1. Introduction
Coral reefs are one of the most diverse marine environments. These species-rich ecosystems are integral to our well-being as they provide a range of cultural, spiritual, and economic services [
1,
2,
3]. In the last century, local and global anthropogenic stressors to coral reefs have increased, mainly due to human population growth, migration, and urbanization [
4,
5,
6].
The extraction of live corals for research causes another anthropogenic pressure on coral reefs [
7]. Since the scientific collection of corals is recognized as a damaging practice, most countries require scientists to acquire special permits to collect corals. During 2019, researchers in the Gulf of Eilat (Israel) requested permission to sample 355 whole coral colonies and 1208 coral fragments (personal communication, Dr. Assaf Zevuloni, ecologist of the Gulf of Eilat, Nature and Parks Authority). Researchers collect corals for two main reasons: they conduct studies that are directly related to corals, such as their genetics, breeding, physiology, diseases, etc., or they conduct studies that are indirectly related to corals, such as interactions with reef organisms, water flow, and wave reduction, recreating reef structures, human interactions, and more.
Coral skeletons, the hard calcium carbonate structures left after the death of corals, are also utilized in marine research for study topics such as: the enhancement of coral settlement through the use of optimized settlement tiles [
8]; coral morphology and sedimentation [
9]; upscaling microalgal cultivation by using calcium carbonate skeletons [
10]; hydrodynamics of coral reefs [
11,
12]; evaluating the activeness of potential marine habitats and marine animals’ behavior [
13].
Many factors affect reef organisms’ recruitment of fish in particular to specific corals or shelters. One important factor is the structural complexity of the coral colony, which in turn provides shelter to residing fish [
14,
15]. It has been shown that corals’ structural complexity correlates with reef fishes’ biodiversity [
16]. Additionally, various studies suggest that different fish populations interact differently with various morphological structures of corals [
15,
17,
18]. Morphological structures of branching corals are often characterized by measurements, such as the number of branches, surface area, planar surface area [
15], branch spacing [
19], volume, ecological volume [
20], and fractal dimension [
21]. These morphological characteristics may have different effects on fishes and other animals’ (such as sponges, mollusks, and worms) recruitment and use of the coral [
15,
17,
21,
22,
23]. Nonetheless, we are yet to understand why specific populations prefer to inhabit one morphological structure over another [
18,
24,
25].
In addition to scientific extraction, a much more severe anthropogenic pressure is caused by the harvesting of live corals for the aquarium trade industry [
26,
27]. In 2003, it was estimated that between 1.5 and 2 million households worldwide maintained a marine aquarium as a hobby, with an annual market value of
$200-
$330 million (US) [
28]. More updated data are difficult to obtain [
29,
30,
31], but the magnitude of the market value can be assumed according to other information. Although the tracking of traded marine wildlife is complicated and has limited accuracy [
32,
33], at least several million pieces of live coral were traded annually between 1988 and 2003 [
28,
34].
Three-dimensional printing (3DP) is a form of additive manufacturing technology that fabricates physical objects from model data. During the printing process, models are digitally sliced into 2D layers, then they are built from the base upwards through the deposition of melted material [
13]. The rapid progress in 3DP technology has opened up research opportunities, making it possible to create multiple identical shelters that mimic a natural coral’s structure [
35,
36]. However, the use of 3DP in coral reef studies started only in the last decade. Several groups have already published their first steps in this field, but it will take some time before 3DP becomes an applicable scientific method in reef research [
13,
25,
37,
38,
39].
This study examines the feasibility of using 3DP corals to investigate and understand the behavior of corals associated reef fishes. Particularly, we aim to answer the following questions: Will reef fishes accept and utilize our custom-made 3DP coral models as shelters? If so, will reef fishes show a preference for a specific color as a factor in choosing a shelter? What will reef fishes favor among an artificial printed model, a live coral, or a coral skeleton for shelter? Additionally, and lastly, what specific coral structure characteristics are preferred by specific reef fish species?
2. Methods
This study constituted a variety of experiments and methodological approaches: following the creation of coral replicas, a preliminary test for fish acceptance was conducted; afterward, a laboratory experiment of fishes’ choices of shelter color was conducted, and finally, in situ fishes’ preferences of shelter structure were examined.
2.1. Creating a Coral Replica
A skeleton of a branching, reef-building, Stylophora pistillata coral was used to create a 3D-printed coral model that possessed the natural coral’s exact morphologic structure. A skeleton size of 160 mm in diameter was chosen after observing S. pistillata corals in a reef that was inhabited by fish. The selected skeleton was cleaned of any settling organisms and algae by soaking it in a bleach–water solution (1:1) for one week.
The coral was scanned using the ‘White light’ method [
40] with an Optima™ scanner, an IDS Imaging Development Systems GmbH camera, and the FlexScan™ software. As part of the scanning method, the coral skeleton was spray coated with SKD-S2 by Manga Flux™ (
https://magnaflux.eu/EU/EN/Products/Liquid-Penetrant-Testing/Consumables/SKD-S2.htm, accessed on 20 April 2023), which creates an opaque, yet nonreflecting white coloration that allows for high-contrast scanning. The scanned model’s precision was high, and challenges in scanning the inner branches were resolved using Rapidform™, Rhino 5.0™, Meshmixer™ and ZBrash™ 3D software (for more details see the
Supplementary Material).
