
Endophytic Fungi of Emersed Halophytes in River Deltas and Tidal Flats of the Korean Ramsar Wetlands



Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Plant Sampling
2.3. Isolation of Endophytic Fungi
2.4. Extraction of Genomic DNA and Target Gene Amplification
2.5. Identification and Phylogeny
2.6. Biodiversity Assessment
3. Results and Discussion
3.1. Taxonomy and Phylogeny
3.2. Endophytic Distribution in Each Emersed Halophyte Species
3.3. Morphological and Alpha Diversity
3.4. Endophytic Distribution and Diversity within Salt Marshes in Estuaries and Tidal Flats
3.5. Wide Distribution of Certain Fungal Genera
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pritchard, D.W. What Is an Estuary: Physical Viewpoint. In Estuaries; Lauf, G.H., Ed.; American Association for the Advancement of Science Publishing 83: Washington, DC, USA, 1967; pp. 3–5. [Google Scholar]
- Adam, P. Saltmarsh Ecology; Cambridge University Press: New York, NY, USA, 1990. [Google Scholar]
- Watson, G.E. Big thicket plant ecology: An introduction. In Temple Big Thicket Series #5, 3rd ed.; University of North Texas Press: Denton, TX, USA, 2006. [Google Scholar]
- National Institute of Environmental (NIE). Research on Coastal Landscape and the Conservational Strategy (II); Case of the West coast, Korea; Ministry of Environment: Sejong, Korea, 2008. [Google Scholar]
- National Institute of Environmental (NIE). Research on Coastal Landscape and the Conservational Strategy (2007–2009); Ministry of Environment: Sejong, Korea, 2009. [Google Scholar]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda Mdel, C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Bloemberg, G.V.; Lugtenberg, B.J. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol. 2001, 4, 343–350. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Rocha-Granados, M.D.C.; Glick, B.R.; Santoyo, G. Microbiome engineering to improve biocontrol and plant growth-promoting mechanisms. Microbiol. Res. 2018, 208, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Park, J.M.; Hong, J.W.; Son, J.S.; Hwang, Y.J.; Cho, H.M.; You, Y.H.; Ghim, S.Y. A strategy for securing unique microbial resources—Focusing on Dokdo islands-derived microbial resources. Israel J. Ecol. Evol. 2018, 64, 1–15. [Google Scholar] [CrossRef]
- Ryan, M.J.; McCluskey, K.; Verkleij, G.; Robert, V.; Smith, D. Fungal biological resources to support international development: Challenges and opportunities. World J. Microbiol. Biotechnol. 2019, 35, 139. [Google Scholar] [CrossRef] [Green Version]
- Cultural Heritage Administration of Korea Website. Available online: http://www.heritage.go.kr/heri/idx/index.do (accessed on 21 October 2020).
- Amanda, S. Species Diversity Patterns at Ecotones. Master’s Thesis, University of North Carolina, Chapel Hill, NC, USA, 2009. [Google Scholar]
- Smith, R.L. Ecology and Field Biology, 2nd ed; Harper & Row: New York, NY, USA, 1974; p. 251. [Google Scholar]
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem: Ecology, Threats and Management; Oxford University Press: New York, NY, USA, 2004. [Google Scholar]
- Du, W.; Yao, Z.; Li, J.; Sun, C.; Xia, J.; Wang, B.; Shi, D.; Ren, L. Diversity and antimicrobial activity of endophytic fungi isolated from Securinega suffruticosa in the Yellow River Delta. PLoS ONE 2020, 15, e0229589. [Google Scholar] [CrossRef] [PubMed]
- Seena, S.; Sridhar, K.R. Endophytic fungal diversity of 2 sand dune wild legumes from the southwest coast of India. Can. J. Microbiol. 2004, 50, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, N.M.; Gutama, A.D.; Aysa, A. Endophytic fungal diversity isolated from different agro-ecosystem of Enset (Ensete ventericosum) in Gedeo zone, SNNPRS, Ethiopia. BMC Microbiol. 2019, 19, 172. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Luo, X.; Fan, M.; Zeng, W.; Yang, C.; Wu, J.; Zhao, C.; Zhang, Y.; Zhao, P. Endophytic fungi from the ranches of Camellia taliensis (W. W. Smith) Melchior, a widely distributed wild tea plant. World J. Microbiol. Biotechnol. 2019, 35, 113. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wei, Z.; Pu, S.; Xiang, M.; Yan, A.; Zhang, Y.; Wang, X. Diversity of endophytic fungi of Paeonialactiflora pallas and screening for fungal paeoniflorin producers. FEMS Microbiol. Lett. 2018, 365. [Google Scholar] [CrossRef]
