
Morphological Traits and Capture Depth of the Norwegian Skate (Dipturus nidarosiensis (Storm, 1881)) from Two Mediterranean Populations

 ,
,  , , , , and
, , , , and
Abstract
:1. Introduction
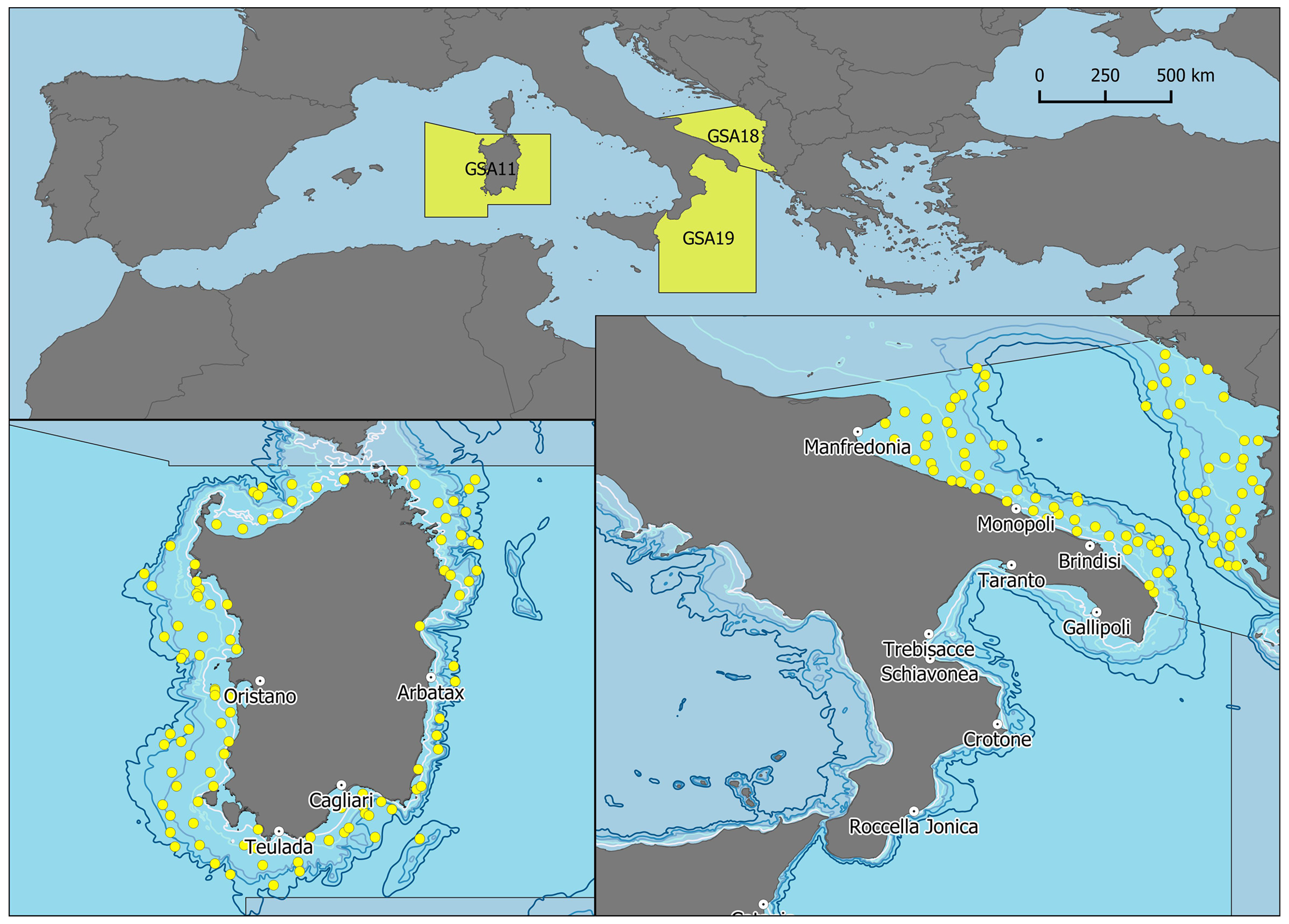
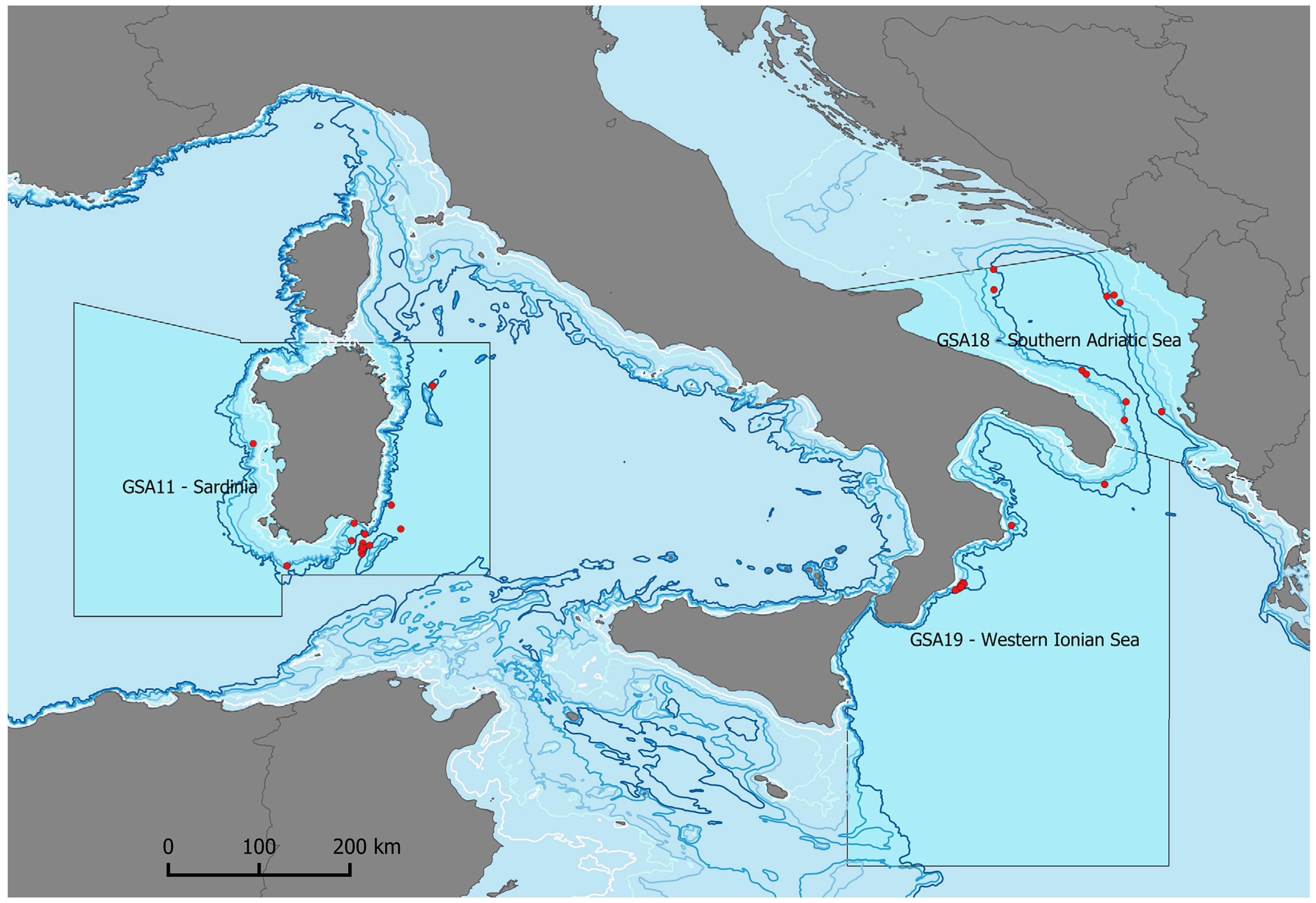
2. Materials and Methods
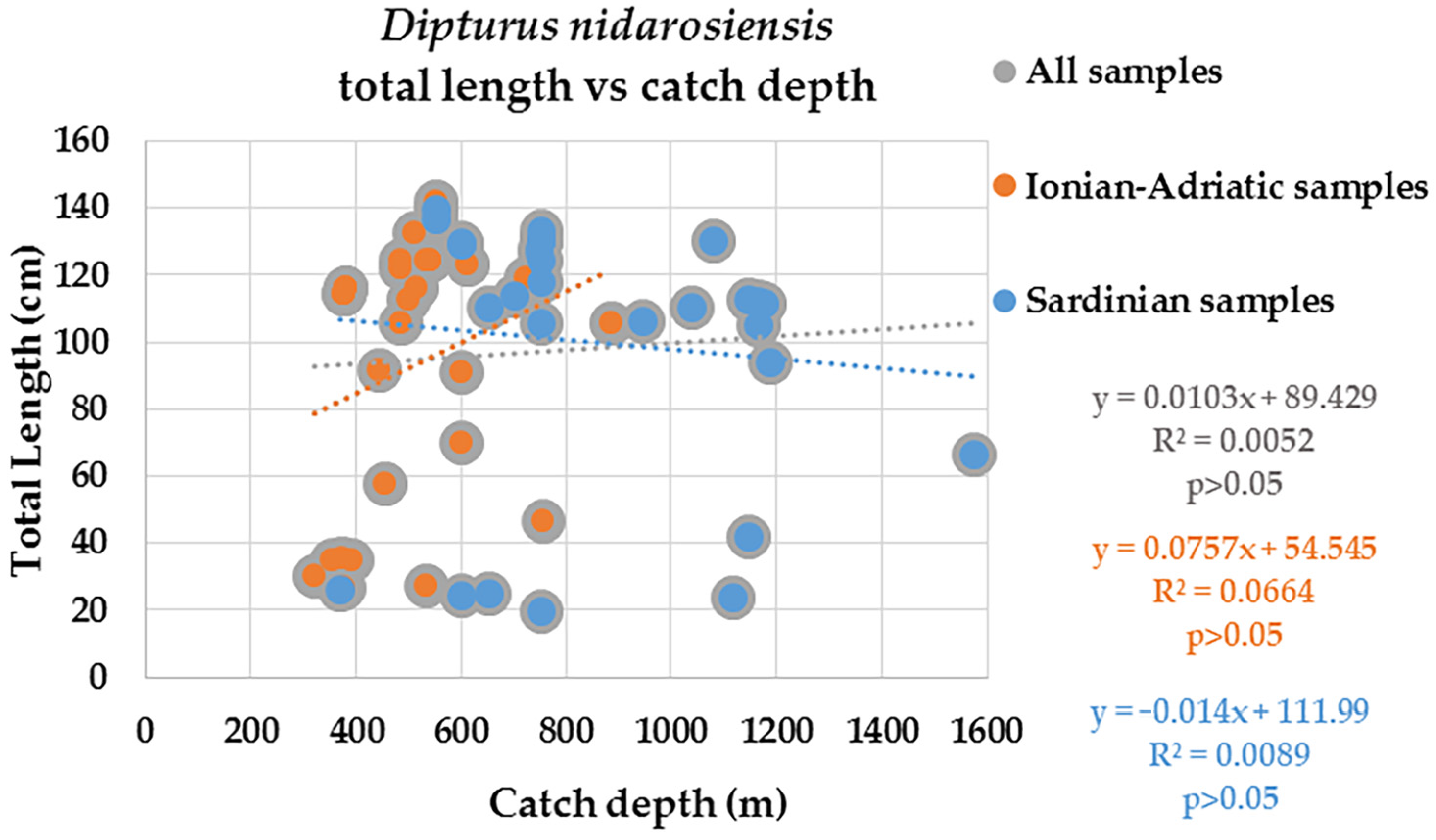
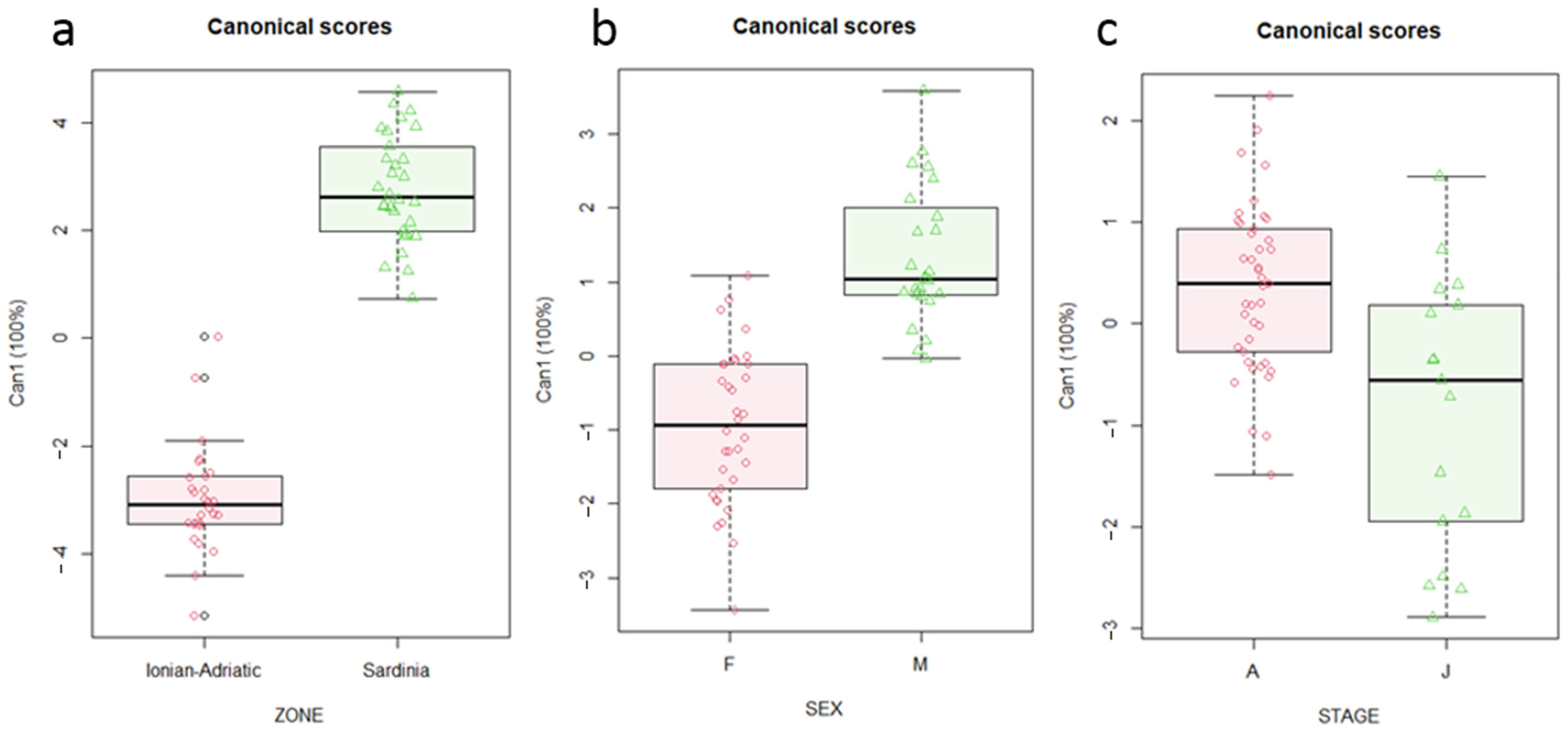
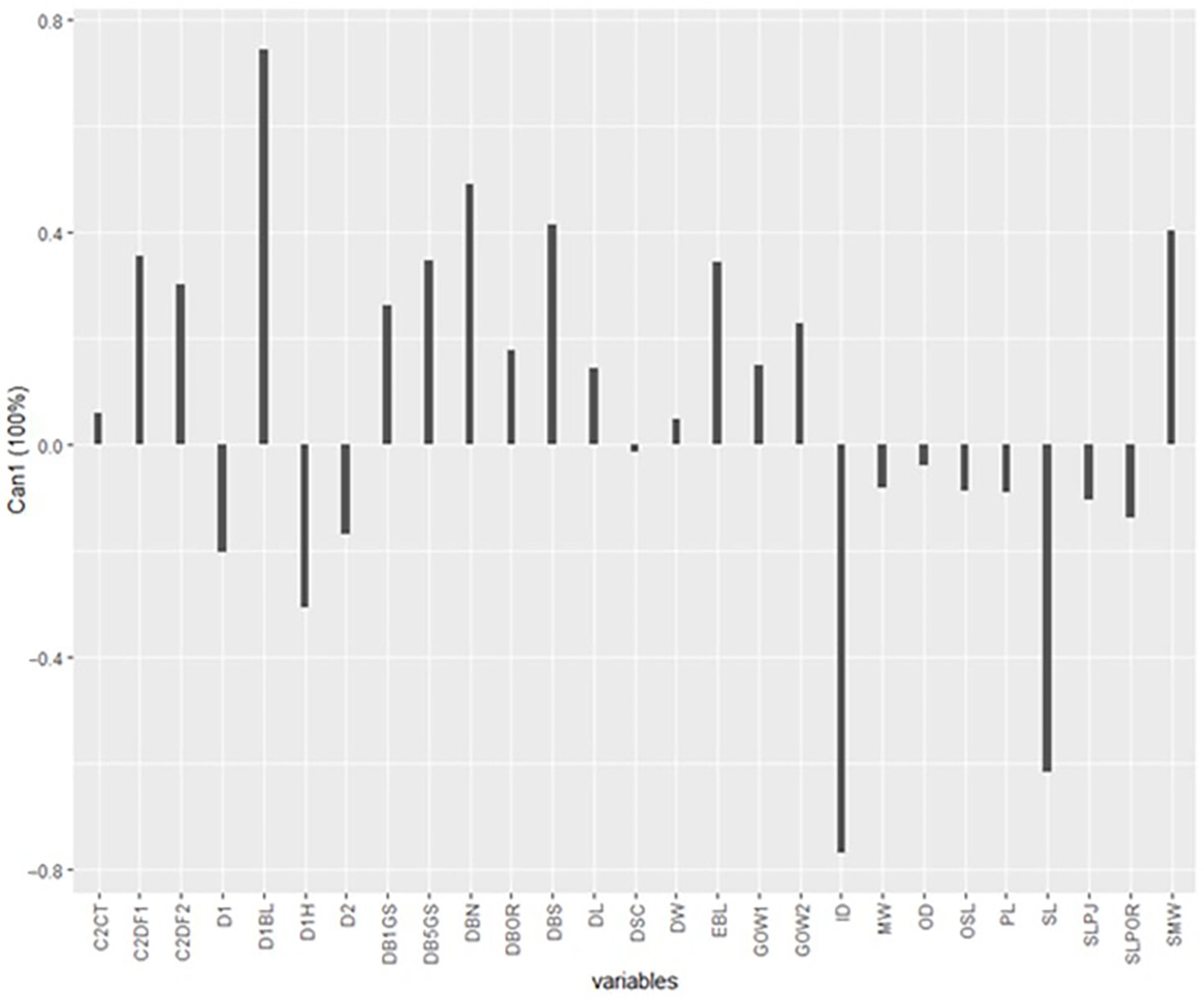
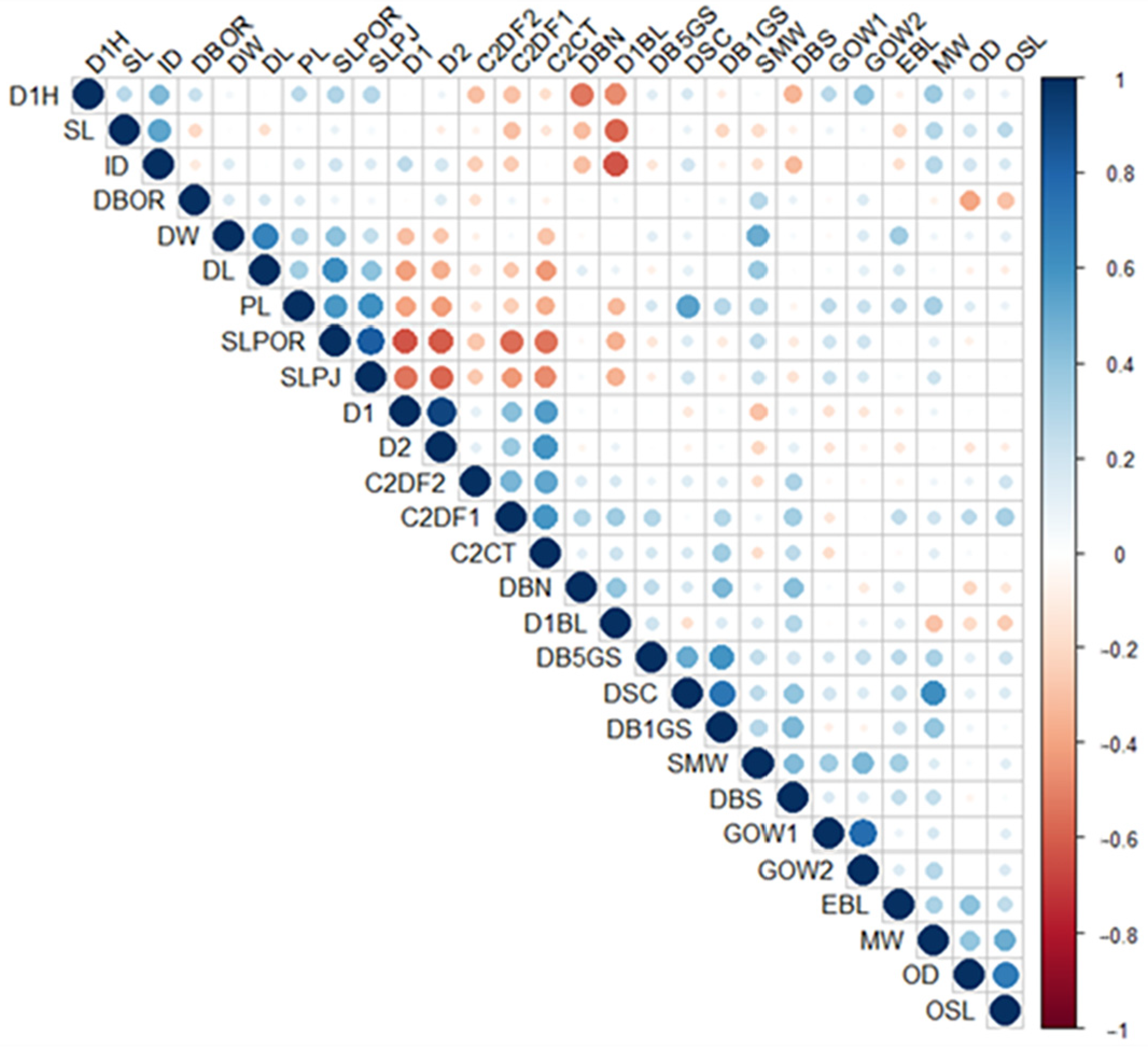
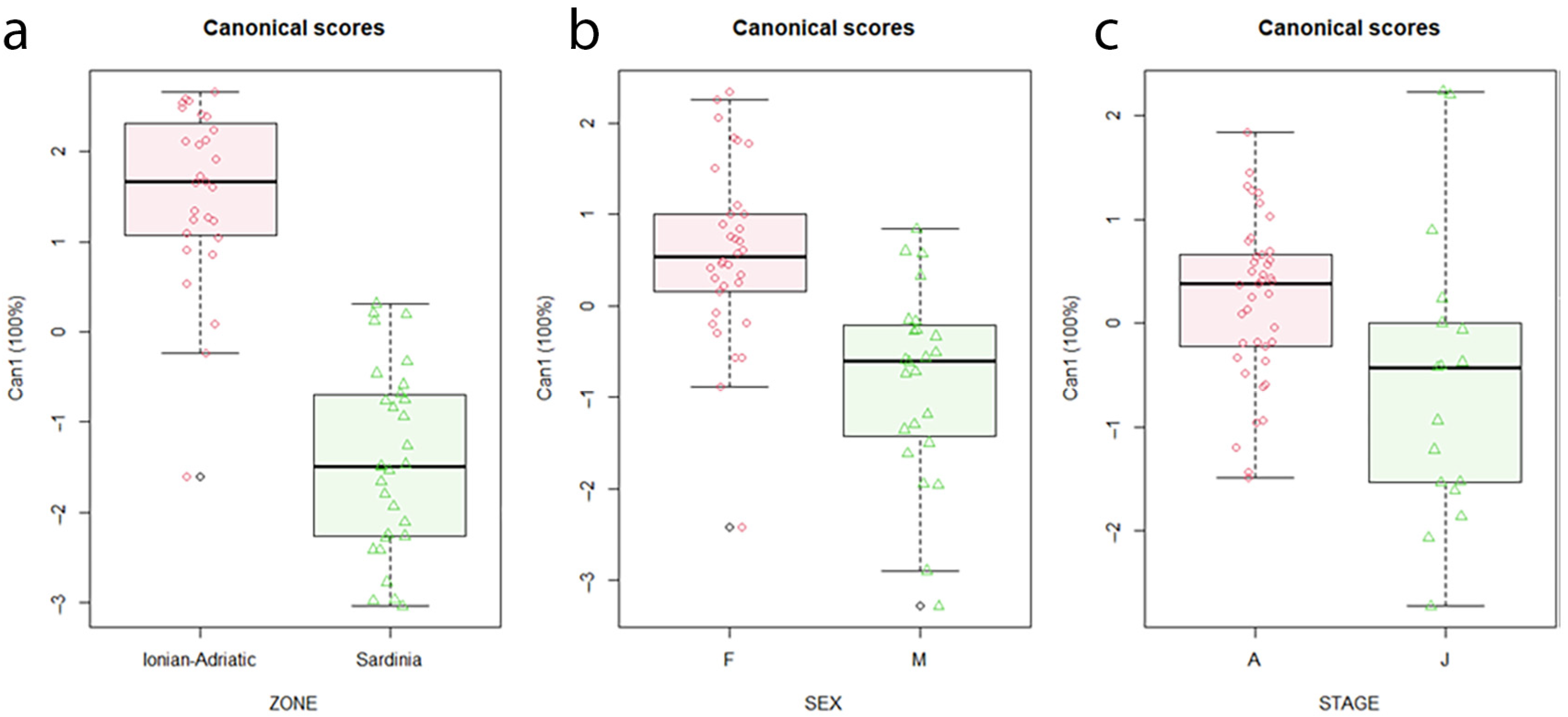
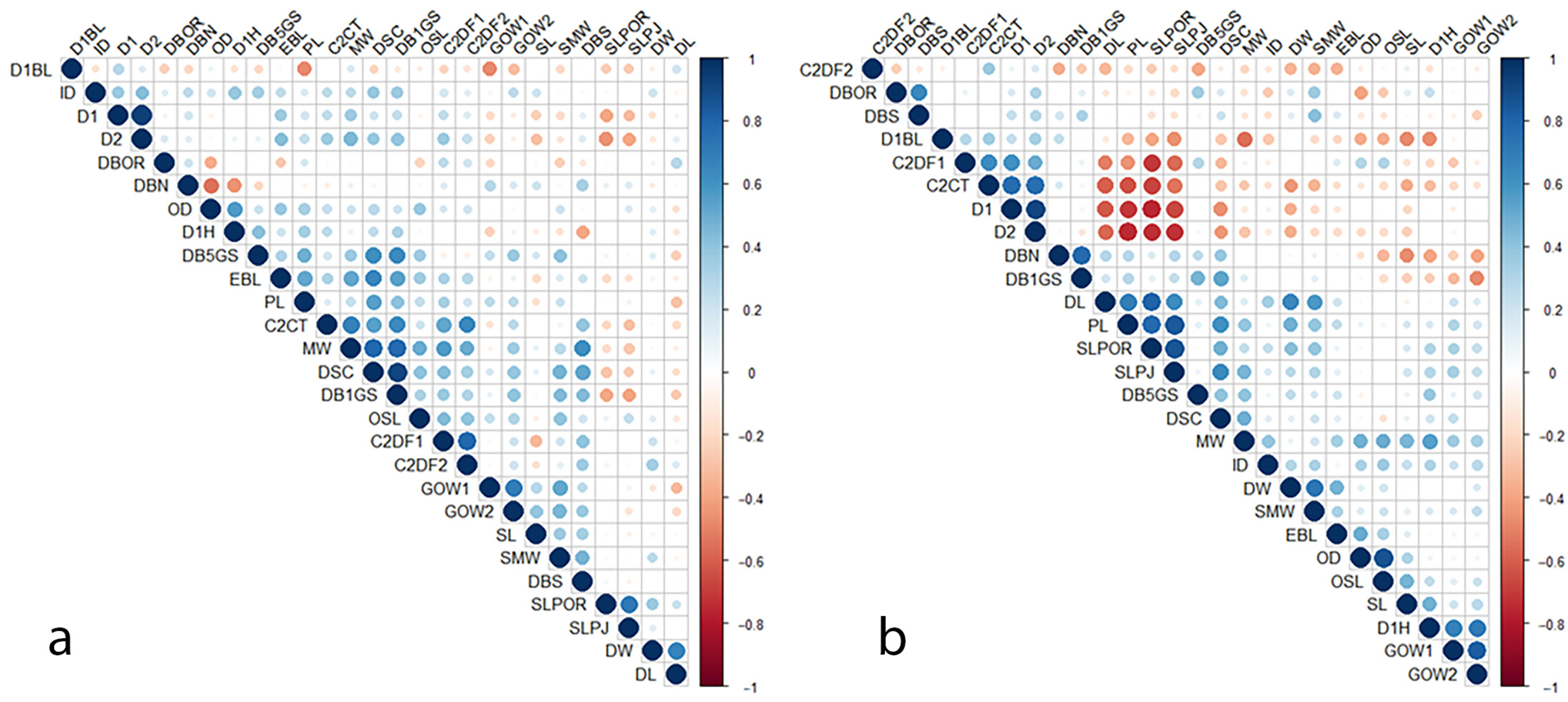
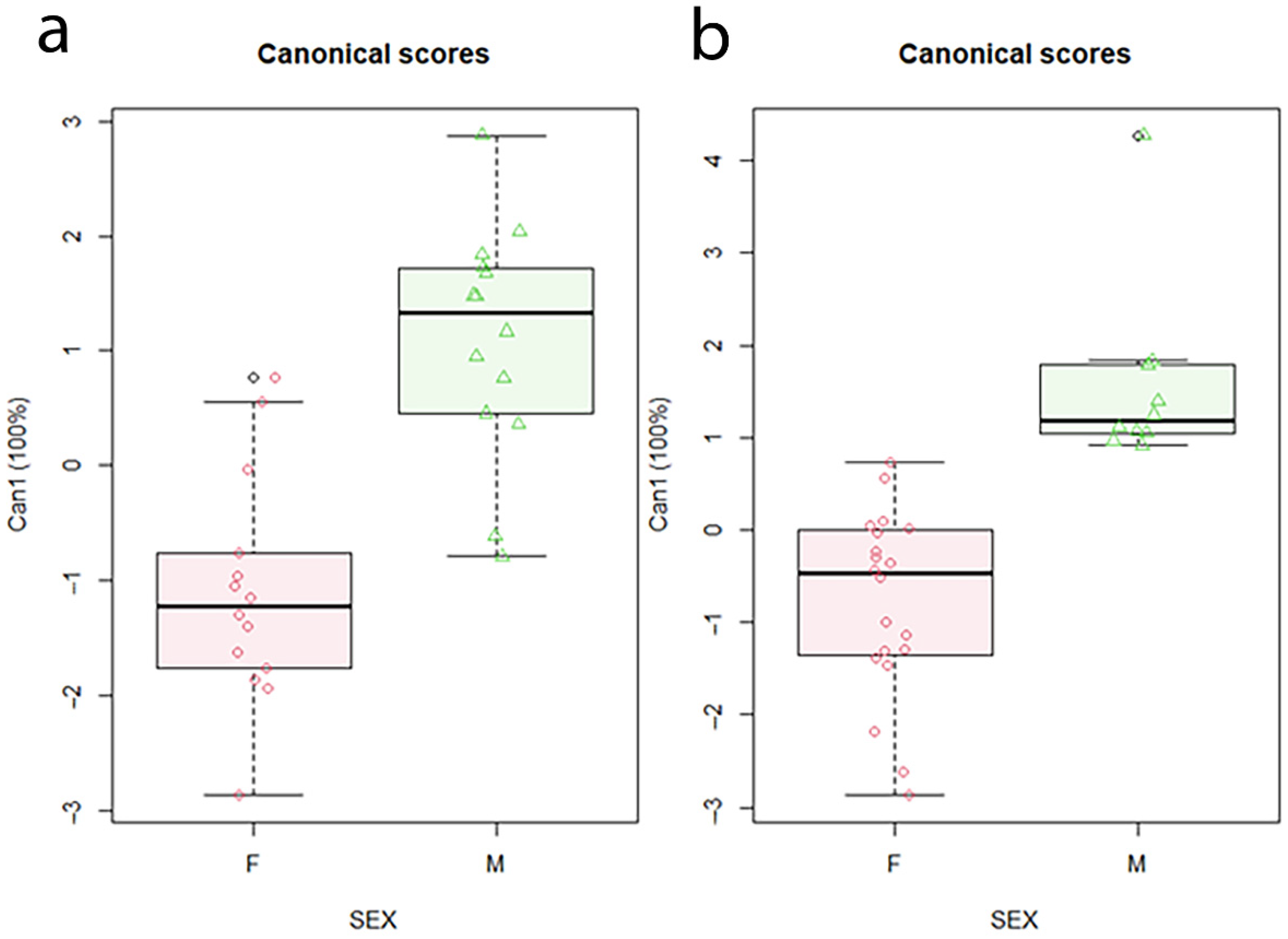
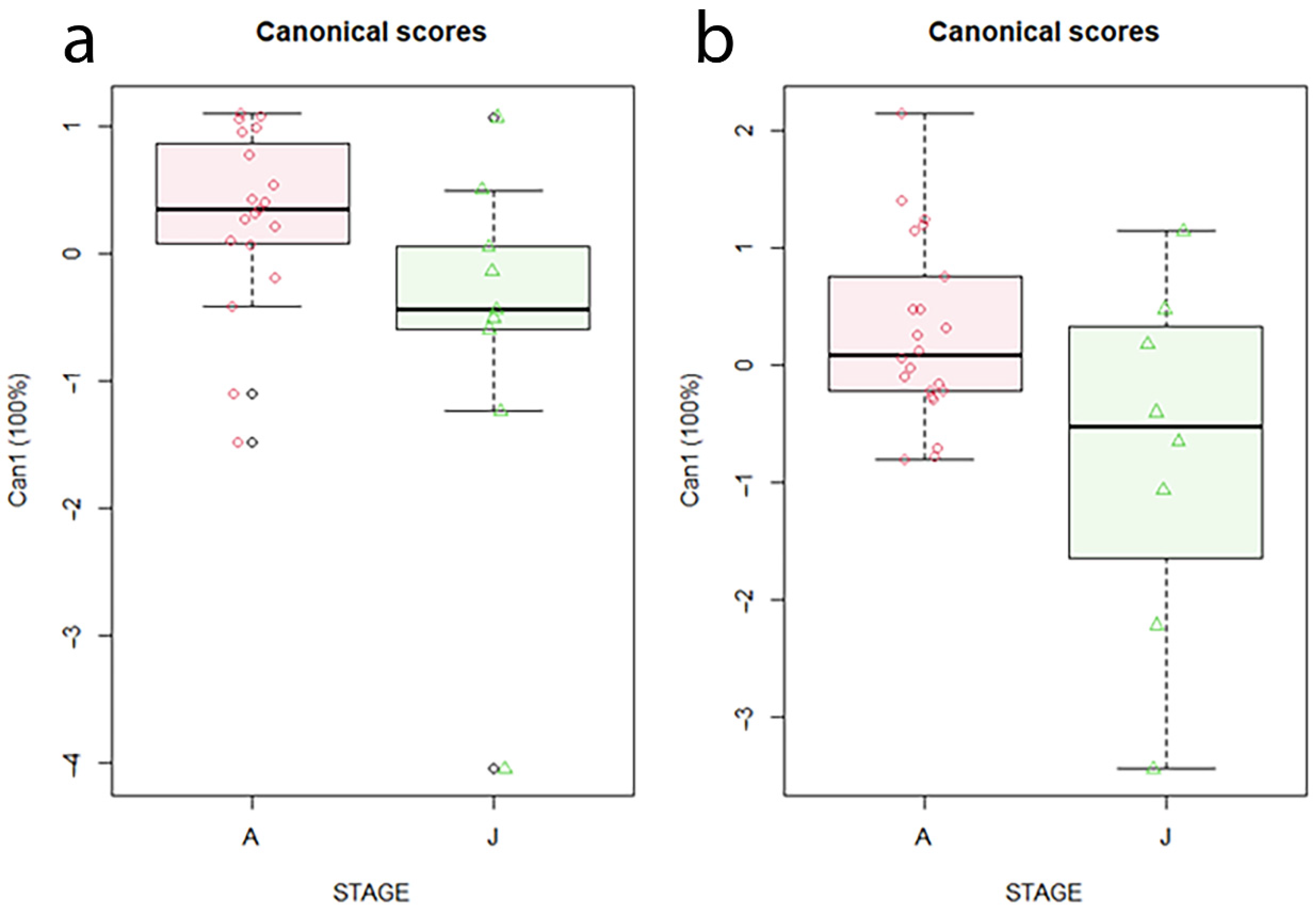
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stehmann, M. Rajidae. In Check-List of the Fishes of the Eastern Tropical Atlantic; Quero, J.C., Hureau, J.C., Karrer, C., Post, A., Saldanha, L., Eds.; Junta Nacional de Investigacao Cientifica e Tecnológica: Lisbon, Portugal, 1990; Volume 1, pp. 29–50. [Google Scholar]
- Stehmann, M.; Bürkel, D.L. Rajidae. In Fishes of the Northeastern Atlantic and Mediterranean; Whitehead, P.J., Bauchot, M.L., Hureau, J.C., Tortonese, E., Eds.; Unesco: Paris, France, 1984; pp. 163–196. [Google Scholar]
- Priede, I.; Godbold, J.A.; King, N.J.; Collins, M.A.; Baile, Y.D.M.; Gordon, J.D.M. Deep-sea demersal fish species richness in the Porcupine Seabight, NE Atlantic Ocean: Global and regional patterns. Mar. Ecol. 2010, 31, 247–260. [Google Scholar] [CrossRef]
- Rodríguez-Cabello, C.; Pérez, M.; Sánchez, F. New records of chondrichthyans species caught in the Cantabrian Sea (southern Bay of Biscay). J. Mar. Biolog. Assoc. 2013, 93, 1929–1939. [Google Scholar] [CrossRef]
