Integrating Three-Dimensional Benthic Habitat Characterization Techniques into Ecological Monitoring of Coral Reefs


Abstract
:1. Introduction
2. Materials and Methods
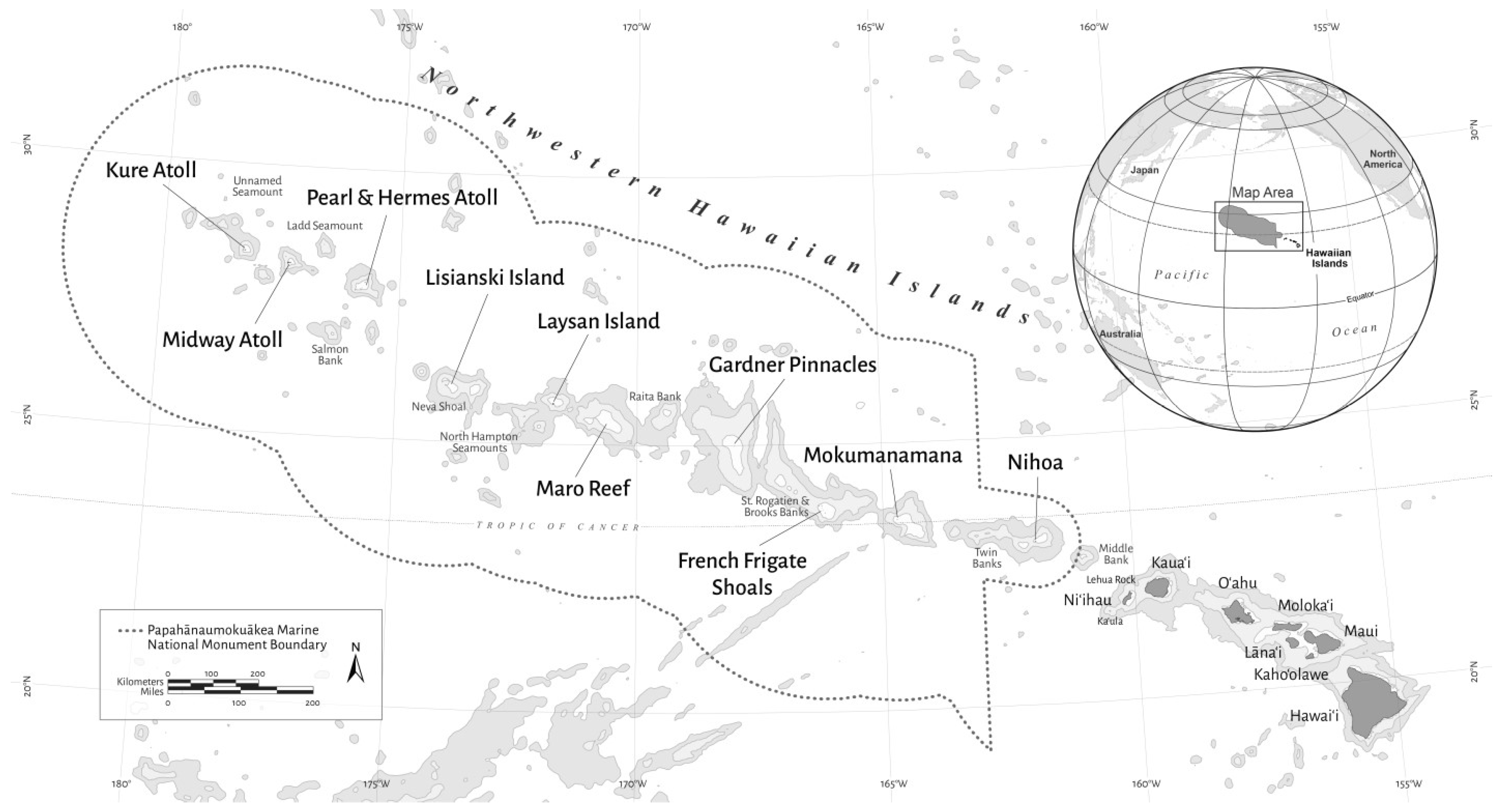
2.1. Image Acquisition
2.2. Generation of 3D Models
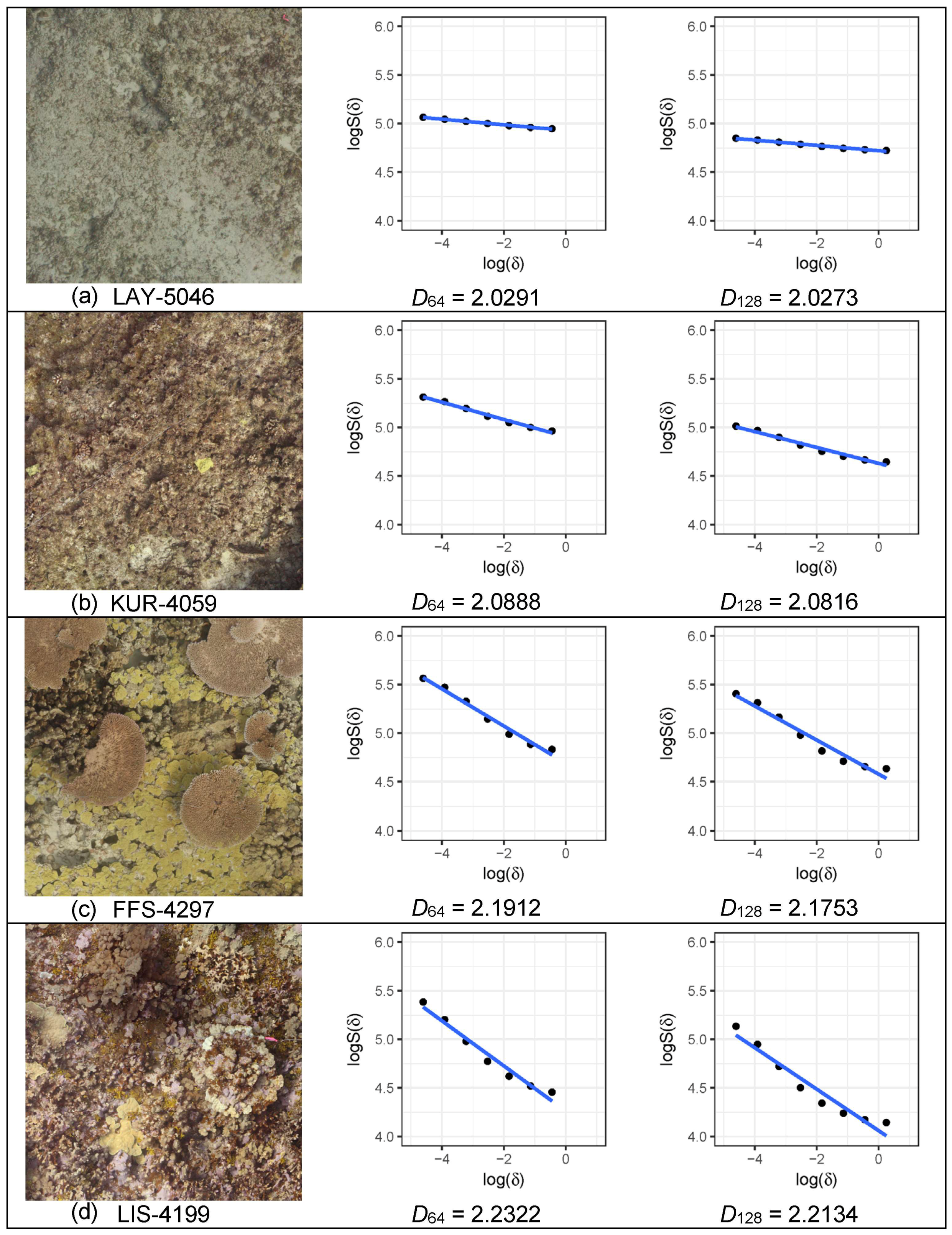
2.3. Quantification of the Habitat Structure
2.3.1. ArcMap Procedure
2.3.2. R Procedure
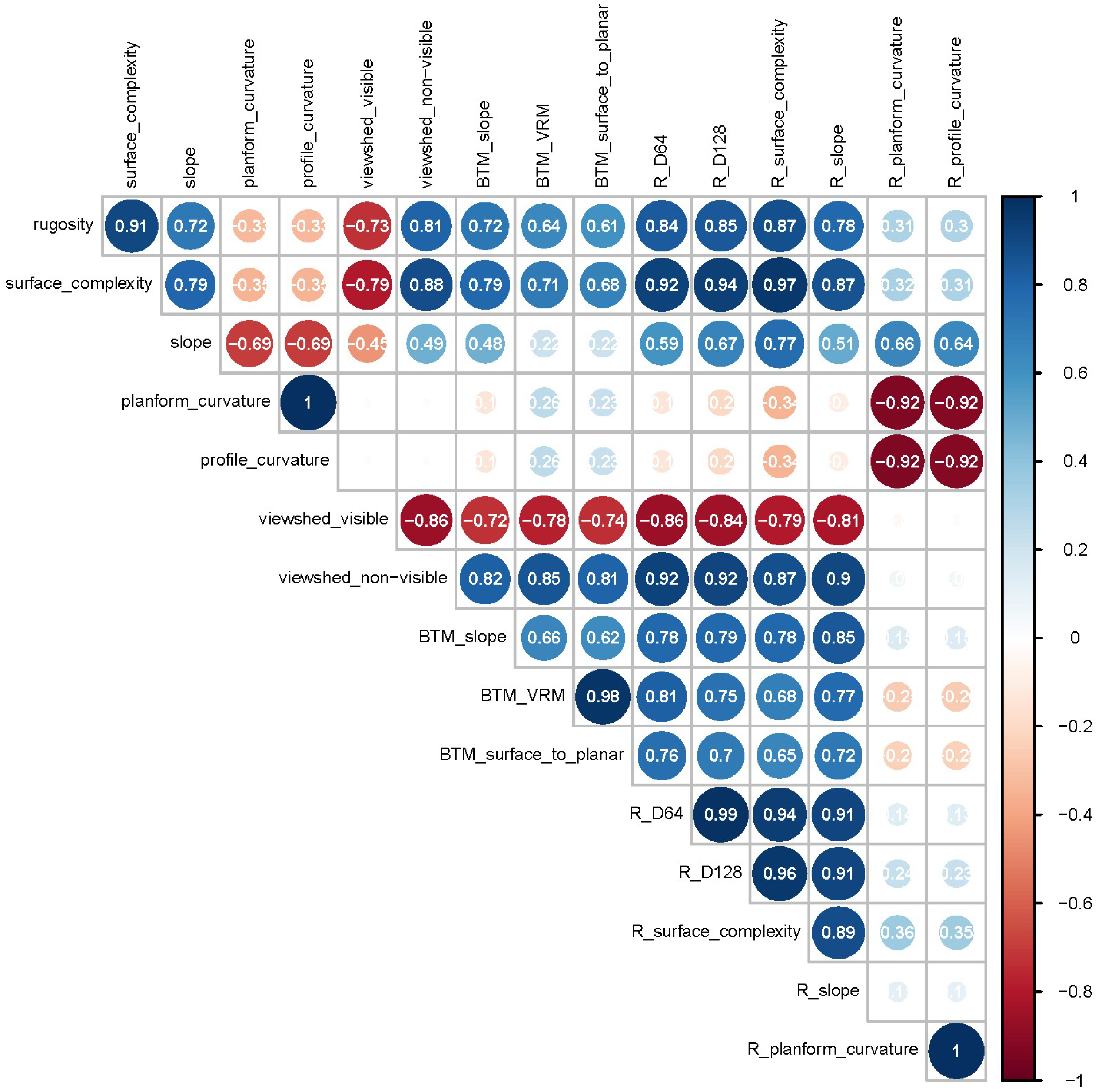
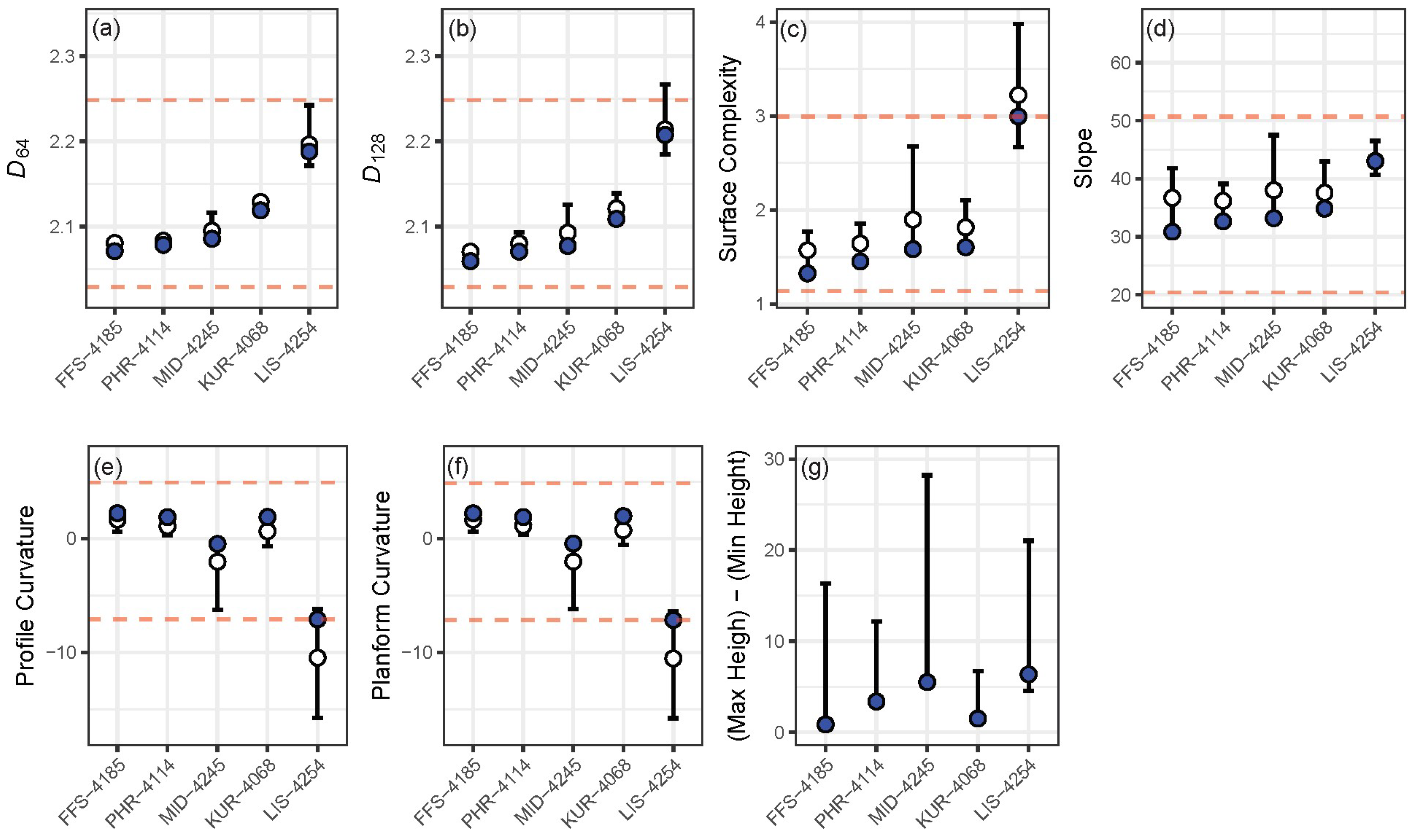
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- McGehee, M.A. Correspondence between assemblages of coral reef fishes and gradients of water motion, depth, and substrate size off Puerto Rico. Mar. Ecol. Prog. Ser. 1994, 105, 243–255. [Google Scholar] [CrossRef]
- Floeter, S.R.; Krohling, W.; Gasparini, J.L.; Ferreira, C.E.L.; Zalmon, I.R. Reef fish community structure on coastal islands of the southeastern Brazil: The influence of exposure and benthic cover. Environ. Biol. Fish. 2007, 78, 147–160. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Parrish, J.D. Habitat characteristics affecting fish assemblages on a Hawaiian coral reef. J. Exp. Mar. Biol. Ecol. 1998, 224, 1–30. [Google Scholar] [CrossRef]
