAT)
2.4. EEG Recording and Pre-Processing
2.5. EEG/ERP Data Analysis
2.5.1. ERP for the Metacontrol Task
2.5.2. EEG Alpha TRP for AUT and RAT
2.6. Statistical Analysis
3. Results
3.1. Metacontrol Task Results
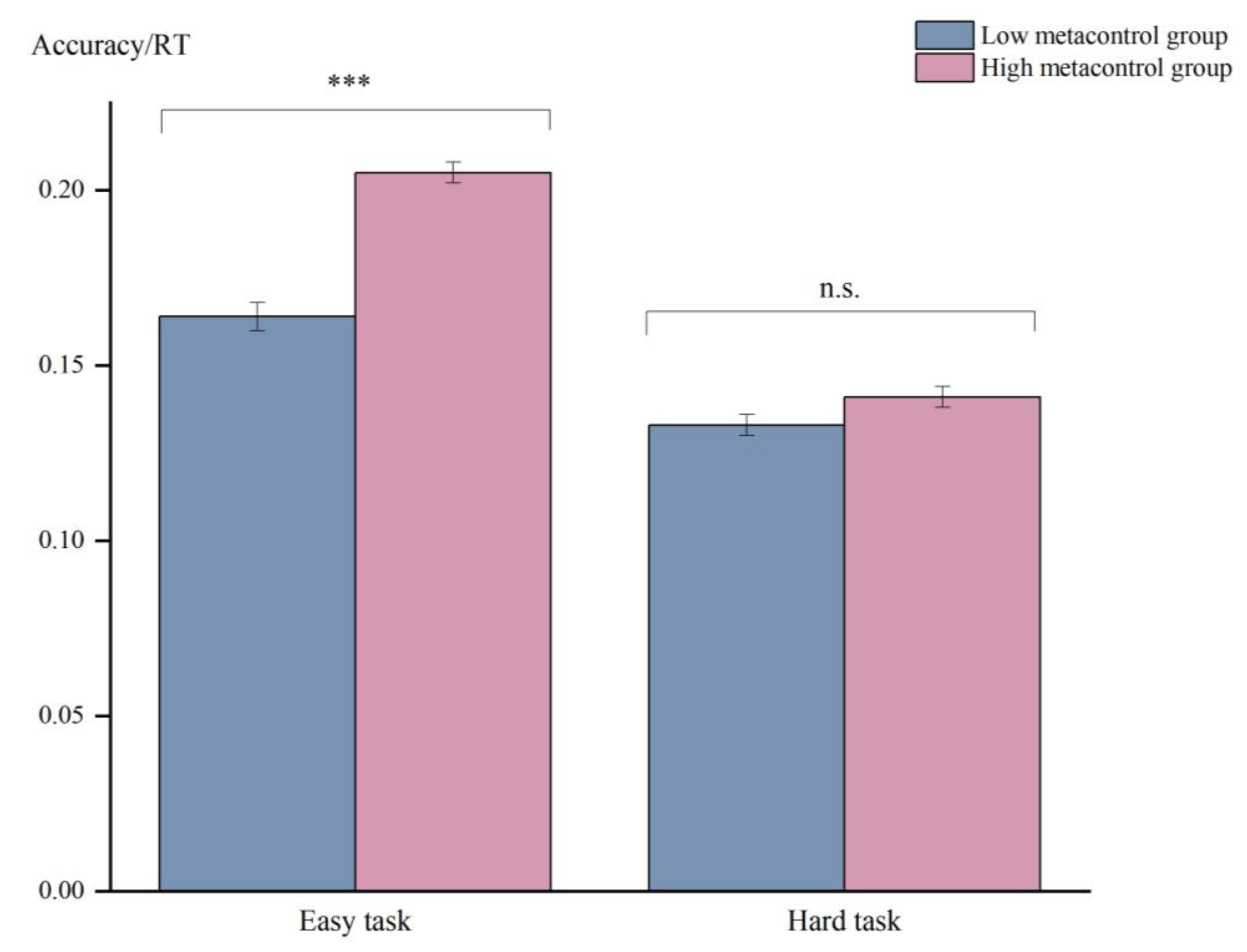
3.1.1. Behavioral Results
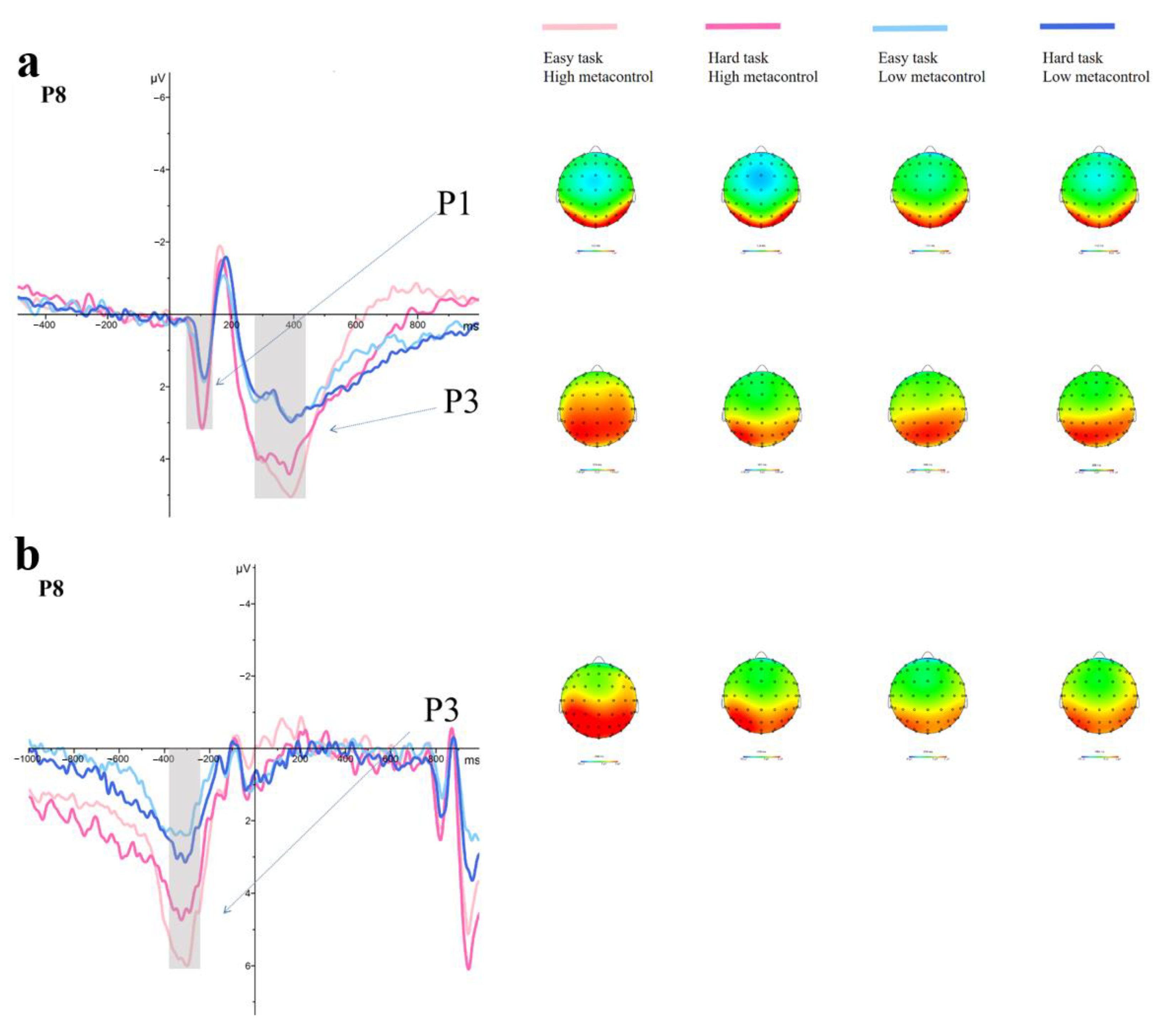
3.1.2. ERP Results
3.2. AUT and RAT Results
3.2.1. Behavioral Results
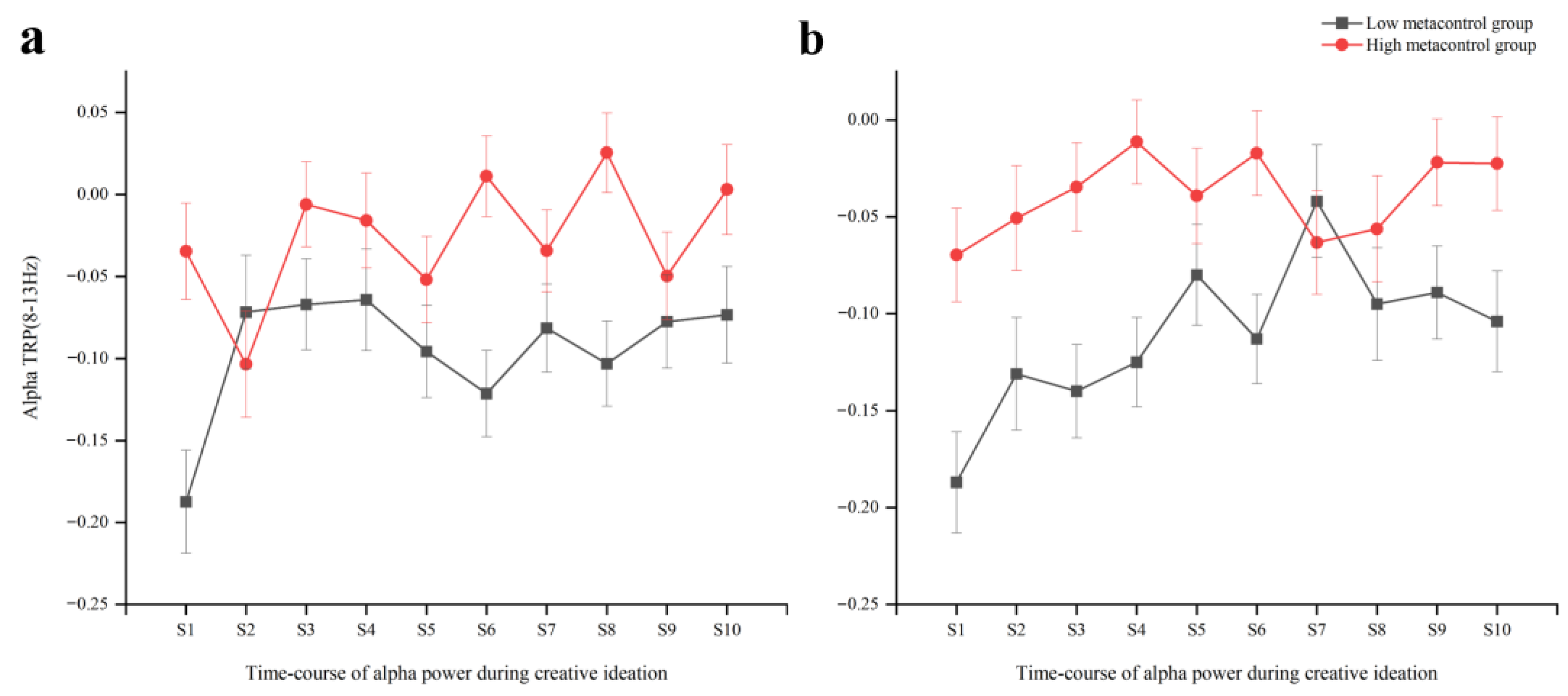
3.2.2. EEG Alpha TRP Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agnoli, Sergio, Marco Zanon, Serena Mastria, Alessio Avenanti, and Giovanni Emanuele Corazza. 2020. Predicting response originality through brain activity: An analysis of changes in EEG alpha power during the generation of alternative ideas. Neuroimage 207: 116385. [Google Scholar] [CrossRef] [PubMed]
- Baas, Matthijs, Carsten K. W. De Dreu, and Bernard A. Nijstad. 2008. A meta-analysis of 25 years of mood-creativity research: Hedonic tone, activation, or regulatory focus? Psychological Bulletin 134: 779–806. [Google Scholar] [CrossRef] [PubMed]
- Benedek, Mathias, Rainer J. Schickel, Emanuel Jauk, Andreas Fink, and Aljoscha C. Neubauer. 2014. Alpha power increases in right parietal cortex reflects focused internal attention. Neuropsychologia 56: 393–400. [Google Scholar] [CrossRef] [PubMed]
- Benedek, Mathias, Sabine Bergner, Tanja Könen, Andreas Fink, and Aljoscha C. Neubauer. 2011. EEG alpha synchronization is related to top-down processing in convergent and divergent thinking. Neuropsychologia 49: 3505–11. [Google Scholar] [CrossRef]
- Bocanegra, Bruno R., and Bernhard Hommel. 2014. When cognitive control is not adaptive. Psychological Science 25: 1249–55. [Google Scholar] [CrossRef]
- Campbell, Donald T. 1960. Blind variation and selective retention in creative thought as in other knowledge processes. Psychological Review 67: 380–400. [Google Scholar] [CrossRef]
- Cheng, Lifang, Weiping Hu, Xiaojuan Jia, and Mark A. Runco. 2016. The different role of cognitive inhibition in early versus late creative problem finding. Psychology of Aesthetics Creativity and the Arts 10: 32–41. [Google Scholar] [CrossRef]
