
Seasonal Dynamics of Marine Bacterial Communities in Aquaculture Farms: The Case of the Northern Ionian Coastal Ecosystem (Mediterranean Sea)
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Microbial DNA Extraction
2.3. Library Construction and Sequencing
2.4. Bioinformatics Analysis
2.5. Diversity and Composition Analysis
2.6. Differential Abundance Analysis
3. Results
3.1. Environmental Variables Characteristics
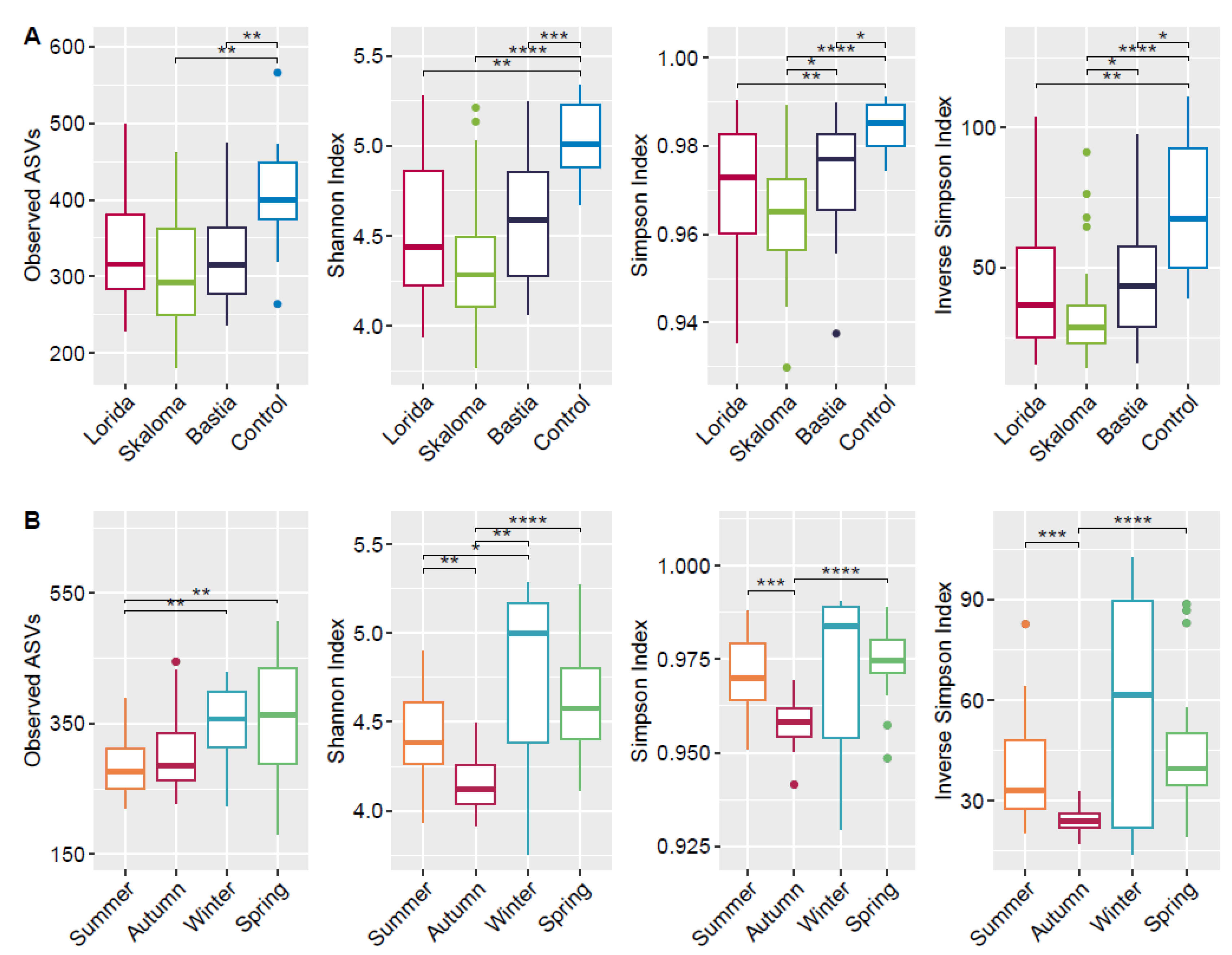
3.2. Sequencing Overview and Alpha Diversity Analysis of Bacterial Communities
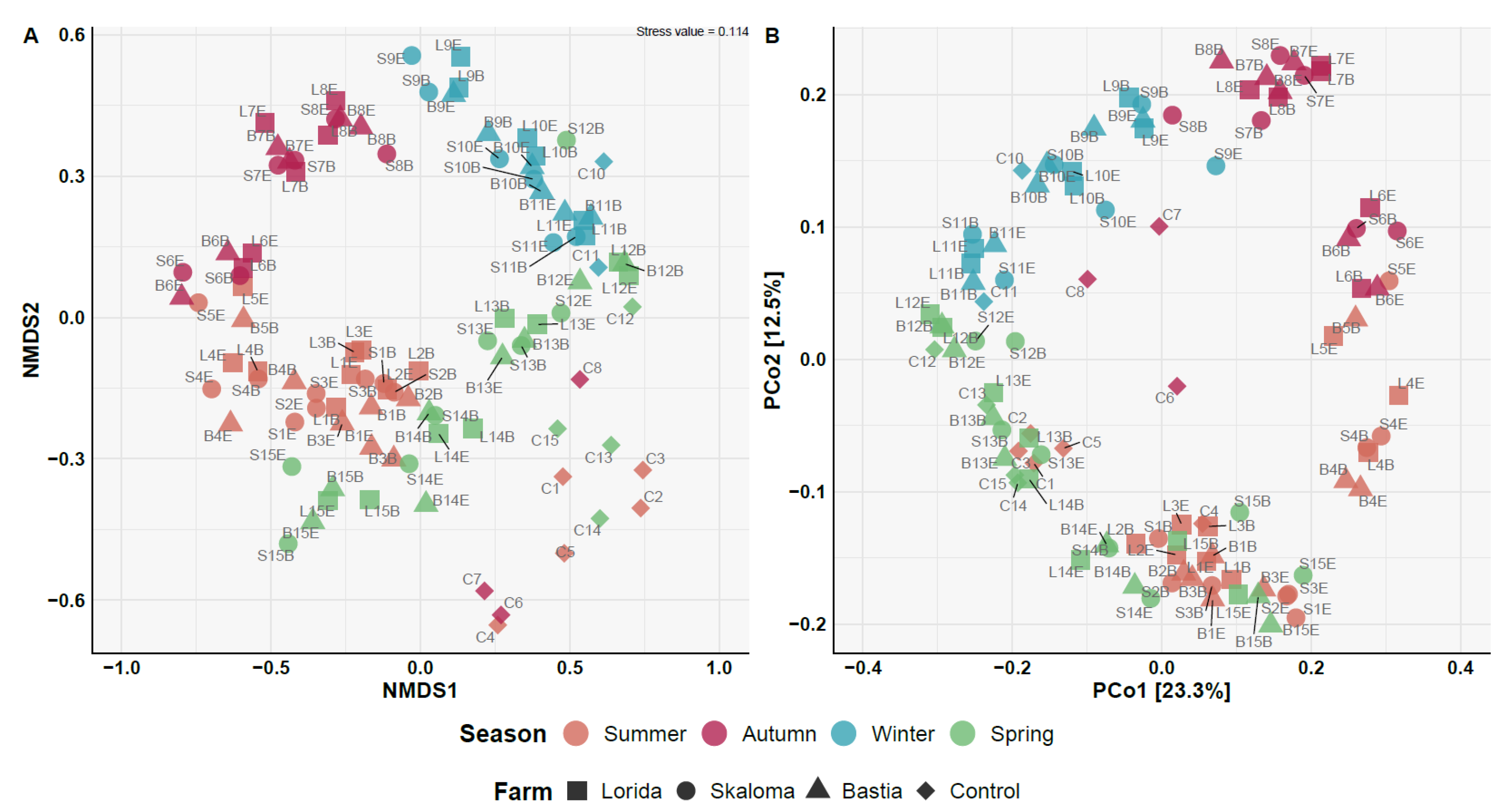
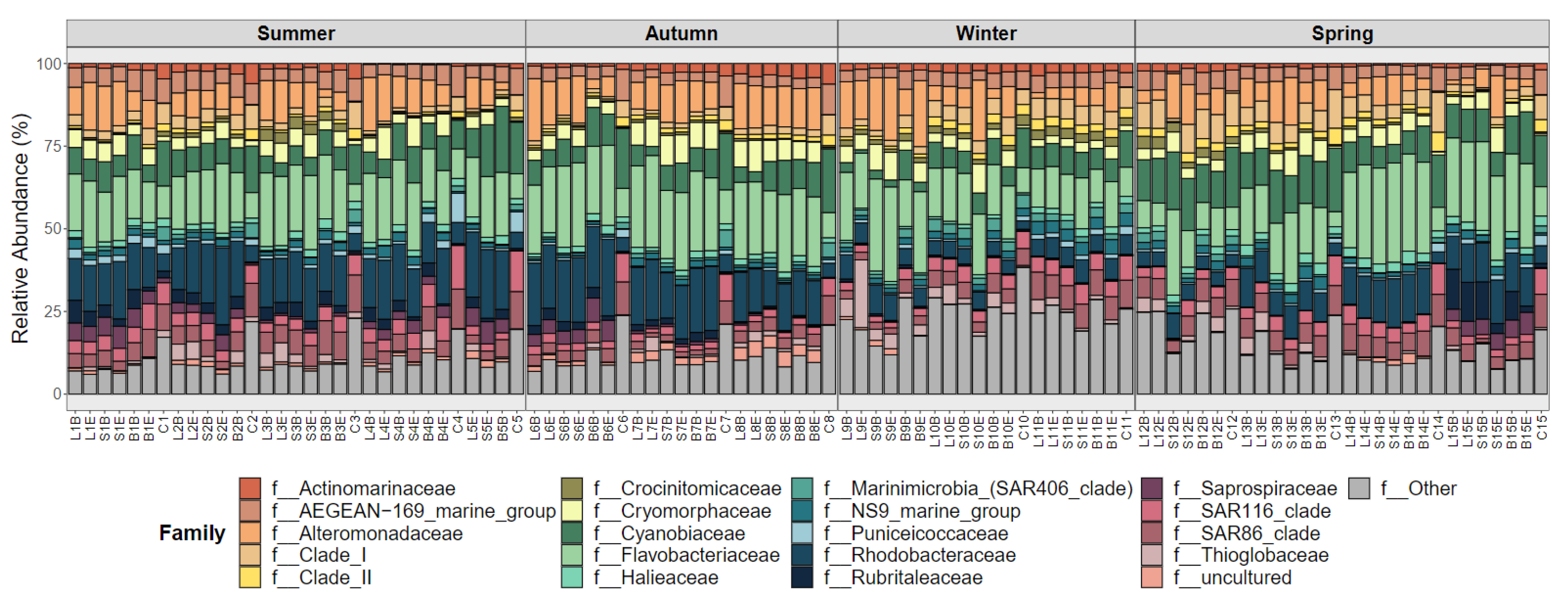
3.3. Beta Diversity Analysis and Bacterial Community Composition
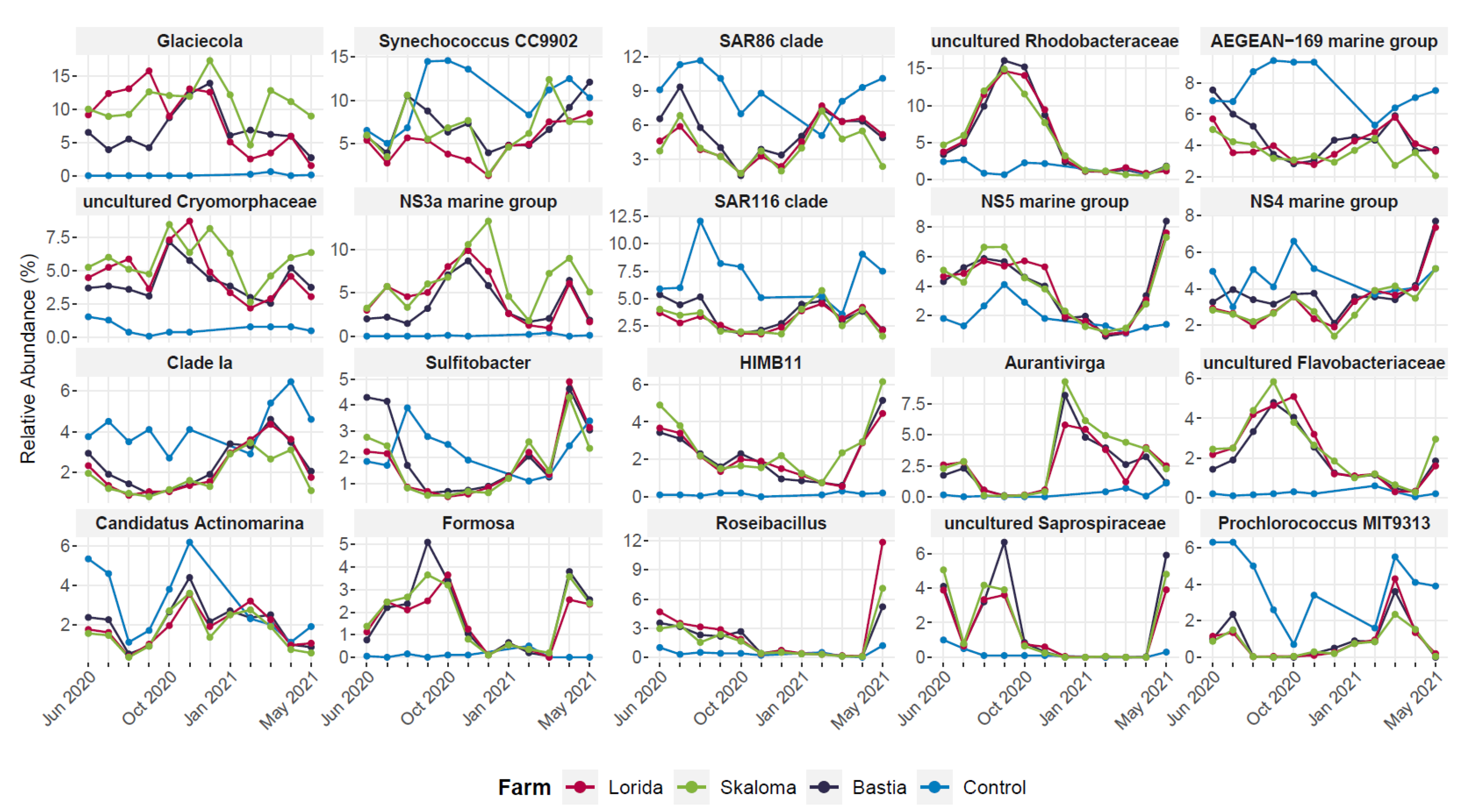
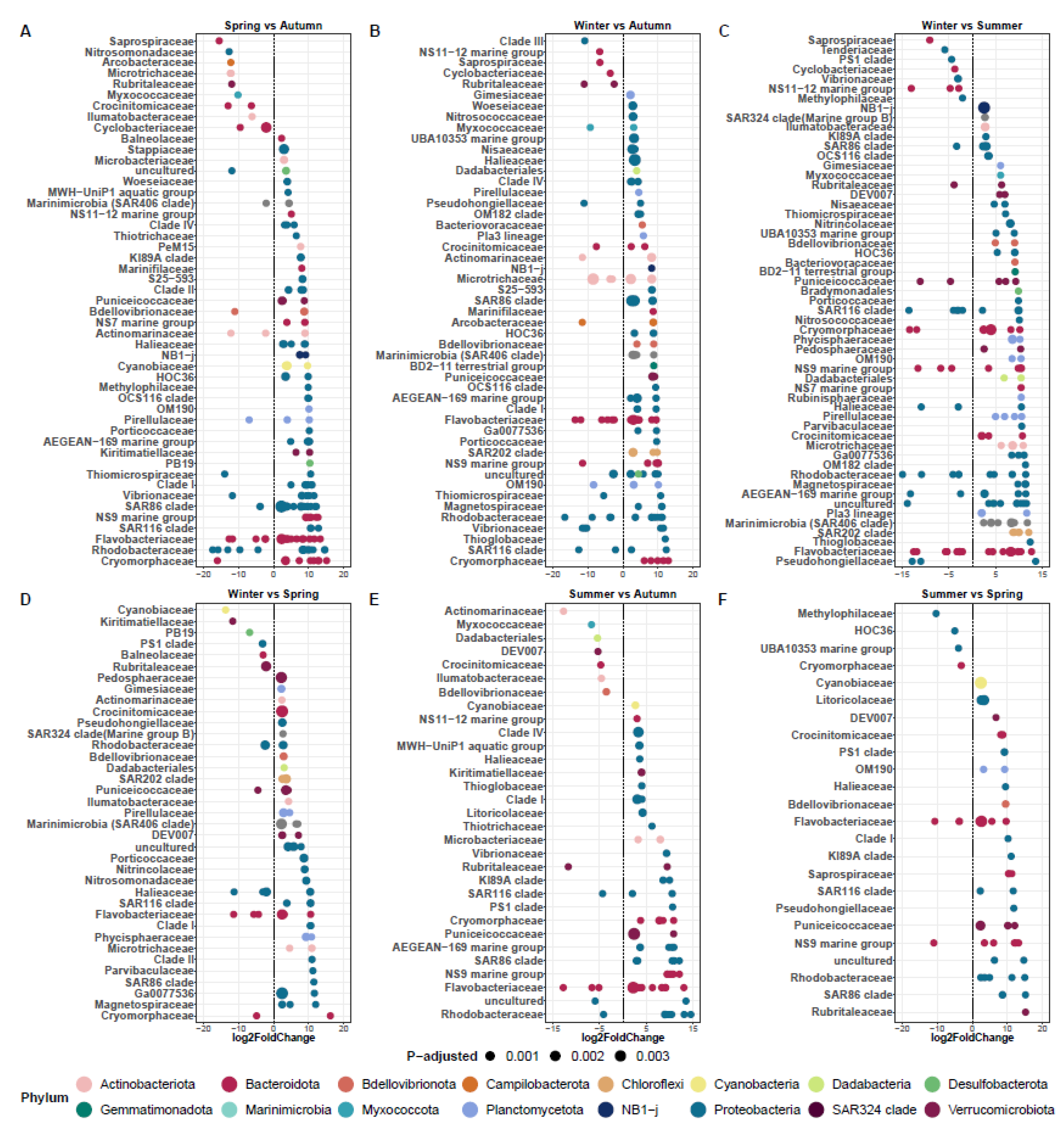
3.4. Seasonal Patterns of Differentially Abundant Taxa in the Marine Aquaculture Systems
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arrigo, K.R. Molecular diversity and ecology of microbial plankton. Nature 2005, 437, 343–348. [Google Scholar] [CrossRef]
- Morris, R.; Nunn, B.; Frazar, C.; Goodlett, D.R.; Ting, Y.S.; Rocap, G. Comparative metaproteomics reveals ocean-scale shifts in microbial nutrient utilization and energy transduction. ISME J. 2010, 4, 673–685. [Google Scholar] [CrossRef] [Green Version]
- Acinas, S.G.; Sánchez, P.; Salazar, G.; Cornejo-Castillo, F.M.; Sebastián, M.; Logares, R.; Royo-Llonch, M.; Paoli, L.; Sunagawa, S.; Hingamp, P.; et al. Deep ocean metagenomes provide insight into the metabolic architecture of bathypelagic microbial communities. Commun. Biol. 2021, 4, 604. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Vogt, M.; Elizondo, U.H.; Righetti, D.; Zimmermann, N.E.; Gruber, N. Major restructuring of marine plankton assemblages under global warming. Nat. Commun. 2021, 12, 5226. [Google Scholar] [CrossRef] [PubMed]