- Jones, G.P.; Syms, C. Disturbance, habitat structure and the ecology of fishes on coral reefs. Austral Ecol. 1998, 23, 287–297. [Google Scholar] [CrossRef]
- Gratwicke, B.; Speight, M.R. The relationship between fish species richness, abundance and habitat complexity in a range of shallow tropical marine habitats. J. Fish Biol. 2005, 66, 650–667. [Google Scholar] [CrossRef]
- Pittman, S.J.; Costa, B.M.; Battista, T.A. Using lidar bathymetry and boosted regression trees to predict the diversity and abundance of fish and corals. J. Coast. Res. 2009, 25, 27–38. [Google Scholar] [CrossRef]
- Komyakova, V.; Munday, P.L.; Jones, G.P. Relative importance of coral cover, habitat complexity and diversity in determining the structure of reef fish communities. PLoS ONE 2013, 8, e83178. [Google Scholar] [CrossRef]
- Connell, J.H. Disturbance and recovery of coral assemblages. Coral Reefs 1997, 16, S101–S113. [Google Scholar] [CrossRef]
- Loya, Y.; Sakai, K.; Yamazato, K.; Nakano, Y.; Sambali, H.; van Woesik, R. Coral bleaching: The winners and the losers. Ecol. Lett. 2001, 4, 122–131. [Google Scholar] [CrossRef]