Models for the tests were printed using different materials: (1) gypsum covered with a thin layer of epoxy glue, which did not affect the shape, (2) red Poly-Lactic Acid (PLA), (3) yellow PLA, (4) orange PLA, (5) blue PLA, and (6) light brown PLA with 30% wood. Gypsum models were printed using a binder jet printer ZPrinter 350 at a 300 dpi resolution setting. Zprinter does not need a support since it creates the object in a powder bed. The gypsum model was created at a 1:1 scale. PLA Models were printed using Flashforge Creator Pro II using the Flash Print Slicer, with a linear support with an overhang threshold and a pillar size 1.5 mm. The printing settings were PLA type, support enabled, raft enabled, standard resolution, with a layer height of 0.18 mm, a first layer height of 0.27 mm, the perimeter shells set to 2, the top solid layer was set to 3, and the bottom solid layers were set to 3. The fill density was 15%; the fill pattern was a hexagon. We combined the infills every 2 layers. The print speed was 60 mm/s, the travel speed was 60 mm/s, the extruder temperature was 205 °C, and the platform temperature was 55 °C. Due to the mechanical limitations of the printer, the brown PLA model was printed at a scale of 82% of the size of the original skeleton.
The dimensions of the model, before alterations, were L-160.1 mm × W-160.1 mm × H-113.3 mm, with a surface area of 840.3 cm2, a material volume of 214.1 cm3, and a net ecological volume of 926 cm3.
2.2. Fish Acceptance Test
2.2.1. Experimental Setup
The goals of this test were (1) to examine fishes’ acceptance of artificial shelters of different colors and (2) to compare fishes’ acceptance of the artificial models to that of live S. pistillata in the area.
The test was conducted on the west shore of the Gulf of Aqaba in front of the Inter University Institute for Marine Sciences (IUI) (N 29°30′ E 34°55′) for eight months between March and June 2017 and between January and March 2018 (permit nos. 2017/41605 and 2018/41851, Israeli Nature and Parks Authority). The original natural skeleton and three replicas made of yellow PLA, red PLA and gypsum were set next to an artificial structure in front of the IUI, Eilat, Israel, between depths of 11 and 13 m. This location was chosen because of the high biodiversity in that area. In addition, one gypsum replica was set on the sea floor at a six-meter depth in front of the IUI pier. This location was chosen for its relative isolation and distance from the reef, where the nearest coral was three meters away and the closest knoll was ten meters away. The knoll limited the number of options available to the organisms in the area [
41].
On 28 March 2017, the replicas were moved to an artificial structure at a depth of 8–9 m. A brown replica was added at the same location as well. The models were extracted from the water 3 months later on 20 June 2017 and were left to dry.
On 28 January 2018, the test was repeated, with the coral replica staying in the water for another 3 months.
2.2.2. Experimental Procedures
At least once a week, at different times of the day, a group of divers observed the models and their surroundings for 30 min and recorded fish and other animals in or near the models (n = 21 surveys in total). In addition, on four occasions, an action camera was set to capture specific models in time-lapse mode, taking a photograph every minute for three-hour periods.
2.3. Fish Choice of Shelter’s Color
2.3.1. Experimental Fish
Eight adult female P. squamipinnis were collected from the shallow waters (3–10 m deep) in front of the Interuniversity Institute for Marine Sciences, Gulf of Aqaba (Permit No. 2017/41736, Israeli Nature and Parks Authority). Fishes were maintained for three days in an isolated acclimation tank (L-30 cm × W-30 cm × H-40 cm V = 31.5 L) containing no other objects and with running natural ambient seawater. Excluding feeding and daily checkups, the fishes were not disturbed to minimize stress. After three days, the fishes were transferred to a test aquarium in pairs. After each experimental session (20 min), the fishes were moved to a temporary holding aquarium to prevent studying the same fish in the same setting. With the exception of when they were tested, the fishes were fed daily with an excess of live Artemia salina nauplii and maintained under a 10:14 h dark:light regime. Each fish was tested once or twice a day for five days a week for three weeks. At the end of each day, the fishes were left in a resting tank with running seawater and were fed. The tests took place only during the middle part of their natural activity times, i.e., between 08.00 and 16.00. When the experiments ended, all animals were returned to the exact location in the reef from where they had been taken.
2.3.2. Shelters’ Color Design
The experiment was a 2 option test and followed the design of Shashar et al. (2005). An experimental aquarium (L-80 cm × W-30 cm × H-40 cm V-96 L) was fitted with a fish holding chamber (L-15 cm × W-10 cm × H-40 cm V-6 L) in the middle on the longest side of the tank (
Figure 1). The tank was visually isolated with a black screen. An internet protocol camera (Microsoft LifeCam Cinema HD. Microsoft Corporation One Microsoft Way Redmond, WA 98052, USA) provided a full view of the fishes’ movements and was remotely controlled from a distance behind the screen, eliminating the potential effects of human presence throughout the experiments [
42].
Coral shelters were located on each side of the tank, with one centimeter of distance from the edge of the shelter to the wall of the tank. Coral shelters were: (1) yellow—yellow Poly-Lactic Acid (PLA); (2) brown—light brown PLA with 30% wood dust in a light brown color, scaleX0.82; (3) blue—blue PLA; (4) orange—orange PLA; (5) skeleton—the original natural skeleton on which the models were based, which was colored white; (6) live—live S. pistillata coral with structural characteristics as similar to the original as possible, which was colored dark green.
