
Potential Detritivorous Diet of the Invasive Apple Snail (Pomacea canaliculata Lamarck, 1822) in Mangroves: The Relationship between Feeding Indicators and Chemical Characteristics of Decaying Leaf Litter

Abstract
:1. Introduction
2. Materials and Methods
2.1. P. canaliculata and Decaying Leaf Litter
2.2. Growth of P. canaliculata Feeding on Decaying Leaf Litter
2.3. Multiple-Choice Experiment of P. canaliculata on Decaying Leaf Litter
2.4. No-Choice Experiment of P. canaliculata Feeding on Decaying Leaf Litter
2.5. Feeding Indicators of P. canaliculata
2.6. Statistical Analyses
3. Results
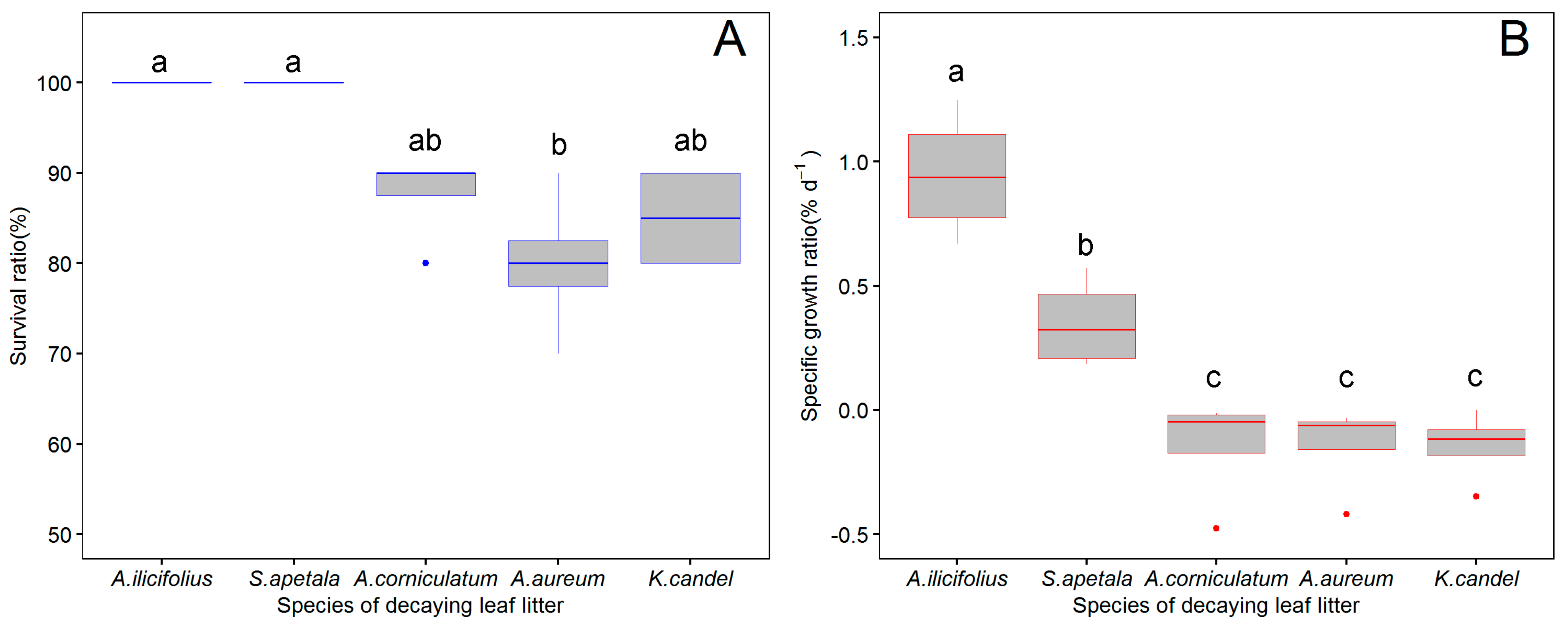
3.1. The Growth of P. canaliculata on Decaying Leaf Litter
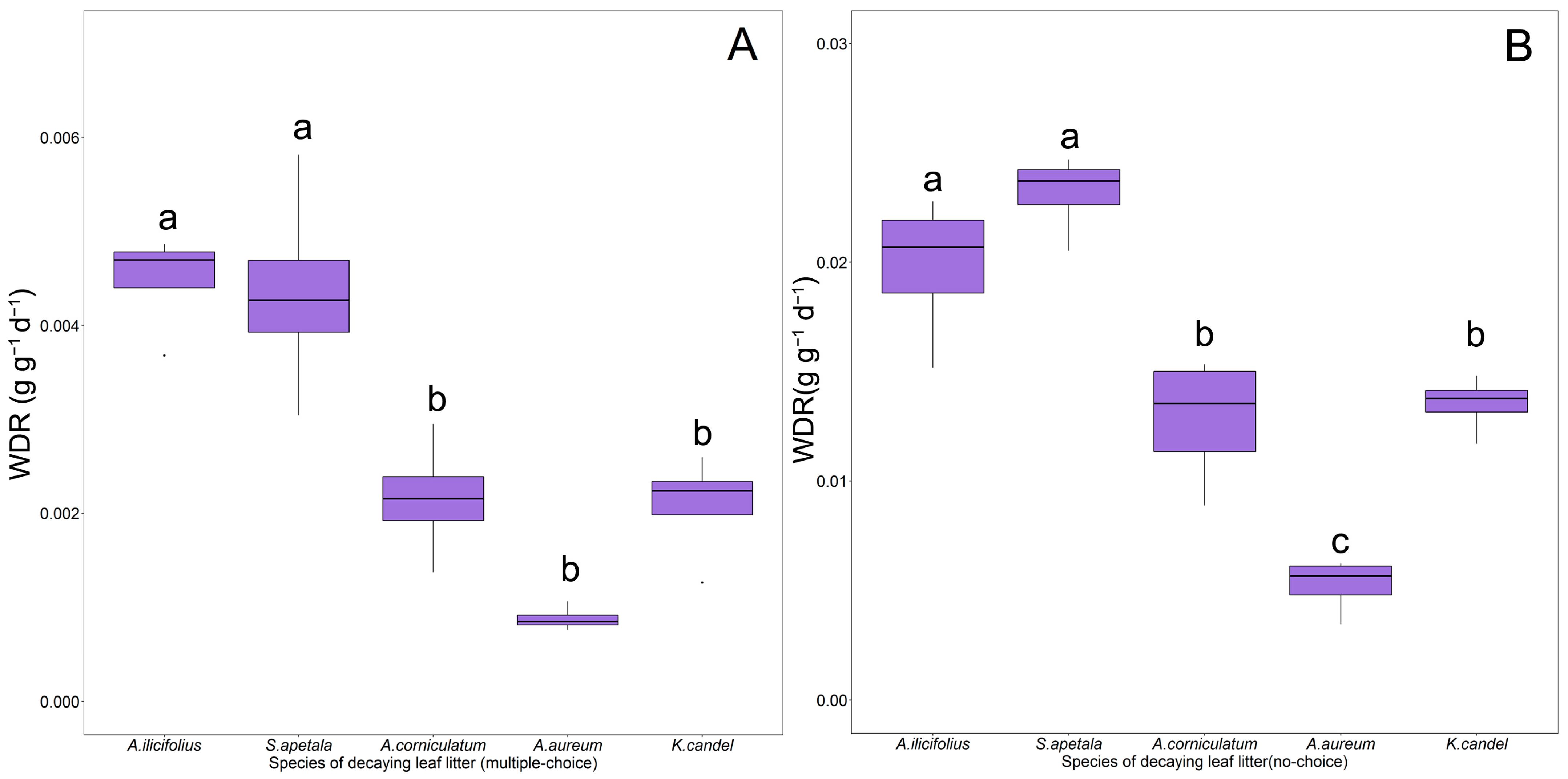
3.2. The Feeding Indicators of P. canaliculata on Mangrove Leaves
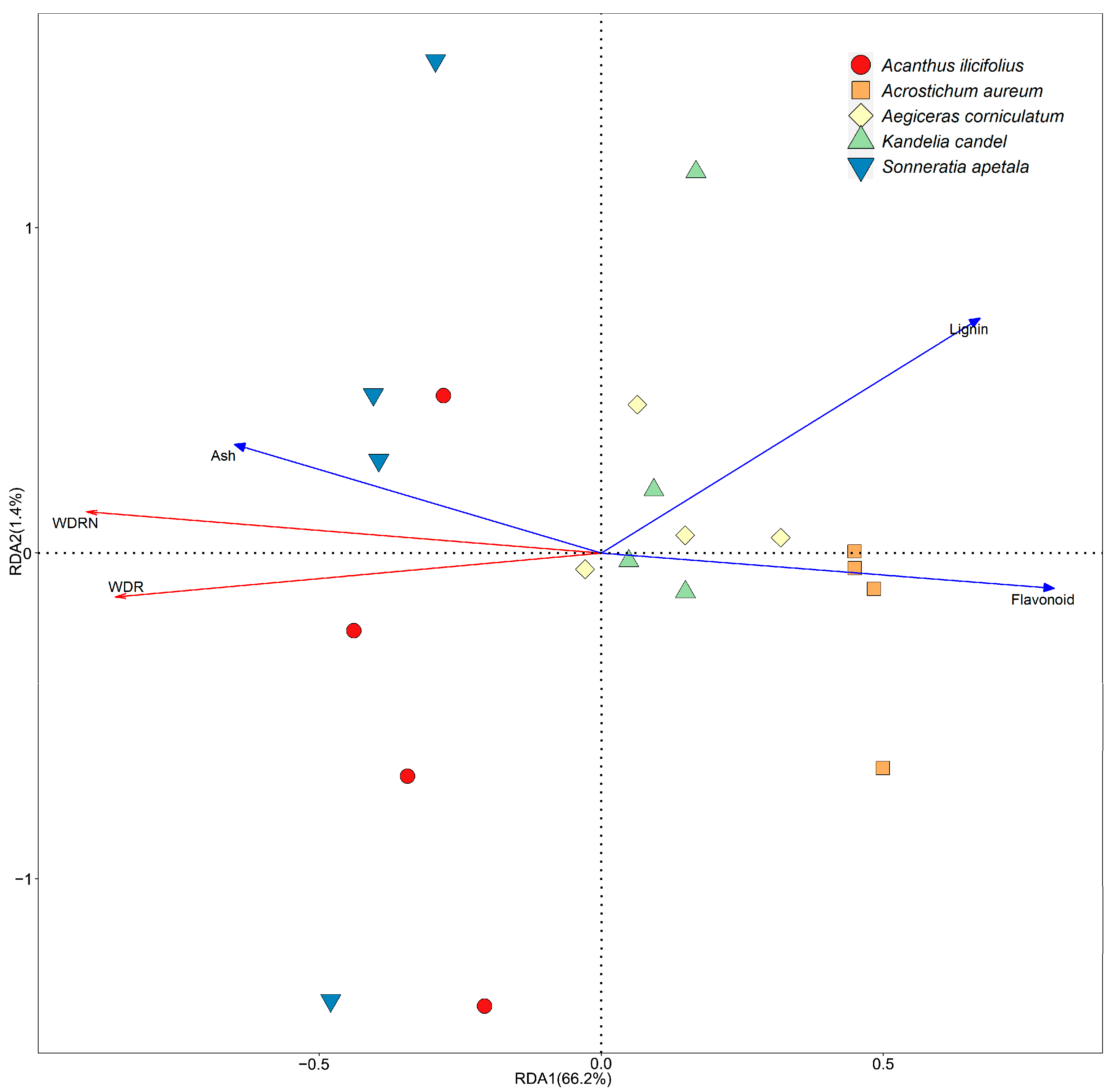
3.3. Correlation Analysis between the Diet and Chemical Characteristics of Decaying Leaf Litter
4. Discussion
4.1. Survival and Growth on Decaying Leaf Litter
4.2. Feeding Preference of P. canaliculata on Decayling Leaf Litter
4.3. Relationship between Feeding Patterns and Characteristics of the Decaying Litter
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruan, L.L.; Yan, M.; Zhang, L.; Fan, X.S.; Yang, H.X. Spatial-temporal NDVI pattern of global mangroves: A growing trend during 2000–2018. Sci. Total Environ. 2022, 844, 157075. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, E.; Valdemarsen, T.; de Moraes, P.C.; Güth, A.Z.; Sumida, P.Y.G.; Quintana, C.O. Anaerobic carbon oxidation in sediment of two Brazilian mangrove forests: The influence of tree roots and crab burrows. Ocean Coast. Res. 2023, 71, e23003. [Google Scholar] [CrossRef]
- Tse, P.; Nip, T.H.M.; Wong, C.K. Nursery function of mangrove: A comparison with mudflat in terms of fish species composition and fish diet. Estuar. Coast. Shelf Sci. 2008, 80, 235–242. [Google Scholar] [CrossRef]
