
Characterizing the Variability of a Physical Driver of North Atlantic Right Whale Foraging Habitat Using Altimetric Indices

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
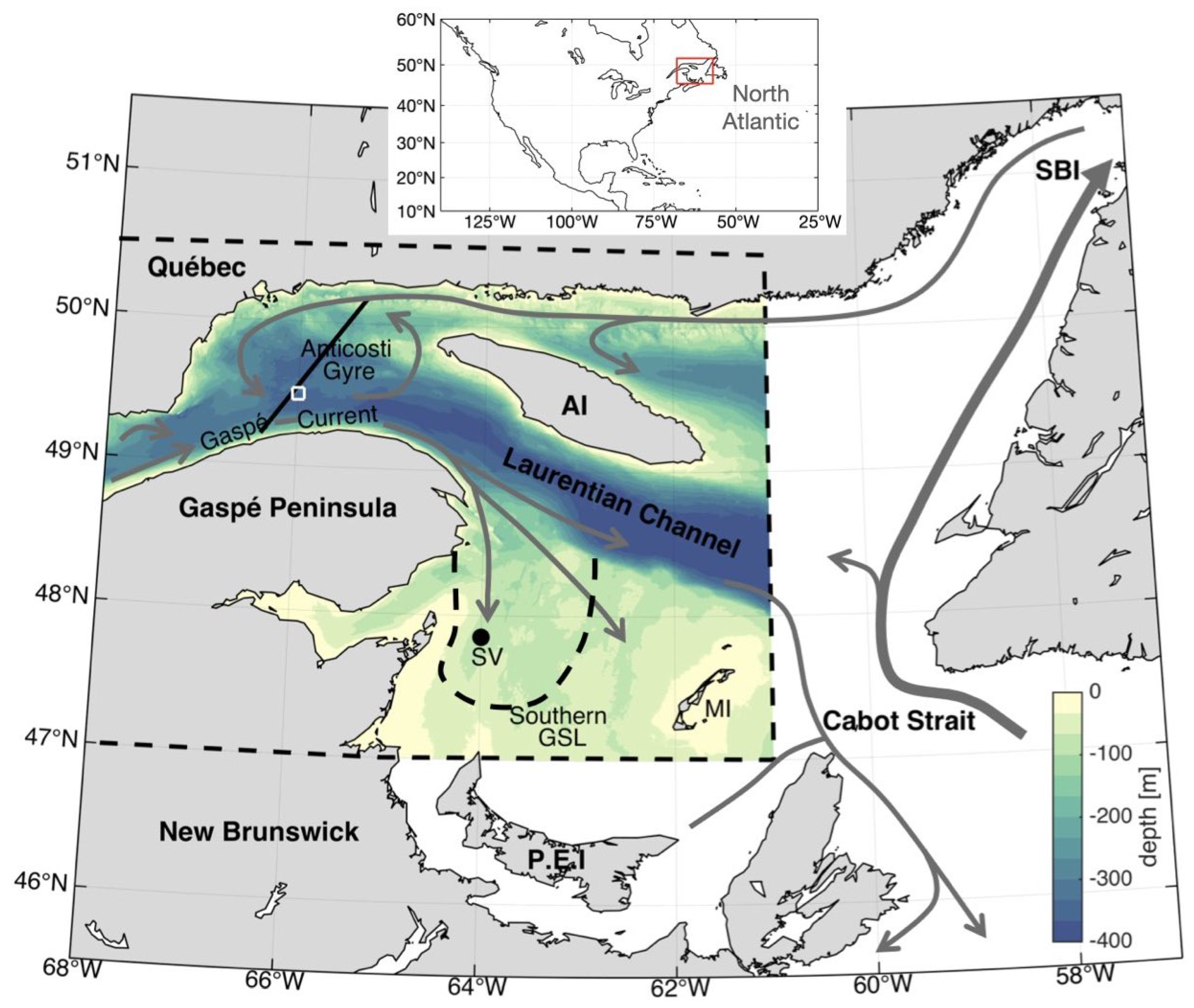
1.1. GSL Circulation
1.2. NARW in the sGSL
1.3. Satellite Altimetry to Monitor Physical Drivers of Prey Supply
2. Materials and Methods
2.1. Jason Altimetry SLA
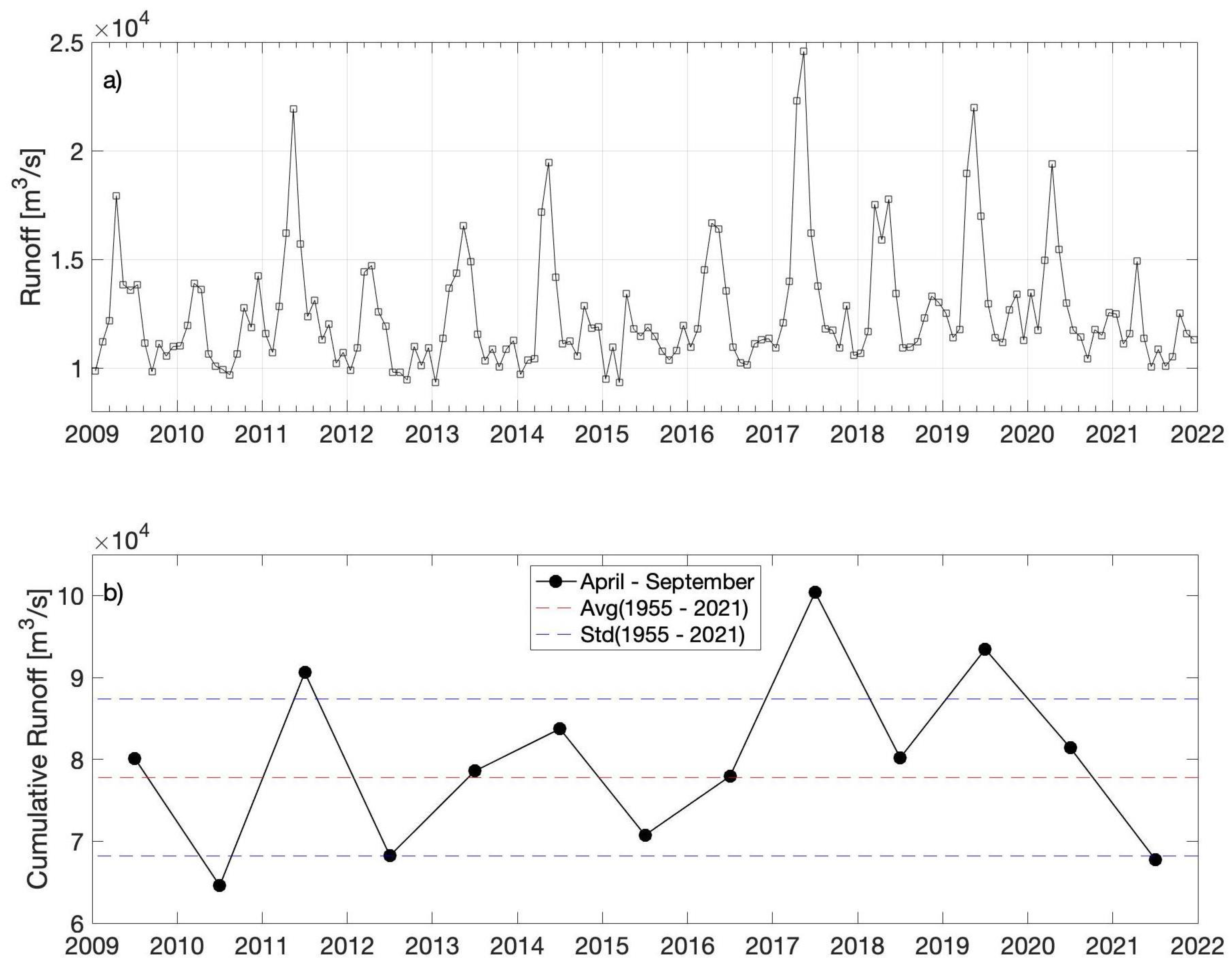
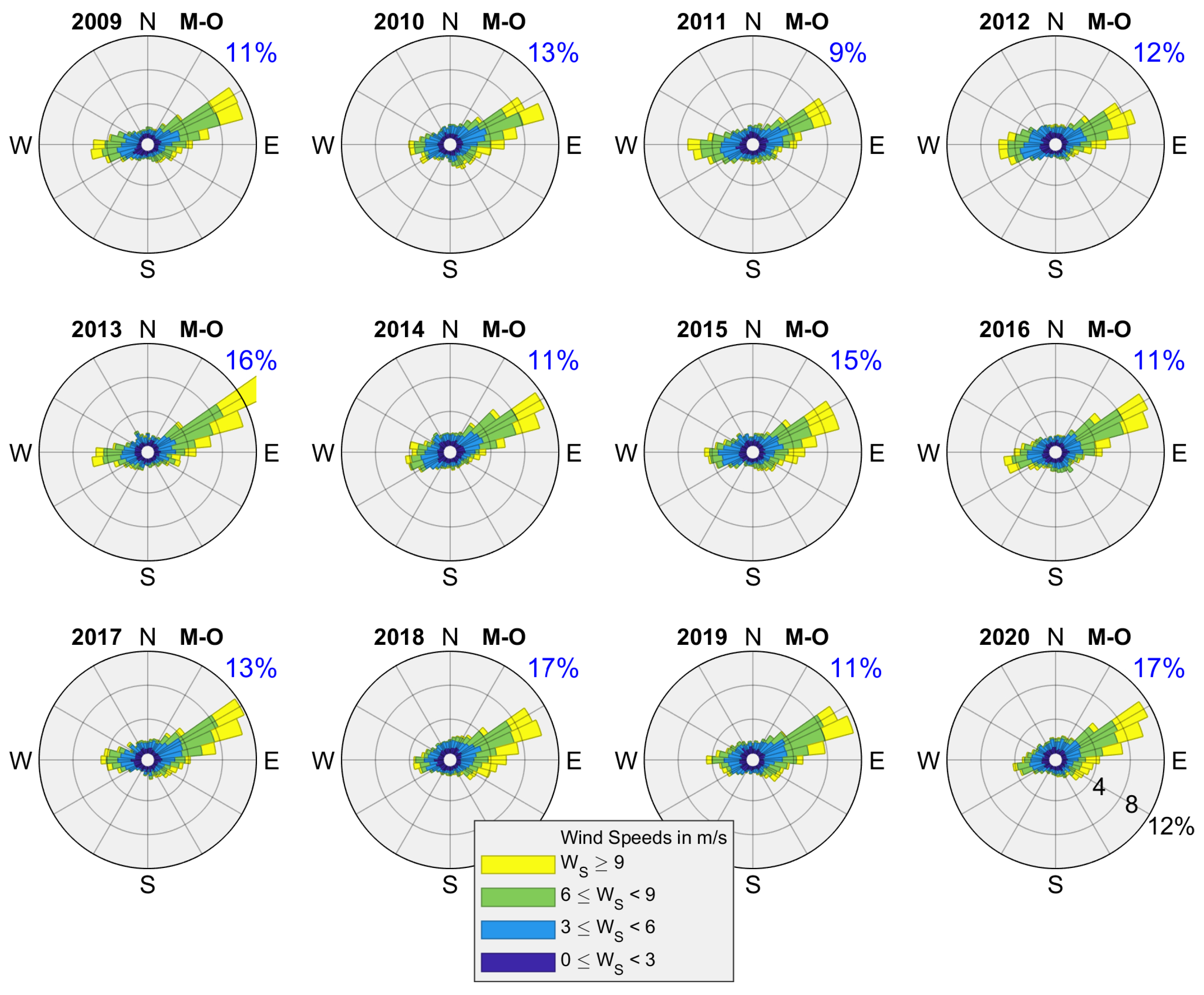
2.2. Freshwater Discharge and Wind Data
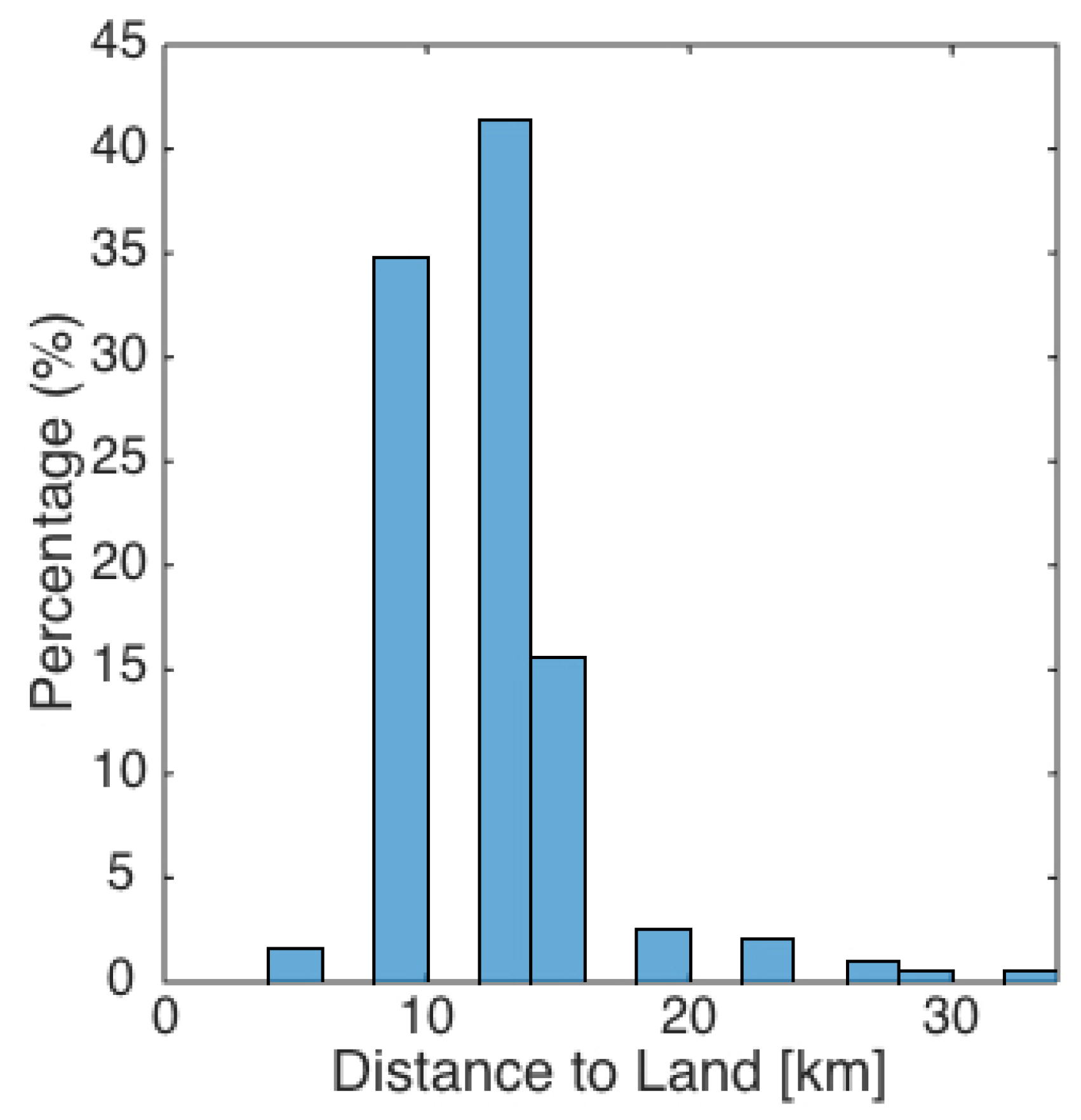
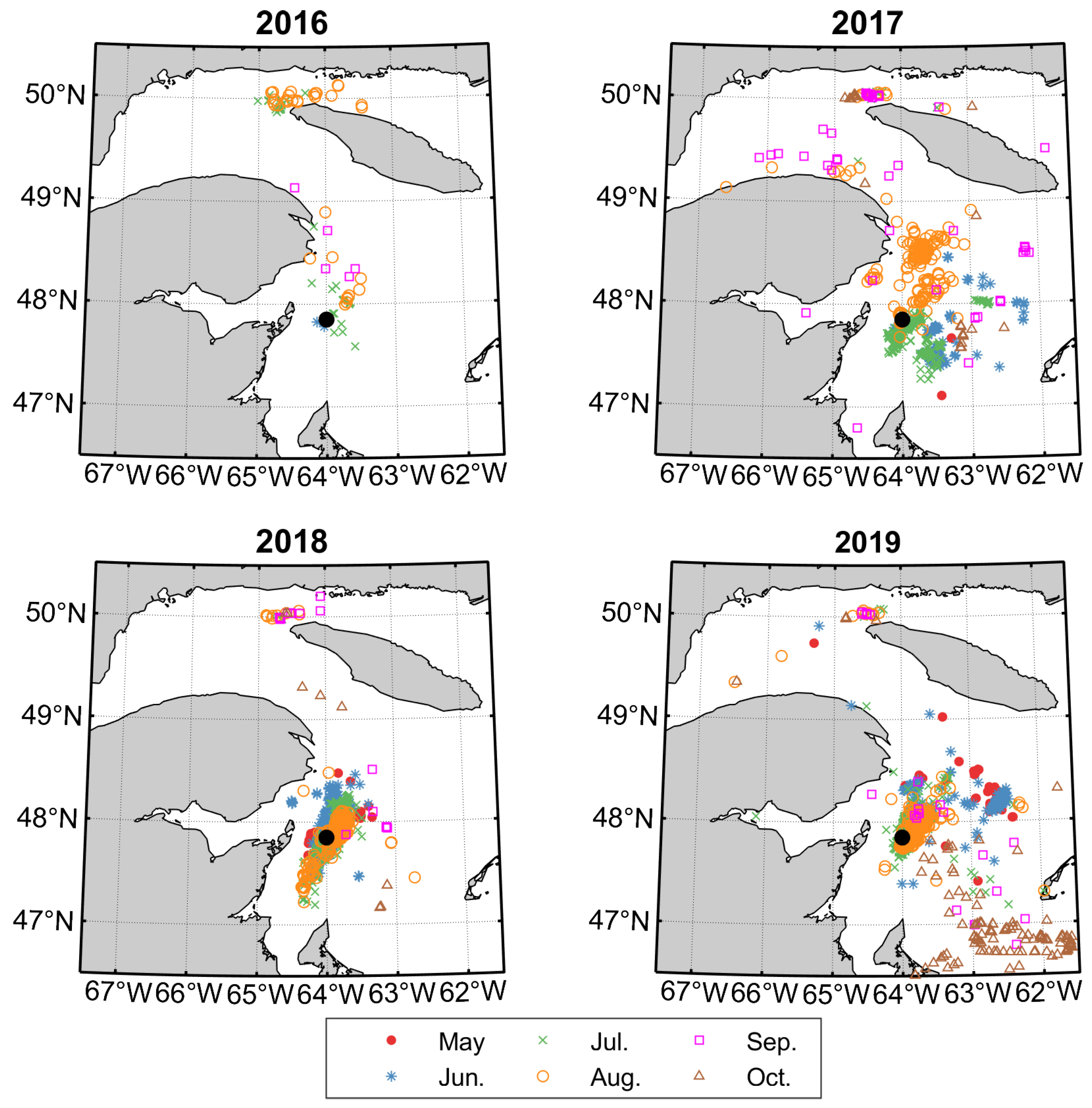
2.3. NARW Sightings and Zooplankton Data
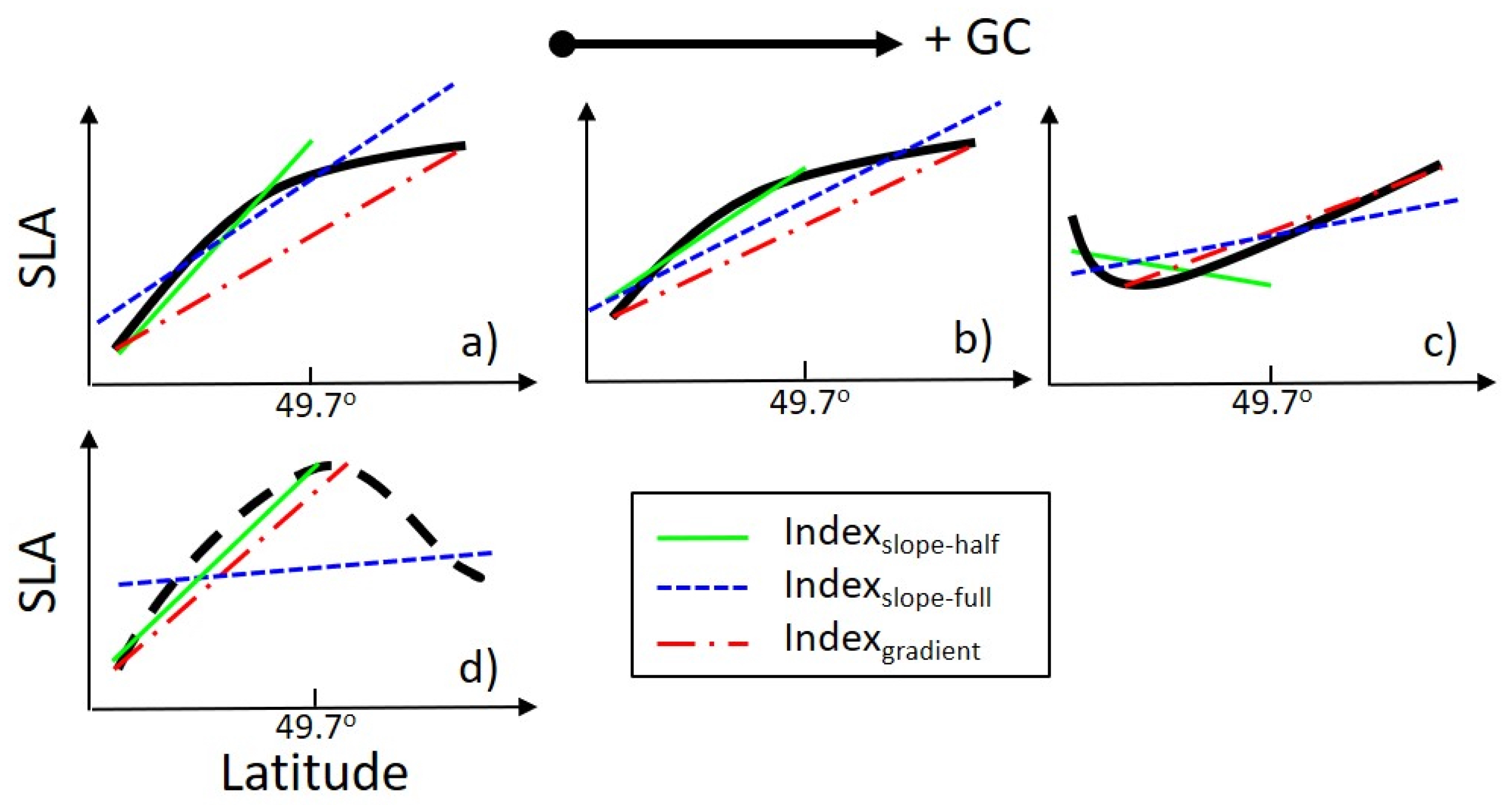
2.4. Altimetric Indices
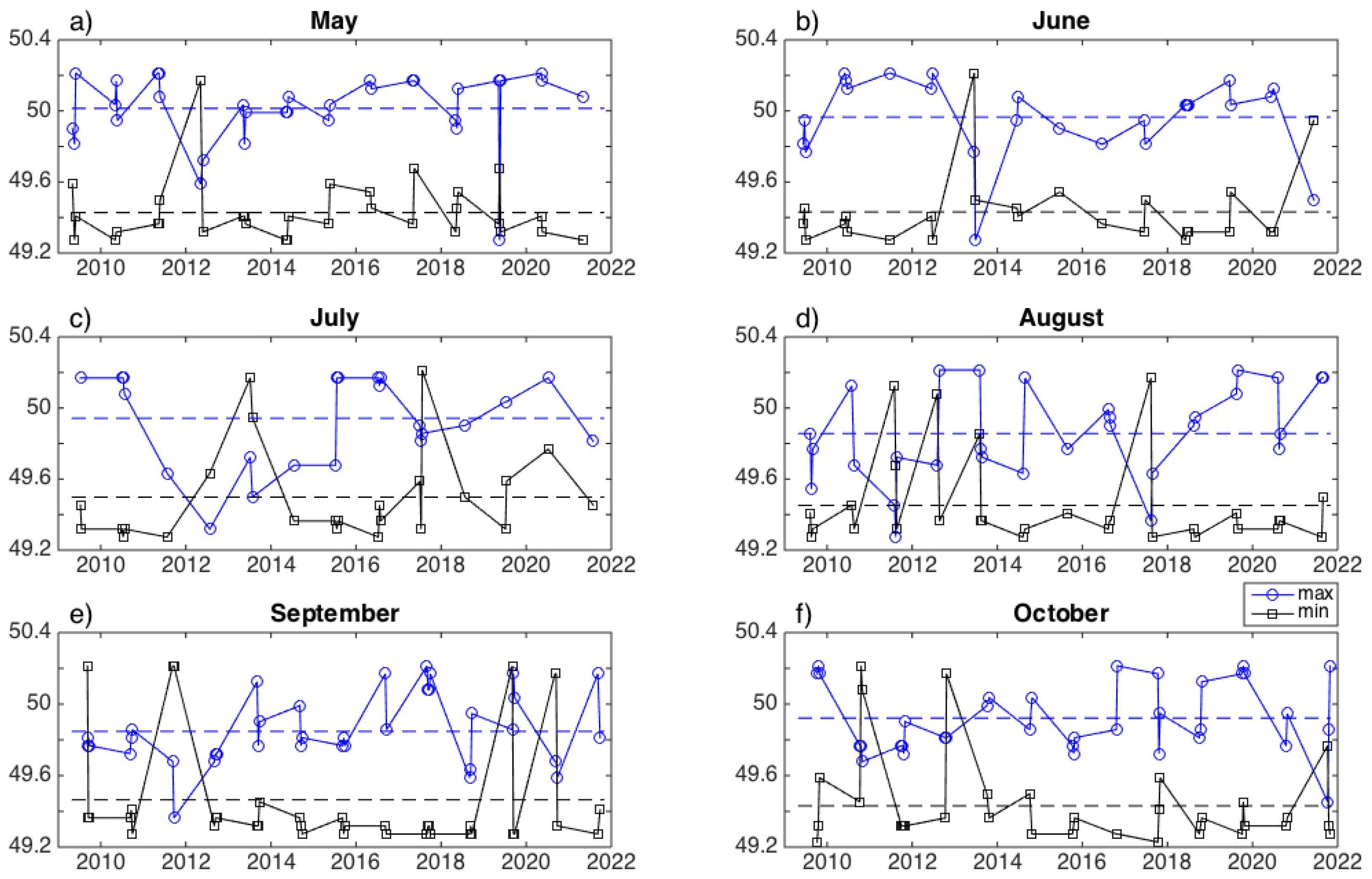
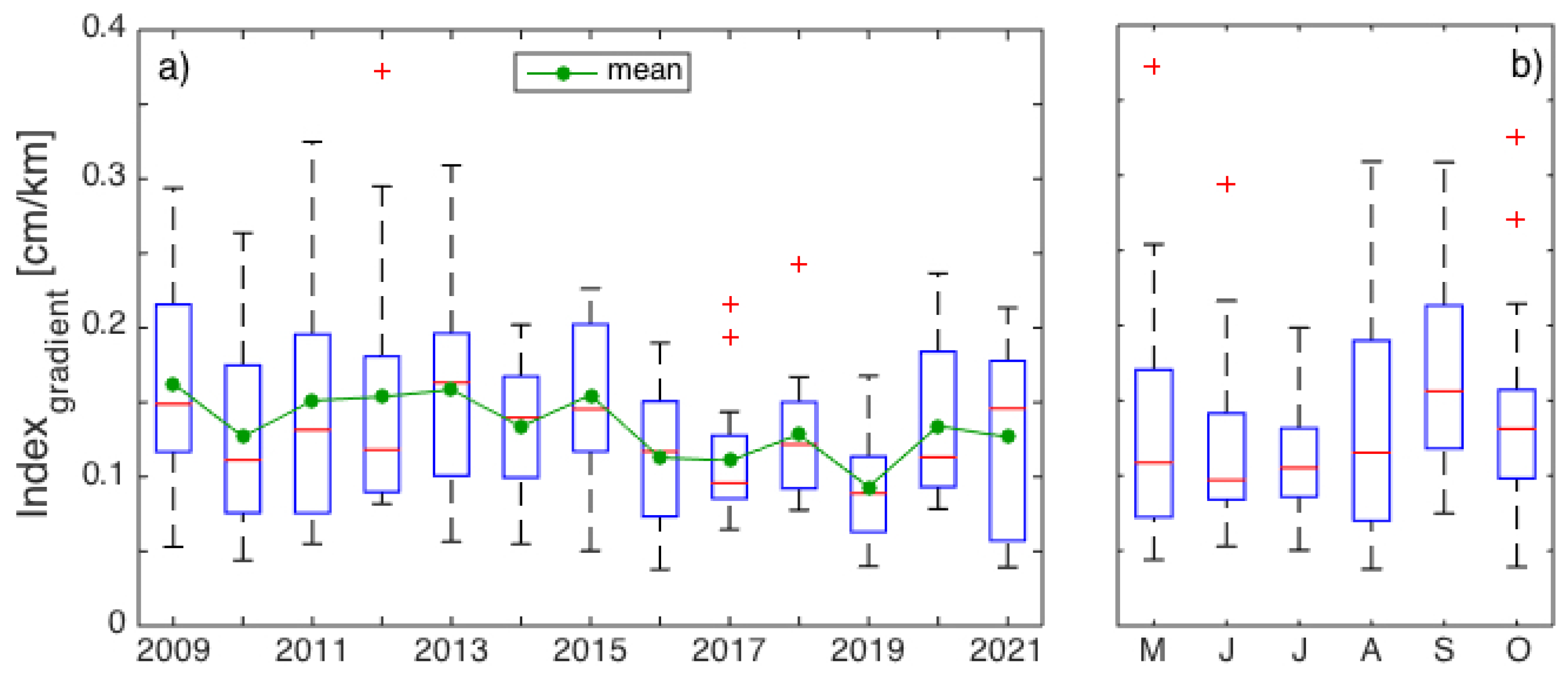
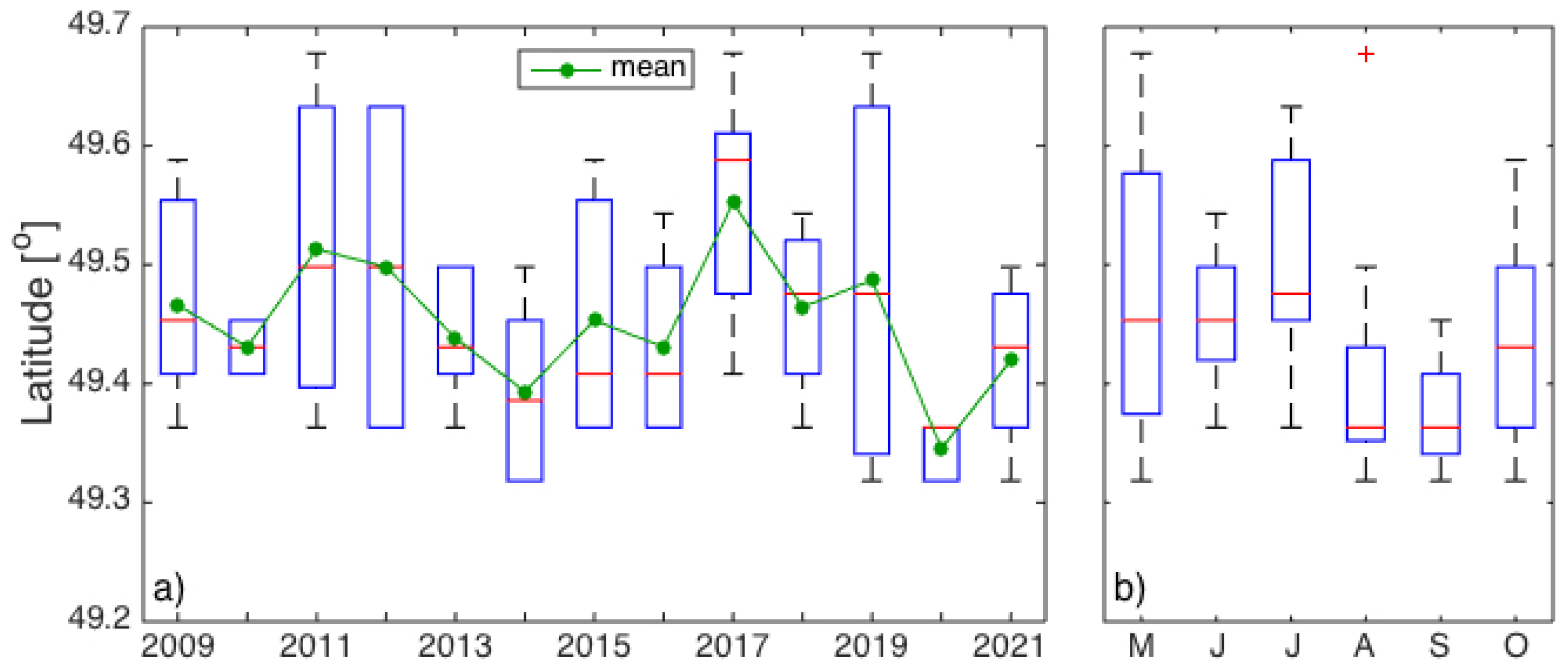
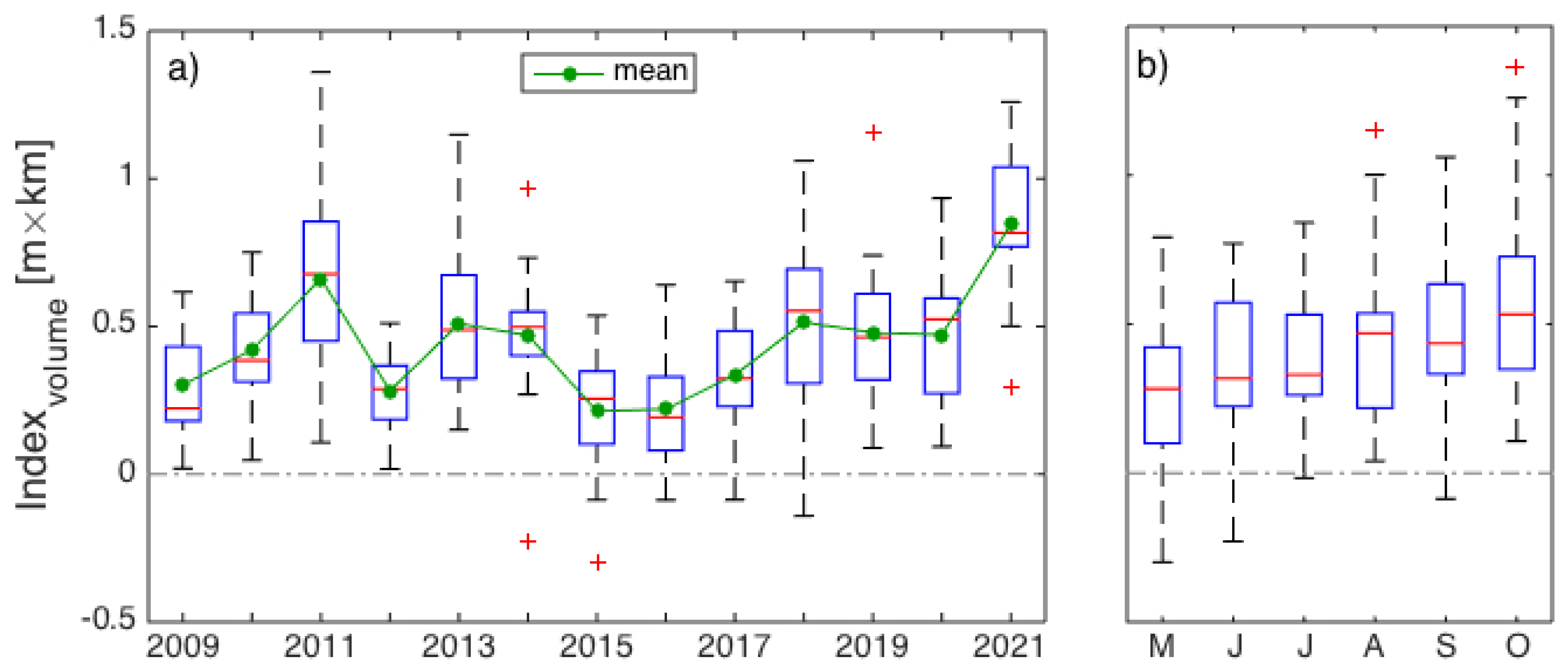
3. Results
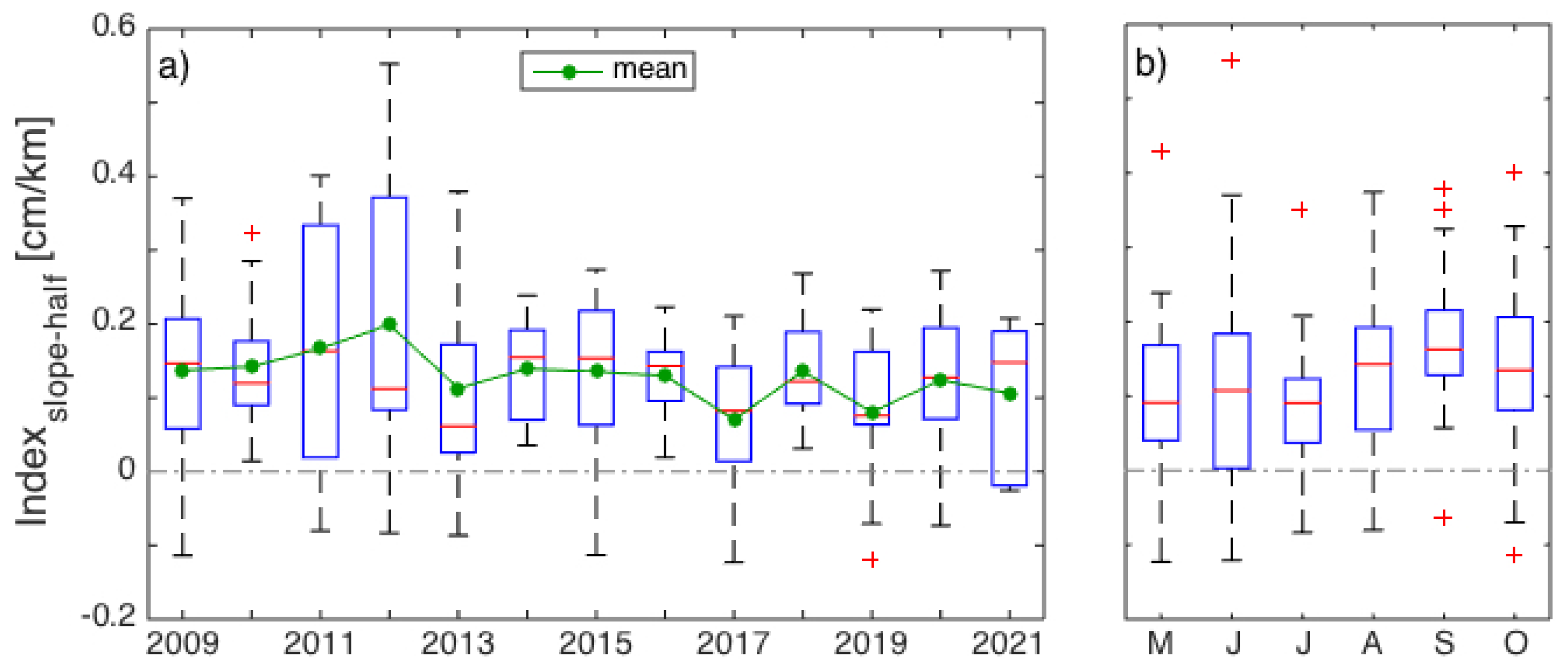
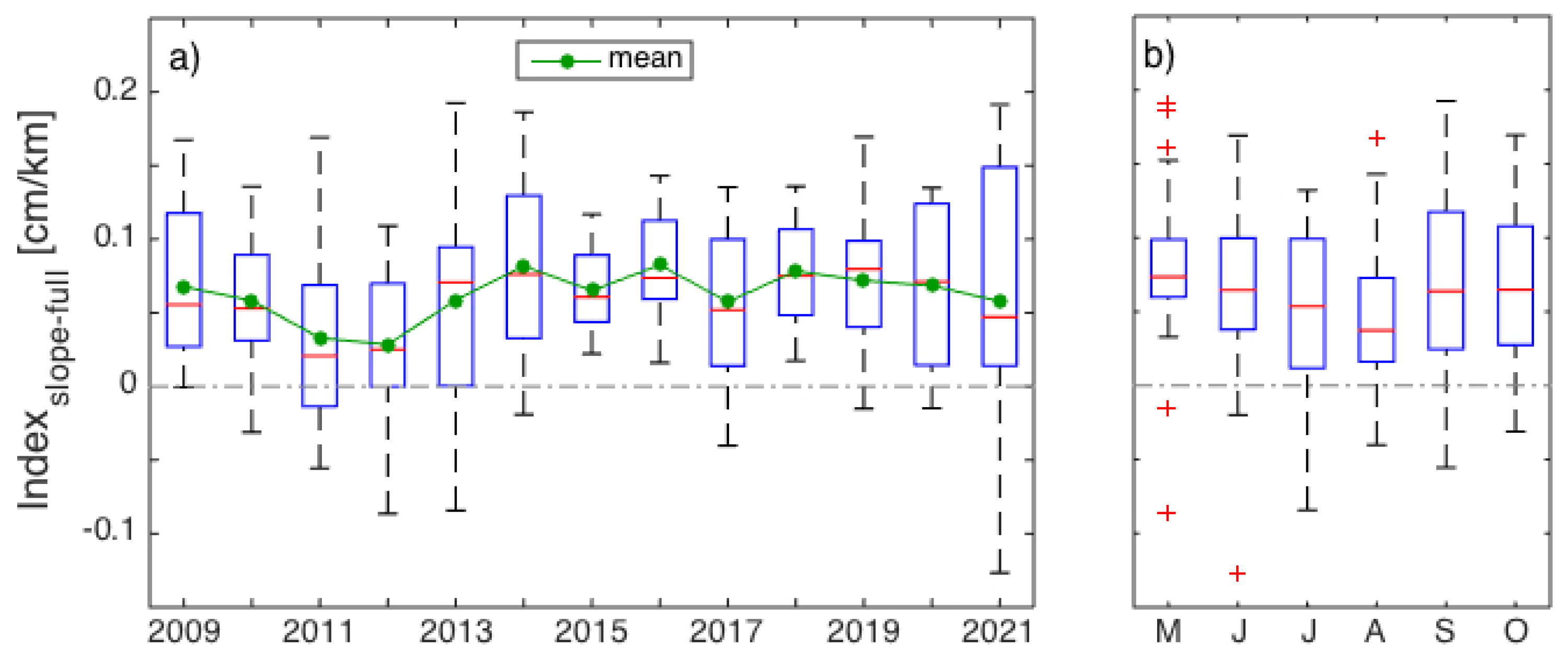
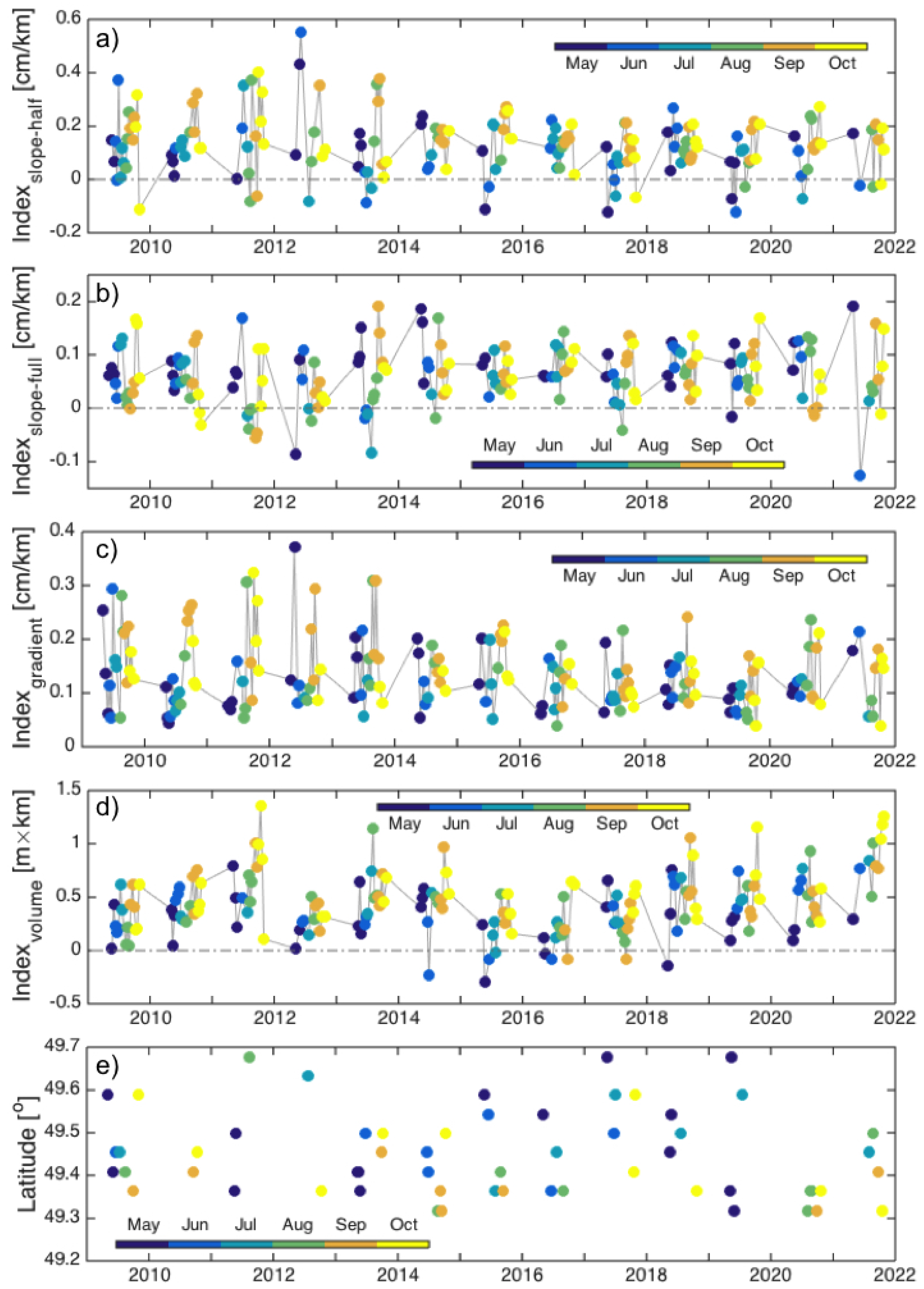
3.1. Interannual Variability in SLA Indices
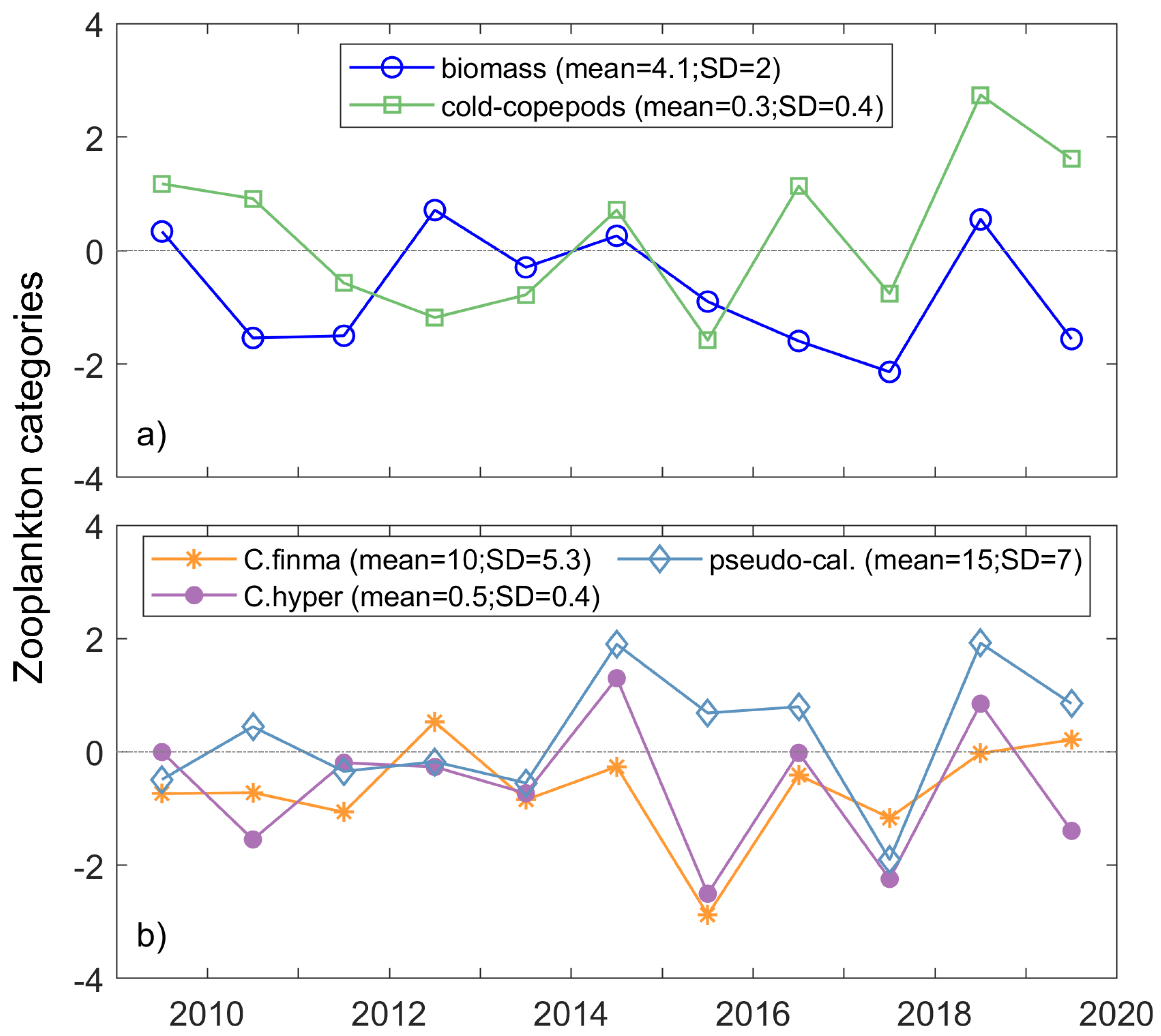
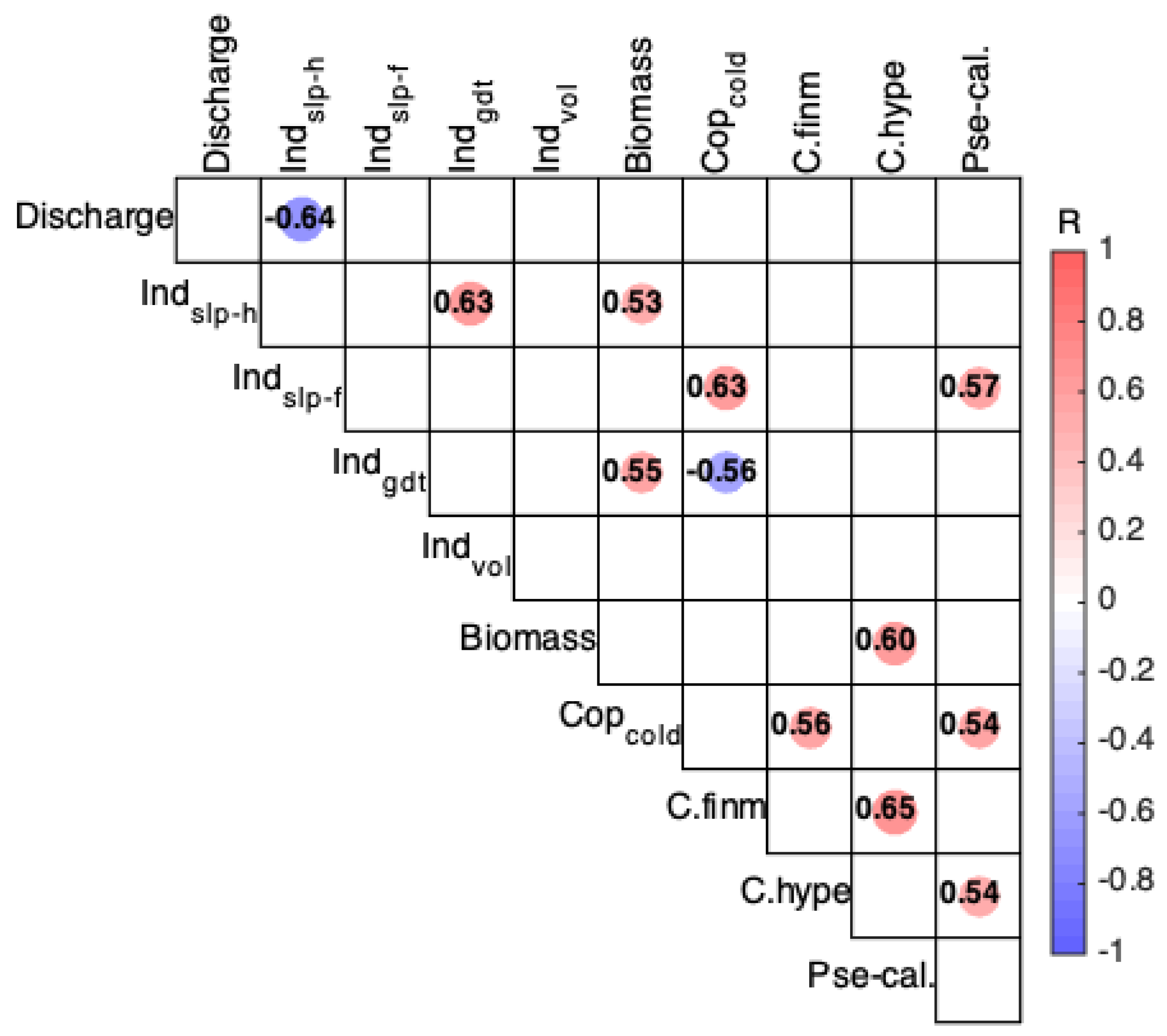
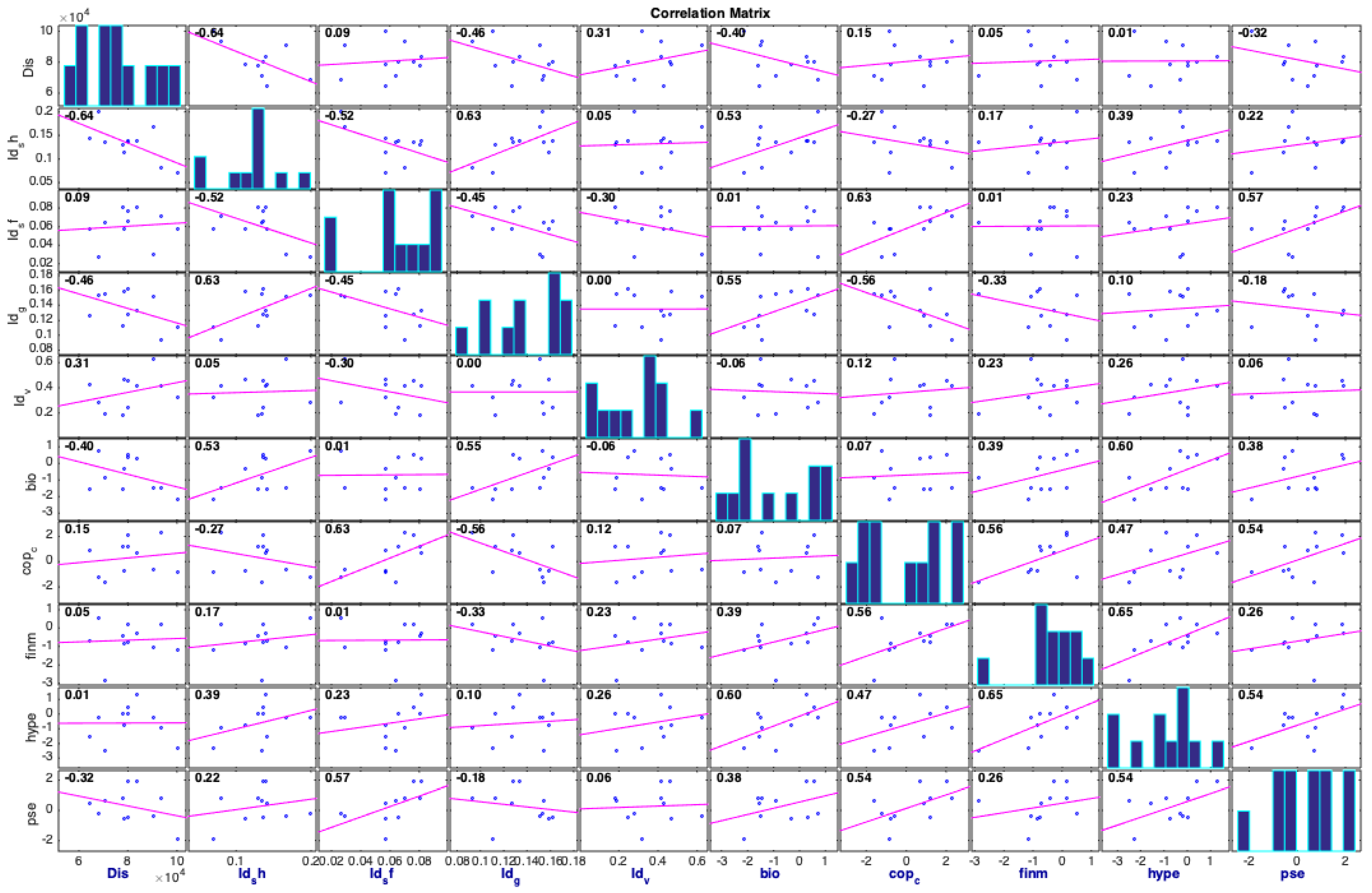
3.2. Correlation Analysis between Physical and Zooplankton Indices
4. Discussion
4.1. Gaspé Current Index Developed from Satellite Altimetry
4.2. Physical Indices Linked to the Zooplankton Variations
4.3. Altimetric Indices to Infer Whale Foraging Habitats