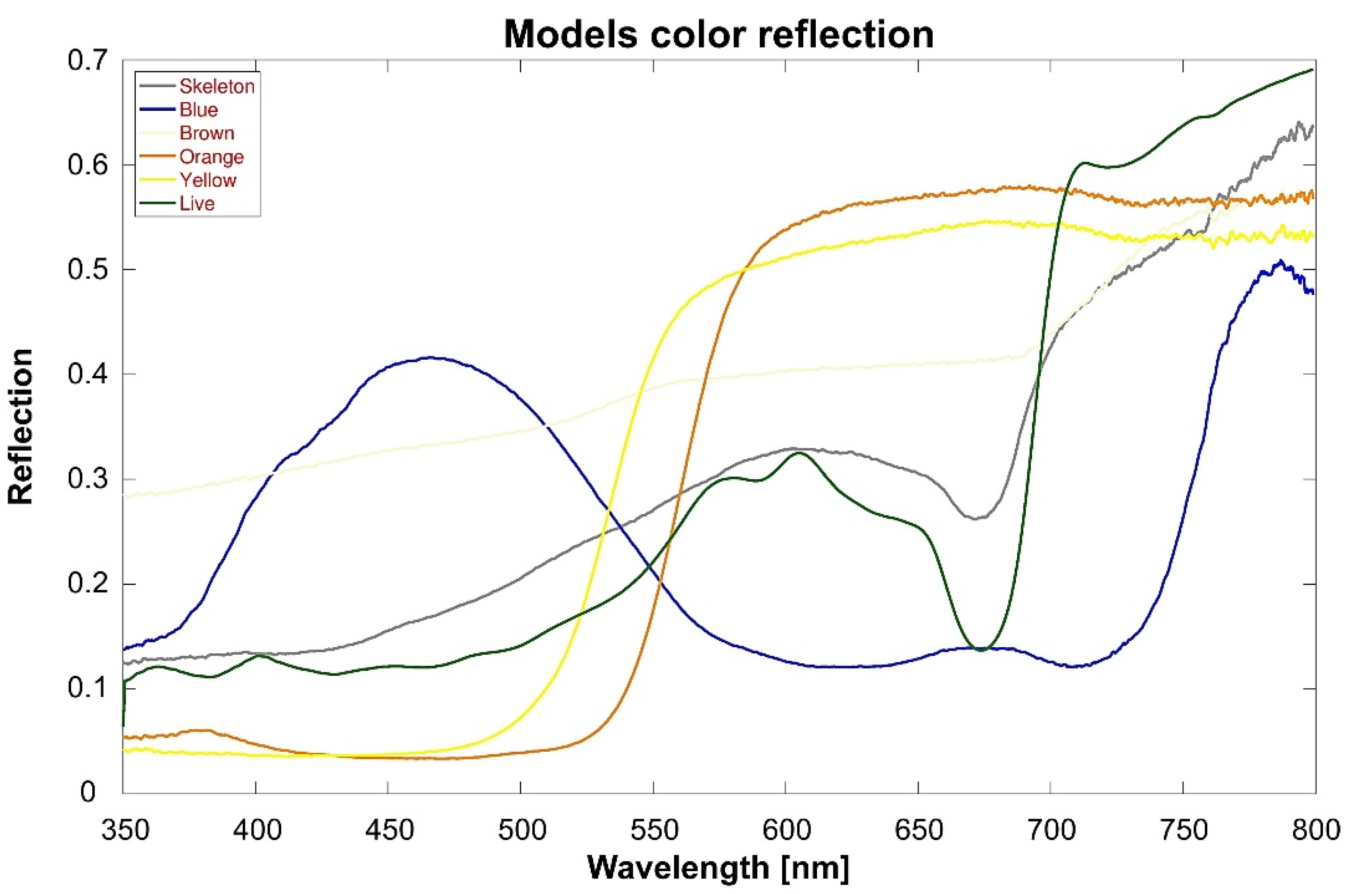
To ascertain the real spectral reflectance and not relay humans’ interpretation of color, the reflectance spectra of the shelters were measured following Stambler and Shashar (2007). A fiber-optics-based calibrated Ocean Optics USB 2000 spectrometer was used to measure the reflectance of the shelters and of a white Teflon standard on a clear (no clouds) day, outdoors, and in the shade. The shelters were set 2 cm from the fiber optic (10 m long; 400 µm in diameter), which was set with a 5° acceptance angle restrictor. All spectra were corrected for baseline electrical noise, and measurements taken in the dark in each experiment were used to subtract additional operational noise. Reflectance was calculated as the ratio between the measurements taken of the corals and those taken of the white standard [
43] (
Figure 2).
During each experiment, two out of the six coral shelters were introduced into the experimental aquarium. The order of the pairs of shelters was randomly chosen until all pair combinations were examined by all fishes.
All experiments were conducted in a laboratory under fluorescent illumination conditions (ca 3.19 µmol/m2/s). Water (unfiltered ambient seawater) in the experimental tank was changed before every experiment (four runs), with the water flow being terminated 10 min before the first run to prevent water motion from affecting the fishes’ behavior.
2.3.3. Experiments on Shelter Color
One of the six shelter options was placed on each side of the experimental tank. Because of P. squamipinnis’ social behavior, fishes were tested in pairs. A pair of fishes was introduced into the center chamber for a 10 min acclimation period. Then, the gate of the chamber was opened using a remote-control lever. In the following 10 min (preliminary behavioral observations suggested that this duration was sufficient for P. squamipinnis to show preference for a shelter), fishes’ movements and positions were recorded on video. At the end of each run, the participating fishes were moved to a separate tank.
2.3.4. Shelter Color—Data Analysis
A total of 30 settings were tested (15 combinations of the 6 shelters, with the locations in the tank switched). Eight fish were tested in each setting, and there was a total of n = 240. Three parameters were analyzed: (a) first choice of a shelter, (b) duration of time spent in/around the shelters, and (c) the number of fishes that preferred each shelter.
For the analysis of the first choice of shelter, a chi-square goodness-of-fit test was used against the null assumption of there being an equal number of choices of each shelter.
To determine the hierarchy of fishes’ preference between the shelters, a chi-square test was conducted between each pair of shelters against a null assumption of there being an equal number of choices. To eliminate potential side effects, both settings of each pair were added (e.g., yellow on the left, and blue on the right, and vice versa) for a total of n = 16.
To analyze the duration of staying in/around the shelters, the data were transformed to a percentage of time in which each fish spent in each shelter. An arcsin transformation was used to meet the assumption of normality and tested with a two-way ANOVA.
