
Age and Depositional Environment of Whale-Bearing Sedimentary Succession from the Lower Pliocene of Tuscany (Italy): Insights from Palaeomagnetism, Calcareous Microfossils and Facies Analyses
 , , , ,
, , , ,
Abstract
:1. Introduction
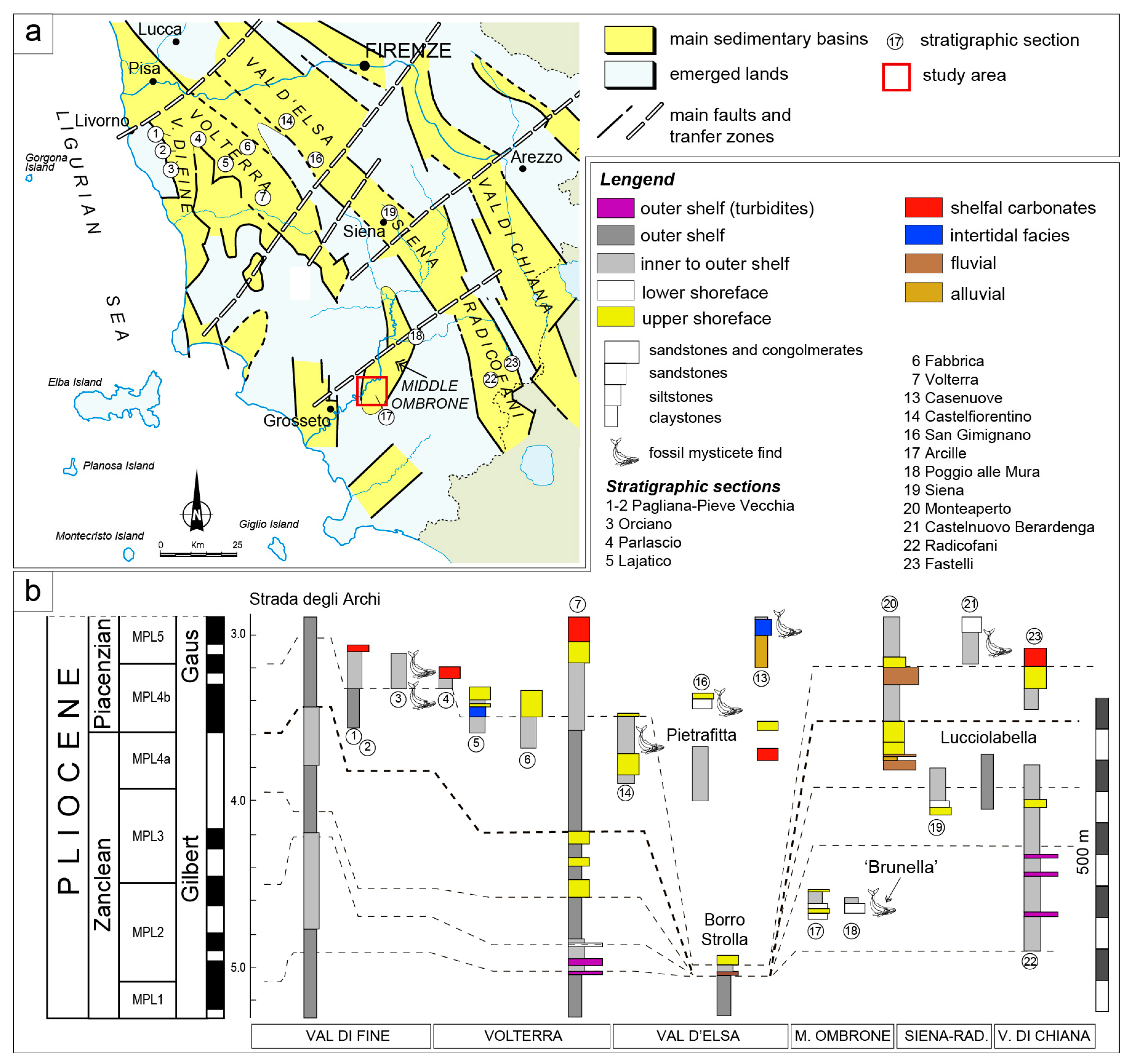
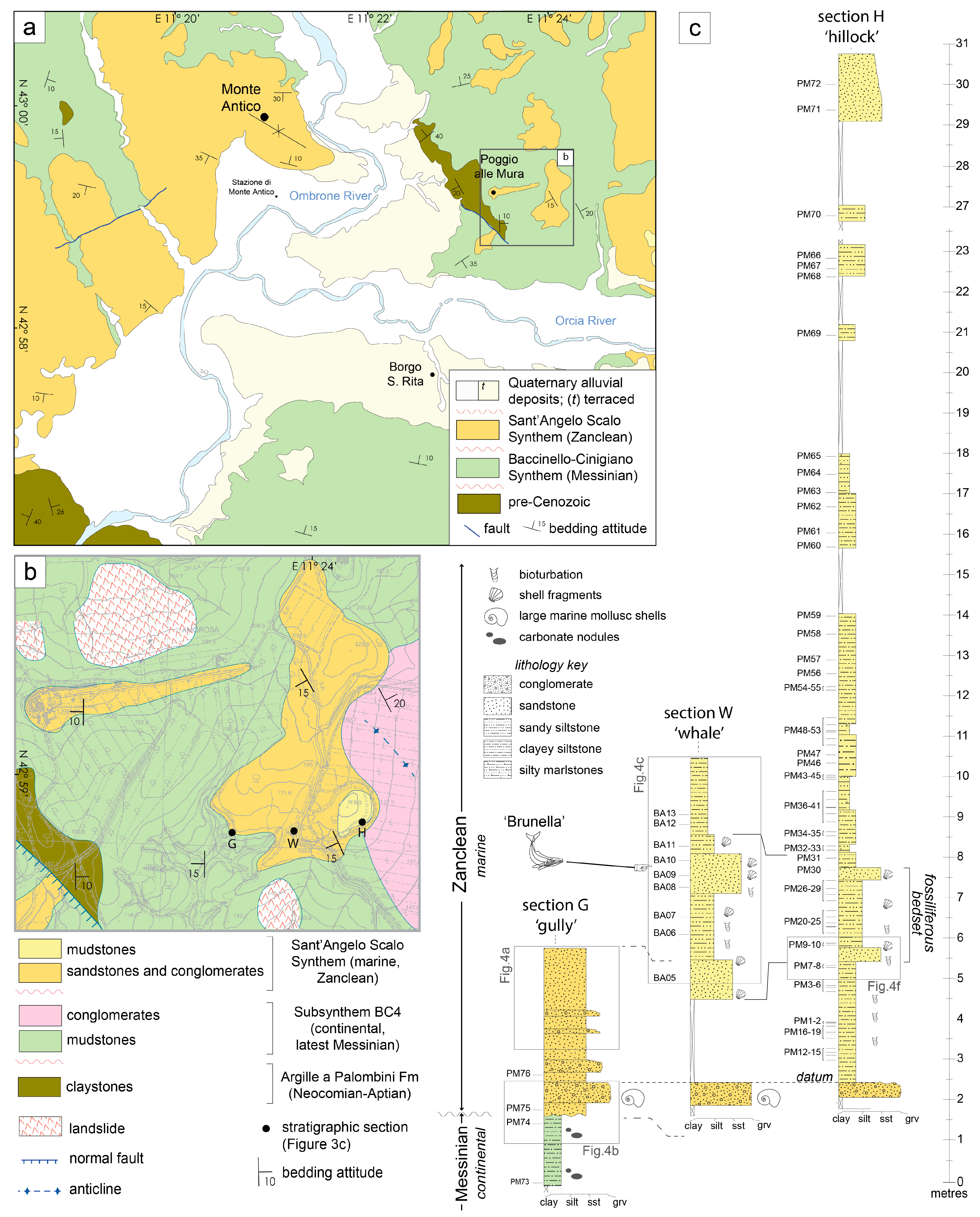
2. Geological Setting


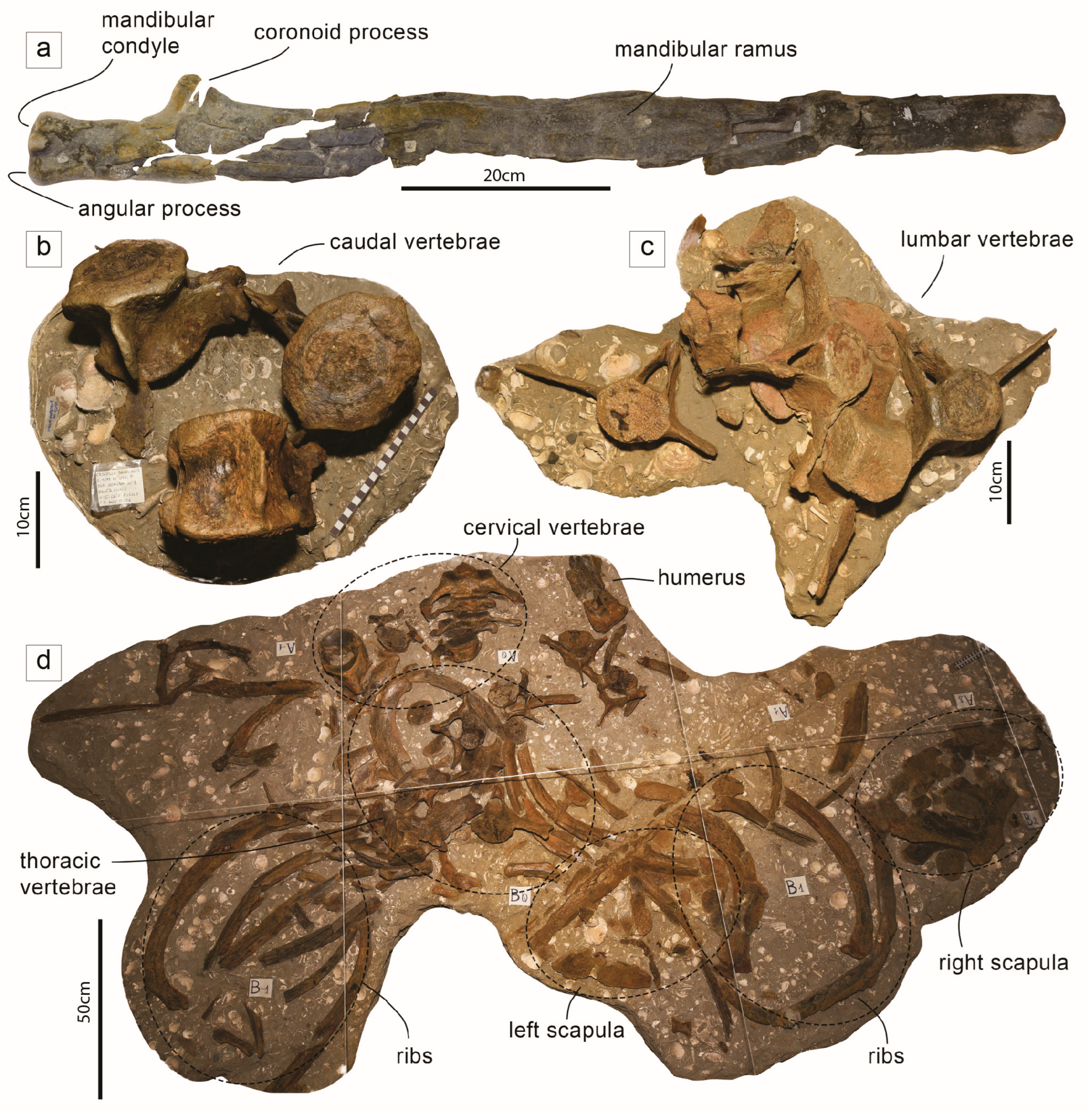
The Whale Brunella and Its Host Sedimentary Rocks
3. Materials and Methods
4. Results
4.1. Sedimentary Facies Association
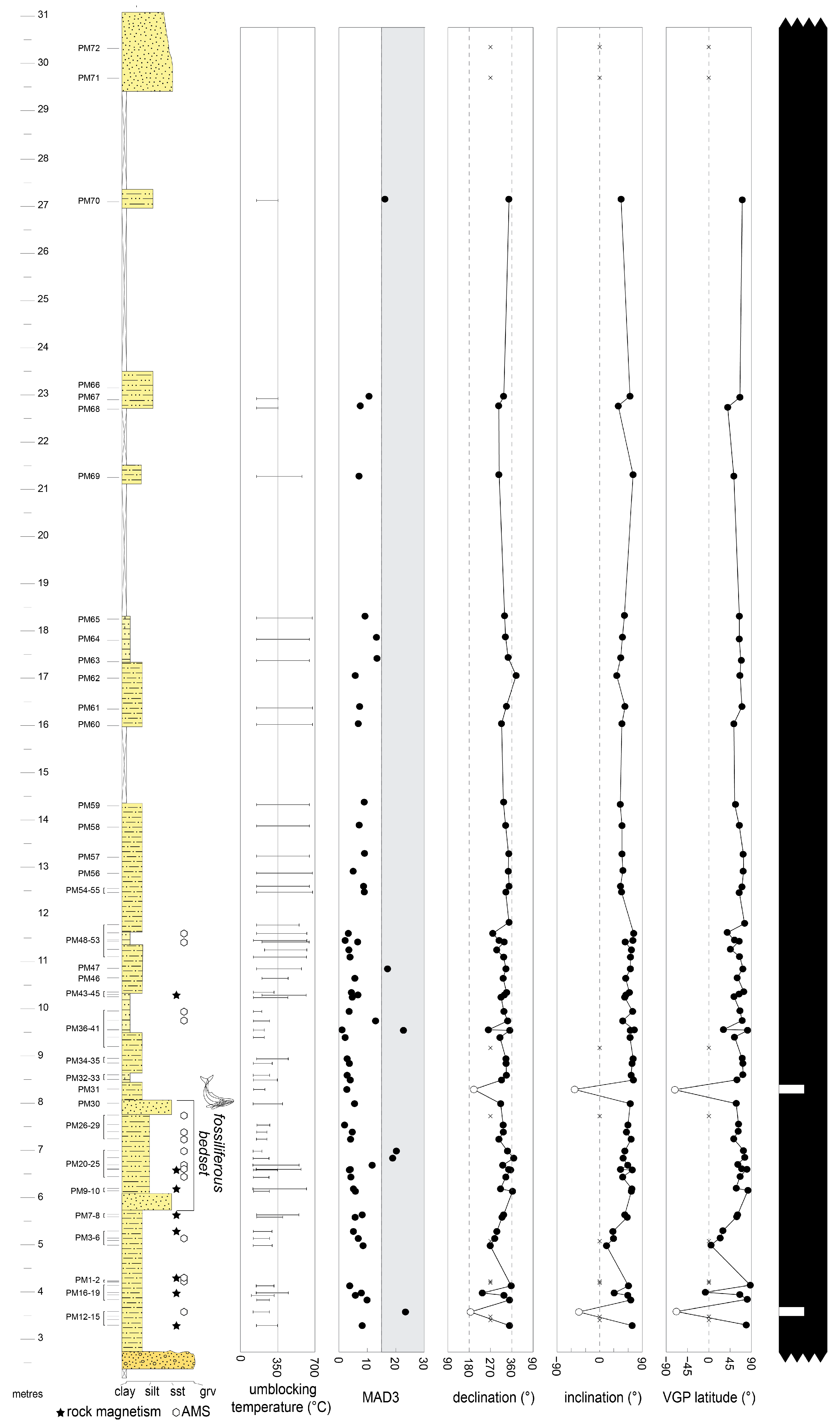
4.2. Palaeomagnetism
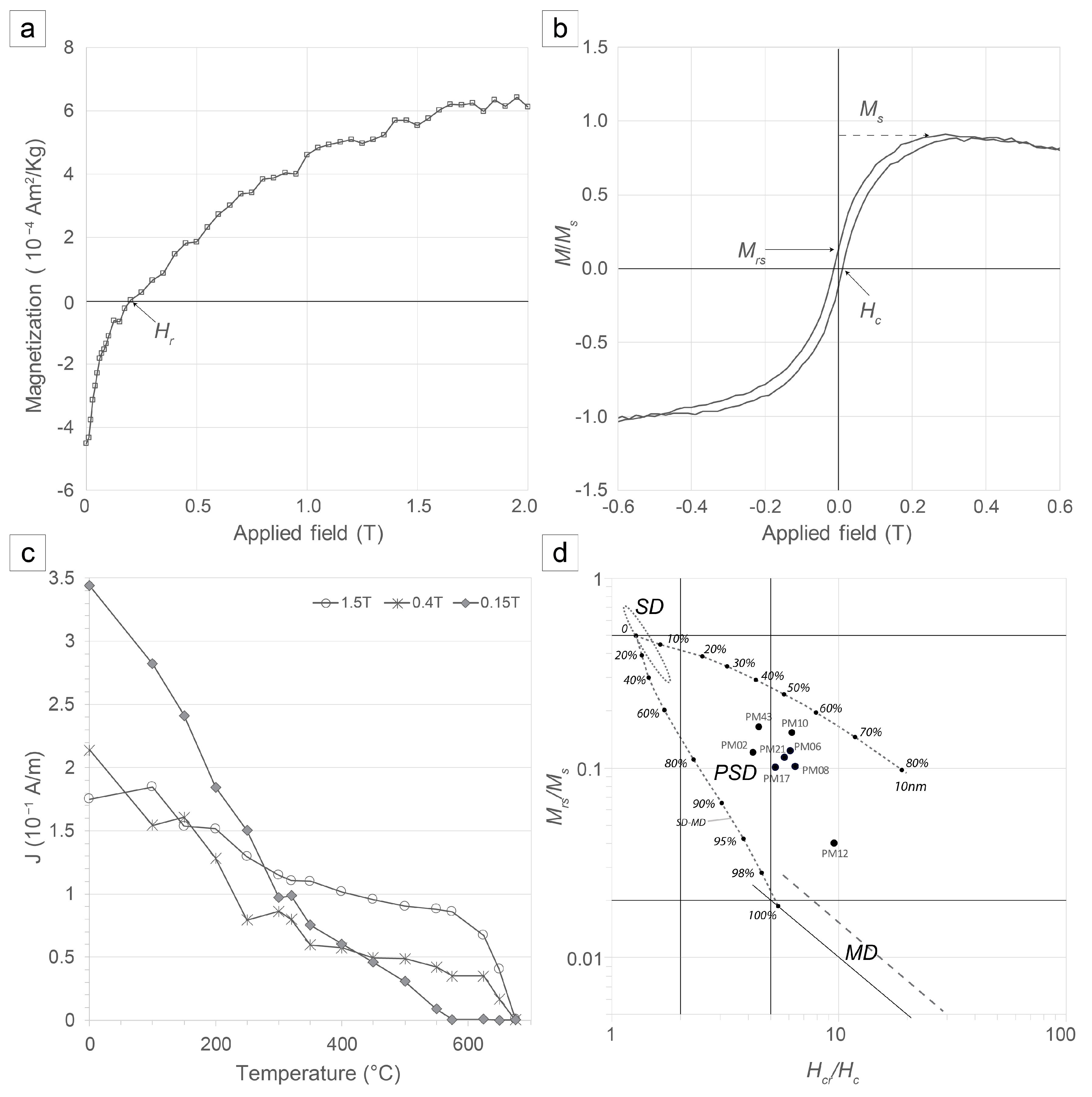
4.2.1. Rock Magnetism

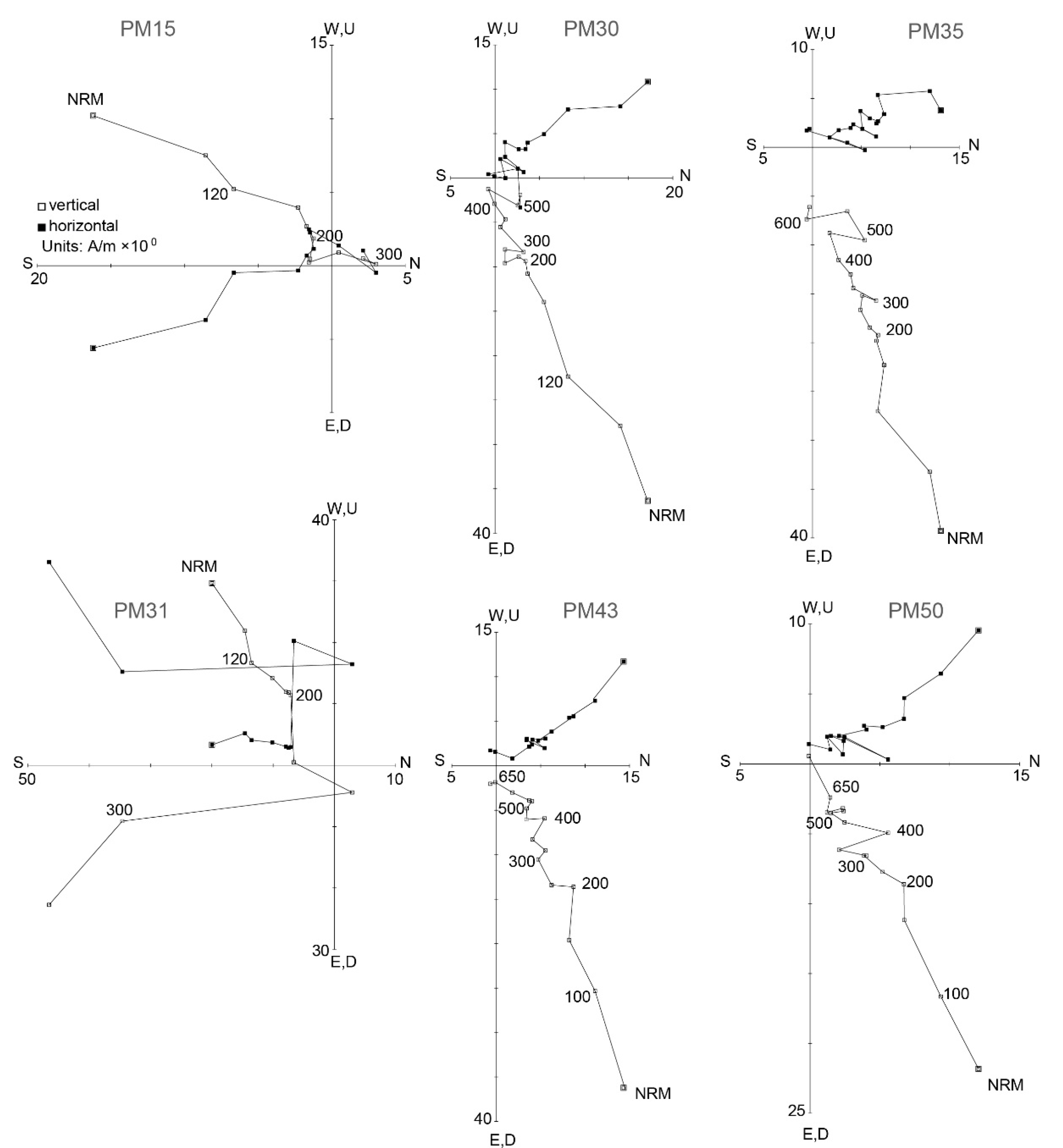
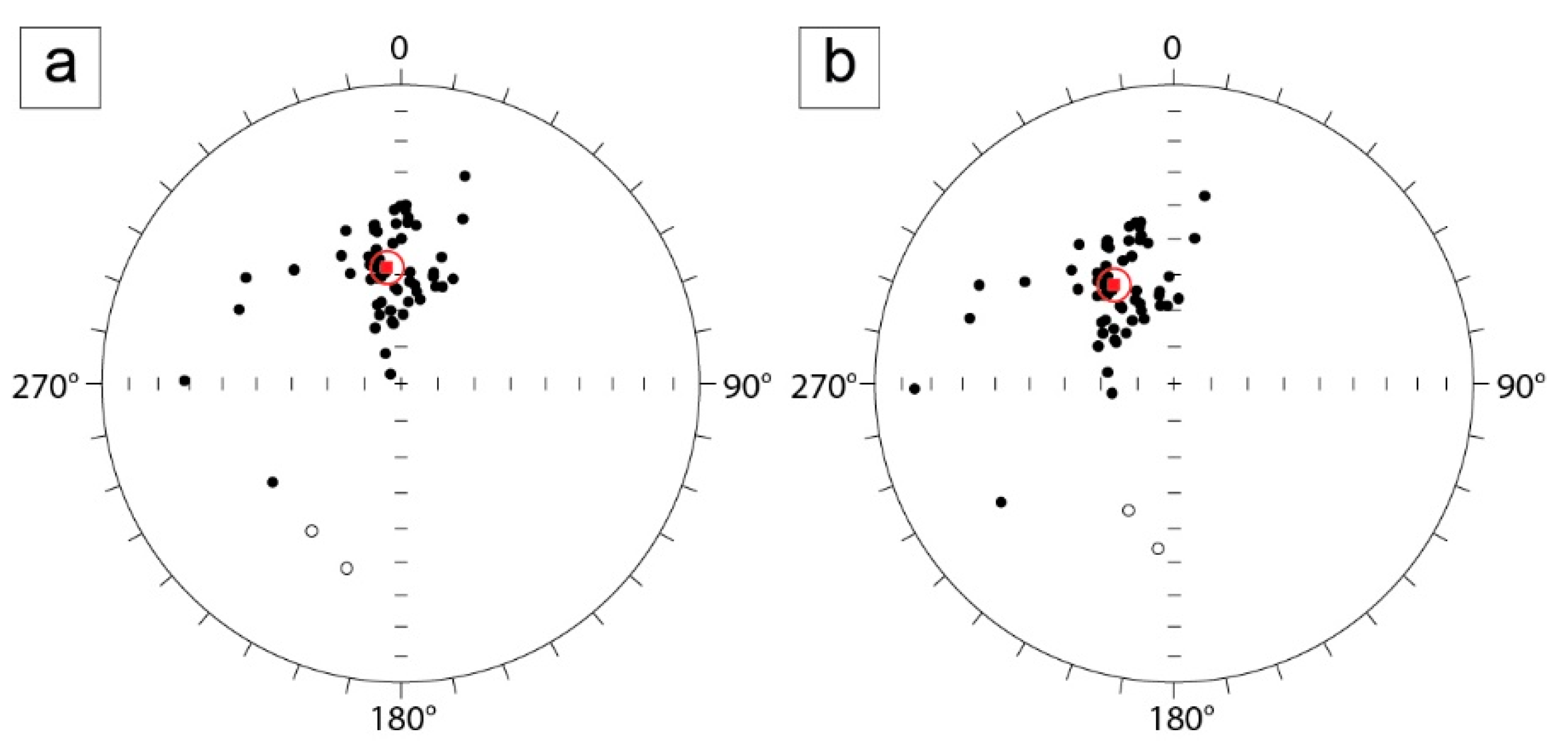
4.2.2. Characteristic Remnant Magnetisation
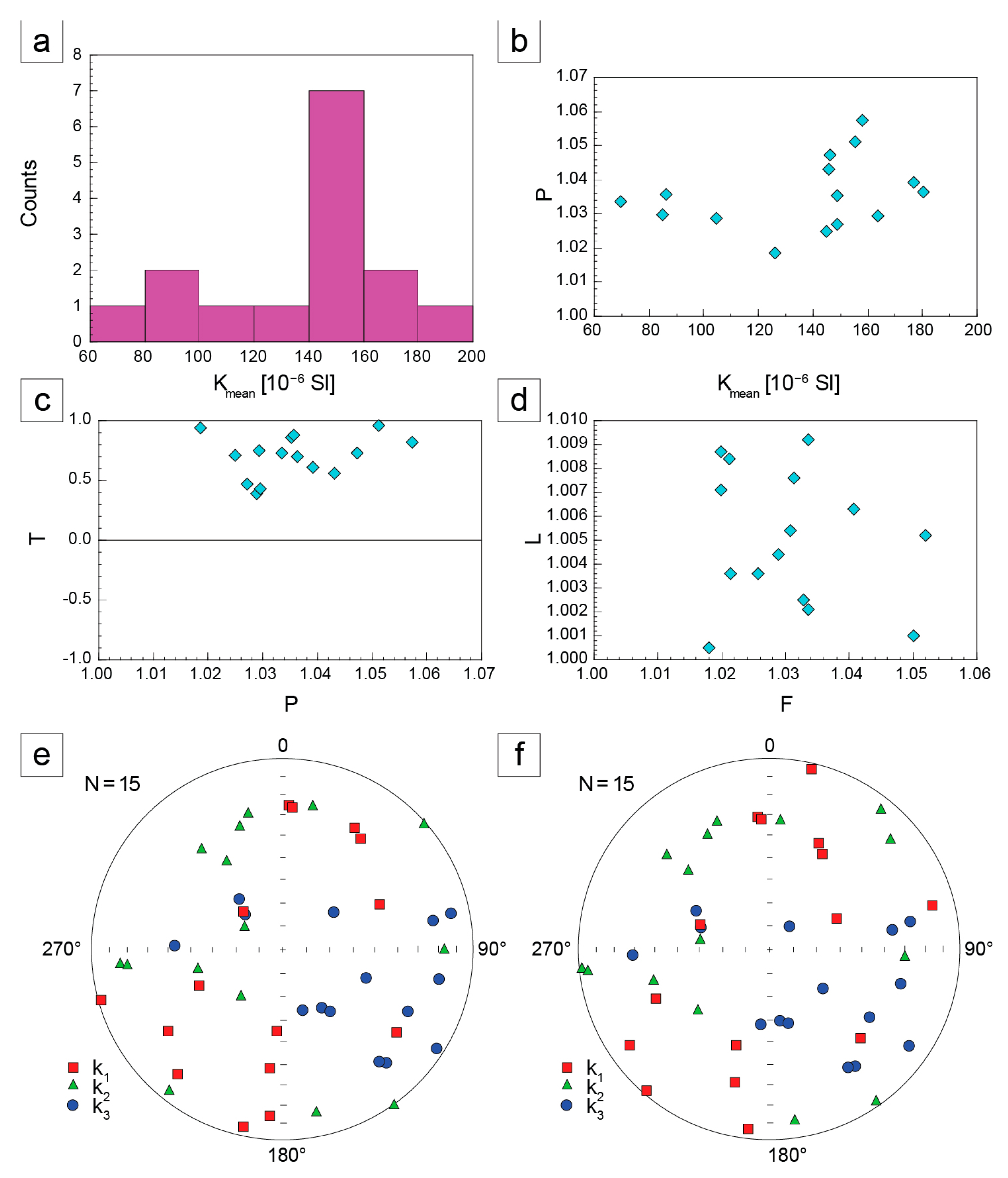
4.2.3. Anisotropy of Magnetic Susceptibility (AMS)
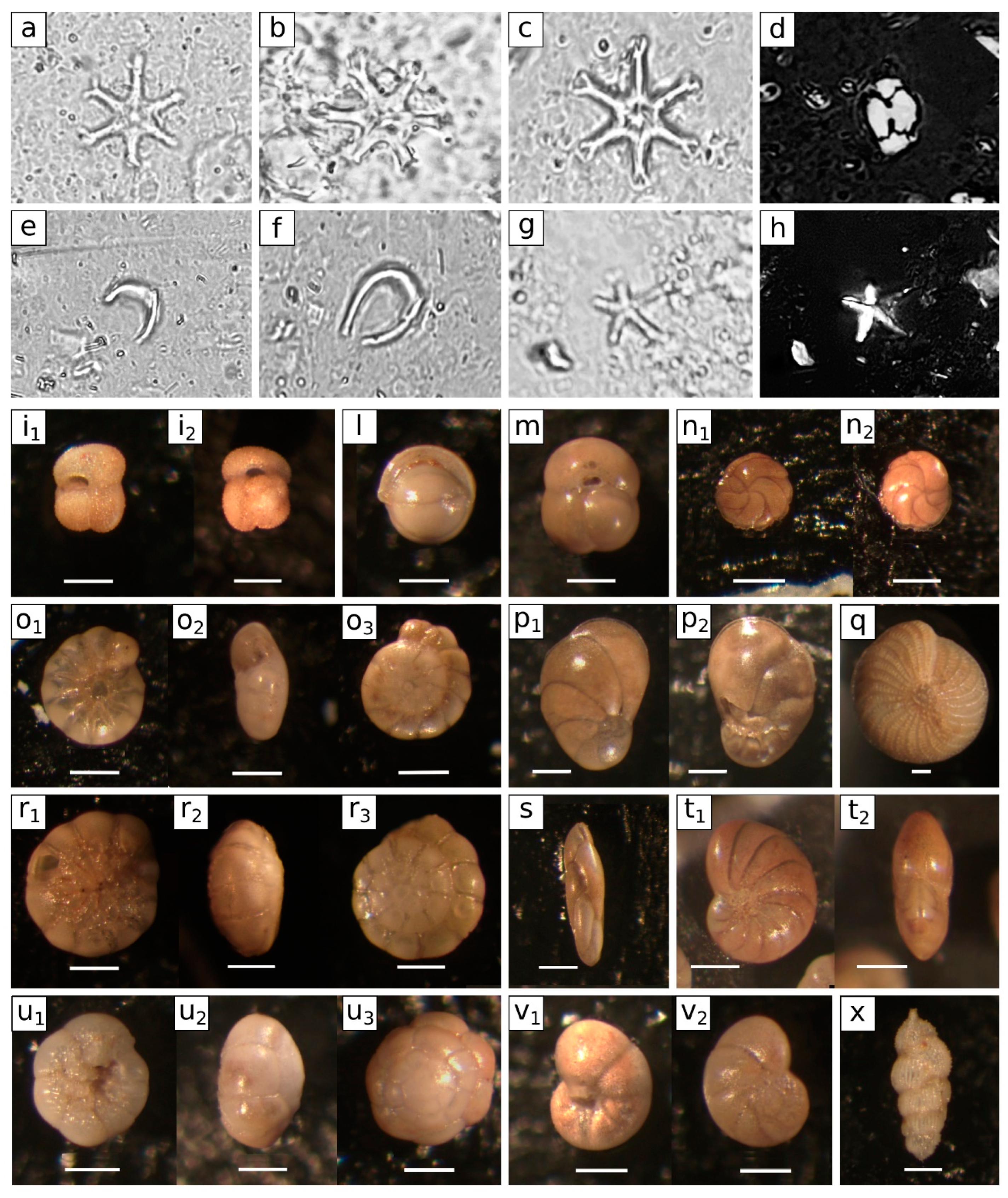
4.3. Micropalaeontology
4.3.1. Calcareous Plankton
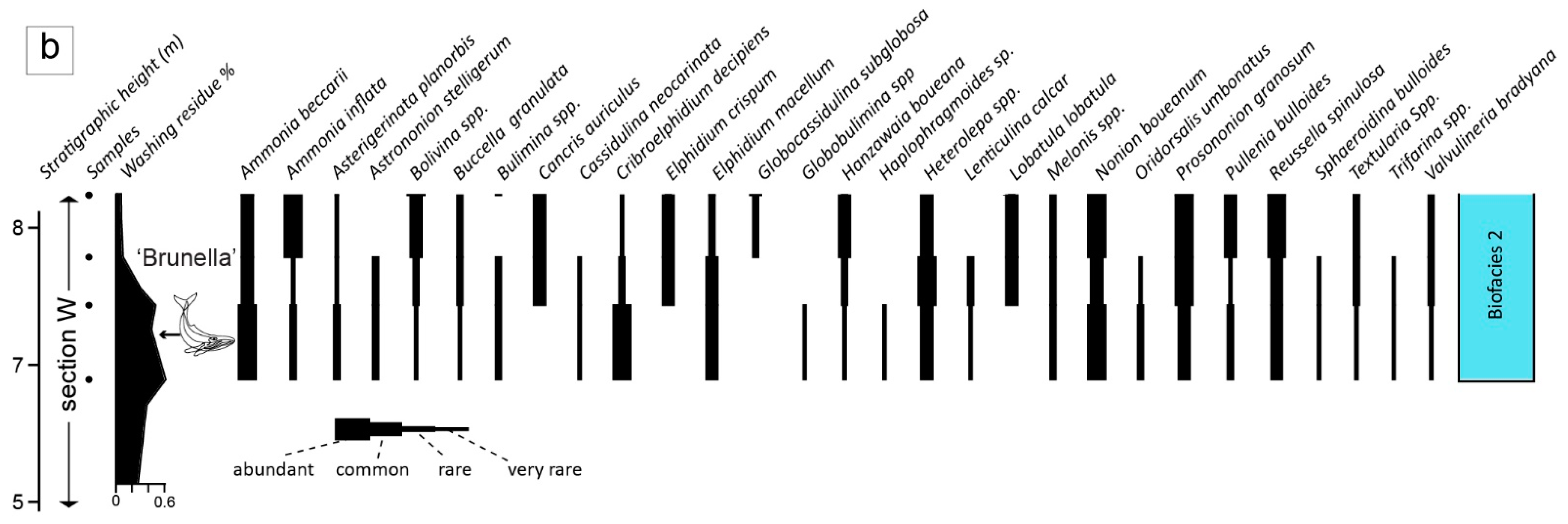
4.3.2. Benthic Foraminifera
5. Discussion
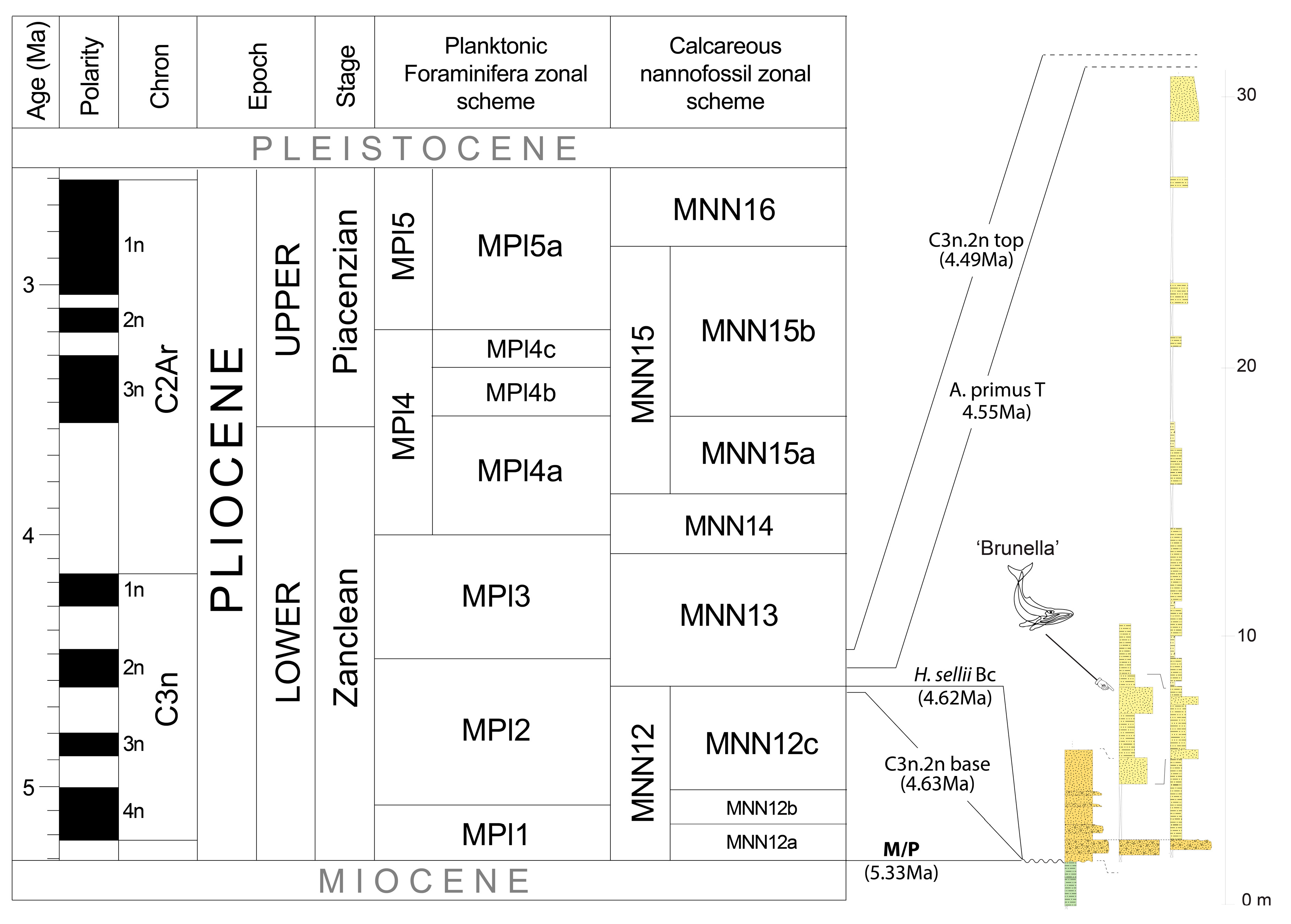
5.1. Depositional Age of Brunella
5.2. Palaeoenvironmental Insights from Sedimentary Facies, AMS and Benthic Foraminifera
6. Conclusions
- Magnetobiochronology results provide evidence that the clastic sediments embedding Brunella were deposited during the polarity Chron C3n.2n and between 4.62 Ma (H. sellii Bc—Base common) and 4.55 Ma (A. primus T—Top), which corresponds to the lowermost part of the calcareous nannofossil MNN13 zone;