- Couch, C.S.; Burns, J.H.R.; Liu, G.; Steward, K.; Gutlay, T.N.; Kenyon, J.; Eakin, C.M.; Kosaki, R.K. Mass coral bleaching due to unprecedented marine heatwave in Papahānaumokuākea Marine National Monument (Northwestern Hawaiian Islands). PLoS ONE 2017, 12, e0185121. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.L.; Shima, J.S.; Phillips, N.E. Effects of microhabitat availability on estimates of density of a reef fish: Implications for assessments of marine protected areas. Hydrobiologia 2012, 685, 173–190. [Google Scholar] [CrossRef]
- Burns, J.H.R.; Delparte, D.; Gates, R.D.; Takabayashi, M. Integrating structure-from-motion photogrammetry with geospatial software as a novel technique for quantifying 3D ecological characteristics of coral reefs. PeerJ 2015, 3, e1077. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.H.R.; Delparte, D.; Kapono, L.; Belt, M.; Gates, R.D.; Takabayashi, M. Assessing the impact of acute disturbances on the structure and composition of a coral community using innovative 3D reconstruction techniques. Methods Oceanogr. 2016, 15–16, 49–59. [Google Scholar] [CrossRef]
- Young, G.C.; Dey, S.; Rogers, A.D.; Exton, D. Cost and time-effective method for multi-scale measures of rugosity, fractal dimension, and vector dispersion from coral reef 3D models. PLoS ONE 2017, 12, e0175341. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.X.; Roelfsema, C.M.; Saunders, M.I.; Phinn, S.R. Measuring coral reef terrain roughness using ‘structure-from-motion’ close-range photogrammetry. Geomorphology 2015, 242, 21–28. [Google Scholar] [CrossRef]
- Agudo-Adriani, E.A.; Cappelletto, J.; Cavada-Blanco, F.; Croquer, A. Colony geometry and structural complexity of the endangered species Acropora cervicornis partly explains the structure of their associated fish assemblage. PeerJ 2016, 4, e1861. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.; Bryson, M.; Bridge, T.; Hustache, J.; Williams, S.B.; Byrne, M.; Figueira, W. Quantifying the response of structural complexity and community composition to environmental change in marine communities. Glob. Chang. Biol. 2016, 22, 1965–1975. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, R.; Figueira, W.F.; Pratchett, M.S.; Boube, T.; Adam, A.; Kobelkowsky-Vidrio, T.; Doo, S.S.; Atwood, T.B.; Byrne, M. 3D photogrammetry quantifies growth and external erosion of individual coral colonies and skeletons. Sci. Rep. 2017, 7, 16737. [Google Scholar] [CrossRef] [Green Version]
