We will contrast the above behaviour with the results discussed below for unsteady insolation.
4.1. Homogeneous Ocean with Background Algal Field Absent
We begin with a highly idealised example in which the ocean is unstratified, (i.e.,
), corresponding to
, in (18). Upon dropping the tilde superfix denoting a non-dimensional quantity, the governing equations for the biofouled microplastic trajectory become:
upon setting
, corresponding to no background algal field.
In analysing the biofouled particle trajectory, a useful diagnostic is the “compensation depth” where production and mortality of the algae are in balance. At this depth,
, and (28) implies that the compensation depth,
satisfies
From (29), the non-dimensional compensation depth,
, is given by
If a background algal field is present (i.e.,
is not identical to zero), we postulate that no modification of (30) is required because
is not associated with “new photosynthetic algae production”.
The governing Equations (27) and (28), and more generally (17) and (21), are solved numerically using Python. A Runge–Kutta–Fehlberg adaptive time step scheme is employed, with a minimum time step of
. The following dimensionless parameter values
,
, and
are used, corresponding to an oscillation period of the clean particle of approximately one day. The particle trajectories, forced by time-varying insolation, are calculated at latitudes
,
and
.
Figure 1 shows the particle trajectory (blue curve) at
over a 5-day interval, where the red line is the compensation depth and the green line is the constant euphotic zone depth. We note that, over such a short time interval, the compensation depth is approximately constant.
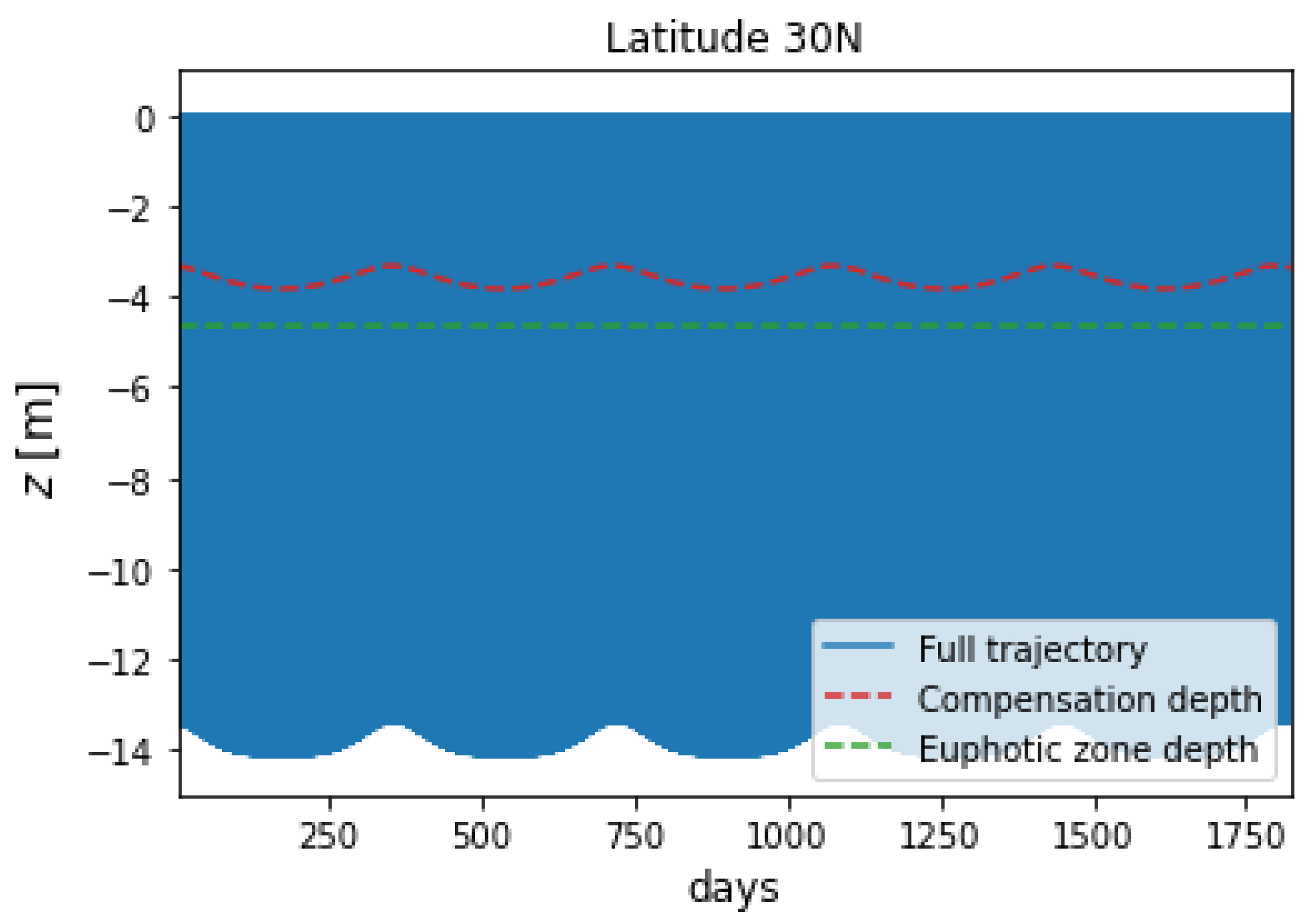
Figure 2 depicts the 5-year trajectory of a microplastic particle at a latitude of 30N, where the seasonal behaviour of the compensation depth is apparent.
Figure 1 reveals that the particle remains at the sea surface for a short period until it acquires sufficient biofilm to become negatively buoyant. It then performs vertical oscillations in the upper water column with a daily period. Once the particle has sunk beneath the euphotic layer, defouling starts to occur, due to a lack of sunlight. In addition, the periods of darkness, when no photosynthesis takes place, also contribute to the decay of the biofilm. As the biofilm dies off, the particle trajectory starts to slow down. The particle reaches a depth where its buoyancy is neutral, and thereafter it begins to rise. Upon reaching the surface, the particle remains there until it acquires sufficient biofilm to become negatively buoyant again. In summer, the particle sinks to a deeper depth than in winter, due to higher amounts of insolation being present during the summer months, supporting greater biofilm mass than in the winter, as seen in
Figure 2.