- The depositional age of the marine interval of the investigated composite section is constrained to be within a very short chronological interval (c. 70 Ky) of the Zanclean, which makes Brunella the earliest baleen whale of Tuscany postdating the Messinian Salinity Crisis;
- Sedimentary facies and benthic Foraminifera assemblages indicate Brunella was buried by sediments accumulating in a nearshore setting above the fair-weather base level, at a pace greater than c. 40 cm/Ky.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dominici, S.; Danise, S.; Benvenuti, M. Pliocene stratigraphic paleobiology in Tuscany and the fossil record of marine megafauna. Earth-Sci. Rev. 2018, 176, 277–310. [Google Scholar] [CrossRef]
- Bisconti, M. Cenozoic environmental changes and evolution of baleen whales. In Whales and Dolphins: Behavior, Biology and Distribution; Murray, C.A., Ed.; Nova Publishing: New York, NY, USA, 2010; pp. 1–46. [Google Scholar]
- Bisconti, M. Storia evolutiva dei Mysticeti: Diversità, estinzioni e conservazione. In Estinzioni di Massa e Biodiversità—XXII Giornata dell’Ambiente; Battaglia, B., Bullini, L., D’Argenio, B., Gerola, F.M., Pignatti, S., Scarascia Mugnozza, G.T., Eds.; Accademia Nazionale dei Lincei, Atti dei Convegni Lincei, Bardi Editore: Roma, Italy, 2005; Volume 220, pp. 95–105. [Google Scholar]
- Hsü, K.J.; Ryan, W.B.F.; Cita, M.B. Late Miocene desiccation of the Mediterranean. Nature 1973, 242, 240–244. [Google Scholar] [CrossRef]