Although fishes were tested in pairs, they were treated as performing independent choices: in a sample of five 10 min runs, the fishes were separated from each other (i.e., at opposite sides of the tank) 45.9% (SD = 10%) of the time.
2.4. In Situ Fish Preference of Shelter Structure
In this test, we examined the preference of fishes for specific features of coral structures. For this task, 3DP corals were especially useful since it was possible to specifically manipulate the structure, while leaving all other parameters constant. The structure of the original model was manipulated, and the different shelters were compared to natural coral in the reef. Two manipulations were chosen to be examined: tabular structure [
15] and the scale of coral modification. For each structural change, a gradient of three models was created; for the scale, the models were 100%, 150%, and 200% of the size of the original coral. For the tabular structure, the branches were stretched to create a disk-shaped top with diameters of 19 cm, 22 cm, and 25 cm (ratios to the original structure diameter of 118.75%, 137.5%, and 156.25%, respectively).
2.4.1. Structure Manipulation
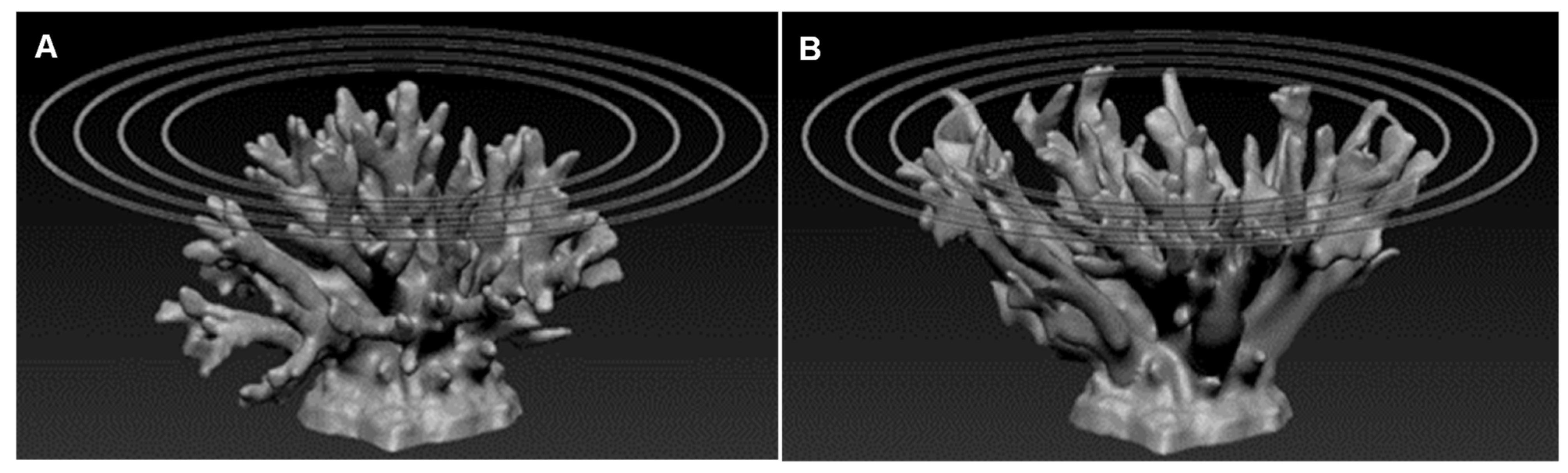
Tabular structure (also known as corymbose [
23])—this series was created by using virtual reference elements built in Rhino 5.0™, with a set of virtual rings in diameters equal to the diameters of the planed circular surface along which the branches were stretched (
Figure 3A). This allowed for the stretching of the upper part of the model alone, while keeping the base constant. Hence, a more open structure with a larger upper “canopy” was created. The diameters were: 19 cm, 22 cm, and 25 cm. After building the rings, the coral model was transferred to ZBrush 4R7™ software, which was used to extend and stretch the branches towards the appropriate reference ring to achieve the final structure (
Figure 3B), while maintaining the branches’ width, i.e., for a tabular structure with a diameter of 25 cm, a ring of 25 cm was used. The final models were transferred to Simplify3D™ V4 software for optimal setting and final preparations to print with a 3D printer Workbench 300 using 3DP™.
Scale manipulating the geometry in this series was achieving by scaling the original model of the coral. The process is straightforward, adding a scale ratio in percentage, and it can be conducted using most 3D modeling software. In this experiment, we used Fusion 360TM.
The coral model was scaled, and three different variations were produced by scaling the model. The model was scaled at 100%, 150%, and 200%. All three variations were then imported into Simplify3D™ V4 software for final preparations and to achieve the correct positioning for 3D printing using a 3DP™ printer.
Different structures and shelters were coded for the ease of the follow-up, as presented in the
Supplementary Material.
2.4.2. Test Preparation
All models were soaked in a tank with running unfiltered seawater that was covered with a shade net to minimize light and prevent macroalgae growth. The models were left in the tank for three weeks to let microalgae, biofilm, and microorganisms settle on them before they were used in the tests.
2.4.3. Experimental Settings
On 03/09/2018, all models were positioned randomly on the area surrounding an artificial reef on a long metal beam, which is part of the structure of the underwater observatory (N29°30′ E34°55′) (
Figure 4) (permit no. 2018/42050, Israeli Nature and Parks Authority). All models were set at a depth of 5 m ± 20 cm, with at least 1 m distance between them. Every month, the models were removed, cleared of any animals in/on them, placed on the sandy bottom (12 m) for the night, and positioned again in a different location the following morning. After 3 weeks of the first round of placements, it was observed that
Chromis viridis were laying eggs on the models. After consideration, it was decided to delay the location change in this round by two weeks to observe this phenomenon. Following that occasion, the models’ location on the beam were changed every month for a total of four changes. After the second round, three coral skeletons and four live corals were marked and used as controls and monitored as well. These were: a skeleton of
Pocillopora sp. (PS), a
Stylophora sp. skeleton (BS), an
Acropora sp. colony that was half-alive but also half-dead or dying (HA), a live
Stylophora sp. (LS), live
Acropora sp. (LA), a large live
Stylophora sp. (27 cm diameter) (LLS), and a large live
Acropora sp. as a representative of the tabular structure (33 cm diameter), which were used to compare them with the printed tabular structures (TSLA).