- Matos, C.R.L.; Berredo, J.F.; Machado, W.; Sanders, C.J.; Metzger, E.; Cohen, M.C.L. Carbon and nutrient accumulation in tropical mangrove creeks, Amazon region. Mar. Geol. 2020, 429, 106317. [Google Scholar] [CrossRef]
- Zhu, J.J.; Yan, B. Blue carbon sink function and carbon neutrality potential of mangroves. Sci. Total Environ. 2022, 822, 153438. [Google Scholar] [CrossRef]
- Hochard, J.P.; Hamilton, S.; Barbier, E.B. Mangroves shelter coastal economic activity from cyclones. Proc. Natl. Acad. Sci. USA 2019, 116, 12232–12237. [Google Scholar] [CrossRef] [Green Version]
- Celis-Hernandez, O.; Giron-Garcia, M.P.; Ontiveros-Cuadras, J.F.; Canales-Delgadillo, J.C.; Perez-Ceballos, R.Y.; Ward, R.D.; Acevedo-Gonzales, O.; Armstrong-Altrin, J.S.; Merino-Ibarra, M. Environmental risk of trace elements in mangrove ecosystems: An assessment of natural vs oil and urban inputs. Sci. Total Environ. 2020, 730, 138643. [Google Scholar] [CrossRef]
- Mosquera, E.; Blanco-Libreros, J.F.; Riascos, J.M. Are urban mangroves emerging hotspots of non-indigenous species? A study on the dynamics of macrobenthic fouling communities in fringing red mangrove prop roots. Biol. Invasions 2023, 25, 787–800. [Google Scholar] [CrossRef]
- Faraco, L.F.D.; Ghisi, C.L.; Marins, M.; Ota, S.; Schühli, G.S. Infestation of mangroves by the invasive moth Hyblaea puera (Cramer, 1777)(lepidoptera: Hyblaeidae). Braz. Arch. Biol. Technol. 2019, 62, e19170516. [Google Scholar] [CrossRef]
- Russell, D.J.; Thuesen, P.A.; Thomson, F.E. Reproductive strategies of two invasive tilapia species Oreochromis mossambicus and Tilapia mariae in northern Australia. J. Fish Biol. 2012, 80, 2176–2197. [Google Scholar] [CrossRef]