- Staley, J.T.; Konopka, A.; Staley Allan, J.T.K. Microorganisms in Aquatic and Terrestrial Habitats. Annu. Rev. Microbiol. 1985, 39, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Steen, A.D.; Crits-Christoph, A.; Carini, P.; DeAngelis, K.M.; Fierer, N.; Lloyd, K.G.; Cameron Thrash, J. High proportions of bacteria and archaea across most biomes remain uncultured. ISME J. 2019, 13, 3126–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilborn, R.; Amoroso, R.O.; Anderson, C.M.; Baum, J.K.; Branch, T.A.; Costello, C.; De Moor, C.L.; Faraj, A.; Hively, D.; Jensen, O.P.; et al. Effective fisheries management instrumental in improving fish stock status. Proc. Natl. Acad. Sci. USA 2020, 117, 2218–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melnychuk, M.C.; Kurota, H.; Mace, P.M.; Pons, M.; Minto, C.; Osio, G.C.; Jensen, O.P.; de Moor, C.L.; Parma, A.M.; Richard Little, L.; et al. Identifying management actions that promote sustainable fisheries. Nat. Sustain. 2021, 4, 440–449. [Google Scholar] [CrossRef]
- Tsikliras, A.C.; Dinouli, A.; Tsiros, V.Z.; Tsalkou, E. The Mediterranean and Black Sea fisheries at risk from overexploitation. PLoS ONE 2015, 10, e0121188. [Google Scholar] [CrossRef] [Green Version]
- Froese, R.; Winker, H.; Coro, G.; Demirel, N.; Tsikliras, A.C.; Dimarchopoulou, D.; Scarcella, G.; Quaas, M.; Matz-Lück, N. Status and rebuilding of European fisheries. Mar. Policy 2018, 93, 159–170. [Google Scholar] [CrossRef]