- Cannas, R.; Follesa, M.C.; Cabiddu, S.; Porcu, C.; Salvadori, S.; Iglesias, S.P.; Deiana, A.M.; Cau, A. Molecular and morphological evidence of the occurrence of the Norwegian skate Dipturus nidarosiensis in the Mediterranean Sea. Mar. Biol. Res. 2010, 6, 341–350. [Google Scholar] [CrossRef]
- Follesa, M.C.; Cannas, R.; Cabiddu, S.; Cau, A.; Mulas, A.; Porcu, C.; Cau, A. Preliminary observations of the reproductive biology and diet for the Norwegian skate Dipturus nidarosiensis (Rajidae) from the Central Western Mediterranean Sea. Cybium 2012, 36, 473–477. [Google Scholar]
- Cariani, A.; Messinetti, S.; Ferrari, A.; Arculeo, M.; Bonello, J.J.; Bonnici, L.; Cannas, R.; Carbonara, P.; Cau, A.; Charilaou, C.; et al. Improving the Conservation of Mediterranean Chondrichthyans: The ELASMOMED DNA Barcode Reference Library. PLoS ONE 2017, 12, 170244. [Google Scholar] [CrossRef]
- Marongiu, M.F.; Porcu, C.; Bellodi, A.; Cannas, R.; Cau, A.; Cuccu, D.; Mulas, A.; Follesa, M.C. Temporal dynamics of demersal chondrichthyan species in the central western Mediterranean Sea: The case study in Sardinia Island. Fish. Res. 2017, 193, 81–94. [Google Scholar] [CrossRef]
- Porcu, C.; Marongiu, M.F.; Bellodi, A.; Cannas, R.; Cau, A.; Melis, R.; Mulas, A.; Soldovilla, G.; Vacca, L.; Follesa, M.C. Morphological descriptions of the eggcases of skates (Rajidae) from the central-western Mediterranean, with notes on their distribution. Helgol. Mar. Res. 2017, 71, 10. [Google Scholar] [CrossRef] [Green Version]
- Porcu, C.; Marongiu, M.F.; Olita, A.; Bellodi, A.; Cannas, R.; Carbonara, P.; Cau, A.; Mulas, A.; Pesci, P.; Follesa, M.C. The demersal bathyal fish assemblage of the Central-Western Mediterranean: Depth distribution, sexual maturation and reproduction. Deep-Sea Res. I Oceanogr. Res. Pap. 2020, 166, 103394. [Google Scholar] [CrossRef]