- Sancho, G.; Kingsley-Smith, P.R.; Morris, J.A.; Toline, C.A.; McDonough, V.; Doty, S.M. Invasive lionfish (Pterois volitans/miles) feeding ecology in Biscayne National Park, Florida, USA. Biol. Invasions 2018, 20, 2343–2361. [Google Scholar] [CrossRef]
- Cao, M.; Cui, L.; Sun, H.; Zhang, X.; Zheng, X.; Jiang, J. Effects of Spartina alterniflora invasion on soil microbial community structure and ecological functions. Microorganisms 2021, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Bigatti, G.; Miloslavich, P.; Penchaszadeh, P.E. Sexual differentiation and size at first maturity of the invasive mussel Perna viridis (Linnaeus, 1758) (mollusca: Mytilidae) at La Restinga Lagoon (Margarita Island, Venezuela). Am. Malacol. Bull. 2005, 20, 65–69. [Google Scholar]
- Ribeiro, R.D.; Rovai, A.S.; Twilley, R.R.; Castaneda-Moya, E. Spatial variability of mangrove primary productivity in the neotropics. Ecosphere 2019, 10, e02841. [Google Scholar] [CrossRef]
- David, F.; Marchand, C.; Nguyen, T.N.; Van, V.T.; Taillardat, P.; Meziane, T. Trophic relationships and basal resource utilisation in the Can Gio mangrove biosphere reserve (southern Vietnam). J. Sea Res. 2019, 145, 35–43. [Google Scholar] [CrossRef]
- Hill, J.M.; Jones, R.W.; Hill, M.P.; Weyl, O.L.F. Comparisons of isotopic niche widths of some invasive and indigenous fauna in a south African river. Freshw. Biol. 2015, 60, 893–902. [Google Scholar] [CrossRef]
- Lowe, S.; Browne, M.; Boudjelas, S.; Poorter, M. 100 of the World’s Worst Invasive Alien Species: A Selection from the Global Invasive Species Database; Invasive Species Specialist Group: Auckland, New Zealand, 2000; pp. 6–7. [Google Scholar]
- Chuong Nguyen, T.H.; Kim Lien, G.T.; Yen, P.H.; Ho, T.T.; Thuy Van, D.T.; Van Kiem, P.; Hung, N.H.; Kuo, P.C.; Setzer, W.N. Molluscicidal activity of compounds from the roots of Aralia armata against the golden apple snail (Pomacea canaliculata). Nat. Prod. Commun. 2022, 17, 1–12. [Google Scholar] [CrossRef]