- Bisconti, M.; Munsterman, D.K.; Post, K. A new balaenopterid whale from the late Miocene of the Southern North Sea Basin and the evolution of balaenopterid diversity (Cetacea, Mysticeti). PeerJ 2019, 7, e6915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitmore, F.C., Jr. Neogene climatic change and the emergence of the modern whale fauna of the North Atlantic Ocean. In Contributions in Marine Mammal Paleontology Honoring Frank C. Whitmore; Berta, A., Deméré, T.A., Eds.; Proceedings of the San Diego Society of Natural History San Diego; San Diego Society of Natural History: San Diego, CA, USA, 1994; Volume 29, pp. 223–227. [Google Scholar]
- Bisconti, M. Evolutionary history of Balaenidae. Cranium 2003, 20, 9–50. [Google Scholar]
- Bisconti, M. Taxonomy and evolution of the Italian Pliocene Mysticeti (Mammalia, Cetacea): A state of the art. Boll. Soc. Paleont. Ital. 2009, 48, 147–156. [Google Scholar]
- Dominici, S.; Danise, S.; Cau, S.; Freschi, A. The awkward record of fossil whales. Earth-Sci. Rev. 2019, 205, 103057. [Google Scholar] [CrossRef]
- Bisconti, M.; Pellegrino, L.; Carnevale, G. Evolution of gigantism in right and bowhead whales (Cetacea, Mysticeti, Balaenidae). Biol. J. Linn. Soc. 2021, 134, 498–524. [Google Scholar] [CrossRef]
- Bisconti, M.; Bosselaers, M. On Plesiocetus Van Beneden, 1859 (Mammalia, Cetacea, Mysticeti). Riv. Ital. Paleontol. Stratigr. 2021, 127, 231–274. [Google Scholar] [CrossRef]
- Bisconti, M. New description, character analysis and preliminary phylogenetic assessment of two Balaenidae skulls from the Italian Pliocene. Palaeontogr. Ital. 2000, 87, 37–66. [Google Scholar]
- Cau, S.; Freschi, A. Revisione cronostratigrafica dei cetacei del Plio-Pleistocene nell’Appennino settentrionale. Parva Nat. 2020, 15, 59–83. [Google Scholar]