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Cullen, J.J.; Franks, P.J.; Karl, D.M.; Longhurst, A. Physical influences on marine ecosystem dynamics. Sea 2002, 12, 297–336. [Google Scholar]
- Cox, S.; Embling, C.; Hosegood, P.; Votier, S.; Ingram, S. Oceanographic drivers of marine mammal and seabird habitat-use across shelf-seas: A guide to key features and recommendations for future research and conservation management. Estuar. Coast. Shelf Sci. 2018, 212, 294–310. [Google Scholar] [CrossRef]
- Brennan, C.E.; Maps, F.; Gentleman, W.C.; Plourde, S.; Lavoie, D.; Chassé, J.; Lehoux, C.; Krumhansl, K.A.; Johnson, C.L. How transport shapes copepod distributions in relation to whale feeding habitat: Demonstration of a new modelling framework. Prog. Oceanogr. 2019, 171, 1–21. [Google Scholar] [CrossRef]
- Pineda, J.; Starczak, V.; da Silva, J.C.; Helfrich, K.; Thompson, M.; Wiley, D. Whales and waves: Humpback whale foraging response and the shoaling of internal waves at Stellwagen Bank. J. Geophys. Res. Ocean. 2015, 120, 2555–2570. [Google Scholar] [CrossRef]
- Franks, P.J. Sink or swim: Accumulation of biomass at fronts. Mar. Ecol. Prog. Ser. 1992, 82, 1–12. [Google Scholar] [CrossRef]
- Pineda, J. Circulation and larval distribution in internal tidal bore warm fronts. Limnol. Oceanogr. 1999, 44, 1400–1414. [Google Scholar] [CrossRef]
- Epstein, A.W.; Beardsley, R.C. Flow-induced aggregation of plankton at a front: A 2-D Eulerian model study. Deep Sea Res. Part II Top. Stud. Oceanogr. 2001, 48, 395–418. [Google Scholar] [CrossRef]