- Chermahini, Soghra Akbari, and Bernhard Hommel. 2010. The (b)link between creativity and dopamine: Spontaneous eye blink rates predict and dissociate divergent and convergent thinking. Cognition 115: 458–65. [Google Scholar] [CrossRef]
- Chermahini, Soghra Akbari, and Bernhard Hommel. 2012. More creative through positive mood? Not everyone! Frontiers in Human Neuroscience 6: 319. [Google Scholar] [CrossRef]
- Colzato, Lorenza S., Ayca Szapora, and Bernhard Hommel. 2012. Meditate to create: The impact of focused-attention and open-monitoring training on convergent and divergent thinking. Frontiers in Psychology 3: 116. [Google Scholar] [CrossRef]
- Colzato, Lorenza S., Bernhard Hommel, Wenxin Zhang, Veit Roessner, and Christian Beste. 2022. The metacontrol hypothesis as diagnostic framework of OCD and ADHD: A dimensional approach based on shared neurobiological vulnerability. Neuroscience and Biobehavioral Reviews 137: 11. [Google Scholar] [CrossRef] [PubMed]
- Colzato, Lorenza S., Roberta Sellaro, Iliana Samara, Matthijs Baas, and Bernhard Hommel. 2015. Meditation-induced states predict attentional control over time. Consciousness and Cognition 37: 57–62. [Google Scholar] [CrossRef] [PubMed]
- Dreisbach, Gesine, and Thomas Goschke. 2004. How positive affect modulates cognitive control: Reduced perseveration at the cost of increased distractibility. Journal of Experimental Psychology: Learning Memory and Cognition 30: 343–53. [Google Scholar] [CrossRef] [PubMed]
- Eppinger, Ben, Thomas Goschke, and Sebastian Musslick. 2021. Meta-control: From psychology to computational neuroscience. Cognitive Affective & Behavioral Neuroscience 21: 447–52. [Google Scholar] [CrossRef]
- Fink, Andreas, Roland H. Grabner, Mathias Benedek, and Aljoscha C. Neubauer. 2006. Divergent thinking training is related to frontal electroencephalogram alpha synchronization. European Journal of Neuroscience 23: 2241–46. [Google Scholar] [CrossRef] [PubMed]
- Finke, Ronald A., Thomas B. Ward, and Steven M. Smith. 1992. Creative Cognition: Theory, Research, and Applications. Cambridge: MIT Press. [Google Scholar]