- United Nations. Transforming Our World: The 2030 Agenda for Sustainable Development. In A New Era in Global Health; Springer Publishing Company: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Wang, Z.; Tsementzi, D.; Williams, T.C.; Juarez, D.L.; Blinebry, S.K.; Garcia, N.S.; Sienkiewicz, B.K.; Konstantinidis, K.T.; Johnson, Z.I.; Hunt, D.E. Environmental stability impacts the differential sensitivity of marine microbiomes to increases in temperature and acidity. ISME J. 2021, 15, 19–28. [Google Scholar] [CrossRef]
- Nikouli, E.; Meziti, A.; Antonopoulou, E.; Mente, E.; Kormas, K.A. Host-Associated Bacterial Succession during the Early Embryonic Stages and First Feeding in Farmed Gilthead Sea Bream (Sparus aurata). Genes 2019, 10, 483. [Google Scholar] [CrossRef] [Green Version]
- Yajima, D.; Fujita, H.; Hayashi, I.; Shima, G.; Suzuki, K.; Toju, H. Core species and interactions prominent in fish-associated microbiome dynamics. Microbiome 2023, 11, 53. [Google Scholar] [CrossRef]
- Trainer, V.L.; Moore, S.K.; Hallegraeff, G.; Kudela, R.M.; Clement, A.; Mardones, J.I.; Cochlan, W.P. Pelagic harmful algal blooms and climate change: Lessons from nature’s experiments with extremes. Harmful Algae 2020, 91, 101591. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wang, Y.; Liu, P.; Sun, Y.; Song, Z.; Hu, X. Characteristics of bacterial community structure and function associated with nutrients and heavy metals in coastal aquaculture area. Environ. Pollut. 2021, 275, 116639. [Google Scholar] [CrossRef] [PubMed]