- Foresi, L.M.; Bambini, A.M. Il Neogene. In Atlante dei Dati Biostratigrafici della Toscana; Cerrina Feroni, A., Ed.; CNR-IGG: Pisa, Italy, 2010. [Google Scholar]
- Scotton, R.; Bigazzi, R.; D’Amore, G.; Di Marco, S.; Koenig, E.; Nannini, P.; Santagati, P.; Tabolli, J.; Tartarelli, G.; Tarantini, M.; et al. Il Progetto “Brunella”: Principali attività preparatorie e strategie di comunicazione intorno ad un balenotteride pliocenico in Toscana. Parva Nat. 2020, 15, 85–133. [Google Scholar]
- Bisconti, M.; Scotton, R.; Santagati, P.; Foresi, L.M.; Ragaini, L.; Tartarelli, G.; Carnevale, G.; Buckeridge, J.; Koenig, E.; Tabolli, J.; et al. A Whale in a Vineyard: Palaeontological Preparation and Education During the ‘Brunella’ Project, a Large-Scale Conservation Effort Focused on a Pliocene Whale in Southern Tuscany, Italy. Geoheritage 2023, 15, 1–19. [Google Scholar] [CrossRef]
- Damiani, A.V.; Gandin, A.; Pannuzzi, L. Il Bacino dell’Ombrone-Orcia nelquadro dell’evoluzione paleografica e tettonica della Toscana meridionale. Mem. Soc. Geol. Ital. 1981, 21, 281–287. [Google Scholar]
- Bossio, A.; Costantini, A.; Foresi, L.M.; Mazzei, R.; Monteforti, B.; Salvatorini, G.; Sandrelli, F. Notizie preliminari sul Pliocene del bacino del medio Ombrone e della zona di Roccastrada. Atti Soc. Toscana Sci. Nat. 1991, 98, 259–269. [Google Scholar]
- Bossio, A.; Costantini, A.; Foresi, L.M.; Mazzei, R.; Monteforti, B.; Radi, L.; Salvatorini, G.; Sandrelli, F. Carta Geologica dell’Area del Medio Ombrone (Provincie di Siena e Grosseto); Cartografia S.E.L.C.A.: Firenze, Italy, 1991. [Google Scholar]