- Mulas, A.; Bellodi, A.; Carbonara, P.; Cau, A.; Marongiu, M.F.; Pesci, P.; Porcu, C.; Follesa, M.C. Bio-Ecological Features Update on Eleven Rare Cartilaginous Fish in the Central-Western Mediterranean Sea as a Contribution for Their Conservation. Life 2021, 11, 871. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Amaro, S.; Ordines, F.; Puerto, M.Á.; García, C.; Ramon, C.; Terrasa, B.; Massutí, E. New morphological and molecular evidence confirm the presence of the Norwegian skate Dipturus nidarosiensis (Storm, 1881) in the Mediterranean Sea and extend its distribution to the western basin. Mediterr. Mar. Scie. 2017, 18, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Carbonara, P.; Cannas, R.; Donnaloia, M.; Melis, R.; Porcu, C.; Spedicato, M.T.; Zupa, W.; Follesa, M.C. On the presence of Dipturus nidarosiensis (Storm, 1881) in the Central Mediterranean area. PeerJ 2019, 7, 7009. [Google Scholar] [CrossRef] [Green Version]
- Isajlović, I.; Dulčić, J.; Piccinetti, C.; Vrgoč, N.; Manfredi, C.; Dragičević, B. Additional records of Norwegian skate Dipturus nidarosiensis (Storm, 1881) (Pisces: Rajidae) in the Adriatic Sea. Acta Adriat. 2020, 61, 217–222. [Google Scholar] [CrossRef]
- Geraci, M.L.; Di Lorenzo, M.; Falsone, F.; Scannella, D.; Di Maio, F.; Colloca, F.; Vitale, S.; Serena, F. The occurrence of norwegian skate, Dipturus nidarosiensis (Elasmobranchii: Rajiformes: Rajidae), in the Strait of Sicily, Central Mediterranean. Acta Ichthyol. Piscat. 2019, 49, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Follesa, M.C.; Porcu, C.; Mulas, A.; Deiana, A.M.; Cau, A. Deep-water fish assemblages in the central-western Mediterranean (south Sardinian deepwaters). J. Appl. Ichthyol. 2011, 27, 129–135. [Google Scholar] [CrossRef]