Figure 2 also shows that the microplastic particle trajectory mirrors the time-varying compensation depth, being shallower in winter and deeper in summer, which is to be expected as the latter is calculated from daily average insolation and represents the depth at which the algal growth is balanced by the mortality rate.
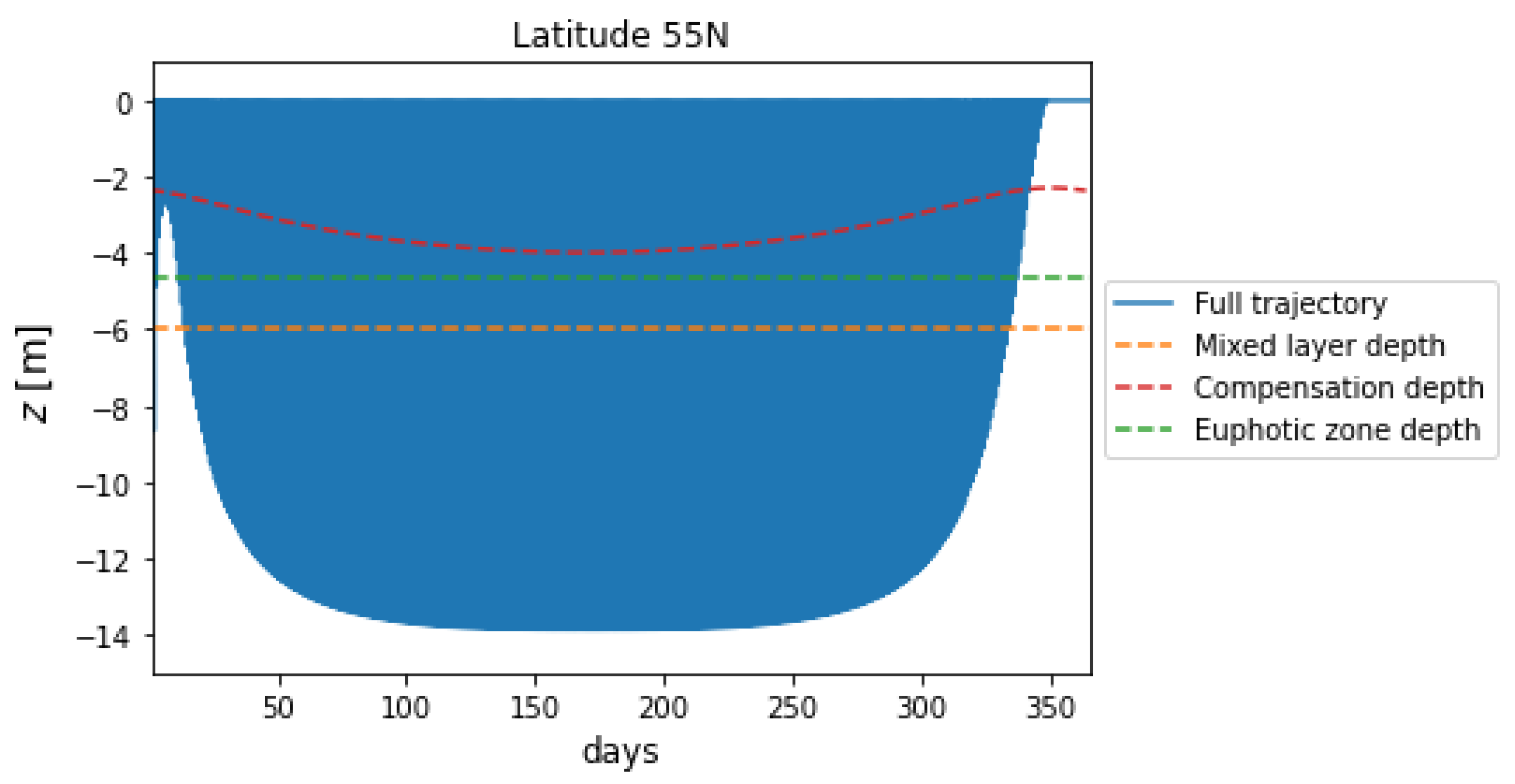
Figure 3 plots the 5-year trajectory of a microplastic particle at a latitude of 55N. The particle trajectory displays the same qualitative behaviour as a particle at 30N. Initially, the particle begins sinking, but to a shallower depth than the particle observed at 30N due to there being less insolation, and thus less biofilm growth, during winter at 55N. We observe that the particle exhibits oscillatory behaviour, resting at the surface for a period of days during the winter months when biofilm growth is at a minimum. As the year progresses from winter to summer, the particle begins to sink deeper, due to an increase in daily integrated insolation supporting greater biofilm mass. Once again, the particle trajectory over each year mirrors the behaviour of the compensation depth.
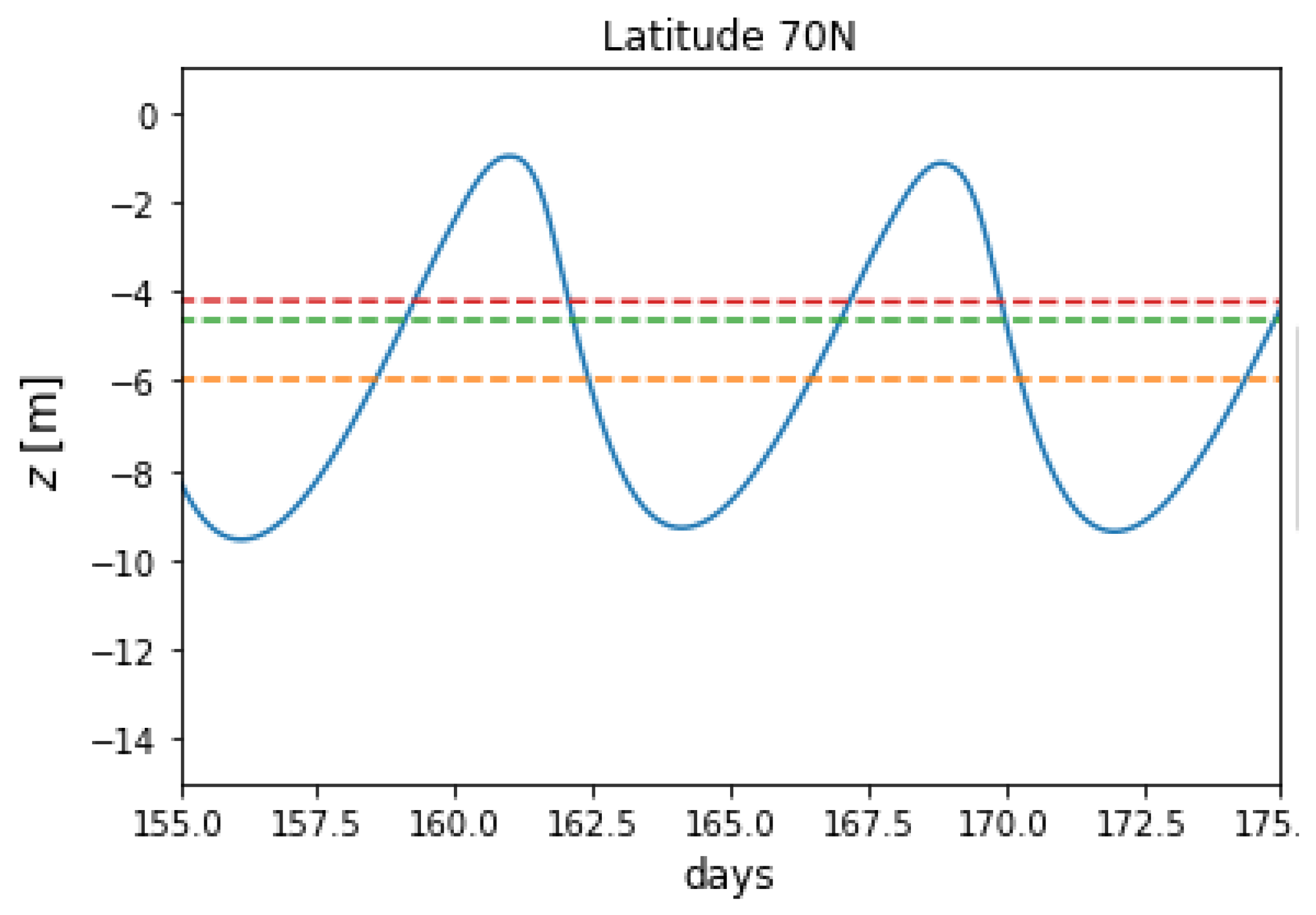
Figure 4 plots the 5-year trajectory of a microplastic particle at a latitude of 70N. Now, the particle displays aperiodic behaviour, unlike the trajectories in
Figure 2 and
Figure 3.
At this high latitude, the particle floats at the ocean surface for a prolonged period of time during the winter and over the early spring, when there is little or no insolation present and hence no biofilm production to make the particle negatively buoyant.
Once there is sufficient insolation for biofilm growth to occur, the particle sinks suddenly. The cause of this sudden drop is revealed in
Figure 5, where we observe that the algal population increases until it reaches a critical level at around day 105, when the particle becomes negatively buoyant. During the summer, there is a period of time during which the microplastic does not resurface. This is because the sun does not set, and so there is always a certain amount of penetrative radiation present in the upper ocean water column. When the biofilm begins to die off below the base of the euphotic layer, the particle begins to rise, and once it has re-entered the euphotic layer there is sufficient summer insolation for photosynthesis to begin biofilm growth once more, thus preventing the particle resurfacing. Unlike
Figure 2 and
Figure 3, the particle trajectory does not mimic the behaviour of the compensation depth. However, it is observed that at times when the compensation depth is at a maximum, the particle begins to sink after long periods of rest at the ocean surface. At these times, the biofilm production is near its maximum, and in the water column above this depth, biofilm production exceeds mortality.