- Xu, Y.X.; Wang, W.S.; Yao, J.M.; Yang, M.L.; Guo, Y.H.; Deng, Z.H.; Mao, Q.; Li, S.Z.; Duan, L.P. Comparative proteomics suggests the mode of action of a novel molluscicide against the invasive apple snail Pomacea canaliculata, intermediate host of Angiostrongylus cantonensis. Mol. Biochem. Parasit. 2022, 247, 111431. [Google Scholar] [CrossRef]
- Zhao, B.; Luo, M.; Zhang, J.; Liu, Y.; Deng, Z.; Gong, X. Genetic diversity of two globally invasive snails in Asia and Americas in relation with agricultural habitats and climate factors. Diversity 2022, 14, 1069. [Google Scholar] [CrossRef]
- Letelier, S.; Rebolledo, A.; Pedro Báez Fabres, A.; Soto-Acuña, S.; Jackson, D.; Mansilla, P.; Collado, G. The highly invasive freshwater apple snail Pomacea canaliculata (gastropoda: Ampullariidae) in northern Chile: Morphological and molecular confirmation. J. Zool. Stud. 2016, 3, 119–128. [Google Scholar]
- Yang, S.; Zhong, J.R.; Zhao, L.L.; Wu, H.; Du, Z.J.; Liu, Q.; Zhang, J.E.; Yan, T.M.; Huang, X.L. The salinity tolerance of the invasive golden apple snail (Pomacea canaliculata). Molluscan Res. 2017, 38, 90–98. [Google Scholar] [CrossRef]
- Tue, N.T.; Quy, T.D.; Nhuan, M.T.; Dung, L.V.; Thai, N.D. Tracing carbon transfer and assimilation by invertebrates and fish across a tropical mangrove ecosystem using stable isotopes. Mar. Ecol. Evol. Persp. 2017, 38, e12460. [Google Scholar] [CrossRef]
- Ma, W.; Wang, M.; Wang, W.; Liu, Y.; Luo, L.; Tang, C. Biodiversity of mangrove mollusks in the west coast of Hainan island, China. Biodivers. Sci. 2018, 26, 707–716. [Google Scholar] [CrossRef]
- Liu, J.L.; Zhao, B.L.; Li, Y.H.; Deng, X.Y.; Yue, Q.; Xu, J.T.; Xu, S.Q. Palatability of mangrove leaves to invasive apple snails: The relation between feeding electivity and multiple plant characteristics. Aquat. Invasions 2022, 17, 277–299. [Google Scholar] [CrossRef]