- Benvenuti, M.; Moratti, G.; Sani, F.; Bonini, M.; Oms, O.L.; Papini, M.; Rook, L.; Cavallina, C.; Cavini, L. Messinian-earliest Zanclean tectonic-depositional dynamics of the Cinigiano-Baccinello and Velona basins (Tuscany, Italy). Ital. J. Geosci. 2015, 134, 237–254. [Google Scholar] [CrossRef]
- Bossio, A.; Cerri, R.; Costantini, A.; Gandin, A.; Lazzarotto, A.; Magi, M.; Sandrelli, F. I Bacini distensivi Neogenici e Quaternari della Toscana. In 76° Riunione Estiva della Società Geologica Italiana—Convegno SIMP; Guida all’Escursione: Firenze, Italy, 1992; Volume B4, pp. 198–227. [Google Scholar]
- Bossio, A.; Costantini, A.; Lazzarotto, A.; Liotta, D.; Mazzanti, R.; Mazzei, R.; Salvatorini, G.; Sandrelli, F. Rassegna delle conoscenze sulla stratigrafia del neoautoctono toscano. Mem. Soc. Geol. Ital. 1993, 49, 17–98. [Google Scholar]
- Martini, I.P.; Sagri, M. Tectono-sedimentary characteristics of Late Miocene-Quaternary extensional basins of the Northern Apennines, Italy. Earth Sci. Rev. 1993, 34, 197–233. [Google Scholar] [CrossRef]
- DeCelles, P.G. Foreland basin systems revisited: Variations in response to tectonic settings. In Tectonics of Sedimentary Basins: Recent Advances; Busby, C., Azor, A., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2012; pp. 405–426. [Google Scholar]
- Martini, I.; Ambrosetti, E.; Brogi, A.; Aldinucci, M.; Zwaan, F.; Sandrelli, F. Polyphase extensional basins: Interplay between tectonics and sedimentation in the Neogene Siena-Radicofani Basin (Northern Apennines, Italy). Int. J. Earth Sci. 2021, 110, 1729–1751. [Google Scholar] [CrossRef]