- Jurelevicius, D.; Cotta, S.R.; Montezzi, L.F.; Dias, A.C.F.; Mason, O.U.; Picão, R.C.; Jansson, J.K.; Seldin, L. Enrichment of potential pathogens in marine microbiomes with different degrees of anthropogenic activity. Environ. Pollut. 2021, 268, 115757. [Google Scholar] [CrossRef]
- Uyaguari-Diaz, M.I.; Chan, M.; Chaban, B.L.; Croxen, M.A.; Finke, J.F.; Hill, J.E.; Peabody, M.A.; Van Rossum, T.; Suttle, C.A.; Brinkman, F.S.L.; et al. A comprehensive method for ampliconbased and metagenomic characterization of viruses, bacteria, and eukaryotes in freshwater samples. Microbiome 2016, 4, 20. [Google Scholar] [CrossRef] [Green Version]
- Debroas, D.; Domaizon, I.; Humbert, J.F.; Jardillier, L.; Lepére, C.; Oudart, A.; Taib, N. Overview of freshwater microbial eukaryotes diversity: A first analysis of publicly available metabarcoding data. FEMS Microbiol. Ecol. 2017, 93, fix023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, M.A.A.; Magdy, M. Metabarcoding profiling of microbial diversity associated with trout fish farming. Sci. Rep. 2021, 11, 421. [Google Scholar] [CrossRef]
- Lee, D.E.; Lee, J.; Kim, Y.M.; Myeong, J.I.; Kim, K.H. Uncultured bacterial diversity in a seawater recirculating aquaculture system revealed by 16S rRNA gene amplicon sequencing. J. Microbiol. 2016, 54, 296–304. [Google Scholar] [CrossRef]
- Kalaitzidou, M.P.; Alvanou, M.V.; Papageorgiou, K.V.; Lattos, A.; Sofia, M.; Kritas, S.K.; Petridou, E.; Giantsis, I.A. Pollution Indicators and HAB-Associated Halophilic Bacteria Alongside Harmful Cyanobacteria in the Largest Mussel Cultivation Area in Greece. Int. J. Environ. Res. Public Health 2022, 19, 5285. [Google Scholar] [CrossRef]