- Fan, M.; Chen, X.; Luo, X.; Zhang, H.; Liu, Y.; Zhang, Y.; Wu, J.; Zhao, C.; Zhao, P. Diversity of endophytic fungi from the leaves of Vaccinium dunalianum. Lett. Appl. Microbiol. 2020, 71, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Ogura, T.; Degawa, Y.; Ohmasa, M. Isolation of Tricholoma matsutake and T. bakamatsutake cultures from field-collected ectomycorrhizas. Mycoscience 2001, 42, 43–50. [Google Scholar] [CrossRef]
- Hasan, H.A. Gibberellin and auxin production plant root fungi and their biosynthesis under salinity-calcium interaction. Rostl. Vyroba. 2002, 48, 101–106. [Google Scholar] [CrossRef]
- Khan, S.A.; Hamayun, M.; Yoon, H.; Kim, H.Y.; Suh, S.J.; Hwang, S.K.; Kim, J.M.; Lee, I.J.; Choo, Y.S.; Yoon, U.H.; et al. Plant growth promotion and Penicillium citrinum. BMC Microbiol. 2008, 8, 231. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- National Center for Biotechnology Information (NCBI) U.S. National Library of Medicine. GenBank Database. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 7 October 2020).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, Y.; Ye, W.; Zhang, Y.; Xu, Y. High speed BLASTN: An accelerated Mega BLAST search tool. Nucleic Acids Res. 2015, 18, 7762–7768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aiyar, A. The use of CLUSTAL W and CLUSTAL X for multiple sequence alignment. Methods Mol. Biol. 2000, 132, 221–241. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Rzhetsky, A.; Nei, M. A simple method for estimating and testing minimum evolution trees. Mol. Biol. Evol. 1992, 9, 945–967. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Tajima, F.; Nei, M. Estimation of evolutionary distance between nucleotide sequences. Mol. Biol. Evol. 1984, 1, 269–285. [Google Scholar] [CrossRef] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Mendes, R.S.; Evangelista, L.R.; Thomaz, S.M.; Agostinho, A.A.; Gomes, L.C. Unified index to measure ecological diversity and species rarity. Ecography 2008, 31, 450–456. [Google Scholar] [CrossRef]
- Mendes, S.M.; Santos, J.; Freitas, H.; Sousa, J.P. Assessing the impact of understory vegetation cut on soil epigeic macrofauna from a cork-oak Montado in South Portugal. Agroforest. Syst. 2011, 82, 139–148. [Google Scholar] [CrossRef]
- Whittaker, R.H. Evolution of species diversity in land communities. Evol. Biol. 1972, 10, 1–67. [Google Scholar]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Science: Oxford, NJ, USA, 2004. [Google Scholar]
- Lambshead, P.J.D.; Platt, H.M.; Shaw, K.M. The detection of differences among assemblages of marine benthic species based on an assessment of dominance and diversity. J. Nat. Hist. 1983, 17, 859–874. [Google Scholar] [CrossRef]
- International Plant Names Index. Available online: http://www.ipni.org (accessed on 7 August 2020).
- Kaiser, M.J.; Attrill, M.J.; Jennings, S.; Thomas, D.N. Marine Ecology: Processes, Systems and Impacts; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- Ma, B.; Lv, X.; Warren, A.; Gong, J. Shifts in diversity and community structure of endophytic bacteria and archaea across root, stem and leaf tissues in the common reed, Phragmites australis, along a salinity gradient in a marine tidal wetland of northern China. Antonie Van Leeuwenhoek 2013, 104, 759–768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.R.; Zhu, Z.N.; Li, Y.Q.; Xiao, M.; Han, M.X.; Wadaan, M.A.M.; Hozzein, W.N.; An, D.D.; Li, W.J. Microbacterium halophytorum sp. nov., a novel endophytic actinobacterium isolated from halophytes. Int. J. Syst. Evol. Microbiol. 2018, 68, 3928–3934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Y.; Liu, F.; Li, Y.Q.; Lei, R.F.; Ma, Q.; Narsing Rao, M.P.; Abudourousuli, D.; Rouzi, Z.; Aosiman, M.; An, D.D.; et al. Sphingobacterium endophyticum sp. nov., a novel endophyte isolated from halophyte. Arch. Microbiol. 2020, 202, 2771–2778. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Zhang, W.D.; Qin, S.; Bian, G.K.; Xing, K.; Li, Y.F.; Cao, C.L.; Jiang, J.H. Saccharopolyspora dendranthemae sp. nov. a halotolerant endophytic actinomycete isolated from a coastal salt marsh plant in Jiangsu, China. Antonie Van Leeuwenhoek 2013, 103, 1369–1376. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.N.; Li, Y.R.; Li, Y.Q.; Xiao, M.; Han, M.X.; Wadaan, M.A.M.; Hozzein, W.N.; An, D.D.; Li, W.J. Microbacterium suaedae sp. nov., isolated from Suaeda aralocaspica. Int. J. Syst. Evol. Microbiol. 2019, 69, 411–416. [Google Scholar] [CrossRef] [PubMed]
- MOE (Ministry of Environment. Korea). The Fifth National Report to the Convention on Biological Diversity. 2014. Available online: https://www.cbd.int/nr5/ (accessed on 23 April 2014).