2.4.4. Experimental Procedure
Models and controls were examined every 2–3 days for a total of at least 11 observations per position change. Forty of the observations were performed during the daytime (08.00–17.00 h), and twenty-one were performed at night. Each model was observed for 1–2 min, and every interaction with the fish population was recorded and filmed if possible. Video filming was conducted with a SJ4000 extreme camera that was used on five separated occasions, each for 2 h recording (total recording time 10 h).
2.4.5. Data analysis
To distinguish between day and night observations, a sign test was used and a Pearson’s Chi-squared test was used to observe the differences between models in species’ preferences. To analyze the differences between the fish communities interacting with the artificial model and the communities interacting with the natural corals, a t-test was used.
Data were analyzed in R version 3.5.1 using RStudio Version 1.0.143.
3. Results
3.1. Preliminary Work
This test was a presence/absence test. The goal was to check if the fishes would accept the artificial models as shelters. In the first two weeks of the test, only day observations were made, and no invertebrates or fishes were observed in any model. Two days after the relocation, a hermit crab was seen between the branches of the yellow model. After a month in which the models were in the water, hermit crabs and brittle stars were seen in all the models except for the isolated gypsum. Only during night observations (25 days after deployment), fishes were spotted within the models for the first time. Later on, when a camera was positioned on the models, fishes were also spotted during the day. In the first set-up, fishes were spotted on the brown and yellow models and the original skeleton. In the brown model, the species seen were Neopomacentrus miryae, Bodianus anthioides, and Chromis viridis. In the yellow model, the fish species observed were N. miryae, Pseudanthias squamipinnis, and Corythoichthys cf schultzi. The fishes that we observed to interact with the natural skeleton were N. miryae and P. squamipinnis. The gypsum model was not interacted with and was removed at the end of the first set.
In the second setting, the observations were focused on fishes only. Fishes were spotted in the brown, yellow, orange, and blue models. The models in which fish were not seen were the red model and the artificial zip wire model. In the skeleton,
N. miryae and
P. squamipinnis were observed. In the yellow model, many
N. miryae and
P. squamipinnis were spotted; at times, three individuals were spotted together. In the brown model, only
N. miryae were seen. In the blue model, only
P. squamipinnis were seen, and in the orange model, the fishes observed were
Meiacanthus nigrolineatus and
N. miryae (
Figure 5).
Fishes observed in the models were usually seen more than once, sometimes even for a whole month. Since none of the fishes were marked or tagged, we do not know whether sightings were of the same individuals or of different specimens.
3.2. Color Choice Results
In this laboratory experiment, on a total of 240 occasions, the fishes were given a choice between two shelters. Out of all runs, on 9 occasions, the fishes did not spend any time in either shelter, while on 36 occasions, the fishes spent time only at one of the shelters, and the rest spent time in both.
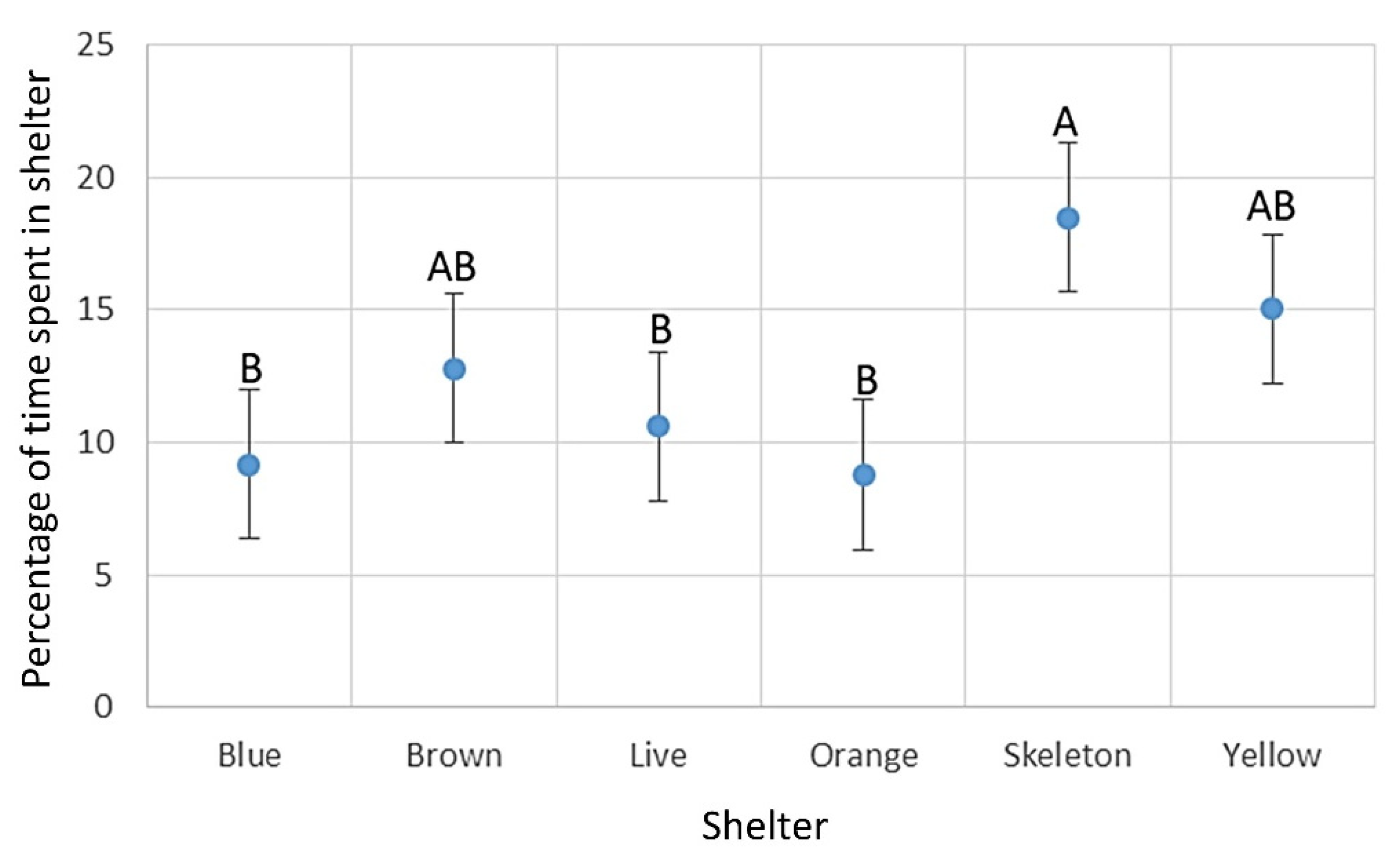
When comparing the total duration of stay in the shelter, fishes showed a significant preference for the skeleton shelter (18.49%) over the blue (9.18%), the live (10.62%), or the orange (8.79%) shelters. The yellow (15.04%) and brown (12.77%) shelters did not show a significant difference from the other shelters (two-way ANOVA; F
5,473 = 3.8544;
p = 0.00196;
Figure 6).