- Jing, X.; Gou, H.; Gong, Y.; Ji, Y.; Su, X.; Zhang, J.; Han, M.; Xu, L.; Wang, T. Seasonal dynamics of the coastal bacterioplankton at intensive fish-farming areas of the Yellow Sea, China revealed by high-throughput sequencing. Mar. Pollut. Bull. 2019, 139, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Zan, S.; Ming, H.; Li, M.; Xu, J.; Xie, Y.; Wang, W.; Fan, J. 16S Next-generation sequencing and quantitative PCR reveal the distribution of potential pathogens in the Liaohe Estuary. Front. Mar. Sci. 2022, 9, 1044260. [Google Scholar] [CrossRef]
- Pavloudi, C.; Tsertou, M.I.; Antonopoulou, E.; Katharios, P. Investigation of systemic granulomatosis in cultured meagre, Argyrosomus regius, using clinical metagenomics. Aquaculture 2023, 567, 739249. [Google Scholar] [CrossRef]
- Stavrakidis-Zachou, O.; Lika, K.; Anastasiadis, P.; Papandroulakis, N. Projecting climate change impacts on Mediterranean finfish production: A case study in Greece. Clim. Change 2021, 165, 67. [Google Scholar] [CrossRef]
- Cascarano, M.C.; Stavrakidis-Zachou, O.; Mladineo, I.; Thompson, K.D.; Papandroulakis, N.; Katharios, P. Mediterranean aquaculture in a changing climate: Temperature effects on pathogens and diseases of three farmed fish species. Pathogens 2021, 10, 1205. [Google Scholar] [CrossRef]
- Katselis, G.; Tsolakos, K.; Theodorou, J.A. Mapping of Greek Marine Finfish Farms and Their Potential Impact on the Marine Environment. J. Mar. Sci. Eng. 2022, 10, 286. [Google Scholar] [CrossRef]
- HAPO. Hellenic Organization of Aquaculture Producers—Aquaculture Annual Report; HAPO: Peania, Greece, 2022; Available online: https://fishfromgreece.com/wp-content/flipbook/nov22/ (accessed on 28 June 2023).
