
Calcareous Nannofossil Biostratigraphy and Biochronology at ODP Site 1123 (Offshore New Zealand): A Reference Section for the Last 20 Myr in the Southern Ocean
 , , , ,
, , , ,
Abstract
:1. Introduction
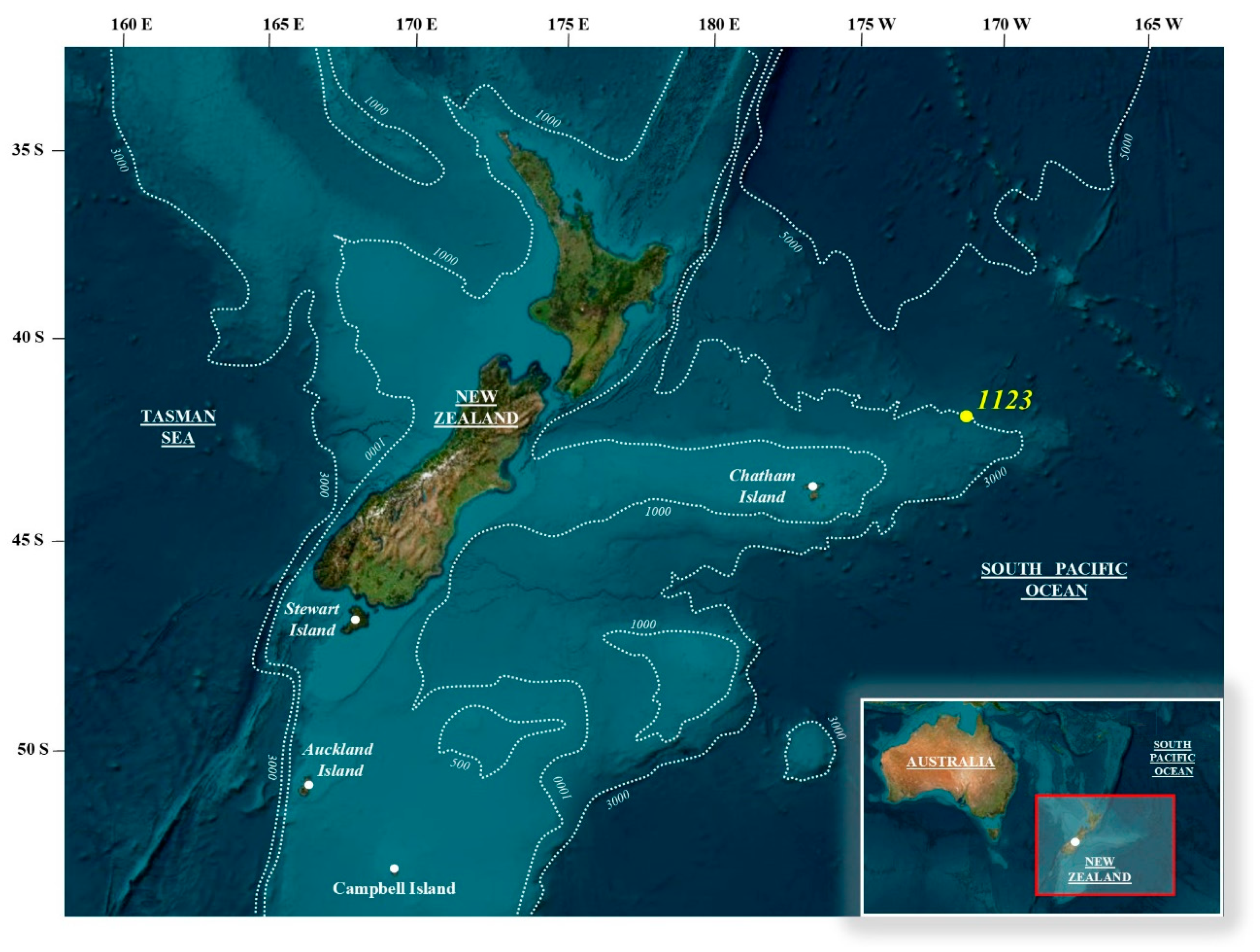
2. Materials and Methods


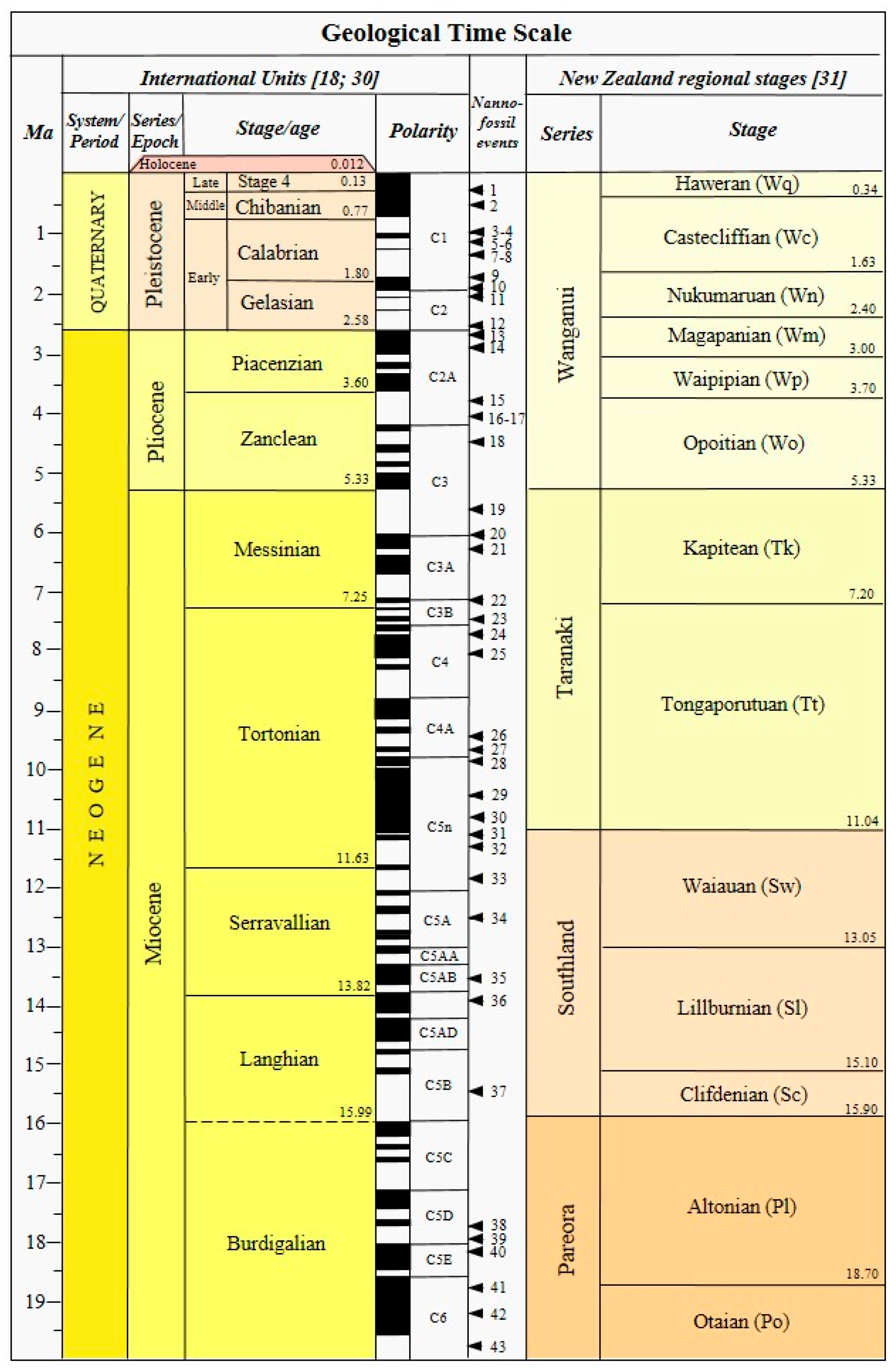
3. Neogene–Quaternary Calcareous Nannofossils Biostratigraphy: State of the Art
3.1. Holocene–Pleistocene (Holocene–Gelasian = Haweran–Nukumaruan)
3.2. Pliocene–Late Miocene (Piacenzian–Tortonian = Mangapanian–Tongaporutuan)
3.3. Middle–Early Miocene (Serravallian–Burdigalian = Waiauan–Otaian)
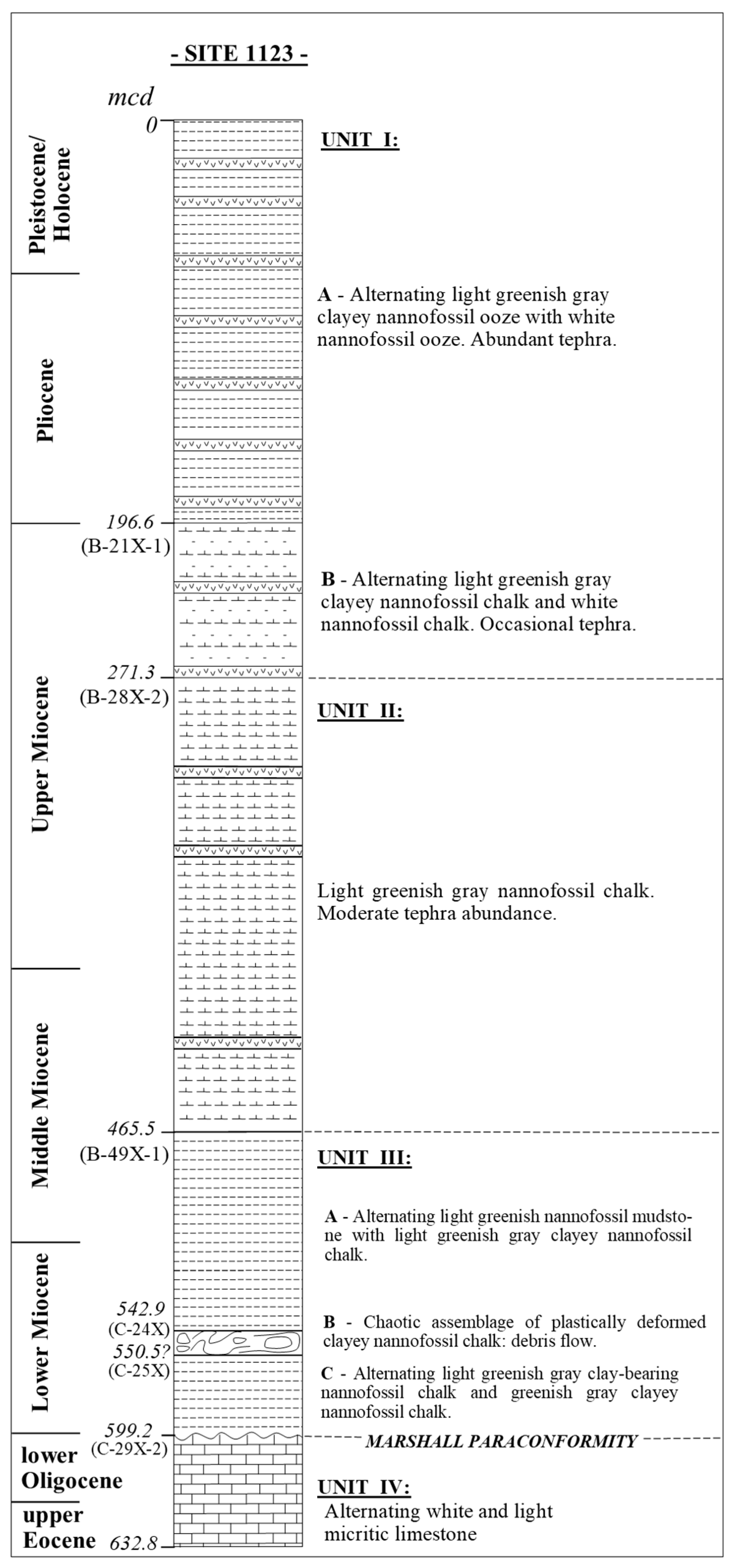
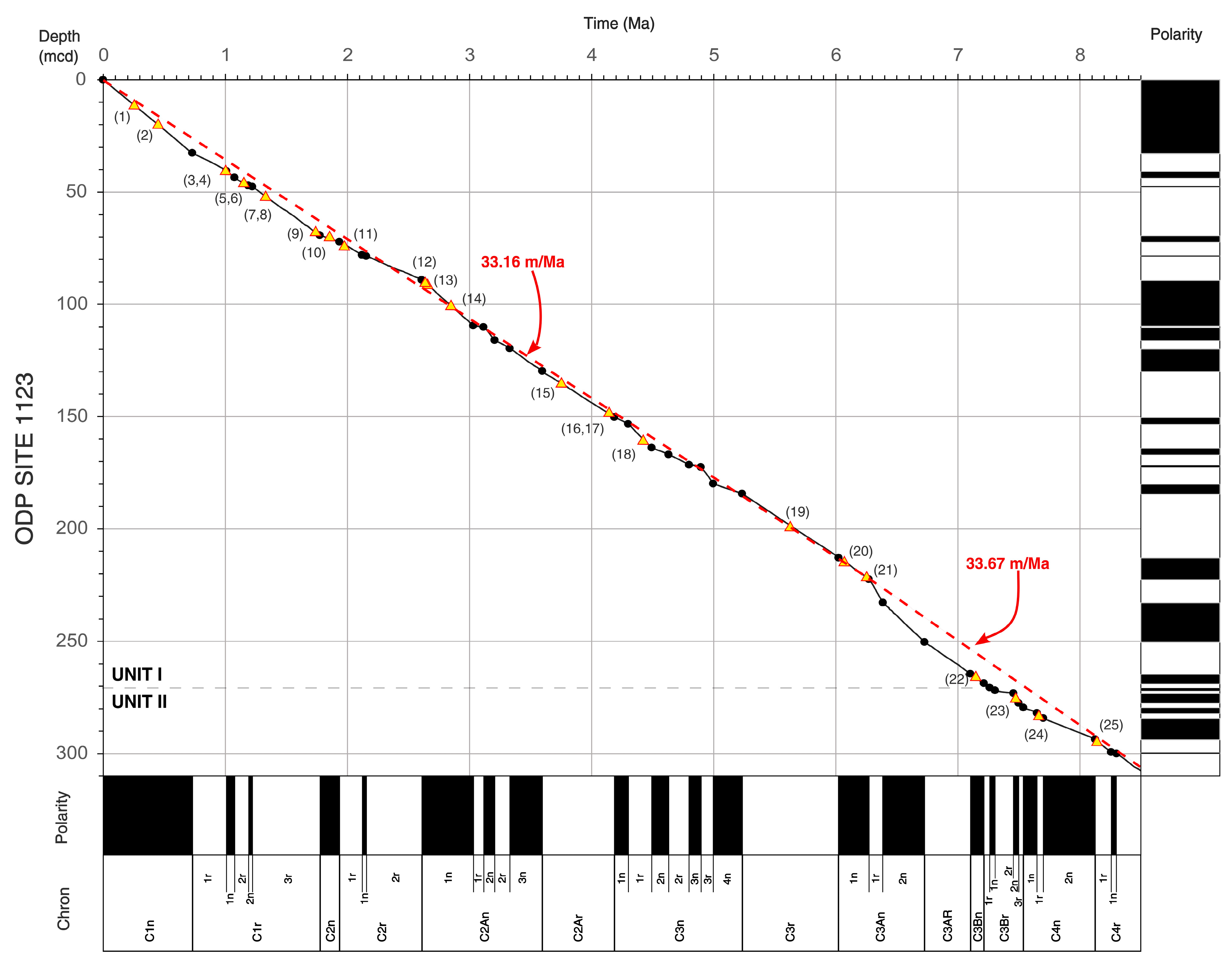
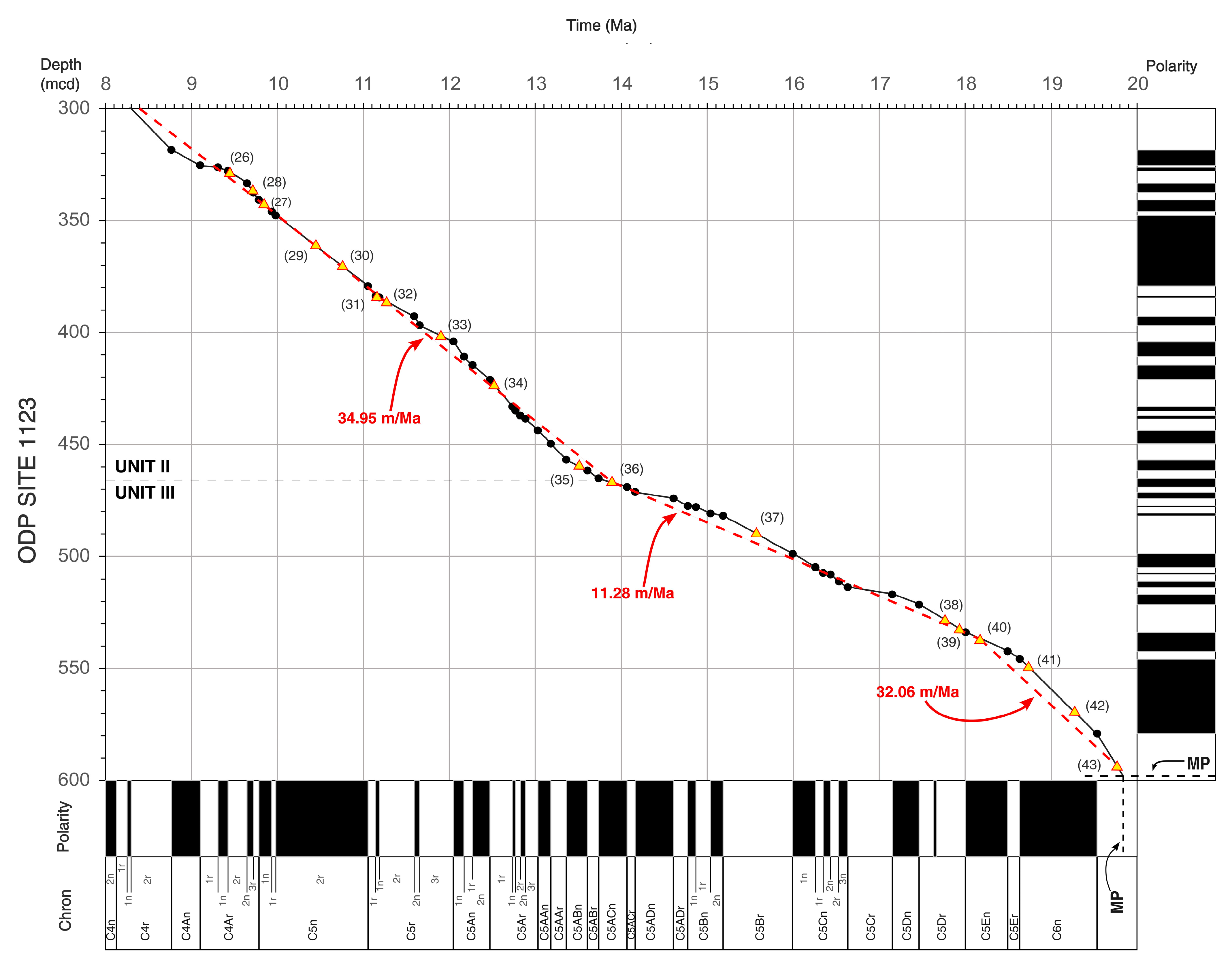
4. Calcareous Nannofossils Biostratigraphy and Biochronology at ODP Site 1123
4.1. Holocene–Pleistocene (Holocene–Gelasian = Haweran–Nukumaruan)
4.2. Pliocene–Late Miocene (Piacenzian–Tortonian = Mangapanian–Tongaporutuan)
4.3. Middle–Early Miocene (Serravallian–Burdigalian = Waiauan–Otaian)