- Figueira, W.; Ferrari, R.; Weatherby, E.; Porter, A.; Hawes, S.; Byrne, M. Accuracy and precision of habitat structural complexity metrics derived from underwater photogrammetry. Remote Sens. 2015, 7, 16883–16900. [Google Scholar] [CrossRef]
- Raoult, V.; Reid-Anderson, S.; Ferri, A.; Williamson, J.E. How reliable is structure from motion (SfM) over time and between observers? A case study using coral reef bommies. Remote Sens. 2017, 9, 740. [Google Scholar] [CrossRef]
- Raoult, V.; David, P.A.; Dupont, S.F.; Mathewson, C.P.; O’Neill, S.J.; Powell, N.N.; Williamson, J.E. GoPros™ as an underwater photogrammetry tool for citizen science. PeerJ 2016, 4, e1960. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, A.; Kosaki, R.K. Use of multivariate control charts to assess the status of reef fish assemblages in the Northwestern Hawaiian Islands. PeerJ 2017, 5, e3651. [Google Scholar] [CrossRef] [PubMed]
- Williams, I.D.; Baum, J.K.; Heenan, A.; Hanson, K.M.; Nadon, M.O.; Brainard, R.E. Human, oceanographic and habitat drivers of central and western Pacific coral reef fish assemblages. PLoS ONE 2015, 10, e0120516. [Google Scholar] [CrossRef]
- Walbridge, S.; Slocum, N.; Pobuda, M.; Wright, D.J. Unified geomorphological analysis workflows with Benthic Terrain Modeler. Geosciences 2018, 8, 94. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; 3.4.4; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Pebesma, E.J.; Bivand, R.S. Classes and methods for spatial data in R. R News 2005, 5, 9–13. [Google Scholar]
- Bivand, R.S.; Pebesma, E.; Gómez-Rubio, V. Applied Spatial Data Analysis with R, 2nd ed.; Springer: New York, NY, USA, 2013. [Google Scholar]
- Hijmans, R.J. raster: Geographic Data Analysis and Modeling, R package version 2.5-8; Geospatial and Farming Systems Research Consortium: Davis, CA, USA, 2016. [Google Scholar]
- Bivand, R.; Rundel, C. rgeos: Interface to Geometry Engine—Open Source (GEOS), R package version 0.3-23; Geospatial and Farming Systems Research Consortium: Davis, CA, USA, 2017. [Google Scholar]
- An Overview of the Surface Toolset. Available online: http://desktop.arcgis.com/en/arcmap/10.4/tools/spatial-analyst-toolbox/an-overview-of-the-surface-tools.htm (accessed on 8 November 2018).