- Shanks, A.L.; Wright, W.G. Internal-wave-mediated shoreward transport of cyprids, megalopae, and gammarids and correlated longshore differences in the settling rate of intertidal barnacles. J. Exp. Mar. Biol. Ecol. 1987, 114, 1–13. [Google Scholar] [CrossRef]
- Lennert-Cody, C.E.; Franks, P.J. Plankton patchiness in high-frequency internal waves. Mar. Ecol. Prog. Ser. 1999, 186, 59–66. [Google Scholar] [CrossRef]
- Prairie, J.C.; Sutherland, K.R.; Nickols, K.J.; Kaltenberg, A.M. Biophysical interactions in the plankton: A cross-scale review. Limnol. Oceanogr. Fluids Environ. 2012, 2, 121–145. [Google Scholar] [CrossRef]
- Kingsford, M.; Wolanski, E.; Choat, J. Influence of tidally induced fronts and Langmuir circulations on distribution and movements of presettlement fishes around a coral reef. Mar. Biol. 1991, 109, 167–180. [Google Scholar] [CrossRef]
- Lavoie, D.; Simard, Y.; Saucier, F.J. Aggregation and dispersion of krill at channel heads and shelf edges: The dynamics in the Saguenay-St. Lawrence Marine Park. Can. J. Fish. Aquat. Sci. 2000, 57, 1853–1869. [Google Scholar] [CrossRef]
- Sorochan, K.; Brennan, C.E.; Plourde, S.; Johnson, C.L. Availability, supply, and aggregation of prey (Calanus spp.) in foraging areas of the North Atlantic right whale (Eubalaena glacialis). ICES J. Mar. Sci. 2021, 78, 3498–3520. [Google Scholar] [CrossRef]