5. Conclusive Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Figures of Nannofossils Index Markers at ODP Site 1123



References
- Shipboard Scientific Party. Site 1123: North Chatham Drift: A 20-Ma Record of the Pacific Deep Western Boundary Current. In Proceedings of the Ocean Drilling Program 181 Initial Reports. 2000. Available online: http://www-odp.tamu.edu/publications/181_IR/chap_07/chap_07.htm (accessed on 8 December 2022).
- Carter, R.M. The Mid-Oligocene Marshall Paraconformity, New Zealand: Coincidence with Global Eustatic Sea-Level Fall or Rise? J. Geol. 1985, 93, 359–371. [Google Scholar] [CrossRef]
- Fulthorpe, C.S.; Carter, R.M.; Miller, K.G.; Wilson, J. Marshall Paraconformity: A Mid-Oligocene Record of Inception of the Antarctic Circumpolar Current and Coeval Glacio-Eustatic Lowstand? Mar. Pet. Geol. 1996, 13, 61–77. [Google Scholar] [CrossRef]
- Mildenhall, D.C.; Hollis, C.J.; Naish, T.R. Orbitally-Influenced Vegetation Record of the Mid-Pleistocene Climate Transition, Offshore Eastern New Zealand (ODP Leg 181, Site 1123). Mar. Geol. 2004, 205, 87–111. [Google Scholar] [CrossRef]
- Fenner, J.; Di Stefano, A. Late Quaternary Oceanic Fronts along Chatham Rise Indicated by Phytoplankton Assemblages, and Refined Calcareous Nannofossil Stratigraphy for the Mid-Latitude SW Pacific. Mar. Geol. 2004, 205, 59–86. [Google Scholar] [CrossRef]
- Weedon, G.; Hall, I. Neogene Palaeoceanography of Chatham Rise (Southwest Pacific) Based on Sediment Geochemistry. Mar. Geol. 2004, 205, 207–225. [Google Scholar] [CrossRef]
- Crundwell, M.; Scott, G.; Naish, T.; Carter, L. Glacial–Interglacial Ocean Climate Variability from Planktonic Foraminifera during the Mid-Pleistocene Transition in the Temperate Southwest Pacific, ODP Site 1123. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 260, 202–229. [Google Scholar] [CrossRef]
- Christiansen, K.J. Paleo-Oceanographic and-Climatic Reconstruction in the Southwest Pacific [ODP Site 1123] during MIS 11. Master’s Thesis, Victoria University of Wellington, Wellington, New Zealand, 2012; pp. 1–137. [Google Scholar]