- Moore, I.D.; Grayson, R.B.; Ladson, A.R. Digital terrain modelling: A review of hydrological, geomorphological, and biological applications. Hydrol. Process. 1991, 5, 3–30. [Google Scholar] [CrossRef]
- Zevenbergen, L.W.; Thorne, C.R. Quantitative analysis of land surface topography. Earth Surf. Process. Landf. 1987, 12, 47–56. [Google Scholar] [CrossRef]
- Basillais, É. Coral surfaces and fractal dimensions: A new method. Comptes Rendus Acad. Sci. Sci. De La Vie 1997, 320, 653–657. [Google Scholar] [CrossRef]
- Martin-Garin, B.; Lathuilière, B.; Verrecchia, E.P.; Geister, J. Use of fractal dimensions to quantify coral shape. Coral Reefs 2007, 26, 541–550. [Google Scholar] [CrossRef] [Green Version]
- Reichert, J.; Backes, A.R.; Schubert, P.; Wilke, T. The power of 3D fractal dimensions for comparative shape and structural complexity analyses of irregularly shaped organisms. Methods Ecol. Evol. 2017, 8, 1650–1658. [Google Scholar] [CrossRef]
- Shary, P.A.; Sharaya, L.S.; Mitusov, A.V. The problem of scale-specific and scale-free approaches in geomorphometry. Geogr. Fis. Dinam. Quat. 2005, 28, 81–101. [Google Scholar]
- Blaga, L. Aspects regarding the significance of the curvature types and values in the studies of geomorphometry assisted by GIS. Ann. Univ. Din Oradea Ser. Geogr. 2012, XXII, 327–337. [Google Scholar]
- Krebs, P.; Stocker, M.; Pezzatti, G.B.; Conedera, M. An alternative approach to transverse and profile terrain curvature. Int. J. Geogr. Inf. Sci. 2015, 29, 643–666. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Process | Settings |
|---|---|
| Align Photos | High accuracy, generic preselection enabled, 50,000 key point limit, 5000 tie point limit. |
| Optimize Camera Alignment | (Use all the ones selected by the software.) |
| Build Dense Cloud | Medium quality, mild depth filtering, reuse depth maps disabled. |
| Build Mesh | Arbitrary surface type, high face count, interpolation enabled, calculate vertex colors enabled. |
| Build Texture | Adaptive orthophoto mapping mode, mosaic blending mode, texture size/count 16,384, enable hole filling. |
| Habitat metric | Software/License | Variable name |
|---|---|---|
| linear rugosity | ArcMap, Functional Surface/3D Analyst | rugosity |
| surface complexity | ArcMap, Functional Surface/3D Analyst | surface complexity |
| R | R surface complexity | |
| slope | ArcMap, Functional Surface/3D Analyst | slope |
| ArcMap, BTM | BTM slope | |
| R | R slope | |
| curvature | ArcMap, Surface/Spatial Analyst R | planform curvature profile curvature R planform curvature R profile curvature |
| viewshed | ArcMap, Surface/Spatial Analyst | viewshed visible viewshed non-visible |
| terrain ruggedness | ArcMap, BTM | BTM VRM |
| surface area to planar area ratio | ArcMap, BTM | BTM surface to planar |
| Fractal dimension | R | R D64 (1 to 64 cm resolution) R D128 (1 to 128 cm resolution) |
| - | Mean | SD | Median | Min–Max |
|---|---|---|---|---|
| Original planar area (m2) | 142.34 | 39.13 | 143.27 | 57.53–284.23 |
| Planar area ×64 (m2) | 109.57 | 37.00 | 111.00 | 38.50–243.30 |
| Planar area ×128 (m2) | 80.76 | 34.64 | 81.92 | 14.75–208.08 |
| Area retained ×64 (%) | 75.61 | 6.31 | 77.3 | 53.7–85.6 |
| Area retained ×128 (%) | 54.39 | 10.71 | 57.0 | 19.2–73.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fukunaga, A.; Burns, J.H.R.; Craig, B.K.; Kosaki, R.K. Integrating Three-Dimensional Benthic Habitat Characterization Techniques into Ecological Monitoring of Coral Reefs. J. Mar. Sci. Eng. 2019, 7, 27. https://doi.org/10.3390/jmse7020027
Fukunaga A, Burns JHR, Craig BK, Kosaki RK. Integrating Three-Dimensional Benthic Habitat Characterization Techniques into Ecological Monitoring of Coral Reefs. Journal of Marine Science and Engineering. 2019; 7(2):27. https://doi.org/10.3390/jmse7020027
Chicago/Turabian StyleFukunaga, Atsuko, John H. R. Burns, Brianna K. Craig, and Randall K. Kosaki. 2019. "Integrating Three-Dimensional Benthic Habitat Characterization Techniques into Ecological Monitoring of Coral Reefs" Journal of Marine Science and Engineering 7, no. 2: 27. https://doi.org/10.3390/jmse7020027