- Mayo, C.A.; Marx, M.K. Surface foraging behaviour of the North Atlantic right whale, Eubalaena glacialis, and associated zooplankton characteristics. Can. J. Zool. 1990, 68, 2214–2220. [Google Scholar] [CrossRef]
- Winn, H.E.; Price, C.A.; Sorensen, P.W. The distributional biology of the right whale (Eubalaena glacialis) in the western North Atlantic. Rep.-Int. Whal. Comm. 1986, 10, 129–138. [Google Scholar]
- Woodley, T.H.; Gaskin, D.E. Environmental characteristics of North Atlantic right and fin whale habitat in the lower Bay of Fundy, Canada. Can. J. Zool. 1996, 74, 75–84. [Google Scholar] [CrossRef]
- Abdalla, S.; Kolahchi, A.A.; Ablain, M.; Adusumilli, S.; Bhowmick, S.A.; Alou-Font, E.; Amarouche, L.; Andersen, O.B.; Antich, H.; Aouf, L.; et al. Altimetry for the future: Building on 25 years of progress. Adv. Space Res. 2021, 68, 319–363. [Google Scholar] [CrossRef]
- Lama, G.F.C.; Sadeghifar, T.; Azad, M.T.; Sihag, P.; Kisi, O. On the indirect estimation of wind wave heights over the southern coasts of Caspian Sea: A comparative analysis. Water 2022, 14, 843. [Google Scholar] [CrossRef]
- Oziel, L.; Baudena, A.; Ardyna, M.; Massicotte, P.; Randelhoff, A.; Sallée, J.B.; Ingvaldsen, R.B.; Devred, E.; Babin, M. Faster Atlantic currents drive poleward expansion of temperate phytoplankton in the Arctic Ocean. Nat. Commun. 2020, 11, 1705. [Google Scholar] [CrossRef]