- Forgeron, S.J.; Quadros, A.F.; Zimmer, M. Crab-driven processing does not explain leaf litter-deposition in mangrove crab burrows. Ecol. Evol. 2021, 11, 8856–8862. [Google Scholar] [CrossRef]
- Behbahani, B.A.; Yazdi, F.T.; Shahidi, F.; Noorbakhsh, H.; Vasiee, A.; Alghooneh, A. Phytochemical analysis and antibacterial activities extracts of mangrove leaf against the growth of some pathogenic bacteria. Microb. Pathog. 2018, 114, 225–232. [Google Scholar] [CrossRef]
- Zimmer, M. Detritus. In Encyclopedia of Ecology, 2nd ed.; Elsevier: Oxford, UK, 2019. [Google Scholar]
- Pradisty, N.A.; Amir, A.A.; Zimmer, M. Plant species- and stage-specific differences in microbial decay of mangrove leaf litter: The older the better? Oecologia 2021, 195, 843–858. [Google Scholar] [CrossRef]
- Fratini, S.; Vigiani, V.; Vannini, M.; Cannicci, S. Terebralia palustris (gastropoda; Potamididae) in a Kenyan mangal: Size structure, distribution and impact on the consumption of leaf litter. Mar. Biol. 2004, 144, 1173–1182. [Google Scholar] [CrossRef]
- Penha-Lopes, G.; Bouillon, S.; Mangion, P.; Macia, A.; Paula, J. Population structure, density and food sources of Terebralia palustris (potamididae: Gastropoda) in a low intertidal Avicennia marina mangrove stand (inhaca island, Mozambique). Estuar. Coast Shelf Sci. 2009, 84, 318–325. [Google Scholar] [CrossRef]
- Yin, Y.; He, Q.; Pan, X.; Liu, Q.; Wu, Y.; Li, X. Predicting current potential distribution and the range dynamics of Pomacea canaliculata in China under global climate change. Biology 2022, 11, 110. [Google Scholar] [CrossRef]
- Guangzhou Nansha District Meteorological Bureau, Nansha Climate Monitoring Bulletin in 2021. Available online: Http://www.Gzns.Gov.Cn/gznsqx/gkmlpt/mindex (accessed on 10 December 2022).