In summary, it can be seen that there are significant variations between the particle trajectories in
Figure 2,
Figure 3 and
Figure 4. Microplastic trajectories at latitudes 30N and 55N exhibit time-periodic behaviour, whereas the trajectory of a microplastic particle at 70N displays aperiodic oscillations. At both 55N and 70N, there are prolonged periods of time in winter where the microplastic rests at the ocean surface, whereas this behaviour is not seen at 30N. It is also shown that there is no year-on-year accumulating effect of insolation on biofilm growth. Each year, the trajectory of the microplastic particle is the same, with the only exception being at a latitude of 70N for the first year, due to the initial conditions for the numerical integration of (27) and (28). The maximum depth achieved by the microplastic at these three latitudes is approximately the same, namely
, or around three times the euphotic zone depth.
We now contrast the particle trajectories plotted in
Figure 3 and
Figure 4 with those forced by constant insolation considered by Kreczak et al. (2021) [
20]. At latitudes 55N and 70N, the annual average insolation is used to force the microplastic particle.
Figure 6 and
Figure 7 show the corresponding particle trajectory over a 30-day time interval at latitudes 55N and 70N, respectively. The 30-day time interval used in these plots clearly resolves the particle oscillation.
In
Figure 6 and
Figure 7, the maximum depth reached by the particle is significantly shallower compared with the summer maximum depth acquired by the particle when insolation varies with time (see
Figure 3 and
Figure 4). This is because the summer biofilm production in
Figure 3 and
Figure 4 exceeds the constant biofilm production rate in
Figure 6 and
Figure 7. In
Figure 6 and
Figure 7, the maximum depth attained by the particle is greater at a latitude of 55N compared with 70N, due to the slightly greater average annual insolation at the lower latitude. Naturally, this behaviour is also reflected in the compensation depth. Note also that the microplastic particle does not float at the ocean surface for a period of time during winter when insolation is constant, due to biofilm growth occurring throughout the year when the particle enters the euphotic zone. With unsteady insolation, there is no biofilm growth in the absence of sunlight, irrespective of whether the particle is in the euphotic layer or not.
4.2. Stratified Ocean with Background Algal Field Absent
In the previous subsection, we assumed that there was no stratification in the ocean by setting . In this section, we introduce a stratified ocean with density profile given by in (20). Once again, we neglect algal attachment () onto the microplastic from a pre-existing field in the upper ocean.
In Kreczak et al. (2021) [
20], a step stratification was adopted, where the pycnocline depth was infinitesimally thin. In this study, the stratification (20) has a more realistic finite-depth pycnocline, which scales as
. Recall from (20) that in dimensionless form this becomes
. We now examine the microplastic trajectories as a function of the pycnocline depth for two cases: (i) mixed layer deeper than the euphotic layer; (ii) mixed layer shallower than the euphotic layer. In the following numerical results,
,
, and
. A euphotic layer depth of 75m is fixed throughout the results using a stratified ocean, corresponding to
. For comparison with Kreczak et al. (2021) [
20], we will first use constant insolation fixed at the annual average value at 55N, followed by results for time-varying insolation at this latitude.
Figure 8 plots the particle trajectory when insolation is constant, with a pycnocline thickness of 50 m and a mixed layer depth below the euphotic zone. Unlike the behaviour observed in
Figure 3, the particle no longer reaches the ocean surface and instead oscillates in a subsurface duct, bounded below by the base of the mixed layer, similar to the behaviour observed in Kreczak et al. (2021) [
20]. This subsurface trapping behaviour is due to the density jump in the pycnocline, displayed in
Figure 9. Once the particle enters the pycnocline, its velocity decreases due to its relative buoyancy decreasing.
Fewer algal cells need to be removed in order to attain neutral buoyancy in the pycnocline. The particle begins to rise, crossing back into the mixed layer, and now weighing more than it did on its descent. When the particle enters the euphotic zone, biofilm growth restarts, and now, less time is required for the particle to attain neutral buoyancy. Thus, the particle does not resurface. This process repeats until the particle becomes trapped above the pycnocline.