- Pascucci, V.; Costantini, A.; Martini, I.P.; Dringoli, R. Tectonosedimentary analysis of a complex, extensional, Neogene basin formed on thrust-faulted, Northern Apennines hinterland: Radicofani Basin, Italy. Sediment. Geol. 2006, 183, 71–97. [Google Scholar] [CrossRef]
- Mattei, M.; Kissel, C.; Funiciello, R. No tectonic rotation of the Tuscan Tyrrhenian margin (Italy) since late Messinian. J. Geophys. Res. Solid Earth 1996, 101, 2835–2845. [Google Scholar] [CrossRef]
- Benvenuti, M.; Del Conte, S.; Scarselli, N.; Dominici, S. Hinterland basin development and infilling through tectonic and eustatic processes: Latest Messinian-Gelasian Valdelsa Basin, Northern Apennines, Italy. Basin Res. 2014, 26, 387–402. [Google Scholar] [CrossRef]
- Bonini, M.; Sani, F.; Stucchi, E.M.; Moratti, G.; Benvenuti, M.; Menanno, G.; Tanini, C. Late Miocene shortening of the Northern Apennines back-arc. J. Geodyn. 2014, 74, 1–31. [Google Scholar] [CrossRef]
- Brogi, A. Bowl-shaped basin related to low-angle detachment during continental extension: The case of the controversial Neogene Siena Basin (central Italy, Northern Apennines). Tectonophysics 2011, 499, 54–76. [Google Scholar] [CrossRef]
- Brogi, A. Late evolution of the inner Northern Apennines from the structure of the Monti del Chianti-Monte Cetona ridge (Tuscany, Italy). J. Struct. Geol. 2020, 141, 104205. [Google Scholar] [CrossRef]
- Regione Toscana. Geological Maps at Scale 1:10,000 Composing the Sheet n. 287 (scale 1:50,000) Montevarchi. 2007. Available online: http://www.regione.toscana.it/ambienteeterritorio/geologia/index.html (accessed on 1 February 2023).