- Chi, L.P.; Li, X.M.; Li, L.; Li, X.; Wang, B.G. Cytotoxic thiodiketopiperazine derivatives from the deep sea-derived fungus Epicoccum nigrum SD-388. Mar. Drugs 2020, 18, 160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Li, Q.; Yang, H.; Cai, X.L.; Xia, X.K.; Chen, S.P.; Li, M.F.; She, Z.G.; Lin, Y.C. Structure elucidation of three diphenyl ether derivatives from the mangrove endophytic fungus SBE-14 from the South China Sea. Magn. Reson. Chem. 2009, 247, 453–455. [Google Scholar] [CrossRef] [PubMed]
- Afiyatullov, S.S.; Leshchenko, E.V.; Berdyshev, D.V.; Sobolevskaya, M.P.; Antonov, A.S.; Denisenko, V.A.; Popov, R.S.; Pivkin, M.V.; Udovenko, A.A.; Pislyagin, E.A.; et al. Zosteropenillines: Polyketides from the marine derived fungus Penicillium thomii. Mar. Drugs 2017, 15, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabello, M.; Irrazabal, G.; Bucsinszky, A.M.; Saparrat, M.; Schalamuk, S. Effect of an arbuscular mycorrhizal fungus, Glomus mosseae, and a rock-phosphate-solubilizing fungus, Penicillium thomii, on Mentha piperita growth in a soilless medium. J. Basic Microbiol. 2005, 45, 182–189. [Google Scholar] [CrossRef]
- Cheng, Z.; Liu, W.; Fan, R.; Han, S.; Li, Y.; Cui, X.; Zhang, J.; Wu, Y.; Lv, X.; Zhang, Y.; et al. Terpenoids from the deep-sea-derived fungus Penicillium thomii YPGA3 and their bioactivities. Mar. Drugs 2020, 18, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicoletti, R.; Vinale, F. Bioactive compounds from marine-derived Aspergillus, Penicillium, Talaromyces and Trichoderma species. Mar. Drugs 2018, 16, 408. [Google Scholar] [CrossRef] [Green Version]
- Park, M.S.; Oh, S.Y.; Fong, J.J.; Houbraken, J.; Lim, Y.W. The diversity and ecological roles of Penicillium in intertidal zones. Sci. Rep. 2019, 9, 13540. [Google Scholar] [CrossRef] [Green Version]
- Zhuravleva, O.I.; Sobolevskaya, M.P.; Leshchenko, E.V.; Kirichuk, N.N.; Denisenko, V.A.; Dmitrenok, P.S.; Dyshlovoy, S.A.; Zakharenko, A.M.; Kim, N.Y.; Afiyatullov, S.S. Meroterpenoids from the alga-derived fungi Penicillium thomii Maire and Penicillium lividum Westling. J. Nat. Prod. 2014, 77, 1390–1395. [Google Scholar] [CrossRef] [PubMed]