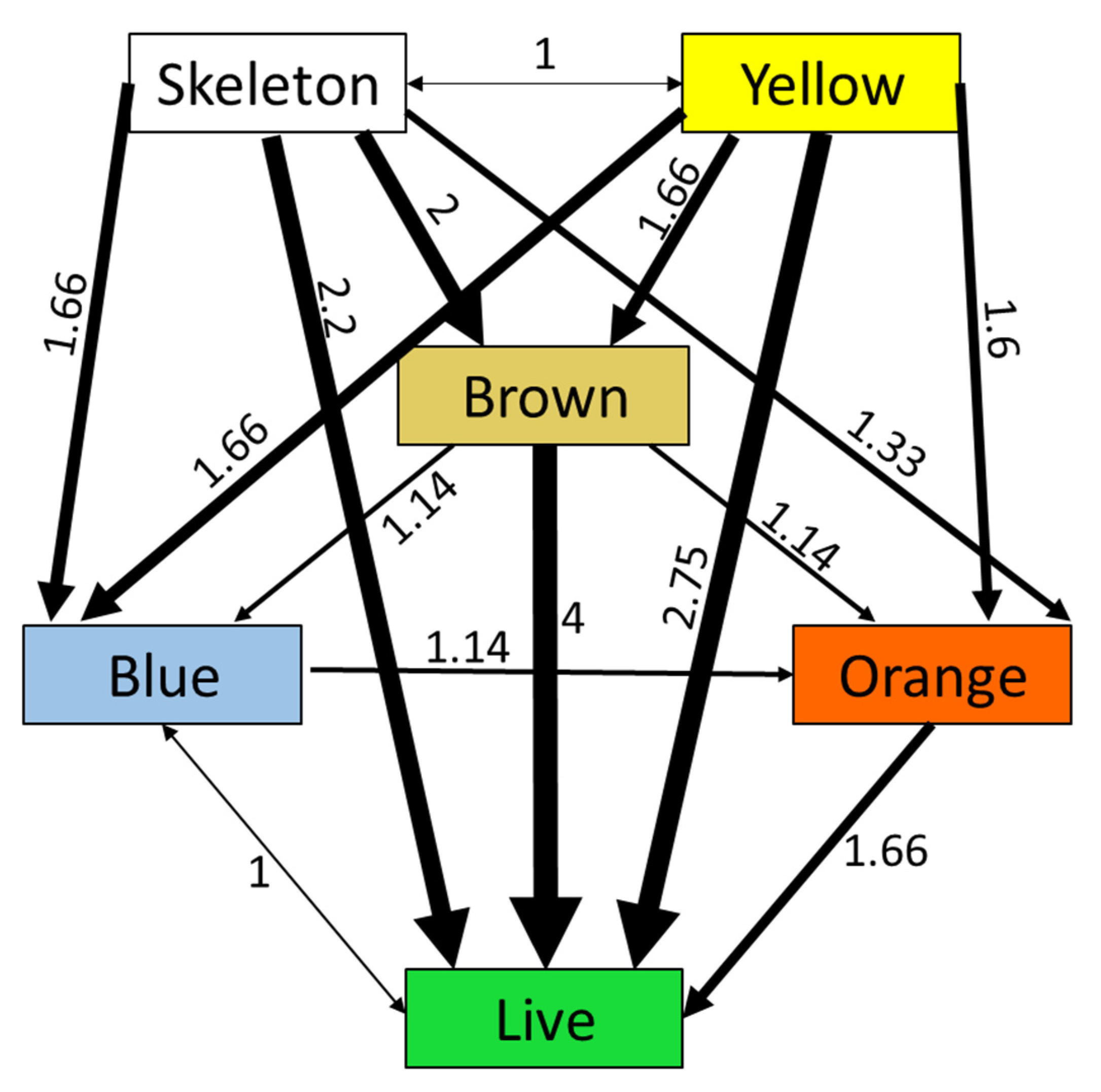
When comparing each fish’s preference as individuals, defined as the shelter in which a specific fish spent most of its time, a somewhat different picture arises (df = 15,
p < 0.001) (
Figure 7). The most preferred shelters were the skeleton and yellow ones (both were preferred 47 times out of 80 presentations). When they are compared against one another, they were each chosen 8 times out of 16 presentations. The least preferred shelter in this comparison was the live coral shelter, which was colored dark green and was chosen 26 times out of 80 presentations.