- De Pretis, F. La Balena che Nuotava nel Mediterraneo Nano. 3 Domande a Michelangelo Bisconti; La Stampa TuttoScienze: Torino, Italy, 2017; p. 7. [Google Scholar]
- Batini, G. La Toscana delle Balene; Edizioni Polistampa: Firenze, Italy, 2009; Available online: https://www.ibs.it/toscana-delle-balene-quando-grandi-libro-giorgio-batini/e/9788859605010 (accessed on 1 February 2023).
- Scotton, R.; Bigazzi, R.; Casati, S.; D’Amore, G.; Di Marco, S.; Foresi, L.M.; Koenig, E.; Ragaini, L.; Tabolli, J.; Tarantini, M.; et al. The “Brunella” Project: Preparation and study of a mysticete from the Early Pliocene of Tuscany. Fossilia 2018, 15, 61–63. [Google Scholar]
- Motta, S. Note Illustrative della Carta Geologica d’Italia alla Scala 1:100,000; Foglio 128 Grosseto: Ercolano, Italy, 1969. [Google Scholar]
- Tinelli, C. Marine Vertebrates from Pliocene Shell Beds from Tuscany (Italy): Prospecting, Taphonomy, Palaeoecology and Systematic Palaeontology. Ph.D. Thesis, Università di Pisa, Pisa, Italy, 2013. [Google Scholar]
- Sprovieri, R. Mediterranean Pliocene biochronology: An high resolution record based on quantitative planktonic foraminifera distribution. Riv. Ital. Paleontol. Stratigr. 1992, 98, 61–100. [Google Scholar]
- Hilgen, F.; Lourens, L.J.; Van Dam, J.A.; Beu, A.G.; Boyes, A.F.; Cooper, R.A.; Krigsman, W.; Ogg, J.G.; Piller, W.E.; Wilson, D.S. The Neogene period. In The Geologic Time Scale 2012; Gradstein, F.M., Ogg, J.G., Schmitz, M., Ogg, G., Eds.; Elsevier Publ.: Amsterdam, The Netherlands, 2012; pp. 923–978. [Google Scholar]
- Tinelli, C.; Ribolini, A.; Bianucci, G.; Bini, M.; Landini, W. Ground penetrating radar and palaeontology: The detection of sirenian fossil bones under a sunflower field in Tuscany (Italy). Comptes Rendus Palevol 2012, 11, 445–454. [Google Scholar] [CrossRef]
- Sorbi, S.; Vaiani, S.C. New sirenian record from Lower Pliocene sediments of Tuscany (Italy). Riv. Ital. Paleontol. Stratigr. 2007, 113, 299–304. [Google Scholar]
- Dominici, S.; Forli, M. Lower Pliocene molluscs from southern Tuscany (Italy). Boll. Soc. Pal. Ital. 2021, 60, 69–98. [Google Scholar]
- Lowrie, W. Identification of ferromagnetic minerals in a rock by coercivity and unblocking temperature properties. Geophys. Res. Lett. 1990, 17, 159–162. [Google Scholar] [CrossRef]
- Hrouda, F. Magnetic anisotropy of rocks and its application in geology and geophysics. Geophys. Surv. 1982, 5, 37–82. [Google Scholar] [CrossRef]
- Tarling, D.H.; Hrouda, F. The Magnetic Anisotropy of Rocks; Chapman & Hall: London, UK, 1993; 217p. [Google Scholar]
- Hrouda, F.; Janak, F. The changes in shape of the magnetic susceptibility ellipsoid during progressive metamorphism and deformation. Tectonophysics 1976, 34, 135–148. [Google Scholar] [CrossRef]
- Kraus, M.J.; Aslan, A. Eocene hydromorphic paleosols: Significance for interpreting ancient floodplain processes. J. Sed. Petrol. 1993, 63, 453–463. [Google Scholar]
- Mancini, M.; Marini, M.; Moscatelli, M.; Stigliano, F.; Cavinato, G.P.; Di Salvo, C.; Simionato, M. Stratigraphy of the Palatine hill (Rome, italy): A record of repeated middle pleistocene-holocene paleovalley incision and infill. Alp. Mediterr. Quat. 2018, 31, 171–194. [Google Scholar]
- Nummendal, D.; Swift, D.J.P. Transgressive Stratigraphy at Sequence-Bounding Unconformities: Some Principles Derived from Holocene and Creataceous Examples; SEPM Special Publication: Tulsa, OK, USA, 1987; Volume 41, pp. 241–260. [Google Scholar]
- Siggerud, E.I.H.; Steel, R.J.; Pollard, J.E. Bored pebbles and ravinement surface clusters in a transgressive systems tract, Sant Llorenç del Munt fan-delta complex, SE Ebro Basin, Spain. Sediment. Geol. 2000, 138, 161–177. [Google Scholar] [CrossRef]
- Hwang, I.G.; Heller, P.L. Anatomy of a transgressive lag: Panther Tongue Sandstone, Star Point Formation, central Utah. Sedimentology 2002, 49, 977–999. [Google Scholar] [CrossRef]
- Cattaneo, A.; Steel, R.J. Transgressive deposits: A review of their variability. Earth-Sci. Rev. 2003, 62, 187–228. [Google Scholar] [CrossRef]
- Ambrosetti, E.; Martini, I.; Sandrelli, F. Shoal-water deltas in high-accommodation settings: Insights from the lacustrine Valimi Formation (Gulf of Corinth, Greece). Sedimentology 2017, 64, 425–452. [Google Scholar] [CrossRef]
- Colombera, L.; Mountney, N.P. On the geological significance of clastic parasequences. Earth-Sci. Rev. 2020, 201, 103062. [Google Scholar] [CrossRef]
- Tauxe, L.; Mullender, T.A.T.; Pick, T. Potbellies, wasp-waists, and superparamagnetism in magnetic hysteresis. J. Geophys. Res. Solid Earth 1996, 101, 571–583. [Google Scholar] [CrossRef]
- Tauxe, L.; Bertram, H.N.; Seberino, C. Physical interpretation of hysteresis loops: Micromagnetic modeling of fine particle magnetite. Geochem. Geophys. Geosyst. 2002, 3, 1–22. [Google Scholar] [CrossRef]
- Day, R.; Fuller, M.; Schmidt, V.A. Hysteresis properties of titanomagnetites: Grain-size and compositional dependence. Phys. Earth Planet. Int. 1977, 13, 260–267. [Google Scholar] [CrossRef]
- Dunlop, D.J. Theory and application of the Day plot (Mrs/Ms versus Hcr/Hc) 1. Theoretical curves and tests using titanomagnetite data. J. Geophys. Res. 2002, 107, 1–22. [Google Scholar]
- Roberts, A.P.; Chang, L.; Rowan, C.J.; Horng, C.S.; Florindo, F. Magnetic properties of sedimentary greigite (Fe3S4): An update. Rev. Geophys. 2011, 49, RG1002. [Google Scholar] [CrossRef] [Green Version]
- Tauxe, L.; Banerjee, S.K.; Butler, R.F.; van der Voo, R. Essentials of Paleomagnetism. 5th Web Edition. 2018. Available online: https://earthref.org/MagIC/books/Tauxe/Essentials/ (accessed on 13 February 2023).