- Eurostat European Commission. Agriculture, Forestry and Fishery Statistics: 2020 Edition; Publications Office of the European Union: Luxembourg, 2020; ISBN 978-92-76-21521-9. [Google Scholar]
- Konstantinidis, E.; Perdikaris, C.; Batzios, C.; Michaelidis, B.; Ganias, K. Estimating cage farming capacity of data-poor mariculture sectors in the Eastern Mediterranean. J. Appl. Aquac. 2021, 33, 111–125. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. A Manual of Chemical and Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984. [Google Scholar] [CrossRef]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 20th ed.; American Public Health Association, American Water Works Association and Water Environmental Federation: Washington, DC, USA, 1998. [Google Scholar]
- Cruaud, P.; Vigneron, A.; Fradette, M.S.; Charette, S.J.; Rodriguez, M.J.; Dorea, C.C.; Culley, A.I. Open the SterivexTM casing: An easy and effective way to improve DNA extraction yields. Limnol. Oceanogr. Methods 2017, 15, 1015–1020. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2020, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing. R 2011. Available online: https://www.r-project.org/ (accessed on 28 June 2023).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A. Ggpubr: “ggplot2” Based Publication Ready Plots. R Package version 0.6.0, 2020. Available online: https://CRAN.R-project.org/package=ggpubr (accessed on 28 June 2023).
- Mikryukov, V. metagMisc: Miscellaneous Functions for Metagenomic Analysis. R Package version 0.5.0, 2020. Available online: https://github.com/vmikk/metagMisc (accessed on 28 June 2023).
- Hsieh, T.; Ma, K.; Chao, A. iNEXT: Interpolation and Extrapolation for Species Diversity. R Package version 3.0.0, 2022. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.S.; Kirkegaard, R.H.; Karst, S.M.; Albertsen, M. ampvis2: An R package to analyse and visualise 16S rRNA amplicon data. bioRxiv 2018, 10–11. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. vegan: Community Ecology Package. R Package version 2.6.4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 28 June 2023).
- Martinez Arbizu, P. pairwiseAdonis: Pairwise Multilevel Comparison Using. R Package version 0.4.1, 2020. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2018, 35, 2084–2092. [Google Scholar] [CrossRef] [Green Version]
- Bouwmeester, M.M.; Goedknegt, M.A.; Poulin, R.; Thieltges, D.W. Collateral diseases: Aquaculture impacts on wildlife infections. J. Appl. Ecol. 2021, 58, 453–464. [Google Scholar] [CrossRef]
- Zhao, J.L.; Liu, Y.S.; Liu, W.R.; Jiang, Y.X.; Su, H.C.; Zhang, Q.Q.; Chen, X.W.; Yang, Y.Y.; Chen, J.; Liu, S.S.; et al. Tissue-specific bioaccumulation of human and veterinary antibiotics in bile, plasma, liver and muscle tissues of wild fish from a highly urbanized region. Environ. Pollut. 2015, 198, 15–24. [Google Scholar] [CrossRef]
- Morsy, A.; Ebeid, M.; Soliman, A.; Halim, A.A.; Ali, A.; Fahmy, M. Evaluation of the water quality and the eutrophication risk in Mediterranean sea area: A case study of the Port Said Harbour, Egypt. Environ. Chall. 2022, 7, 100484. [Google Scholar] [CrossRef]
- Wang, K.; Wei, Q.; Jian, H.; Gao, Z.; Yao, Q. Variations in the nutrient concentration and composition in Liaodong Bay under long-term human activities. Mar. Pollut. Bull. 2022, 182, 114016. [Google Scholar] [CrossRef]
- Sarà, G. Ecological effects of aquaculture on living and non-living suspended fractions of the water column: A meta-analysis. Water Res. 2007, 41, 3187–3200. [Google Scholar] [CrossRef]
- Bouwman, A.F.; Beusen, A.H.W.; Overbeek, C.C.; Bureau, D.P.; Pawlowski, M.; Glibert, P.M. Hindcasts and future projections of global inland and coastal nitrogen and phosphorus loads due to finfish aquaculture. Rev. Fish. Sci. 2013, 21, 112–156. [Google Scholar] [CrossRef]
- Rubio-Portillo, E.; Villamor, A.; Fernandez-Gonzalez, V.; Antón, J.; Sanchez-Jerez, P. Exploring changes in bacterial communities to assess the influence of fish farming on marine sediments. Aquaculture 2019, 506, 459–464. [Google Scholar] [CrossRef]
- Fodelianakis, S.; Papageorgiou, N.; Pitta, P.; Kasapidis, P.; Karakassis, I.; Ladoukakis, E.D. The pattern of change in the abundances of specific bacterioplankton groups is consistent across different nutrient-enriched habitats in crete. Appl. Environ. Microbiol. 2014, 80, 3784–3792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalimeris, A.; Kassis, D. Sea surface circulation variability in the Ionian-Adriatic Seas. Prog. Oceanogr. 2020, 189, 102454. [Google Scholar] [CrossRef]