- Fischer, Rico, and Bernhard Hommel. 2012. Deep thinking increases task-set shielding and reduces shifting flexibility in dual-task performance. Cognition 123: 303–7. [Google Scholar] [CrossRef] [PubMed]
- Fu, Lei, Jia Zhao, Jiangzhou Sun, Yuchi Yan, Mujie Ma, Qunlin Chen, Jiang Qiu, and Wenjing Yang. 2022. Everyday creativity is associated with increased frontal electroencephalography alpha activity during creative ideation. Neuroscience 503: 107–17. [Google Scholar] [CrossRef]
- Goschke, Thomas, and Annette Bolte. 2014. Emotional modulation of control dilemmas: The role of positive affect, reward, and dopamine in cognitive stability and flexibility. Neuropsychologia 62: 403–23. [Google Scholar] [CrossRef]
- Guilford, Joy Paul. 1967. The Nature of Human Intelligence. New York: McGraw-Hill Press. [Google Scholar]
- Herrmann, Christoph S., and Robert T. Knight. 2001. Mechanisms of human attention: Event-related potentials and oscillations. Neuroscience and Biobehavioral Reviews 25: 465–76. [Google Scholar] [CrossRef]
- Hommel, Bernhard. 2015. Between persistence and flexibility: The Yin and Yang of action control. Advances in Motivation Science 2: 33–67. [Google Scholar] [CrossRef]
- Hommel, Bernhard. 2019. Theory of Event Coding (TEC) V2.0: Representing and controlling perception and action. Attention Perception & Psychophysics 81: 2139–54. [Google Scholar] [CrossRef]
- Hommel, Bernhard, and Lorenza S. Colzato. 2017a. Meditation and metacontrol. Journal of Cognitive Enhancement 1: 115–21. [Google Scholar] [CrossRef]
- Hommel, Bernhard, and Lorenza S. Colzato. 2017b. The social transmission of metacontrol policies: Mechanisms underlying the interpersonal transfer of persistence and flexibility. Neuroscience and Biobehavioral Reviews 81: 43–58. [Google Scholar] [CrossRef] [PubMed]
- Hommel, Bernhard, and Reinout W. Wiers. 2017. Towards a unitary approach to human action control. Trends in Cognitive Sciences 21: 940–49. [Google Scholar] [CrossRef] [PubMed]
- Kang, Moon Sun, and Yu-Chin Chiu. 2021. Proactive and reactive metacontrol in task switching. Memory & Cognition 49: 1617–32. [Google Scholar] [CrossRef]