These results demonstrate that color plays a role in the site preference of fishes.
3.3. Fishes’ Preference for Manipulated Structures
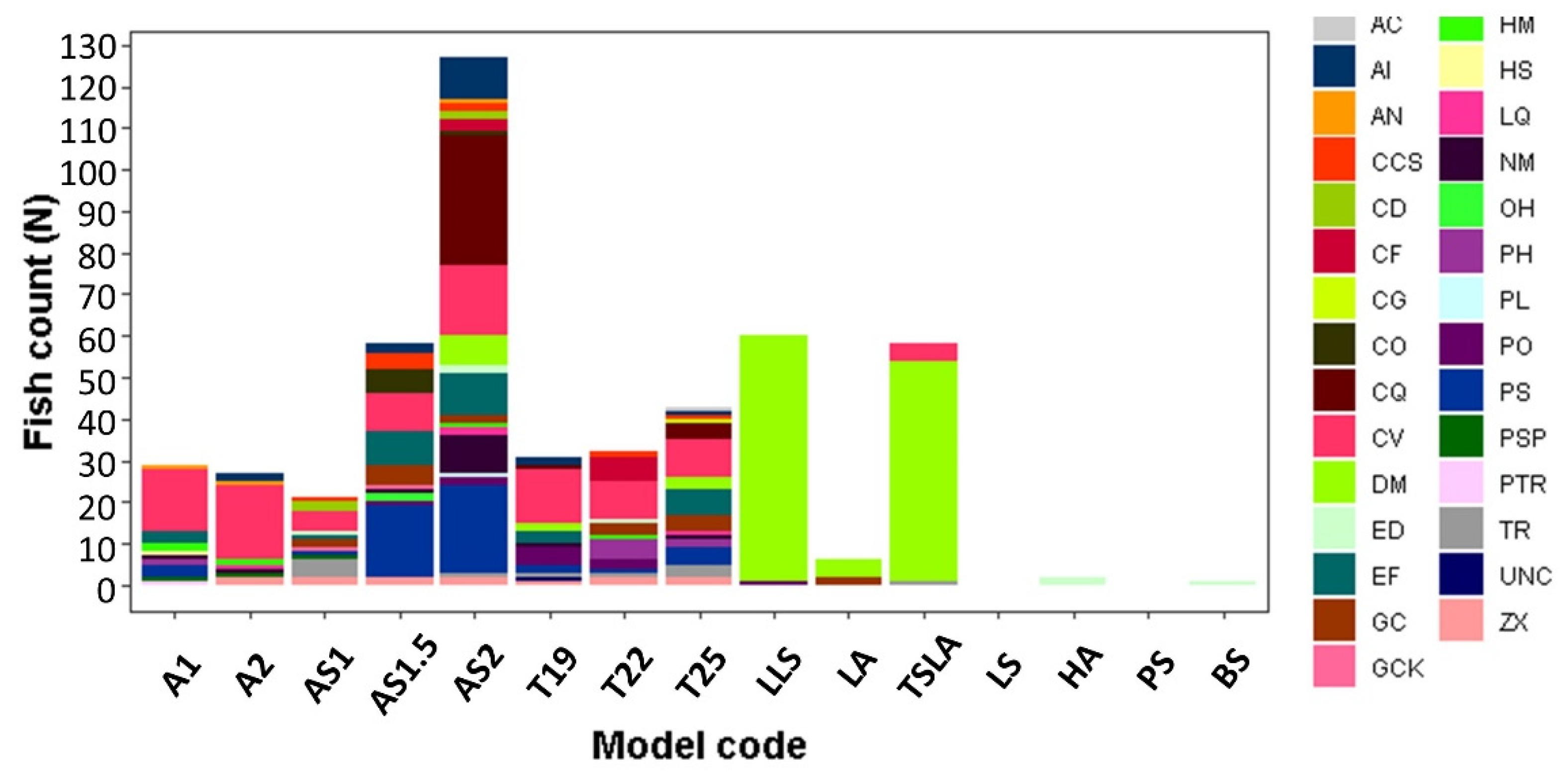
The test was conducted in open water, and in an attempt to cause a minimum amount of disturbance to the environment, fishes were not marked. Hence, it is likely that in many cases we recorded repeated measures with the same fish being recorded on different days. However, since hardly any fishes stayed in any of the shelters during both day and night, we could not ascertain these repeated measurements.
A difference was found in the fish communities that inhabited the different shelters examined (
Figure 8). A larger variety of fish species was recorded in 3DP artificial corals than those in the natural corals, whether they were alive or dead. Although the large natural corals had a large abundance of fishes, most of the fishes observed in these corals were
D. marginatus (
Figure 8).
Fish communities were significantly different between day and night in both abundance and species richness (z = 3.5, p < 0.001). During the day, 92 individuals from 6 different species were observed in the natural shelters, as compared to 306 individuals from 27 species in the artificial ones. During the night, the trend was similar, with 35 individuals (all D. marginatus) in the natural shelters and 63 individuals from 12 different species in the artificial shelters.
Overall, more species and individuals interacted with larger structures than they did with their smaller versions. When comparing the interactions between an artificial structure and a live coral of the same size (T25 vs. TSLA), we found that although the number of interactions was the same, there was a significant difference in the numbers of species (15 in the model and 3 in the live coral).
4. Discussion
This study presents, for the first time, the use of modified and manipulated 3D-printed coral colony structures for the examination of reef fish species’ preferences, as investigated by three different approaches. First, a positive reaction of marine life to several materials (bioplastics in various colors) and the rejection of others (gypsum) were demonstrated. Second, we showed that certain fish species have a preference for a specific shelter colors. Third, manipulating the natural structure of a branched coral model by using various design tools improved the understanding of which structural characteristics are the most appealing to reef fish as shelters.