- Reich, I.; Ijaz, U.Z.; Gormally, M.; Smith, C.J. 16S rRNA sequencing reveals likely beneficial core microbes within faecal samples of the EU protected slug Geomalacus maculosus. Sci. Rep. 2018, 8, 10402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, C.; Salvador, S.; Arrojado, C.; Silva, Y.; Santos, A.L.; Cunha, Â.; Gomes, N.; Almeida, A. Evaluating seasonal dynamics of bacterial communities in marine fish aquaculture: A preliminary study before applying phage therapy. J. Environ. Monit. 2011, 13, 1053–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolda, A.; Gavrilović, A.; Jug-Dujaković, J.; Ljubešić, Z.; El-Matbouli, M.; Lillehaug, A.; Lončarević, S.; Perić, L.; Knežević, D.; Vukić Lušić, D.; et al. Profiling of bacterial assemblages in the marine cage farm environment, with implications on fish, human and ecosystem health. Ecol. Indic. 2020, 118, 106785. [Google Scholar] [CrossRef]
- Deng, M.; Hou, J.; Song, K.; Chen, J.; Gou, J.; Li, D.; He, X. Community metagenomic assembly reveals microbes that contribute to the vertical stratification of nitrogen cycling in an aquaculture pond. Aquaculture 2020, 520, 734911. [Google Scholar] [CrossRef]
- Fuchsman, C.A.; Palevsky, H.I.; Widner, B.; Duffy, M.; Carlson, M.C.G.; Neibauer, J.A.; Mulholland, M.R.; Keil, R.G.; Devol, A.H.; Rocap, G. Cyanobacteria and cyanophage contributions to carbon and nitrogen cycling in an oligotrophic oxygen-deficient zone. ISME J. 2019, 13, 2714–2726. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, M.; Chen, X.; Zhai, W.; Tan, E.; Tang, K. Transcriptomic evidences for microbial carbon and nitrogen cycles in the deoxygenated seawaters of Bohai Sea. Environ. Int. 2022, 158, 106889. [Google Scholar] [CrossRef]
- Fernández-Gómez, B.; Richter, M.; Schüler, M.; Pinhassi, J.; Acinas, S.G.; González, J.M.; Pedrós-Alió, C. Ecology of marine bacteroidetes: A comparative genomics approach. ISME J. 2013, 7, 1026–1037. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Stevens, S.L.R.; Chan, L.-K.; Bertilsson, S.; Glavina del Rio, T.; Tringe, S.G.; Malmstrom, R.R.; Mcmahon, K.D. Ecophysiology of Freshwater Verrucomicrobia Inferred from. mSphere 2017, 2, e00277-17. [Google Scholar] [CrossRef] [Green Version]
- Palladino, G.; Rampelli, S.; Scicchitano, D.; Musella, M.; Quero, G.M.; Prada, F.; Mancuso, A.; Seyfarth, A.M.; Turroni, S.; Candela, M.; et al. Impact of marine aquaculture on the microbiome associated with nearby holobionts: The case of patella caerulea living in proximity of sea bream aquaculture cages. Microorganisms 2021, 9, 455. [Google Scholar] [CrossRef]
- Tsikopoulou, I.; Lampa, M.; Tsiola, A.; Pitta, P.; Tsapakis, M.; Karakassis, I. Functional adaptations of benthic communities to organic matter enrichment at the edge of an allowable zone of effect (AZE). Estuar. Coast. Shelf Sci. 2021, 262, 107596. [Google Scholar] [CrossRef]
- Quero, G.M.; Ape, F.; Manini, E.; Mirto, S.; Luna, G.M. Temporal Changes in Microbial Communities Beneath Fish Farm Sediments Are Related to Organic Enrichment and Fish Biomass Over a Production Cycle. Front. Mar. Sci. 2020, 7, 524. [Google Scholar] [CrossRef]
- Qin, Q.L.; Xie, B.B.; Yu, Y.; Shu, Y.L.; Rong, J.C.; Zhang, Y.J.; Zhao, D.L.; Chen, X.L.; Zhang, X.Y.; Chen, B.; et al. Comparative genomics of the marine bacterial genus Glaciecola reveals the high degree of genomic diversity and genomic characteristic for cold adaptation. Environ. Microbiol. 2014, 16, 1642–1653. [Google Scholar] [CrossRef]
- Bae, S.; Yoo, K. Microplastic contamination and microbial colonization in coastal area of Busan City, Korea. Front. Mar. Sci. 2022, 9, 1030476. [Google Scholar] [CrossRef]
- Sinden, A.; Sinang, S. Cyanobacteria in aquaculture systems: Linking the occurrence, abundance and toxicity with rising temperatures. Int. J. Environ. Sci. Technol. 2016, 13, 2855–2862. [Google Scholar] [CrossRef]
- Gavriilidou, A.; Gutleben, J.; Versluis, D.; Forgiarini, F.; Van Passel, M.W.J.; Ingham, C.J.; Smidt, H.; Sipkema, D. Comparative genomic analysis of Flavobacteriaceae: Insights into carbohydrate metabolism, gliding motility and secondary metabolite biosynthesis. BMC Genom. 2020, 21, 569. [Google Scholar] [CrossRef]
- Kirchman, D.L. The ecology of Cytophaga-Flavobacteria in aquatic environments. FEMS Microbiol. Ecol. 2002, 39, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Habib, C.; Houel, A.; Lunazzi, A.; Bernardet, J.F.; Olsen, A.B.; Nilsen, H.; Toranzo, A.E.; Castro, N.; Nicolas, P.; Duchaud, E. Multilocus sequence analysis of the marine bacterial genus Tenacibaculum suggests parallel evolution of fish pathogenicity and endemic colonization of aquaculture systems. Appl. Environ. Microbiol. 2014, 80, 5503–5514. [Google Scholar] [CrossRef] [Green Version]