Figure 10 plots the particle trajectory for a pycnocline thickness of 500 m, for comparison with
Figure 8. There are some notable differences between the two figures. For instance, the adjustment time for the particle in
Figure 10 to exhibit trapped subsurface oscillations of fixed amplitude, bounded below by the base of the mixed layer, is longer compared to
Figure 8. With the smaller pycnocline density gradient in
Figure 11, compared with
Figure 9, the particle reaches neutral buoyancy at a greater depth, thus extending the damping timescale.
We now turn to the case when the mixed layer depth is shallower than the euphotic layer and examine the particle trajectories forced by constant, annually averaged insolation at 55N, as shown in
Figure 12 and
Figure 13.
Unlike
Figure 8 and
Figure 10, the particle’s oscillation is no longer bounded below by the mixed layer depth, and occurs within the pycnocline. The particle now displays rapidly damped oscillations about the average compensation depth, whereas in Kreczak et al. (2021) [
20], the particle oscillated about the base of the euphotic layer. Indeed, the large-time behaviour of the particle is one of equilibrium, as predicted by (17). From (17), this equilibrium depth within the pycnocline is given in dimensionless form as
Again, it can be seen that for a thicker pycnocline,
Figure 13, the adjustment time to reach equilibrium is longer compared with the thinner pycnocline example shown in
Figure 12.
We now introduce time-varying insolation, to assess its impact on the particle trajectory in a stratified ocean. For brevity, we set the pycnocline thickness to 500 m in all the results discussed in the remaining part of this section.
Figure 14 and
Figure 15 plot the particle trajectory at latitudes 55N and 70N, respectively, when the mixed layer is deeper than the euphotic layer.
Comparing
Figure 10 and
Figure 14, we see that the particle is no longer performing trapped subsurface oscillations when forced by unsteady insolation. However, at 70N (see
Figure 15), the particle displays three distinct dynamical regimes: (i) floating at the surface throughout the winter, (ii) spring and autumn large-amplitude surface-to-pycnocline oscillations; (iii) subsurface trapped oscillations about the compensation depth. In the summer, when the insolation is quasi-steady and at maximum, there is sufficient biofilm attached to the microplastic to prevent surfacing. At its deepest point within the pynocline during the summer, biofilm remains on the particle. Thus, on its ascent into the euphotic zone, it does not reach the surface because new, vigorous biofilm has grown once the particle is above the compensation depth. If the insolation were to remain fixed at its summer value throughout the following autumn and winter, the particle would perform damped subsurface oscillations trapped around the compensation depth. Of course, this does not occur in
Figure 15 because as summer gives way to autumn and biofilm production tails off, the particle reverts to oscillations that reach the sea surface.
We now consider the trajectory behaviour throughout the year at 70N when the mixed layer is shallower than the euphotic layer, more typical of that in summer. For brevity, we omit the plot for 55N using this stratification, because it is very similar to that of
Figure 14.
Figure 16 plots the annual trajectory of a particle forced by time-varying insolation at 70N and with a shallow mixed layer. Comparing
Figure 15 and
Figure 16, we see that the shallower mixed layer leads to more strongly damped subsurface oscillations about the compensation depth. Once again, the particle in
Figure 16 exhibits three regimes of trajectory behaviour throughout the year.
4.3. Varying the Clean Microplastic Particle Size
In all of the above results, the clean microplastic particle size was set so that its oscillation period was equal to one day. In this section, we now examine the particle trajectory behaviour at 70N for smaller clean microplastic particles which have slower rise velocity [
4]. The following results include stratification with the mixed layer depth greater than the euphotic layer depth, a pycnocline thickness of 500 m and time-varying insolation. For simplicity, we again neglect the background algal field
. We keep the growth and mortality rate ratio,
, and the algal population and particle motion timescale ratio,
, both fixed.
Figure 17 plots the trajectory of a particle that is half the original particle size used in
Section 4.1 and
Section 4.2 (
). There are three distinct timescales at play in
Figure 17 that result in a highly non-linear oscillation. These are the diurnal and seasonal timescales of the insolation and the clean particle oscillation period. Qualitatively, the particle trajectories in
Figure 15 and
Figure 17 are identical, albeit with significant fluctuations in the amplitude seen in the former plot. During the summer, the slower rise speed for the biofouled particle in
Figure 17 leads to larger amounts of biofilm when it is in the euphotic zone compared with
Figure 15, which leads to a longer period of subsurface oscillations about the compensation depth.