- Klimesch, Wolfgang, Paul Sauseng, and Simon Hanslmayr. 2007. EEG alpha oscillations: The inhibition-timing hypothesis. Brain Research Reviews 53: 63–88. [Google Scholar] [CrossRef] [PubMed]
- Kok, Albert. 2001. On the utility of P3 amplitude as a measure of processing capacity. Psychophysiology 38: 557–77. [Google Scholar] [CrossRef]
- Luck, Steven J., Geoffrey F. Woodman, and Edward K. Vogel. 2000. Event-related potential studies of attention. Trends in Cognitive Sciences 4: 432–40. [Google Scholar] [CrossRef]
- Ma, Ke, and Bernhard Hommel. 2020. Metacontrol and body ownership: Divergent thinking increases the virtual hand illusion. Psychological Research-Psychologische Forschung 84: 120–27. [Google Scholar] [CrossRef]
- Ma, Ke, Na Deng, and Bernhard Hommel. 2021. Meditation-induced cognitive-control states regulate working memory task performance. Quarterly Journal of Experimental Psychology 74: 1465–76. [Google Scholar] [CrossRef]
- Mekern, Vera N., Bernhard Hommel, and Zsuzsika Sjoerds. 2019a. Computational models of creativity: A review of single-process and multi-process recent approaches to demystify creative cognition. Current Opinion in Behavioral Sciences 27: 47–54. [Google Scholar] [CrossRef]
- Mekern, Vera N., Zsuzsika Sjoerds, and Bernhard Hommel. 2019b. How metacontrol biases and adaptivity impact performance in cognitive search tasks. Cognition 182: 251–59. [Google Scholar] [CrossRef]
- Pfurtscheller, Gert, and Colin Andrew. 1999. Event-related changes of band power and coherence: Methodology and interpretation. Journal of Clinical Neurophysiology 16: 512–19. [Google Scholar] [CrossRef] [PubMed]
- Polich, John. 2007. Updating P300: An integrative theory of P3a and P3b. Clinical Neurophysiology 118: 2128–48. [Google Scholar] [CrossRef] [PubMed]
- Rominger, Christian, Ilona Papousek, Corinna M. Perchtold, Bernhard Weber, Elisabeth M. Weiss, and Andreas Fink. 2018. The creative brain in the figural domain: Distinct patterns of EEG alpha power during idea generation and idea elaboration. Neuropsychologia 118: 13–19. [Google Scholar] [CrossRef]