- Iglesias, S.P.; Toulhoat, L.; Sellos, D.Y. Taxonomic confusion and market mislabeling of threatened skates: Important consequences for their conservation status. Aquat. Conserv. 2010, 20, 319–333. [Google Scholar] [CrossRef]
- Griffiths, A.M.; Sims, D.W.; Cotterell, S.P.; El Nagar, A.; Ellis, J.R.; Lynghammar, A.; McHugh, M.; Neat, F.C.; Pade, N.G.; Queiroz, N.; et al. Molecular markers reveal spatially segregated cryptic species in a critically endangered fish, the common skate (Dipturus batis). Proc. Royal Soc. 2010, 277, 1497–1503. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, F.; Osio, G.C.; Jenkins, C.J.; Rosenberg, A.A.; Lotze, H.K. Long-term change in a meso-predator community in response to prolonged and heterogeneous human impact. Sci. Rep. 2013, 3, 1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.; Fordham, S.V.; Francis, M.P.; et al. Extinction risk and conservation of the world’s sharks and rays. eLife 2014, 3, 00590. [Google Scholar] [CrossRef] [Green Version]
- Frodella, N.; Cannas, R.; Velonà, A.; Carbonara, P.; Farrell, E.D.; Fiorentino, F.; Follesa, M.C.; Garofalo, G.; Hemida, F.; Mancusi, C.; et al. Population connectivity and phylogeography of the Mediterranean endemic skate Raja polystigma and evidence of its hybridization with the parapatric sibling R. montagui. Mar. Ecol. Prog. Ser. 2016, 554, 99–113. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; deWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B 2003, 270, 96–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spouge, J.L. Measurement of a barcode’s accuracy in identifying species. In DNA Barcoding in Marine Perspectives; Trivedi, S., Ansari, A., Ghosh, S., Rehman, H., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Ball, R.E.; Serra-Pereira, B.; Ellis, J.; Genner, M.J.; Iglésias, S.; Johnson, A.F.; Jones, C.S.; Leslie, R.; Lewis, J.; Mariani, S.; et al. Resolving taxonomic uncertainty in vulnerable elasmobranchs: Are the Madeira skate (Raja maderensis) and the thornback ray (Raja clavata) distinct species? Conserv. Genet. 2016, 17, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Cerutti-Pereyra, F.; Meekan, M.G.; Wei, N.W.; O’Shea, O.; Bradshaw, C.J.; Austin, C.M. Identification of rays through DNA barcoding: An application for ecologists. PLoS ONE 2012, 7, 36479. [Google Scholar] [CrossRef] [Green Version]
- Ordines, F.; Baro, J.; Ramírez-Amaro, S.; Serena, F.; Sobrino, I. First substantiated record of Raja asterias Delaroche, 1809 (Elasmobranchii: Rajiformes: Rajidae) in the Gulf of Cádiz, North-eastern Atlantic. Acta Ichthyol. Piscat. 2017, 47, 101–106. [Google Scholar] [CrossRef] [Green Version]
- Vella, A.; Vella, N.; Schembri, S. A molecular approach towards taxonomic identification of elasmobranch species from Maltese fisheries landings. Mar. Genom. 2017, 36, 17–23. [Google Scholar] [CrossRef]
- Ward, R.D.; Holmes, B.H.; White, W.T.; Last, P.R. DNA barcoding Australasian chondrichthyans: Results and potential uses in conservation. Mar. Freshw. Res. 2008, 59, 57–71. [Google Scholar] [CrossRef]
- Stehmann, M.F.W.; Ellis, J.; Walls, R.; Lynghammar, A. Dipturus nidarosiensis . In The IUCN Red List of Threatened Species; 2015; e.T161729A48927468; Available online: https://www.iucnredlist.org/resources/media (accessed on 2 December 2021).