- Cotté, C.; d’Ovidio, F.; Chaigneau, A.; Lévy, M.; Taupier-Letage, I.; Mate, B.; Guinet, C. Scale-dependent interactions of Mediterranean whales with marine dynamics. Limnol. Oceanogr. 2011, 56, 219–232. [Google Scholar] [CrossRef]
- Koutitonsky, V.G.; Budgen, G. The physical oceanography of the Gulf of St. Lawrence: A review with emphasis on the synoptic variability of the motion. The Gulf St. Lawrence: Small ocean or big estuary? Can. Spec. Publ. Fish. Aquat. Sci. 1991, 113, 57–90. [Google Scholar]
- Han, G. Sea level and surface current variability in the Gulf of St Lawrence from satellite altimetry. Int. J. Remote Sens. 2004, 25, 5069–5088. [Google Scholar] [CrossRef]
- Benoit, J.; El-Sabh, M.; Tang, C. Structure and seasonal characteristics of the Gaspé Current. J. Geophys. Res. Ocean. 1985, 90, 3225–3236. [Google Scholar] [CrossRef]
- Lavoie, D.; Chassé, J.; Simard, Y.; Lambert, N.; Galbraith, P.S.; Roy, N.; Brickman, D. Large-scale atmospheric and oceanic control on krill transport into the St. Lawrence estuary evidenced with three-dimensional numerical modelling. Atmos.-Ocean 2016, 54, 299–325. [Google Scholar] [CrossRef]
- El-Sabh, M.I. Surface circulation pattern in the Gulf of St. Lawrence. J. Fish. Board Can. 1976, 33, 124–138. [Google Scholar] [CrossRef]
- Sheng, J. Dynamics of a buoyancy-driven coastal jet: The Gaspe Current. J. Phys. Oceanogr. 2001, 31, 3146–3162. [Google Scholar] [CrossRef]