Decreasing the size of the clean particle to one tenth of the original size,
, used in
Section 4.1 and
Section 4.2 drastically alters the trajectory behaviour, as shown in
Figure 18 and
Figure 19. With the onset of spring biofilm growth, the particle sinks and exhibits subsurface oscillations with an 8-day period (see
Figure 19) about the compensation depth. Now, the length of time during which the particle exhibits subsurface oscillations is approximately 140 days, following the trend noted in
Figure 17. In the autumn, the oscillations are confined to the mixed layer and reach the surface. As expected, during the winter months the oscillations cease entirely and the particle floats at the sea surface.
4.4. Discussion
In this study, the impact of time-dependent insolation on the vertical trajectory of a biofouled microplastic particle in the ocean is explored using the model of Kreczak et al. (2021) [
20], which in turn is a simplified version of the model by Kooi et al. (2017) [
19]. The latter study introduced the first one-dimensional deterministic model for predicting the vertical motion of a biofouled microplastic particle in the ocean. There are two notable departures from the model formulation of Kreczak et al. (2021) [
20] employed in this study. First, the daily timescale is used in formulating the dimensionless governing equations for the particle trajectory, rather than the time taken for a clean (buoyant) microplastic particle to traverse the euphotic layer. Insolation, which drives the particle motion by supporting biofilm production, varies on the daily and seasonal timescales, and it is natural to choose one of these to non-dimensionalise time. Second, a more realistic density profile with a non-zero finite depth pycnocline is used in this study, whereas Kreczak et al. (2021) [
20] employed a step density profile. The dimensionless governing equations for the microplastic particle motion reveal that seven dimensionless parameters govern its behaviour when neglecting a pre-existing algal field in the upper ocean. Fortunately, once the stratification and densities of the clean plastic and the biofilm are prescribed, the number of dimensionless parameters reduces to four.
A new diagnostic variable is introduced in this study, namely, the compensation depth, where biofilm growth is balanced by its mortality. Because growth is a function of irradiance, the compensation depth has the same time-dependent behaviour as the insolation. We demonstrate that, under certain circumstances, microplastic particles oscillate about the compensation depth, as discussed below.
Initially, the study examines the microplastic trajectories driven by unsteady insolation in a homogeneous ocean. In this case, the particle exhibits vertical oscillations that reach the sea surface, as in Kreczak et al. (2021) [
20], although with amplitude that varies seasonally. In the summer, the amplitude is at a maximum, reflecting the fact that biofilm production is at a maximum. Indeed, the seasonal behaviour of the oscillation amplitude is reflected in the seasonal behaviour of the compensation depth. Moving poleward, the particle spends an increasing amount of time floating at the sea surface, and the oscillations become aperiodic. For example, at 30N, the particle spends several hours at the sea surface during the night when biofilm production ceases. Throughout the year, at this latitude the oscillations persist. Moving to 55N, the oscillations are absent entirely during a period centred around the solstice, because there is insufficient biofilm production to make the particle negatively buoyant. This trend continues at 70N, where during the winter, insolation is absent altogether for several weeks, and the particle floats on the sea surface. When the oscillations commence at this high latitude, they are highly modulated, exhibiting maximum amplitude in the spring and fall. In both of these seasons, the particle reaches the sea surface during each oscillation cycle. However, in a neighbourhood centred around the summer solstice, the particle oscillations briefly become subsurface in response to the large quasi-steady biofilm production.
For comparison with Kreczak et al. (2021) [
20], results are also presented for a particle driven by constant, annual average insolation at 55N and 70N in a homogeneous ocean. In both cases, the particle exhibits periodic oscillations that reach the sea surface during every cycle, as noted in Kreczak et al. (2021) [
20]. The depth of these oscillations decreases with increasing latitude. Also of note, for a given latitude, the maximum summer amplitude of the oscillations driven by unsteady insolation is greater than that driven by the annual average insolation.