- Bowman, J.P. Out from the Shadows—Resolution of the Taxonomy of the Family Cryomorphaceae. Front. Microbiol. 2020, 11, 795. [Google Scholar] [CrossRef] [PubMed]
- Pohlner, M.; Dlugosch, L.; Wemheuer, B.; Mills, H.; Engelen, B.; Reese, B.K. The majority of active Rhodobacteraceae in marine sediments belong to uncultured genera: A molecular approach to link their distribution to environmental conditions. Front. Microbiol. 2019, 10, 659. [Google Scholar] [CrossRef] [PubMed]
- Roda-Garcia, J.J.; Haro-Moreno, J.M.; Huschet, L.A.; Rodriguez-Valera, F.; López-Pérez, M. Phylogenomics of SAR116 Clade Reveals Two Subclades with Different Evolutionary Trajectories and an Important Role in the Ocean Sulfur Cycle. mSystems 2021, 6, e00944-21. [Google Scholar] [CrossRef] [PubMed]
- Asami, H.; Aida, M.; Watanabe, K. Accelerated sulfur cycle in coastal marine sediment beneath areas of intensive shellfish aquaculture. Appl. Environ. Microbiol. 2005, 71, 2925–2933. [Google Scholar] [CrossRef] [Green Version]
- Needham, D.M.; Fichot, E.B.; Wang, E.; Berdjeb, L.; Cram, J.A.; Fichot, C.G.; Fuhrman, J.A. Dynamics and interactions of highly resolved marine plankton via automated high-frequency sampling. ISME J. 2018, 12, 2417–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, S.A.M.; Shedid, E.S.; Saied, E.M.; Jassbi, A.R.; Jamebozorgi, F.H.; Rateb, M.E.; Du, M.; Abdel-Daim, M.M.; Kai, G.Y.; Al-Hammady, M.A.M.; et al. Cyanobacteria—From the oceans to the potential biotechnological and biomedical applications. Mar. Drugs 2021, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Eshetu, F.; Faassen, E.J.; Kosten, S.; Huszar, V.L.M. Comparison of cyanobacterial and green algal growth rates at different temperatures. Freshw. Biol. 2013, 58, 552–559. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [Green Version]
- Santic, N.; Solic, M.; Kuspilic, G.; Kuspilic, D. Distribution of Synechococcus and Prochlorococcus in the central Adriatic Sea. Acta Adriat. 2011, 52, 101–114. [Google Scholar]
- Makris, C.; Galiatsatou, P.; Tolika, K.; Anagnostopoulou, C.; Kombiadou, K.; Prinos, P.; Velikou, K.; Kapelonis, Z.; Tragou, E.; Androulidakis, Y.; et al. Climate change effects on the marine characteristics of the Aegean and Ionian Seas. Ocean Dyn. 2016, 66, 1603–1635. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | Temperature (°C) | Salinity (psu) | NO3− (μM) | NO2− (μM) | NH4+ (μM) | Chl-a (μg/L) |
|---|---|---|---|---|---|---|
| Summer | 20.77 ± 0.32 | 38.80 ± 0.01 | 5.41 ± 2.91 | 0.12 ± 0.03 | 2.66 ± 0.56 | 0.66 ± 0.03 |
| Autumn | 22.18 ± 0.10 | 38.82 ± 0.01 | 2.22 ± 0.89 | 0.15 ± 0.03 | 2.89 ± 1.11 | 1.53 ± 0.46 |
| Winter | 16.62 ± 0.07 | 38.04 ± 0.08 | 1.28 ± 0.52 | 0.14 ± 0.02 | 3.05 ± 0.49 | 0.38 ± 0.07 |
| Spring | 16.59 ± 0.09 | 38.49 ± 0.04 | 1.14 ± 0.78 | 0.10 ± 0.01 | 1.24 ± 0.55 | 0.88 ± 0.11 |
| ANOVA | p < 0.05 | p < 0.05 | p < 0.05 | p > 0.05 | p > 0.05 | p < 0.05 |
| Season | Temperature (°C) | Salinity (psu) | NO3− (μM) | NO2− (μM) | NH4+ (μM) | Chl-a (μg/L) |
|---|---|---|---|---|---|---|
| Summer | 19.87 | 38.86 | 3.45 | 0.13 | 1.08 | 0.09 |
| Autumn | 21.71 | 38.91 | 0.87 | 0.06 | 1.22 | 0.15 |
| Winter | 16.77 | 38.52 | 2.12 | 0.09 | 2.63 | 0.25 |
| Spring | 16.44 | 38.65 | 1.25 | 0.06 | 0.42 | 0.39 |
| ANOVA | p < 0.05 | p < 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kyritsi, M.; Tsoureki, A.; Koukaras, K.; Kamidis, N.; Krey, G.; Michailidou, S.; Argiriou, A. Seasonal Dynamics of Marine Bacterial Communities in Aquaculture Farms: The Case of the Northern Ionian Coastal Ecosystem (Mediterranean Sea). J. Mar. Sci. Eng. 2023, 11, 1332. https://doi.org/10.3390/jmse11071332
Kyritsi M, Tsoureki A, Koukaras K, Kamidis N, Krey G, Michailidou S, Argiriou A. Seasonal Dynamics of Marine Bacterial Communities in Aquaculture Farms: The Case of the Northern Ionian Coastal Ecosystem (Mediterranean Sea). Journal of Marine Science and Engineering. 2023; 11(7):1332. https://doi.org/10.3390/jmse11071332
Chicago/Turabian StyleKyritsi, Maria, Antiopi Tsoureki, Konstantinos Koukaras, Nikolaos Kamidis, Grigorios Krey, Sofia Michailidou, and Anagnostis Argiriou. 2023. "Seasonal Dynamics of Marine Bacterial Communities in Aquaculture Farms: The Case of the Northern Ionian Coastal Ecosystem (Mediterranean Sea)" Journal of Marine Science and Engineering 11, no. 7: 1332. https://doi.org/10.3390/jmse11071332