- Rominger, Christian, Ilona Papousek, Corinna M. Perchtold, Mathias Benedek, Elisabeth M. Weiss, Andreas Schwerdtfeger, and Andreas Fink. 2019. Creativity is associated with a characteristic U-shaped function of alpha power changes accompanied by an early increase in functional coupling. Cognitive Affective & Behavioral Neuroscience 19: 1012–21. [Google Scholar] [CrossRef]
- Schneider, Daniel, Christian Beste, and Edmund Wascher. 2012. On the time course of bottom-up and top-down processes in selective visual attention: An EEG study. Psychophysiology 49: 1660–71. [Google Scholar] [CrossRef]
- Schwab, Daniela, Mathias Benedek, Ilona Papousek, Elisabeth M. Weiss, and Andreas Fink. 2014. The time-course of EEG alpha power changes in creative ideation. Frontiers in Human Neuroscience 8: 310. [Google Scholar] [CrossRef]
- Shen, Wangbing, Yuan Yuan, Chang Liu, Baoshu Yi, and Kai Dou. 2016. The Development and Validity of a Chinese Version of the Compound Remote Associates Test. The American Journal of Psychology 129: 245–58. [Google Scholar] [CrossRef]
- Simonton, Dean Keith. 2010. Creative thought as blind-variation and selective-retention: Combinatorial models of exceptional creativity Reply. Physics of Life Reviews 7: 156–79. [Google Scholar] [CrossRef]
- Sternberg, Robert J., and Todd I. Lubart. 1996. Invest in creativity. American Psychologist 51: 677–88. [Google Scholar] [CrossRef]
- Stock, Ann-Kathrin, Laura Steenbergen, Lorenza Colzato, and Christian Beste. 2016. The system neurophysiological basis of non-adaptive cognitive control: Inhibition of implicit learning mediated by right prefrontal regions. Human Brain Mapping 37: 4511–22. [Google Scholar] [CrossRef] [PubMed]
- Twomey, Deirdre M., Peter R. Murphy, Simon P. Kelly, and Redmond G. O’Connell. 2015. The classic P300 encodes a build-to-threshold decision variable. European Journal of Neuroscience 42: 1636–43. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, Sarah, Lorenza S. Colzato, Nicole Wolff, and Christian Beste. 2021. Short-term focused attention meditation restricts the retrieval of stimulus-response bindings to relevant information. Mindfulness 12: 1272–81. [Google Scholar] [CrossRef]