- Ek-Ramos, M.J.; Zhou, W.; Valencia, C.U.; Antwi, J.B.; Kalns, L.L.; Morgan, G.D.; Kerns, D.L.; Sword, G.A. Spatial and temporal variation in fungal endophyte communities isolated from cultivated cotton (Gossypium hirsutum). PLoS ONE 2013, 8, e66049. [Google Scholar] [CrossRef] [PubMed]
- Pan, F.; Liu, Z.Q.; Chen, Q.; Xu, Y.W.; Hou, K.; Wu, W. Endophytic fungus strain 28 isolated from Houttuynia cordata possesses wide-spectrum antifungal activity. Braz. J. Microbiol. 2016, 47, 480–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species | Sampling Sites | GPS Coordinates | Fungal Isolates and Taxon | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Total Isolate Number | Division | Class | Order | Family | Genus | ||||
| Phragmites australis | Delta | 1 | N 34°53′00.20″/E 127°30′49.60″ | 25 | 1 | 4 | 8 | 14 | 18 |
| 2 | N 34°52′55.78″/E 127°30′50.43″ | 25 | 1 | 4 | 7 | 11 | 17 | ||
| 3 | N 34°52′54.36″/E 127°30′53.09″ | 36 | 3 | 6 | 8 | 13 | 16 | ||
| 4 | N 34°52′53.69″/E 127°30′55.98″ | 9 | 1 | 3 | 4 | 4 | 5 | ||
| 5 | N 34°52′55.63″/E 127°30′52.68″ | 22 | 1 | 3 | 6 | 8 | 11 | ||
| 6 | N 34°52′59.46″/E 127°30′52.38″ | 6 | 1 | 2 | 3 | 4 | 4 | ||
| Tidal flat | 7 | N 34°52′38.90″/E 127°30′47.23″ | 9 | 1 | 3 | 5 | 6 | 6 | |
| 8 | N 34°52′34.81″/E 127°30′27.08″ | 13 | 1 | 2 | 4 | 9 | 9 | ||
| 9 | N 34°52′27.65″/E 127°30′14.77″ | 11 | 1 | 3 | 4 | 5 | 6 | ||
| 10 | N 34°52′27.46″/E 127°29′55.06″ | 16 | 1 | 3 | 5 | 7 | 10 | ||
| 11 | N 34°52′27.51″/E 127°29′46.53″ | 8 | 1 | 3 | 4 | 5 | 6 | ||
| Suaeda japonica | Tidal flat | 1 | N 34°51′45.94″/E 127°31′10.12″ | 15 | 2 | 5 | 7 | 8 | 10 |
| 2 | N 34°51′46.41″/E 127°31′09.65″ | 18 | 1 | 3 | 7 | 8 | 11 | ||
| 3 | N 34°51′49.84″/E 127°31′09.49″ | 31 | 2 | 5 | 8 | 12 | 15 | ||
| 4 | N 34°51′56.41″/E 127°31′06.25″ | 29 | 2 | 5 | 8 | 10 | 13 | ||
| 5 | N 34°51′59.83″/E 127°31′06.25″ | 23 | 2 | 5 | 8 | 11 | 15 | ||