- Tang, C. Mixing and circulation in the northwestern Gulf of St. Lawrence: A study of a buoyancy-driven current system. J. Geophys. Res. Ocean. 1980, 85, 2787–2796. [Google Scholar] [CrossRef]
- Ohashi, K.; Sheng, J. Influence of St. Lawrence River discharge on the circulation and hydrography in Canadian Atlantic waters. Cont. Shelf Res. 2013, 58, 32–49. [Google Scholar] [CrossRef]
- Lehoux, C.; Plourde, S.; Lesage, V. Significance of Dominant Zooplankton Species to the North Atlantic Right Whale Potential Foraging Habitats in the Gulf of St. Lawrence: A Bio-Energetic Approach; Canadian Science Advisory Secretariat (CSAS) 2020, 2020/033, vi+44p. Available online: https://epe.lac-bac.gc.ca/100/201/301/weekly_acquisitions_list-ef/2020/20-35/publications.gc.ca/collections/collection_2020/mpo-dfo/fs70-5/Fs70-5-2020-033-eng.pdf (accessed on 2 February 2022).
- Crowe, L.M.; Brown, M.W.; Corkeron, P.J.; Hamilton, P.K.; Ramp, C.; Ratelle, S.; Vanderlaan, A.S.; Cole, T.V. In plane sight: A mark-recapture analysis of North Atlantic right whales in the Gulf of St. Lawrence. Endanger. Species Res. 2021, 46, 227–251. [Google Scholar] [CrossRef]
- DFO. Updated Information on the Distribution of North Atlantic Right Whale in Canadian Waters. Canadian Science Advisory Secretariat (CSAS). Science Advisory Report 2020/037. 2020. Available online: https://waves-vagues.dfo-mpo.gc.ca/library-bibliotheque/40938554.pdf (accessed on 2 February 2022).