Particle trajectories are calculated in a stratified ocean when: (i) the pycnocline thickness varies, (ii) the depth of the mixed layer is deeper/shallower than the fixed euphotic layer depth. In contrast with this study, the step-density profile used by Kreczak et al. (2021) [
6] did not permit an investigation of how the pycnocline density gradient impacts on the particle motion. Forcing the particle motion with annually averaged insolation at 55N leads to large-time, constant-amplitude subsurface oscillations bounded below by the base of the mixed layer, irrespective of the pycnocline thickness. With a thick pycnocline, corresponding to a smaller vertical density gradient, the particle penetrates the pycnocline during the initial adjustment phase. When the mixed layer is shallower than the euphotic layer, significant biofilm growth can take place in the pycnocline. In this case, forcing the particle with annually averaged insolation at 55N leads to highly damped oscillations around the compensation depth, and for large times, the particle is trapped at this depth.
Particle trajectories are calculated using unsteady insolation at 55N and 70N for a fixed pycnocline thickness of 500 m and with a mixed layer deeper/shallower than the euphotic layer. At 55N, the particle exhibits surface-to-depth oscillations with amplitude varying seasonally. However, at 70N, the particle dynamics displays three regimes: (i) floating at the sea surface in winter, (ii) surface-to-depth oscillations in spring/fall with time-varying amplitude, (iii) subsurface oscillations around the compensation depth. The time interval during which subsurface oscillations persist increases/decreases if the mixed layer is shallower/deeper than the euphotic layer. Subsurface oscillation amplitude increases/decreases when the mixed layer is deeper/shallower than the euphotic layer.
Finally, the study investigates how decreasing the clean particle size impacts its trajectory when forced with time-varying insolation in a stratified fluid. As the clean particle size decreases, its rise speed decreases, leading to longer-period oscillations in the presence of biofilm attachment. This is borne out in the results. In addition, as the clean particle size decreases, the particle transitions from exhibiting the three dynamical regimes noted above to one, in which it either floats at the sea surface or performs subsurface oscillations about the compensation depth with seasonally varying amplitude.
In this study, we have not considered the impact of a pre-existing algal field on the microplastic trajectories (i.e., allowing
to be non-zero in Equation (
21)), because this warrants a study in its own right. The specification of the vertical distribution of this field, captured by
in (15), is highly uncertain and depends on location and season. Preliminary investigations indicate that earlier sinking of the biofouled microplastic particle to a greater depth is a consequence of a non-zero
. We have also assumed that the stratification is steady throughout the year, again to aid understanding of the dynamics of the biofouled particle forced by unsteady insolation. Local wind and buoyancy forcing gives rise to an upper mixed layer depth that varies on the diurnal and seasonal timescales [
30,
31]. Coupling a one-dimensional mixed layer model to the model described in this study to assess the impact of time-dependent stratification on the particle trajectory would be worthwhile.
The clean microplastic density has not been altered in this study, with the focus, instead, on time-varying insolation as a function of latitude driving the variable buoyancy of the biofouled particle. Increasing the clean particle density from below towards the density of the ocean surface mixed layer will lead to smaller-amplitude, shorter-period oscillations. Conversely, decreasing the clean particle density will lengthen the oscillation period and increase the oscillation amplitude. Clearly, these predictions are dependent on the rates of growth/decay of the biofilm, water column stratification and the depth of the euphotic layer.
We have also neglected the impact of a background ocean velocity field on the particle dynamics. Biofouled microplastic particles are expected to be strongly coupled to three-dimensional upper ocean turbulence. Kreczak et al. (2023) [
32] have addressed one aspect of this complex problem by examining the statistics of a large ensemble of biofouled microplastic particles in the presence of a Taylor–Green vortex flow. This latter flow is a prototypical representation of a Langmuir cell. The key question in such studies is whether the vortical cell(s) delay the descent of the microplastic particles, and if so, by how long. A worthwhile future study would be to couple the microplastic trajectory model presented in this paper to a numerical model for upper ocean three-dimensional turbulence in order to address the question of delayed particle fall-out.






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}