Artificial 3D-printed coral shelters were meant to provide fishes with the same structure as live corals do, only without the live tissue, which may repel some fish species. However, we could not predict whether or not the fishes would accept this structure/material as a shelter. Additionally, authors of other works, such as Pérez-Pagán and Mercado-Molina (2018), Ruhl and Dixson (2019), and Albalawi et al. (2021), applied a similar method to study fishes’ behavior using a 3D-printed coral colony structure (based on scanned natural corals). Their results were similar: the fishes accepted the artificial structure as shelters. Nonetheless, here we manipulated and modified coral structures to investigate fish species’ preferences for colors and structures. We further present ways of using 3DP corals in science; hence, only examples of the different manipulations and tests performed are presented and discussed, demonstrating the wide scope of this tool.
When checking for fishes’ shelter color preference, our hypotheses were that color plays a part in the reef fishes’ shelter choice and that a given fish species will have a clear preference for a specific color. The results were inconclusive; although the results show that color does matter in making a decision about shelter choice, which color is favored by P. squamipinnis is still not clear. It is possible that individual fishes have individual preferences. Additionally, examining a range of species in different conditions will be beneficial.
Finally, when manipulating the natural structure to emphasize structural characteristics, the hypothesis was that manipulations will affect which fished interact with the structure according to their characteristics. For example, we assumed that bigger spaces between the branches will encourage bigger fish to hide between them, or that a larger structure will simply encourage more fish. The results showed that certain changes did follow our assumptions, and more interactions were observed in the larger structures. As for fishes’ size, no difference was seen. A surprising result was found in the comparison between the live corals and the artificial structures. The number of species interacting with the artificial structures was significantly higher than the number of those in natural corals. A possible explanation for this result is that the live corals were already occupied by Dascyllus marginatus, and different species could not interact with those fishes, while the artificial structures were free of inhabitants, and so, various species could take shelter between their branches. Another possibility is that certain fish species suffer damage from the live coral stinging cells and cannot take shelter inside it, while the artificial structure only provides structural protection and any fishes that fit into it can find a welcoming shelter in it.
Three-dimensional scanning and printing technology is developing exponentially, allowing researchers to conduct tests on imitated natural environments. The natural reef is a multi-level habitat with a variety of substrates, materials, and structures. Every community, and even each individual, might have its own preference for habitat. Creating artificial habitats, which are a good imitation of the natural ones, allowed us to examine defined parameters of the shelters, while keeping others constant.
Notwithstanding, the applicability of 3DP technology still has a few disadvantages. While scanning solutions are available for simple coral colony structures, they lack dexterity for more complicated coral reef structures. Yet, high-resolution 3D scanners (or scanning techniques) are being developed. In the course of this study, the formatting and production of coral models required many attempts and various methods in order to achieve satisfying small coral models. Three-dimensional printing technology presents further challenges that must be addressed before it can be considered as a viable method for creating artificial coral structures. One of the primary concerns is cost. Although 3D printers are becoming more accessible, they are still relatively expensive, making it difficult for many researchers and organizations to invest in the technology. Additionally, the materials used for 3D printing are still being debated, and their long-term impact on the environment must be taken into account. Furthermore, creating large-scale coral structures using 3D printing can be challenging. While there is still much work to be conducted to improve 3D printing technology and its applicability to coral structures, we believe that these challenges can be overcome with continued research and development. We hope that during the course of time and due to improved technologies, using 3DP corals will replace the need for harvesting corals for ornamental use and for research.
We expect that the 3DP of resembling structures will have a broader scope as the technology develops: (A) In the habitat restoration realm, developing 3DP shelters for endangered fish species as nurseries for juvenile fish or as an appealing substrate for fish eggs may be of great interest. (B) As custom-made materials for 3DP are becoming easier to produce, one can examine marine organisms’ preferences and use of different senses in shelter detection and choice making. Here, we studied the use of color, but chemical sensitivity (“smell”), texture sensitivity, and even rheo sensing (sensitivity to water flow and wave reflections) or magneto sensing could be examined. (C) For hydrodynamic inspections in situ and in flow-tanks, 3DP structures may be appropriate substitute models. (D) The use of micro-fragmentation to rapidly create “adult” coral colonies and accelerate reef rehabilitation has been demonstrated for massive corals [
44,
45,
46]; 3DP may produce a complex structure, allowing the application of this approach to branching corals.
For scientific studies, 3DP, together with different Computer Aided-Design software, holds the advantage of being able to fabricate the modified morphology of a single parameter, while keeping others intact. For example, we were able to change only the scale of a printed coral, while maintaining its structure proportions. Further, it is possible to perform artificial parametric gradient adjustment in a controlled and known way. We made these modifications to create a tabular structure out of a basically sphere-shaped coral. Similarly, one can change the branch width, rugosity, or make other unsymmetrical alterations.
Fishes’ responses to 3D coral structures are encouraging for coral replacement in non-scientific applications, such as live coral ornamental aquariums. According to CITES, between 1998 and 2007, a total of 17.83 million pieces of coral and 2.36 million kg of live corals were traded in Southeast Asia [
34]. Advanced 3DP technology can produce suitable replacements for some of these live corals.
With the rapid decline in coral reefs worldwide [
6], researchers and marine protection organizations are desperately pursuing new, efficient approaches to enhance coral reef conservation, rehabilitation and restoration, including using 3DP structures [
10,
13,
25,
38,
39,
47,
48,
49]. The current results demonstrate that utilizing modified and manipulated 3DP coral structures is highly applicable and particularly beneficial for a range of scientific endeavors. Moreover, we hope and believe that 3DP coral models will be useful as substitutes and will decrease the demand for live coral colonies.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