- Yoshida, K.; Hoshikawa, K.; Wada, T.; Yusa, Y. Life cycle of the apple snail Pomacea canaliculata (caenogastropoda: Ampullariidae) inhabiting japanese paddy fields. Appl. Entomol. Zool. 2009, 44, 465–474. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.X.; Ren, H.; Hui, D.F.; Wang, W.Q.; Liao, B.W.; Cao, Q.X. Carbon stocks and potential carbon storage in the mangrove forests of China. J. Environ. Manag. 2014, 133, 86–93. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Ann. Chem. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Masoud, M.S.; Abdel-Halim, A.M.; El Ashmawy, A.A. Seasonal variation of nutrient salts and heavy metals in mangrove (Avicennia marina) environment, Red Sea, Egypt. Environ. Monit. Assess. 2019, 191, 425. [Google Scholar] [CrossRef]
- Islam, M.N.; Ratul, S.B.; Sharmin, A.; Rahman, K.; Ashaduzzaman, M.; Uddin, G.M. Comparison of calorific values and ash content for different woody biomass components of six mangrove species of Bangladesh sundarbans. J. Indian Acad. Wood Sci. 2019, 16, 110–117. [Google Scholar] [CrossRef]
- Fukushima, R.S.; Kerley, M.S.; Ramos, M.H.; Porter, J.H.; Kallenbach, R.L. Comparison of acetyl bromide lignin with acid detergent lignin and klason lignin and correlation with in vitro forage degradability. Anim. Feed Sci. Technol. 2015, 201, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Nierop, K.G.J.; Verstraten, J.M. Fate of tannins in corsican pine litter. J. Chem. Ecol. 2006, 32, 2709–2719. [Google Scholar] [CrossRef]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. The Principle and Technology of Plant Physiology and Biochemistry Experiment; Higher Education Press: Beijing, China, 2000. [Google Scholar]
- Öztaşkent, C.; Ak, İ. Effect of led light sources on the growth and chemical composition of brown seaweed Treptacantha barbata. Aquac. Int. 2020, 29, 193–205. [Google Scholar] [CrossRef]
- Roa, R. Design and analysis of multiple-choice feeding-preference experiments. Oecologia 1992, 89, 509–515. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: Https://www.R-project.Org/ (accessed on 10 December 2022).
- Blanchet, F.G.; Legendre, P.; Borcard, D. Forward selection of explanatory variables. Ecology 2008, 89, 2623–2632. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-6. Available online: Https://cran.R-project.Org/package=vegan (accessed on 10 December 2022).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Slowikowski, K. Ggrepel: Automatically Position Non-Overlapping Text Labels with ‘ggplot2′. R Package Version 0.8.2. Available online: Https://cran.R-project.Org/package=ggrepel (accessed on 10 December 2022).
- Lê, S.; Josse, J.; Husson, F. Factominer: An r package for multivariate analysis. J Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.5. Available online: https://cran.R-project.Org/package=factoextra (accessed on 10 December 2022).