- Verleger, Rolf, Lin Marlena Hamann, Dariusz Asanowicz, and Kamila Śmigasiewicz. 2015. Testing the S-R link hypothesis of P3b: The oddball effect on S1-evoked P3 gets reduced by increased task relevance of S2. Biological Psychology 108: 25–35. [Google Scholar] [CrossRef]
- Verleger, Rolf, Piotr Jaśkowski, and Edmund Wascher. 2005. Evidence for an integrative role of P3b in linking reaction to perception. Journal of Psychophysiology 19: 165–81. [Google Scholar] [CrossRef]
- Zhang, Weitao, Zsuzsika Sjoerds, and Bernhard Hommel. 2020. Metacontrol of human creativity: The neurocognitive mechanisms of convergent and divergent thinking. Neuroimage 210: 116572. [Google Scholar] [CrossRef]
- Zink, Nicolas, Ann-Kathrin Stock, Amirali Vahid, and Christian Beste. 2018. On the neurophysiological mechanisms underlying the adaptability to varying cognitive control demands. Frontiers in Human Neuroscience 12: 411. [Google Scholar] [CrossRef]
- Zink, Nicolas, Wiebke Bensmann, Larissa Arning, Christian Beste, and Ann-Kathrin Stock. 2019. Apolipoprotein ε4 is associated with better cognitive control allocation in healthy young adults. Neuroimage 185: 274–85. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Lin, Y.; Ye, C.; Yang, J.; He, W. Alpha ERS-ERD Pattern during Divergent and Convergent Thinking Depends on Individual Differences on Metacontrol. J. Intell. 2023, 11, 74. https://doi.org/10.3390/jintelligence11040074
Liu C, Lin Y, Ye C, Yang J, He W. Alpha ERS-ERD Pattern during Divergent and Convergent Thinking Depends on Individual Differences on Metacontrol. Journal of Intelligence. 2023; 11(4):74. https://doi.org/10.3390/jintelligence11040074
Chicago/Turabian StyleLiu, Chunlei, Yuhong Lin, Chaoqun Ye, Jiaqin Yang, and Wenguang He. 2023. "Alpha ERS-ERD Pattern during Divergent and Convergent Thinking Depends on Individual Differences on Metacontrol" Journal of Intelligence 11, no. 4: 74. https://doi.org/10.3390/jintelligence11040074