- Fabbrini, A.; Baldassini, N.; Caricchi, C.; Foresi, L.M.; Sagnotti, L.; Dinarés-Turell, J.; Di Stefano, A.; Lirer, F.; Menichetti, M.; Winkler, A.; et al. In search of the Burdigalian GSSP: New evidence from the Contessa Section (Italy). Ital. J. Geosci. 2019, 138, 274–295. [Google Scholar] [CrossRef]
- Di Stefano, A.; Baldassini, N.; Raffi, I.; Fornaciari, E.; Incarbona, A.; Negri, A.; Bonomo, S.; Villa, G.; Di Stefano, E.; Rio, D. Neogene-Quaternary Mediterranean Calcareous Nannofossil Biozonation and Biochronology. Stratigraphy, 2023; in press. [Google Scholar]
- Iaccarino, S.; Castradori, D.; Cita, M.B.; Di Stefano, E.; Gaboardi, S.; McKenzie, J.A.; Spezzaferri, S.; Sprovieri, R. The Miocene/Pliocene boundary and the significance of the earliest Pliocene flooding in the Mediterranean. Mem. Soc. Geol. Ital. 1999, 54, 109–131. [Google Scholar]
- Maniscalco, R.; Casciano, C.I.; Distefano, S.; Grossi, F.; Di Stefano, A. Facies analysis in the Second Cycle Messinian evaporites predating the early Pliocene reflooding: The Balza Soletta section (Corvillo Basin, central Sicily). Ital. J. Geosci. 2019, 138, 301–316. [Google Scholar] [CrossRef]
- Cita, M.B. Micropaleontologia; Cisalpino Goliardica: Milano, Italy, 1983. [Google Scholar]
- Van der Zwaan, G.J.; Jorissen, F.J.; De Stigter, H.C. The depth dependency of planktonic/benthic foraminiferal ratios: Constraints and applications. Mar. Geol. 1990, 95, 1–16. [Google Scholar] [CrossRef]
- Murray, J.W. Ecology and Applications of Benthic Foraminifera; Cambridge University Press: New York, NY, USA, 2006; pp. 1–426. [Google Scholar]
- Dominici, S.; Mazzanti, R.; Nencini, C. Geologia dei dintorni di San Miniato tra l’Arno, l’Elsa e l’Era. Quaderni del Museo di Storia Naturale di Livorno 1995, 14 (Suppl. S1), 1–35. [Google Scholar]
- Bossio, A.; Foresi, L.M.; Mazzanti, R.; Mazzei, R.; Salvatorini, G. Note micropaleontologiche sulla successione miocenica del Torrente Morra e su quella Pliocenica del Bacino dei Fiumi Tora e Fine (Provincie di Livorno e Pisa). Atti Soc. Toscana Sci. Nat. 1997, 104, 85–134. [Google Scholar]
- Lirer, F.; Antonio, C.; Claudia, C.; Elena, T.; Francisco, S.; Salvatorini, G.; Turco, E.; Cosentino, C.; Sierro, F.J.; Caruso, A. Mediterranean Neogene planktonic foraminifer biozonation and biochronology. Earth Sci. Rev. 2019, 196, 102869. [Google Scholar] [CrossRef]
- Raffi, I.; Wade, B.S.; Palike, H.; Beu, A.G.; Cooper, R.; Crundwell, M.P.; Krijgsman, W.; Moore, T.; Raine, I.; Sardella, R.; et al. The Neogene Period. In The Geological Time Scale 2020; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 29, pp. 1141–1215. [Google Scholar]
- Lourens, L.J.; Hilgen, F.J.; Shackleton, N.J.; Laskar, J.; Wilson, D. The Neogene Period. In A Geologic Time Scale 2004; Gradstein, F., Ed.; Cambridge University Press: Cambridge, UK, 2004; Volume 21, pp. 409–440. [Google Scholar]
- Florindo, F.; Sagnotti, L. Palaeomagnetism and rock magnetism in the upper Pliocene Valle Ricca (Rome, Italy) section. Geophys. J. Int. 1995, 123, 340–354. [Google Scholar] [CrossRef] [Green Version]
- Channel, J.E.T.; Poli, M.S.; Rio, D.; Sprovieri, R.; Villa, G. Magnetic stratigraphy and biostratigraphy of Pliocene “argille azzurre” (Northern Apennines, Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 1994, 110, 83–102. [Google Scholar] [CrossRef]
- Bossio, A.; Costantini, A.; Foresi, L.M.; Lazzarotto, A.; Mazzanti, R.; Mazzei, R.; Pascucci, V.; Salvatorini, G.; Sandrelli, F.; Terzuoli, A. Space-time sedimentary evolution of the Neogene-Quaternary sediments of the western side of the Northern Apennines (Italy). Mem. Soc. Geol. Ital. 1998, 52, 513–525. [Google Scholar]
- Blanc-Vernet, L. Contribution à l'étude des Foraminifères de Mèditerranée. Rec. Trav. St. Mar. Endoume 1969, 48, 5–281. [Google Scholar]
- Jorissen, F.J. The distribution of benthic foraminifera in the Adriatic Sea. Mar. Micropaleontol. 1987, 12, 21–48. [Google Scholar] [CrossRef]
- Sen Gupta, B.K. Foraminifera in marginal marine environments. In Modern Foraminifera; Sen Gupta, B.K., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; pp. 141–159. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Directions | ||||||||
|---|---|---|---|---|---|---|---|---|
| In Situ | Tilt-Corrected | |||||||
| N | k | a95 | Dec. | Inc. | k | a95 | Dec. | Inc. |
| 60 | 17.4 | 4.5° | 353.1°E | 57.9° | 17.4 | 4.5° | 328.7° | 58.3° |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marini, M.; Foresi, L.M.; Barbagallo, V.; Bisconti, M.; Di Stefano, A.; Muttoni, G.; Martini, I. Age and Depositional Environment of Whale-Bearing Sedimentary Succession from the Lower Pliocene of Tuscany (Italy): Insights from Palaeomagnetism, Calcareous Microfossils and Facies Analyses. J. Mar. Sci. Eng. 2023, 11, 455. https://doi.org/10.3390/jmse11020455
Marini M, Foresi LM, Barbagallo V, Bisconti M, Di Stefano A, Muttoni G, Martini I. Age and Depositional Environment of Whale-Bearing Sedimentary Succession from the Lower Pliocene of Tuscany (Italy): Insights from Palaeomagnetism, Calcareous Microfossils and Facies Analyses. Journal of Marine Science and Engineering. 2023; 11(2):455. https://doi.org/10.3390/jmse11020455
Chicago/Turabian StyleMarini, Mattia, Luca Maria Foresi, Viviana Barbagallo, Michelangelo Bisconti, Agata Di Stefano, Giovanni Muttoni, and Ivan Martini. 2023. "Age and Depositional Environment of Whale-Bearing Sedimentary Succession from the Lower Pliocene of Tuscany (Italy): Insights from Palaeomagnetism, Calcareous Microfossils and Facies Analyses" Journal of Marine Science and Engineering 11, no. 2: 455. https://doi.org/10.3390/jmse11020455