- Azmi, W.A.; Khoo, S.C.; Ng, L.C.; Baharuddin, N.; Abd Aziz, A.; Ma, N.L. The current trend in biological control approaches in the mitigation of golden apple snail pomacea spp. Biol. Control 2022, 175, 105060. [Google Scholar] [CrossRef]
- Printrakoon, C.; Bullangpoti, V. Efficiency of monoterpene compounds for control of rice pest Pomacea canaliculata. Agric. Nat. Resour. 2021, 55, 7–14. [Google Scholar]
- Tanaka, K.; Watanabe, T.; Higuchi, H.; Miyamoto, K.; Yusa, Y.; Kiyonaga, T.; Kiyota, H.; Suzuki, Y.; Wada, T. Density-dependent growth and reproduction of the apple snail, Pomacea canaliculata: A density manipulation experiment in a paddy field. Res. Popul. Ecol. 1999, 41, 253–262. [Google Scholar] [CrossRef]
- Yusa, Y.; Wada, T.; Takahashi, S. Effects of dormant duration, body size, self-burial and water condition on the long-term survival of the apple snail, Pomacea canaliculata (gastropoda: Ampullariidae). Appl. Entomol. Zool. 2006, 41, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Horgan, F.G.; Stuart, A.M.; Kudavidanage, E.P. Impact of invasive apple snails on the functioning and services of natural and managed wetlands. Acta Oecol. 2014, 54, 90–100. [Google Scholar] [CrossRef]
- Morrison, W.E.; Hay, M.E. Feeding and growth of native, invasive and non-invasive alien apple snails (Ampullariidae) in the united states: Invasives eat more and grow more. Biol. Invasions 2011, 13, 945–955. [Google Scholar] [CrossRef]
- Yusa, Y.; Sugiura, N.; Wada, T. Predatory potential of freshwater animals on an invasive agricultural pest, the apple snail Pomacea canaliculata (gastropoda: Ampullariidae), in southern Japan. Biol. Invasions 2006, 8, 137–147. [Google Scholar] [CrossRef]
- Fratini, S.; Cannicci, S.; Vannini, M. Competition and interaction between Neosarmatium smithi (crustacea: Grapsidae) and Terebralia palustris (mollusca: Gastropoda) in a Kenyan mangrove. Mar. Biol. 2000, 137, 309–316. [Google Scholar] [CrossRef]
- Takeichi, M.; Hirai, Y.; Yusa, Y. A water-borne sex pheromone and trail following in the apple snail, Pomacea canaliculata. J. Molluscan Stud. 2007, 73, 275–278. [Google Scholar] [CrossRef] [Green Version]
- Noghabi, N.A.; Shojaei, M.G.; Farahani, M.M.; Weigt, M. Stable isotopes reveal the food sources of benthic macroinvertebrates in the arid mangrove ecosystem of the persian gulf. Estuar. Coast 2022, 45, 2241–2253. [Google Scholar] [CrossRef]
- Ariyanto, D.; Bengen, D.G.; Prartono, T.; Wardiatno, Y. The relationship between content of particular metabolites of fallen mangrove leaves and the rate at which the leaves decompose over time. Biodiversitas 2018, 19, 730–735. [Google Scholar] [CrossRef]
- Praveen, V.P.; Shanij, K.; Suresh, S.; Oommen, M.M.; Nayar, T.S. Species preference in seedling predation by the sesarmid crab Neosarmatium malabaricum (Henderson, 1893) in a mangrove forest in India. Wetlands 2017, 37, 59–66. [Google Scholar] [CrossRef]
- Li, M.T.; Liu, J.L.; Zhao, B.L.; Deng, X.Y.; Chen, H.Y.; Tang, Y.J. Research progress on characteristics and applications of endogenous cellulase in apple snail. Chin. J. Ecol. 2021, 40, 559–567. [Google Scholar]
- Dai, L.P.; Qian, X.W.; Nan, X.Y.; Zhang, Y.J. Effect of cardiac glycosides from nerium indicum on feeding rate, digestive enzymes activity and ultrastructural alterations of hepatopancreas in Pomacea canaliculata. Environ. Toxicol. Phar. 2014, 37, 220–227. [Google Scholar] [CrossRef] [PubMed]
- An, T.C.; Lyu, J.C.; Jia, W.X.; Wang, M.; Wei, S.S.; Zhang, Y.X. Role of macrobenthic fauna in mangrove carbon fluxes indicated by their cellulase and hemicellulase activities. Mar. Biol. Res. 2014, 10, 934–940. [Google Scholar] [CrossRef]