- DFO. Whalesightings Database; Team Whale, Fisheries and Oceans Canada: Dartmouth, NS, Canada, 2022. [Google Scholar]
- Durette-Morin, D.; Evers, C.; Johnson, H.D.; Kowarski, K.; Delarue, J.; Moors-Murphy, H.; Maxner, E.; Lawson, J.W.; Davies, K.T. The distribution of North Atlantic right whales in Canadian waters from 2015–2017 revealed by passive acoustic monitoring. Front. Mar. Sci. 2022, 9, 976044. [Google Scholar] [CrossRef]
- Simard, Y.; Roy, N.; Giard, S.; Aulanier, F. North Atlantic right whale shift to the Gulf of St. Lawrence in 2015, revealed by long-term passive acoustics. Endanger. Species Res. 2019, 40, 271–284. [Google Scholar] [CrossRef]
- Gavrilchuk, K.; Lesage, V.; Fortune, S.; Trites, A.; Plourde, S. A Mechanistic Approach to Predicting Suitable Foraging Habitat for Reproductively Mature North Atlantic Right Whales in the Gulf of St. Lawrence; Canadian Science Advisory Secretariat (CSAS): Ottawa, ON, Canada, 2020; Volume 2020/034, Available online: https://publications.gc.ca/site/eng/9.890978/publication.html (accessed on 2 February 2022).
- Blais, M.; Galbraith, P.; Plourde, S.; Devine, L.; Lehoux, C. Chemical and Biological Oceanographic Conditions in the Estuary and Gulf of St. Lawrence during 2019; DFO Canadian Science Advisory Secretariat Research Document; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2021; Volume 2021/002, p. iv + 66. Available online: https://publications.gc.ca/site/eng/9.896897/publication.html (accessed on 31 August 2023).