- Williams, T.; Helle, K.; Aschan, M. The distribution of chondrichthyans along the northern coast of Norway. ICES J. Mar. Sci. 2008, 65, 1161–1174. [Google Scholar] [CrossRef] [Green Version]
- Chaklader, M.R.; Siddik, M.A.B.; Nahar, A.; Hanif, M.A.; Alam, M.J.; Mahmud, M. Morphometric parameters and allometric growth in paradise threadfin Polynemus paradiseus (Linnaeus, 1758) from a coastal river of Bangladesh. J. Aquac. Res. Dev. 2016, 7, 417. [Google Scholar]
- Mir, J.I.; Saxena, N.; Patiyal, R.S.; Sahoo, P.K. Phenotypic differentiation of Barilius bendelisis (Cypriniformes: Cyprinidae) in four rivers from Central Indian Himalaya. Rev. Biol. Trop. (Int. J. Trop. Biol.) 2015, 63, 165–173. [Google Scholar]
- Vieira, A.R.; Rodrigues, A.S.B.; Sequeira, V.; Neves, A.; Paiva, R.B.; Paulo, O.S.; Serrano Gordo, L. Genetic and Morphological Variation of the Forkbeard, Phycis phycis (Pisces, Phycidae): Evidence of Panmixia and Recent Population Expansion along Its Distribution Area. PLoS ONE 2016, 11, e0167045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Lu, D.; Ma, L. Morphological differences between close populations discernible by multivariate analysis: A case study of genus Coilia (Teleostei: Clupeiforms). Aquat. Living Resour. 2005, 18, 187–192. [Google Scholar] [CrossRef]
- Siddik, M.A.B.; Hanif, M.A.; Chaklader, M.R.; Nahar, A.; Fotedar, R. A multivariate morphometric investigation to delineate stock structure of gangetic whiting, Sillaginopsis panijus (Teleostei: Sillaginidae). SpringerPlus 2016, 5, 520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AnvariFar, H.; Khyabani, A.; Farahmand, H.; Vatandoust, S.; AnvariFar, H.; Jahageerdar, S. Detection of morphometric differentiation between isolated up- and downstream populations of Siah Mahi (Capoeta capoeta gracilis) (Pisces: Cyprinidae) in the Tajan River (Iran). Hydrobiologia 2011, 673, 41–52. [Google Scholar] [CrossRef]
- Muchlisin, Z.A.; Zulkarnaini, B.; Purnawan, S.; Muhadjier, A.; Fadli, F.; Cheng, S.H. Morphometric variations of three species of harvested cephalopods found in northern sea of Aceh Province, Indonesia. Biodiversitas 2014, 15, 142–146. [Google Scholar] [CrossRef]
- Carugati, L.; Melis, R.; Cariani, A.; Cau, A.; Crobe, V.; Ferrari, A.; Follesa, M.C.; Geraci, M.L.; Iglésias, S.P.; Pesci, P.; et al. Combined COI barcode-based methods to avoid mislabelling of threatened species of deep-sea skates. Anim. Conserv. 2021. Available online: https://zslpublications.onlinelibrary.wiley.com/doi/full/10.1111/acv.12716 (accessed on 2 December 2021). [CrossRef]
- Cabral, H.N.; Marques, J.F.; Rego, A.L.; Catarino, A.L.; Figueiredo, J.; Garcia, J. Genetic and morphological variation of Synaptura lusitanica Capello, 1868, along the Portuguese coast. J. Sea. Res. 2003, 50, 167–175. [Google Scholar] [CrossRef]
- Bacha, M.; Jemaa, S.; Hamitouche, A.; Rabhi, K.; Amara, R. Population structure of the European anchovy, Engraulis encrasicolus, in the SW Mediterranean Sea, and the Atlantic Ocean: Evidence from otolith shape analysis. ICES J. Mar. Sci. 2014, 71, 2429–2435. [Google Scholar] [CrossRef]
- Pita, A.; Casey, J.; Hawkins, S.J.; Villarreal, M.R.; Gutiérrez, M.; Cabral, H.; Carocci, F.; Abaunza, P.; Pascual, P.; Presa, P. Conceptual and practical advances in fish stock delineation. Fish. Res. 2016, 173, 185–193. [Google Scholar] [CrossRef]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Ebert, D.A.; Fowler, S.; Compagno, L.J.V. Sharks of the World; Wild Nature Press: Plymouth, UK, 2013; p. 528. [Google Scholar]
- Follesa, M.C.; Carbonara, P. Atlas of the Maturity Stages of Mediterranean Fishery Resources. In Studies and Reviews N. 99; FAO: Rome, Italy, 2019; p. 268. [Google Scholar]
- Follesa, M.C.; Agus, B.; Bellodi, A.; Cannas, R.; Capezzuto, F.; Casciaro, L.; Cau, A.; Cuccu, D.; Donnaloia, M.; Fernandez-Arcaya, U.; et al. The MEDITS maturity scales as a useful tool for investigating the reproductive traits of key species in the Mediterranean Sea. Sci. Mar. 2019, 83S1, 235–256. [Google Scholar] [CrossRef] [Green Version]
- Froese, R. Cube law, condition factor and weight-length relationships: History, meta-analysis and recommendations. J. Appl. Ichthyol. 2006, 22, 241–253. [Google Scholar] [CrossRef]
- Oliveira, E.F.; Goulart, E.; Breda, L.; Minte-Vera, C.V.; de Souza Paiva, L.R.; Vismara, M.R. Ecomorphological patterns of the fish assemblage in a tropical floodplain: Effects of trophic, spatial and phylogenetic structures. Neotrop. Ichthyol. 2010, 8, 569–586. [Google Scholar] [CrossRef]
- Fisher, R. The use of multiple measurements in taxonomic problems. Ann. Eugen. 1936, 7, 179–188. [Google Scholar] [CrossRef]
- Elliott, N.G.; Haskard, K.; Koslow, J.A. Morphometric analysis of orange roughy (Hoplostethus atlanticus) off the continental slope of southern Australia. J. Fish Biol. 1995, 46, 202–220. [Google Scholar] [CrossRef]
- Asuero, A.G.; Sayago, A.; González, A.G. The Correlation Coefficient: An Overview. Crit. Rev. Anal. Chem. 2006, 36, 41–59. [Google Scholar] [CrossRef]
- Ellis, J.R.; Cruz-Martínez, A.; Rackham, B.D.; Rogers, S.I. The distribution of chondrichthyan fishes around the British Isles and implications for conservation. J. Northwest Atl. Fish. Sci. 2005, 37, 195–213. [Google Scholar] [CrossRef]
- Follesa, M.C.; Locci, I.; Pesci, P.; Floris, E.; Cau, A. Lo stato delle risorse nei mari italiani: GSA11–mari di Sardegna. In Lo Stato della Pesca e Dell’acquacoltura nei Mari Italiani, Monografie Scientifiche; Cataudella, S., Spagnolo, M., Eds.; Ministero delle Politiche Agricole, Alimentari e Forestali: Rome, Italy, 2011; pp. 132–140. [Google Scholar]
- Russo, T.; Bitetto, I.; Carbonara, P.; Carlucci, R.; D’Andrea, L.; Facchini, M.T.; Lembo, G.; Maiorano, P.; Sion, L.; Spedicato, M.T.; et al. A holistic approach to fishery management: Evidence and insights from a Central Mediterranean case study (Western Ionian Sea). Front. Mar. Sci. 2017, 4, 193. [Google Scholar] [CrossRef] [Green Version]
- Bensch, A.; Gianni, M.; Gréboval, D.; Sanders, J.S.; Hjort, A. Worldwide Review of Bottom Fisheries in the High Seas, FAO Fisheries and Aquaculture Technical Paper; Food and Agriculture Organization of the United Nations: Rome, Italy, 2009; Volume 1, p. 522. [Google Scholar]
- Artegiani, A.; Bregant, D.; Paschini, E.; Pinardi, N.; Raicich, F.; Russo, A. The Adriatic Sea general circulation. Part I: Air-sea interaction and water mass structure. J. Phys. Oceanogr. 1997, 27, 1492–1514. [Google Scholar] [CrossRef]
- Vilibic, I.; Orlic, M. Adriatic water masses, their rates of formation and transport through the Otranto Strait. Deep-Sea Res. I Oceanogr. Res. Pap. 2002, 16, 171–178. [Google Scholar] [CrossRef]