- Vassilev, S.V.; Baxter, D.; Andersen, L.K.; Vassileva, C.G. An overview of the composition and application of biomass ash. Part 1. Phase-mineral and chemical composition and classification. Fuel 2013, 105, 40–76. [Google Scholar] [CrossRef]
- Glass, N.H.; Darby, P.C. The effect of calcium and pH on Florida apple snail, Pomacea paludosa (gastropoda: Ampullariidae), shell growth and crush weight. Aquat. Ecol. 2009, 43, 1085–1093. [Google Scholar] [CrossRef]
- Nabeelah Bibi, S.; Fawzi, M.M.; Gokhan, Z.; Rajesh, J.; Nadeem, N.; Rengasamy Kannan, R.R.; Albuquerque, R.D.D.G.; Pandian, S.K. Ethnopharmacology, phytochemistry, and global distribution of mangroves-a comprehensive review. Mar. Drugs 2019, 17, 231. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.W.; Chan, M.T.; Kwong, K.L.; Sun, J. Consumption, survival and growth in the invasive freshwater snail Pomacea canaliculata: Does food freshness matter? J. Molluscan Stud. 2011, 77, 189–195. [Google Scholar] [CrossRef]
- Zhang, J.L.; Lin, Q.L.; Peng, Y.S.; Pan, L.H.; Chen, Y.; Zhang, Y.; Chen, L.Z. Distributions of the non-native mangrove Sonneratia apetala in China: Based on google earth imagery and field survey. Wetlands 2022, 42, 35. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristics | Measurement Method |
|---|---|
| Carbon | Potassium dichromate oxidation method [35] |
| Phosphorous | Ascorbic acid-antimony reduced phosphate colorimetric method [36] |
| Calcium | EDTA titrimetric method [37] |
| Protein and nitrogen content | National Standards of China (GB 5009.5-2016) |
| Ash | Dry ash method [38] |
| Crude fiber | National Standards of China (GB/T 5009.10-2003) |
| Lignin | Klason method [39] |
| Tannin and phenol | EDTA method [40] |
| Flavonoid | Ethanol extraction method [41] |
| Indicator (g g−1 d−1) | Carbon (g kg−1) | Nitrogen (g kg−1) | C/N Ratio | Calcium (%) | Phosphate (g kg−1) |
|---|---|---|---|---|---|
| WDR(multiple-choice) | −0.284 | 0.485 * | −0.396 | 0.109 | 0.685 ** |
| WDR(no-choice) | −0.044 | 0.345 | −0.257 | 0.275 | 0.544 * |
| Indicator (g g−1 d−1) | Ash (g kg−1) | Fiber (g kg−1) | Flavonoid (g kg−1) | Lignin (g kg−1) | Protein (g kg−1) | Tannin (g kg−1) | Phenolic (g kg−1) |
|---|---|---|---|---|---|---|---|
| WDR(multiple-choice) | 0.531 * | −0.169 | −0.697 ** | −0.695 ** | 0.485 * | −0.606 ** | −0.552 * |
| WDR(no-choice) | 0.644 ** | −0.176 | −0.755 ** | −0.527 * | 0.345 | −0.474 * | −0.425 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Chen, Z.; Li, Y.; Chen, D.; He, Y.; Zhao, B.; Liao, Y.; Guo, J. Potential Detritivorous Diet of the Invasive Apple Snail (Pomacea canaliculata Lamarck, 1822) in Mangroves: The Relationship between Feeding Indicators and Chemical Characteristics of Decaying Leaf Litter. J. Mar. Sci. Eng. 2023, 11, 672. https://doi.org/10.3390/jmse11030672
Liu J, Chen Z, Li Y, Chen D, He Y, Zhao B, Liao Y, Guo J. Potential Detritivorous Diet of the Invasive Apple Snail (Pomacea canaliculata Lamarck, 1822) in Mangroves: The Relationship between Feeding Indicators and Chemical Characteristics of Decaying Leaf Litter. Journal of Marine Science and Engineering. 2023; 11(3):672. https://doi.org/10.3390/jmse11030672
Chicago/Turabian StyleLiu, Jinling, Zhihua Chen, Yunhui Li, Danying Chen, Yulin He, Benliang Zhao, Yuexin Liao, and Jing Guo. 2023. "Potential Detritivorous Diet of the Invasive Apple Snail (Pomacea canaliculata Lamarck, 1822) in Mangroves: The Relationship between Feeding Indicators and Chemical Characteristics of Decaying Leaf Litter" Journal of Marine Science and Engineering 11, no. 3: 672. https://doi.org/10.3390/jmse11030672