| 6 | N 34°51′59.73″/E 127°31′06.36″ | 28 | 1 | 3 | 6 | 7 | 11 | ||
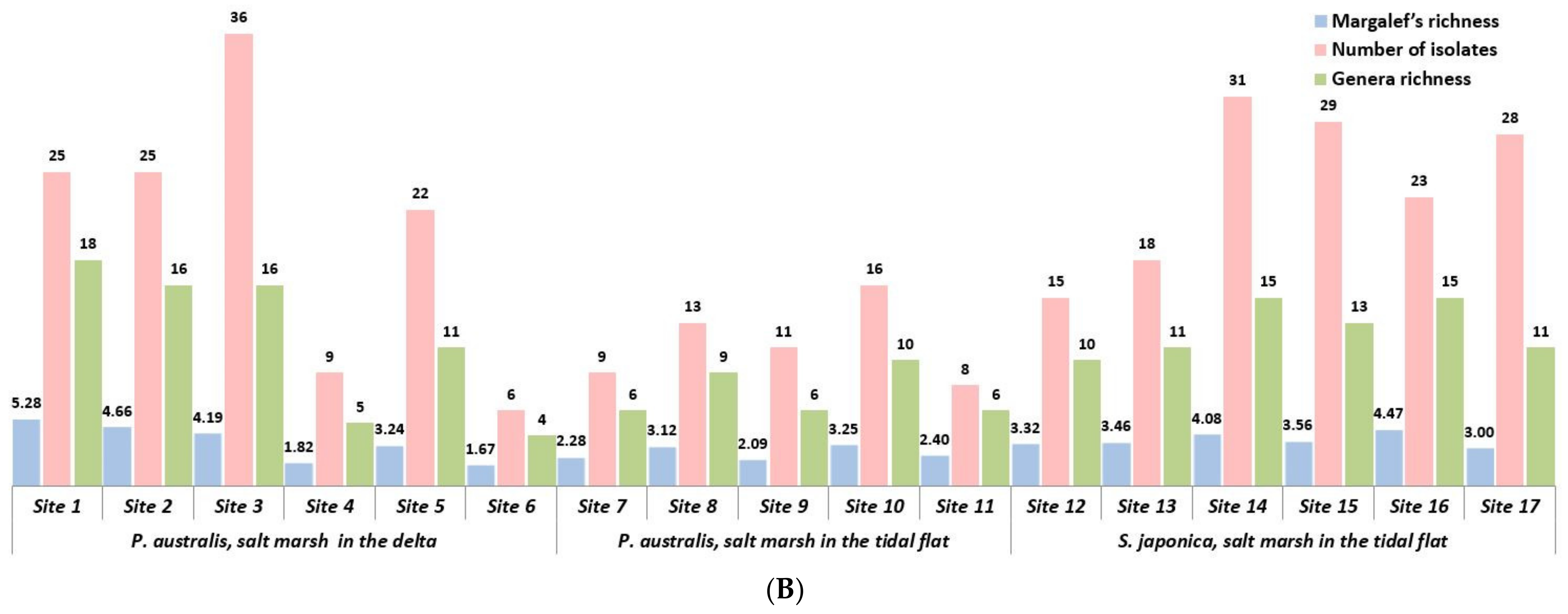
| Host Species | Sampling Site Character | Site Number | Number of Isolates | Genera Richness | Margalef’s Richness | Menhinick’s Index (Dmn) | Shannon’s Index (H’) | Simpson’s Index (D) |
|---|---|---|---|---|---|---|---|---|
| Phragmites australis | Delta | 1 | 25 | 18 | 5.281 | 3.600 | 2.080 | 0.027 |
| 2 | 25 | 16 | 4.660 | 1.919 | 1.919 | 0.037 | ||
| 3 | 36 | 16 | 4.186 | 2.667 | 1.434 | 0.081 | ||
| 4 | 9 | 5 | 1.820 | 1.667 | 1.971 | 0.139 | ||
| 5 | 22 | 11 | 3.235 | 2.345 | 1.940 | 0.082 | ||
| 6 | 6 | 4 | 1.674 | 1.633 | 1.663 | 0.133 | ||
| Tidal flat | 7 | 9 | 6 | 2.276 | 2.000 | 1.739 | 0.111 | |
| 8 | 13 | 9 | 3.119 | 2.496 | 1.623 | 0.064 | ||
| 9 | 11 | 6 | 2.085 | 1.809 | 1.838 | 0.145 | ||
| 10 | 6 | 10 | 3.246 | 2.500 | 1.595 | 0.083 | ||
| 11 | 8 | 6 | 2.404 | 2.121 | 1.507 | 0.107 | ||
| Suaeda japonica | Tidal flat | 12 | 15 | 10 | 3.323 | 2.582 | 2.011 | 0.086 |
| 13 | 18 | 11 | 3.460 | 2.593 | 1.572 | 0.118 | ||
| 14 | 31 | 15 | 4.077 | 2.694 | 1.293 | 0.075 | ||
| 15 | 29 | 13 | 3.564 | 2.414 | 1.724 | 0.076 | ||
| 16 | 23 | 15 | 4.465 | 3.128 | 1.502 | 0.047 | ||
| 17 | 28 | 11 | 3.001 | 2.079 | 1.458 | 0.106 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-M.; Hong, J.-W.; You, Y.-H.; Kim, J.-G. Endophytic Fungi of Emersed Halophytes in River Deltas and Tidal Flats of the Korean Ramsar Wetlands. J. Mar. Sci. Eng. 2021, 9, 430. https://doi.org/10.3390/jmse9040430
Park J-M, Hong J-W, You Y-H, Kim J-G. Endophytic Fungi of Emersed Halophytes in River Deltas and Tidal Flats of the Korean Ramsar Wetlands. Journal of Marine Science and Engineering. 2021; 9(4):430. https://doi.org/10.3390/jmse9040430
Chicago/Turabian StylePark, Jong-Myong, Ji-Won Hong, Young-Hyun You, and Jong-Guk Kim. 2021. "Endophytic Fungi of Emersed Halophytes in River Deltas and Tidal Flats of the Korean Ramsar Wetlands" Journal of Marine Science and Engineering 9, no. 4: 430. https://doi.org/10.3390/jmse9040430