- Marra, A.; Mona, S.; Sà, R.M.; D’Onghia, G.; Maiorano, P. Population Genetic History of Aristeus antennatus (Crustacea: Decapoda) in the Western and Central Mediterranean Sea. PLoS ONE 2015, 10, e0128609. [Google Scholar]
- D’Onghia, G.; Maiorano, P.; Capezzuto, F.; Carlucci, R.; Battista, D.; Giove, A.; Sion, L.; Tursi, A. Further evidences of deep-sea recruitment of Aristeus antennatus (Crustacea: Decapoda) and its role in the population renewal on the exploited bottoms of the Mediterranean. Fish. Res. 2009, 95, 236–245. [Google Scholar] [CrossRef]
- Mannini, A.; Sabatella, R.F. Annuario sullo stato delle risorse e sulle strutture produttive dei mari italiani. Biol. Mar. Mediterr. 2015, 22, 358. [Google Scholar]
- Badyaev, A.V. Growing apart: An ontogenetic perspective on the evolution of sexual size dimorphism. Trends Ecol. Evol. 2002, 17, 369–378. [Google Scholar] [CrossRef]
- Castillo Geniz, J.L.; Nishizaki, O.S.; Perez Jiménez, J.C. Morphological variation and sexual dimorphism in the California skate, Raja inornata Jordan and Gilbert, 1881 from the Gulf of California, Mexico. Zootaxa 2007, 1545, 1–16. [Google Scholar] [CrossRef]
- Martinez, C.M.; Kao, B.H.; Sparks, J.S.; Wainwright, P.C. Pectoral Dimorphism Is a Pervasive Feature of Skate Diversity and Offers Insight into their Evolution. Integr. Org. Biol. 2019, 1, obz012. [Google Scholar] [CrossRef] [Green Version]
- Bolnick, D.I.; Doebeli, M. Sexual dimorphism and adaptive speciation: Two sides of the same ecological coin. Evolution 2003, 57, 2433–2449. [Google Scholar] [CrossRef]
- Orlov, A.M.; Cotton, C.F. Sexually dimorphic morphological characters in five north Atlantic deepwater skates (Chondrichthyes: Rajiformes). J. Mar. Sci. 2011, 2011, 842821. Available online: https://www.hindawi.com/journals/jmb/2011/842821/ (accessed on 2 October 2021). [CrossRef] [Green Version]
- Frisk, M.G. Life history strategies of batoids. In Sharks and Their Relatives II: Biodiversity, Adaptive Physiology and Conservation; Carrier, J.C., Musick, J.A., Heithaus, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2010; pp. 283–316. [Google Scholar]
- Sosebee, K.A. Maturity of Skates in Northeast United States Waters. J. Northwest Atl. Fish. Sci. 2005, 35, 141–153. [Google Scholar] [CrossRef]
- Poulet, N.; Berrebi, P.; Crivelli, A.J.; Lek, S.; Argillier, C. Genetic and morphometric variations in the pikeperch (Sander lucioperca L.) of a fragmented delta. Arch. Hydrobiol. 2004, 159, 531–554. [Google Scholar] [CrossRef]
- Hendry, A.P.; Kelly, M.L.; Kinnison, M.T.; Reznick, D.N. Parallel evolution of the sexes? Effects of predation and habitat features on the size and shape of wild guppies. J. Evol. Biol. 2006, 19, 741–754. [Google Scholar] [CrossRef]
- Bhagat, Y.; Fox, M.G.; Ferreira, M.T. Morphological differentiation in introduced pumpkinseed Lepomis gibbosus (L.) occupying different habitat zones in Portuguese reservoirs. J. Fish. Biol. 2006, 69, 79–94. [Google Scholar] [CrossRef]
- Ihssen, P.E.; Booke, H.E.; Casselman, J.M.; Mc-Glade, J.M.; Payne, N.R.; Utter, E.M. Stock identification: Materials and methods. Can. J. Fish. Aquat. Sci. 1981, 38, 1838–1855. [Google Scholar] [CrossRef]
- Abaunza, P.; Murta, A.G.; Campbell, N.; Cimmaruta, R.; Comesaña, A.S.; Dahle, G.; Gallo, E.; García Santamaría, M.T.; Gordo, L.S.; Iversen, S.A.; et al. Considerations on sampling strategies for an holistic approach to stock identification: The example of the HOMSIR project. Fish. Res. 2008, 89, 104–113. [Google Scholar] [CrossRef]
- Sharker, M.R.; Siddik, M.A.B.; Nahar, A.; Shahjahan, M.; Faroque, A.A. Genetic differentiation of wild and hatchery populations of Indian major carp Cirrhinus cirrhosis in Bangladesh. J. Environ. Biol. 2015, 36, 1223–1227. [Google Scholar]
- Misawa, R.; Narimatsu, Y.; Endo, H.; Kai, Y. Population structure of the ocellate spot skate (Okamejei kenojei) inferred from variations in mitochondrial DNA (mtDNA) sequences and from morphological characters of regional populations. Fish. Bull. NOAA 2018, 117, 24–36. [Google Scholar] [CrossRef]
- Costa, L.J.; De Almeida, P.R.; Costa, M.J. A morphometric and meristic investigation of Lusitanian toadfish Halobatrachus didactylus (Bloch and Schneider 1801): Evidence of population fragmentation on Portuguese coast. Sci. Mar. 2003, 67, 219–231. [Google Scholar] [CrossRef] [Green Version]
- Murta, A.G. Morphological variation of horse mackerel (Trachurus trachurus) in the Iberian and North African Atlantic: Implications for stock identifications. ICES J. Mar. Sci. 2000, 57, 1240–1248. [Google Scholar] [CrossRef]
- Turan, C. Stock identification of Mediterranean horse mackerel (Trachurus trachurus) using morphometric and meristic characters. ICES J. Mar. Sci. 2004, 61, 774–781. [Google Scholar] [CrossRef] [Green Version]
- Wimberger, P.H. Trophic polymorphisms, plasticity, and speciation in vertebrates. In Advances in Fish Foraging Theory and Ecology; Stouder, D.J., Fresh, K., Eds.; Belle Baruch Press: Columbia, SC, USA, 1994; pp. 19–43. [Google Scholar]
- Carbonara, P.; Bellodi, A.; Palmisano, M.; Mulas, A.; Porcu, C.; Zupa, W.; Donnaloia, M.; Carlucci, R.; Sion, L.; Follesa, M.C. Growth and Age Validation of the Thornback Ray (Raja clavata Linnaeus, 1758) in the South Adriatic Sea (Central Mediterranean). Front. Mar. Sci. 2020, 7, 1100. [Google Scholar] [CrossRef]
- Farrugia, T.J.; Goldman, K.J.; Tribuzio, C.; Seitz, A.C. First use of satellite tags to examine movement and habitat use of big skates Beringraja binoculata in the Gulf of Alaska. Mar. Ecol. Prog. Ser. 2016, 556, 209–221. [Google Scholar] [CrossRef]
- Spreitzer, M.L.; Mautner, S.; Makasa, L.; Sturmbauer, C. Genetic and morphological population differentiation in the rock-dwelling and specialized shrimp-feeding cichlid fish species Altolamprologus compressiceps from Lake Tanganyika, East Africa. Hydrobiologia 2012, 682, 143–154. [Google Scholar] [CrossRef] [Green Version]
- Koester, D.M. Anatomy and motor pathways of the electric organ of skates. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2003, 273, 648–662. [Google Scholar] [CrossRef]
- Capapé, C.; Ali, M.; Saad, A.; Alkusairy, H.; Reynaud, C. Atypical characteristics in the longnosed skate Dipturus oxyrinchus (Linnaeus, 1758) from the coast of Syria (Eastern Mediterranean). Thalassia Salent. 2015, 37, 71–80. [Google Scholar]
- Burns, J.G.; Di Nardo, P.; Rood, F.H. The role of predation in variation in body shape in guppies Poecilia reticulata: A comparison of field and common garden phenotypes. J. Fish. Biol. 2009, 75, 1144–1157. [Google Scholar] [CrossRef]