- Conover, R. Comparative life histories in the genera Calanus and Neocalanus in high latitudes of the northern hemisphere. Hydrobiologia 1988, 167, 127–142. [Google Scholar] [CrossRef]
- Brennan, C.E.; Maps, F.; Gentleman, W.C.; Lavoie, D.; Chassé, J.; Plourde, S.; Johnson, C.L. Ocean circulation changes drive shifts in Calanus abundance in North Atlantic right whale foraging habitat: A model comparison of cool and warm year scenarios. Prog. Oceanogr. 2021, 197, 102629. [Google Scholar] [CrossRef]
- Plourde, S.; Lehoux, C.; Johnson, C.; Perrin, G.; Lesage, V. North Atlantic right whale (Eubalaena glacialis) Its Food: A (I) spatial climatology of Calanus biomass and new potential foraging habitats in Canadian waters. J. Plankton Res. 2019, 41, 667–685. [Google Scholar] [CrossRef]
- Sorochan, K.; Brennan, C.E.; Plourde, S.; Johnson, C.L. Spatial variation and transport of abundant copepod taxa in the southern Gulf of St. Lawrence in autumn. J. Plankton Res. 2021, 43, 908–926. [Google Scholar] [CrossRef]
- Maps, F.; Zakardjian, B.A.; Plourde, S.; Saucier, F.J. Modeling the interactions between the seasonal and diel migration behaviors of Calanus finmarchicus and the circulation in the Gulf of St. Lawrence (Canada). J. Mar. Syst. 2011, 88, 183–202. [Google Scholar] [CrossRef]
- Benkort, D.; Lavoie, D.; Plourde, S.; Dufresne, C.; Maps, F. Arctic and Nordic krill circuits of production revealed by the interactions between their physiology, swimming behaviour and circulation. Prog. Oceanogr. 2020, 182, 102270. [Google Scholar] [CrossRef]
- Plourde, S.; Runge, J.A. Reproduction of the planktonic copepod Calanus finmarchicus in the Lower St. Lawrence Estuary: Relation 682 to the cycle of phytoplankton production and evidence for a Calanus pump. Mar. Ecol. Prog. Ser. 1993, 102, 217–227. [Google Scholar] [CrossRef]
- Fu, L.L.; Chelton, D.B. Large-scale ocean circulation. In International Geophysics; Elsevier: Berlin/Heidelberg, Germany, 2001; Volume 69, pp. 133–169, iii–viii. [Google Scholar] [CrossRef]
- Bonjean, F.; Lagerloef, G.S. Diagnostic model and analysis of the surface currents in the tropical Pacific Ocean. J. Phys. Oceanogr. 2002, 32, 2938–2954. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, L.; Li, Z.; Zhou, X. Geostrophic current estimation using altimetric cross-track method in northwest Pacific Ocean. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2014; Volume 17, p. 012105. [Google Scholar]
- Joseph, A. Measuring Ocean Currents: Tools, Technologies, and Data; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Han, G.; Tang, C.; Smith, P. Annual variations of sea surface elevation and currents over the Scotian Shelf and slope. J. Phys. Oceanogr. 2002, 32, 1794–1810. [Google Scholar] [CrossRef]
- Rio, M.H.; Mulet, S.; Picot, N. Beyond GOCE for the ocean circulation estimate: Synergetic use of altimetry, gravimetry, and in situ data provides new insight into geostrophic and Ekman currents. Geophys. Res. Lett. 2014, 41, 8918–8925. [Google Scholar] [CrossRef]
- Mertz, G.; El-Sabh, M.I.; Proulx, D.; Condal, A.R. Instability of a buoyancy-driven coastal jet: The Gaspé Current and its St. Lawrence precursor. J. Geophys. Res. Ocean. 1988, 93, 6885–6893. [Google Scholar] [CrossRef]
- Mertz, G.; El-sabh, M.I. An Autumn Instability Event in the Gaspé: Current. J. Phys. Oceanogr. 1989, 19, 148–156. [Google Scholar] [CrossRef]
- Lavoie, D.; Starr, M.; Zakardjian, B.; Larouche, P. Identification of Ecologically and Biologically Significant Areas (EBSA) in the Estuary and Gulf of St. Lawrence: Primary Production; Fisheries and Oceans: Langley, BC, Canada, 2008.
- Bugden, G. Freshwater runoff effects in the marine environment: The Gulf of St Lawrence example. Can. Tech. Rep. Fish. Aquat. Sci. 1982, 1078, 1–89. [Google Scholar]
- Bourgault, D.; Koutitonsky, V.G. Real-time monitoring of the freshwater discharge at the head of the St. Lawrence Estuary. Atmos.-Ocean 1999, 37, 203–220. [Google Scholar] [CrossRef]
- Galbraith, P.; Chassé, J.; Shaw, J.-L.; Dumas, J.; Caverhill, C.; Lefaivre, D.; Lafleur, C. Physical Oceanographic Conditions in the Gulf of St. Lawrence during 2020; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2021; Volume 2021/045, pp. 1919–5044. Available online: https://publications.gc.ca/site/eng/9.900562/publication.html (accessed on 2 February 2022).
- Hersbach, H.; Bell, B.; Berrisford, P.; Hirahara, S.; Horányi, A.; Muñoz-Sabater, J.; Nicolas, J.; Peubey, C.; Radu, R.; Schepers, D.; et al. The ERA5 global reanalysis. Q. J. R. Meteorol. Soc. 2020, 146, 1999–2049. [Google Scholar] [CrossRef]
- Kenney, R.D. The North Atlantic Right Whale Consortium Database: A Guide for Users and Contributors. 2019. Available online: https://www.narwc.org/uploads/1/1/6/6/116623219/narwc_users_guide__v_6_.pdf (accessed on 2 February 2022).
- NARWC. Identification and Sightings Data of the North Atlantic Right Whale Consortium as of January 2021; New England Aquarium: Boston, MA, USA, 2021. [Google Scholar]
- DFO. Review of North Atlantic Right Whale Occurrence and Risk of Entanglements in Fishing Gear and Vessel Strikes in Canadian Waters; Canadian Science Advisory Secretariat: Ottawa, ON, Canada, 2019; Volume 2019/028, Available online: https://publications.gc.ca/site/eng/9.875486/publication.html (accessed on 2 February 2022).
- Cole, T.V.; Crowe, L.M.; Corkeron, P.J.; Vanderlaan, A.S. North Atlantic Right Whale Abundance, Demography and Residency in the Southern Gulf of St. Lawrence Derived from Directed Aerial Surveys. Canadian Science Advisory Secretariat, 2020/063. 2020. Available online: https://publications.gc.ca/site/eng/9.901892/publication.html (accessed on 2 February 2022).
- Mitchell, M.R.; Harrison, G.; Pauley, K.; Gagné, A.; Maillet, G.; Strain, P. Atlantic Zontal Monitoting Program Sampling Protocol. In Canadian Technical Report of Hydrography and Ocean Sciences; Fisheries & Oceans Canada, Maritimes Region, Ocean Sciences Division, Bedford Institute of Oceanography: Dartmouth, NS, Canada, 2002; Volume 223, Available online: https://publications.gc.ca/site/eng/9.562291/publication.html (accessed on 2 February 2022).
- Williamson, D.F.; Parker, R.A.; Kendrick, J.S. The box plot: A simple visual method to interpret data. Ann. Intern. Med. 1989, 110, 916–921. [Google Scholar] [CrossRef]
- Halimi, A.; Mailhes, C.; Tourneret, J.Y.; Thibaut, P.; Boy, F. Parameter estimation for peaky altimetric waveforms. IEEE Trans. Geosci. Remote Sens. 2012, 51, 1568–1577. [Google Scholar] [CrossRef]
- Bobanović, J.; Thompson, K.R. The influence of local and remote winds on the synoptic sea level variability in the Gulf of Saint Lawrence. Cont. Shelf Res. 2001, 21, 129–144. [Google Scholar] [CrossRef]
- Neu, H.J.A. Man-made changes–The potential benefits and liabilities. In Bedford Institute, Gulf of St Lawrence Workshop; Bedford Institute of Oceanography: Dartmouth, NS, Canada, 1968; pp. 75–77. [Google Scholar]
- Petrie, B.; Toulany, B.; Garrett, C. The transport of water, heat and salt through the Strait of Belle Isle. Atmos.-Ocean 1988, 26, 234–251. [Google Scholar] [CrossRef]
- Hare, J.A.; Kane, J. Zooplankton of the Gulf of Maine—A changing perspective. In American Fisheries Society Symposium; American Fisheries Society: Bethesda, MD, USA, 2012; Volume 79, pp. 115–137. [Google Scholar]
- Zakardjian, B.A.; Sheng, J.; Runge, J.A.; McLaren, I.; Plourde, S.; Thompson, K.R.; Gratton, Y. Effects of temperature and circulation on the population dynamics of Calanus finmarchicus in the Gulf of St. Lawrence and Scotian Shelf: Study with a coupled, three-dimensional hydrodynamic, stage-based life history model. J. Geophys. Res. Ocean. 2003, 108. [Google Scholar] [CrossRef]
- Morrow, R.; Fu, L.L.; Ardhuin, F.; Benkiran, M.; Chapron, B.; Cosme, E.; d’Ovidio, F.; Farrar, J.T.; Gille, S.T.; Lapeyre, G.; et al. Global observations of fine-scale ocean surface topography with the Surface Water and Ocean Topography (SWOT) mission. Front. Mar. Sci. 2019, 6, 232. [Google Scholar] [CrossRef]
- Han, G.; Perrie, W.; Hannah, C.; Bianucci, L.; Grisouard, N.; Straub, D.; Klymak, J.; Smith, G.; Matte, P. Integration of SWOT Measurements in Canadian Oceanographic Research and Operation. Surface Water and Ocean Topography (SWOT) Mission Document. 2020. Available online: https://swot.jpl.nasa.gov/system/documents/files/4175_SWOT_Canada_Project_Summary.pdf?list=projects (accessed on 2 February 2022).














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tao, J.; Shen, H.; Danielson, R.E.; Perrie, W. Characterizing the Variability of a Physical Driver of North Atlantic Right Whale Foraging Habitat Using Altimetric Indices. J. Mar. Sci. Eng. 2023, 11, 1760. https://doi.org/10.3390/jmse11091760
Tao J, Shen H, Danielson RE, Perrie W. Characterizing the Variability of a Physical Driver of North Atlantic Right Whale Foraging Habitat Using Altimetric Indices. Journal of Marine Science and Engineering. 2023; 11(9):1760. https://doi.org/10.3390/jmse11091760
Chicago/Turabian StyleTao, Jing, Hui Shen, Richard E. Danielson, and William Perrie. 2023. "Characterizing the Variability of a Physical Driver of North Atlantic Right Whale Foraging Habitat Using Altimetric Indices" Journal of Marine Science and Engineering 11, no. 9: 1760. https://doi.org/10.3390/jmse11091760