- Webster, M.M.; Atton, N.; Hart, P.J.B.; Ward, A.J.W. Habitat-specific morphological variation among threespine sticklebacks (Gasterosteus aculeatus) within a drainage basin. PLoS ONE 2011, 6, e21060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzarotto, H.; Barros, T.; Louvise, J.; Pellegrini Caramaschi, E. Morphological variation among populations of Hemigrammus coeruleus (Characiformes: Characidae) in a Negro River tributary, Brazilian Amazon. Neotrop. Ichthyol. 2017, 15, e160152. [Google Scholar] [CrossRef] [Green Version]
- van Rijssel, J.C.; Witte, F. Adaptive responses in resurgent Lake Victoria cichlids over the past 30 years. Evol. Ecol. 2013, 27, 253–267. [Google Scholar] [CrossRef]
- Araújo, M.S.; Perez, S.I.; Magazoni, M.J.C.; Petry, A.C. Body size and allometric shape variation in the molly Poecilia vivipara along a gradient of salinity and predation. BMC Evol. Biol. 2014, 15, 1–11. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Equation | a | b | R2 | Equation | a | b | R2 |
|---|---|---|---|---|---|---|---|
| TW = a + TLb (Sardinia) | 0.0009 | 3.387 | 0.970 | ID = a + b TL (Sardinia) | 0.032 | 0.029 | 0.870 |
| TW = a + TLb (Ionian-Adriatic) | 0.0007 | 3.355 | 0.996 | ID = a + b TL (Ionian-Adriatic) | 0.038 | 0.311 | 0.948 |
| TW = a + TLb (Combined) | 0.0007 | 3.388 | 0.975 | ID = a + b TL (Combined) | 0.033 | 0.270 | 0.817 |
| DW = a + b TL (Sardinia) | 0.719 | −0.766 | 0.991 | C2CT = a + b TL (Sardinia) | 0.400 | 3.525 | 0.976 |
| DW = a + b TL (Ionian-Adriatic) | 0.709 | −0.295 | 0.995 | C2CT = a + b TL (Ionian-Adriatic) | 0.405 | 3.185 | 0.967 |
| DW = a + b TL (Combined) | 0.715 | −0.601 | 0.993 | C2CT = a + b TL (Combined) | 0.402 | 3.378 | 0.972 |
| DL = a + b TL (Sardinia) | 0.611 | −1.495 | 0.986 | SLPJ = a + b TL (Sardinia) | 0.195 | 0.196 | 0.960 |
| DL = a + b TL (Ionian-Adriatic) | 0.596 | −1.349 | 0.984 | SLPJ = a + b TL (Ionian-Adriatic) | 0.190 | 0.652 | 0.956 |
| DL = a + b TL (Combined) | 0.606 | −1.577 | 0.985 | SLPJ = a + b TL (Combined) | 0.193 | 0.415 | 0.959 |
| SMW = a + b TL (Sardinia) | 0.363 | −0.731 | 0.981 | PL = a + b TL (Sardinia) | 0.179 | 0.132 | 0.960 |
| SMW = a + b TL (Ionian-Adriatic) | 0.311 | 1.499 | 0.964 | PL = a + b TL (Ionian-Adriatic) | 0.179 | 0.215 | 0.949 |
| SMW = a + b TL (Combined) | 0.342 | 0.027 | 0.960 | PL = a + b TL (Combined) | 0.179 | 0.181 | 0.956 |
| SLPOR = a + b TL (Sardinia) | 0.200 | −0.282 | 0.965 | MW = a + b TL (Sardinia) | 0.089 | −0.179 | 0.957 |
| SLPOR = a + b TL (Ionian-Adriatic) | 0.194 | 0.449 | 0.980 | MW = a + b TL (Ionian-Adriatic) | 0.089 | −0.074 | 0.953 |
| SLPOR = a + b TL (Combined) | 0.197 | 0.081 | 0.972 | MW = a + b TL (Combined) | 0.089 | −0.115 | 0.956 |
| EBL = a + b TL (Sardinia) | 0.022 | 0.161 | 0.801 | DBN = a + b TL (Sardinia) | 0.093 | 0.247 | 0.960 |
| EBL = a + b TL (Ionian-Adriatic) | 0.020 | 0.125 | 0.936 | DBN = a + b TL (Ionian-Adriatic) | 0.095 | −0.721 | 0.964 |
| EBL = a + b TL (Combined) | 0.022 | 0.112 | 0.839 | DBN = a + b TL (Combined) | 0.095 | −0.308 | 0.955 |
| OD = a + b TL (Sardinia) | 0.029 | 0.138 | 0.879 | GOW1 = a + b TL (Sardinia) | 0.017 | −0.043 | 0.944 |
| OD = a + b TL (Ionian-Adriatic) | 0.031 | 0.109 | 0.946 | GOW1 = a + b TL (Ionian-Adriatic) | 0.016 | −0.044 | 0.902 |
| OD = a + b TL (Combined) | 0.030 | 0.134 | 0.911 | GOW1 = a + b TL (Combined) | 0.017 | −0.054 | 0.924 |
| OSL = a + b TL (Sardinia) | 0.043 | 0.184 | 0.951 | GOW2 = a + b TL (Sardinia) | 0.015 | −0.114 | 0.949 |
| OSL = a + b TL (Ionian-Adriatic) | 0.044 | 0.149 | 0.953 | GOW2 = a + b TL (Ionian-Adriatic) | 0.012 | −0.028 | 0.929 |
| OSL = a + b TL (Combined) | 0.043 | 0.183 | 0.951 | GOW2 = a + b TL (Combined) | 0.014 | −0.088 | 0.925 |
| SL = a + b TL (Sardinia) | 0.017 | −0.095 | 0.874 | DB1GS = a + b TL (Sardinia) | 0.162 | −0.680 | 0.978 |
| SL = a + b TL (Ionian-Adriatic) | 0.022 | −0.136 | 0.923 | DB1GS = a + b TL (Ionian-Adriatic) | 0.158 | −0.803 | 0.957 |
| SL = a + b TL (Combined) | 0.019 | −0.067 | 0.832 | DB1GS = a + b TL (Combined) | 0.161 | −0.800 | 0.968 |
| DBOR = a + b TL (Sardinia) | 0.061 | −0.660 | 0.970 | DB5GS = a + b TL (Sardinia) | 0.113 | −0.723 | 0.972 |
| DBOR = a+ b TL (Ionian-Adriatic) | 0.051 | −0.209 | 0.973 | DB5GS = a + b TL (Ionian-Adriatic) | 0.114 | −1.296 | 0.950 |
| DBOR = a + b TL (Combined) | 0.057 | −0.496 | 0.956 | DB5GS = a + b TL (Combined) | 0.114 | −1.058 | 0.960 |
| DBS = a + b TL (Sardinia) | 0.075 | −0.369 | 0.986 | D1BL = a + b TL (Sardinia) | 0.044 | 0.359 | 0.804 |
| DBS = a + b TL (Ionian-Adriatic) | 0.071 | −0.391 | 0.951 | D1BL = a + b TL (Ionian-Adriatic) | 0.034 | 0.073 | 0.889 |
| DBS = a + b TL (Combined) | 0.074 | −0.434 | 0.965 | D1BL = a + b TL (Combined) | 0.041 | 0.071 | 0.736 |
| DSC = a + b TL (Sardinia) | 0.556 | −1.671 | 0.981 | D1H = a + b TL (Sardinia) | 0.034 | −0.512 | 0.852 |
| DSC = a + b TL (Ionian-Adriatic) | 0.562 | −2.190 | 0.955 | D1H = a + b TL (Ionian-Adriatic) | 0.030 | 0.025 | 0.933 |
| DSC = a + b TL (Combined) | 0.558 | −1.915 | 0.970 | D1H = a + b TL (Combined) | 0.032 | −0.235 | 0.879 |
| C2DF1 = a + b TL (Sardinia) | 0.248 | 1.449 | 0.972 | D1 = a + b TL (Sardinia) | 0.167 | 1.567 | 0.942 |
| C2DF1 = a + b TL (Ionian-Adriatic) | 0.252 | −0.292 | 0.971 | D1 = a + b TL (Ionian-Adriatic) | 0.174 | 1.772 | 0.938 |
| C2DF1 = a + b TL (Combined) | 0.252 | 0.441 | 0.968 | D1 = a + b TL (Combined) | 0.169 | 1.778 | 0.936 |
| C2DF2 = a + b TL (Sardinia) | 0.301 | 2.974 | 0.984 | D2 = a + b TL (Sardinia) | 0.102 | 0.979 | 0.917 |
| C2DF2 = a + b TL (Ionian-Adriatic) | 0.323 | 0.414 | 0.981 | D2 = a + b TL (Ionian-Adriatic) | 0.102 | 1.463 | 0.921 |
| C2DF2 = a + b TL (Combined) | 0.312 | 1.678 | 0.982 | D2 = a + b TL (Combined) | 0.101 | 1.271 | 0.917 |
| Actual | |||
| Ionian-Adriatic | Sardinia | ||
| Predicted | Ionian-Adriatic | 27 | 3 |
| Sardinia | 1 | 27 | |
| Actual | |||
| Ionian-Adriatic | Sardinia | ||
| Predicted | Ionian-Adriatic | 25 | 3 |
| Sardinia | 3 | 27 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbonara, P.; Bellodi, A.; Zupa, W.; Donnaloia, M.; Gaudio, P.; Neglia, C.; Follesa, M.C. Morphological Traits and Capture Depth of the Norwegian Skate (Dipturus nidarosiensis (Storm, 1881)) from Two Mediterranean Populations. J. Mar. Sci. Eng. 2021, 9, 1462. https://doi.org/10.3390/jmse9121462
Carbonara P, Bellodi A, Zupa W, Donnaloia M, Gaudio P, Neglia C, Follesa MC. Morphological Traits and Capture Depth of the Norwegian Skate (Dipturus nidarosiensis (Storm, 1881)) from Two Mediterranean Populations. Journal of Marine Science and Engineering. 2021; 9(12):1462. https://doi.org/10.3390/jmse9121462
Chicago/Turabian StyleCarbonara, Pierluigi, Andrea Bellodi, Walter Zupa, Marilena Donnaloia, Palma Gaudio, Cosmidano Neglia, and Maria Cristina Follesa. 2021. "Morphological Traits and Capture Depth of the Norwegian Skate (Dipturus nidarosiensis (Storm, 1881)) from Two Mediterranean Populations" Journal of Marine Science and Engineering 9, no. 12: 1462. https://doi.org/10.3390/jmse9121462