- Ford, H.L.; Raymo, M.E. Regional and global signals in seawater δ18O records across the mid-Pleistocene transition. Geology 2020, 48, 113–117. [Google Scholar] [CrossRef]
- Elderfield, H.; Ferretti, P.; Greaves, M.; Crowhurst, S.; McCave, I.N.; Hodell, D.; Piotrowski, A.M. Evolution of Ocean Temperature and Ice Volume Through the Mid-Pleistocene Climate Transition. Science (1979) 2012, 337, 704–709. [Google Scholar] [CrossRef]
- Venuti, A.; Florindo, F.; Michel, E.; Hall, I.R. Magnetic Proxy for the Deep (Pacific) Western Boundary Current Variability across the Mid-Pleistocene Climate Transition. Earth Planet. Sci. Lett. 2007, 259, 107–118. [Google Scholar] [CrossRef]
- Cobianchi, M.; Mancin, N.; Lupi, C.; Bordiga, M.; Bostock, H.C. Effects of Oceanic Circulation and Volcanic Ash-Fall on Calcite Dissolution in Bathyal Sediments from the SW Pacific Ocean over the Last 550ka. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 429, 72–82. [Google Scholar] [CrossRef]
- Horn, M.; Uenzelmann-Neben, G. The Deep Western Boundary Current at the Bounty Trough, East of New Zealand: Indications for Its Activity Already before the Opening of the Tasmanian Gateway. Mar Geol 2015, 362, 60–75. [Google Scholar] [CrossRef]
- Beltran, C.; Ohneiser, C.; Hageman, K.J.; Scanlan, E. Evolution of the Southwestern Pacific Surface Waters during the Early Pleistocene. New Zealand J. Geol. Geophys. 2016, 59, 514–521. [Google Scholar] [CrossRef]
- Toyos, M.H.; Lamy, F.; Lange, C.B.; Lembke-Jene, L.; Saavedra-Pellitero, M.; Esper, O.; Arz, H.W. Antarctic Circumpolar Current Dynamics at the Pacific Entrance to the Drake Passage Over the Past 1.3 Million Years. Paleoceanogr. Paleoclimatol. 2020, 35, e2019PA003773. [Google Scholar] [CrossRef]
- Mcgonigal, K.; Di Stefano, A. Calcareous Nannofossil Biostratigraphy of the Eocene–Oligocene Transition, ODP Sites 1123 and 1124. Ocean Drill. Program Sci. Results 2002, 181, 1–22. [Google Scholar] [CrossRef]
- Crundwell, M.; Nelson, C. A Magnetostratigraphically-Constrained Chronology for Late Miocene Bolboformids and Planktic Foraminifers in the Temperate Southwest Pacific. Stratigraphy 2007, 4, 1–34. [Google Scholar]
- Raffi, I.; Wade, B.S.; Pälike, H.; Beu, A.G.; Cooper, R.; Crundwell, M.P.; Krijgsman, W.; Moore, T.; Raine, J.I.; Sardella, R.; et al. The Neogene Period. In Geologic Time Scale; Elsevier B.V.: Amsterdam, The Netherlands, 2020; pp. 1141–1215. [Google Scholar]
- Bown, P.R.; Young, J. Introduction. In Calcareous Nannofossil Biostratigraphy; Bown, P.R., Ed.; Kluwer Academic Publisher: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1998; pp. 1–15. [Google Scholar]
- Aubry, M.-P. Handbook of Cenozoic Calcareous Nannoplankton; Micropaleontology Press: New York, NY, USA, 1984–1990; Volume 1–5. [Google Scholar]
- Perch-Nielsen, K. Cenozoic Calcareous Nannofossils. In Plankton Stratigraphy; Bolli, H.M., Saunders, J.B., Perch-Nielsen, K., Eds.; Cambridge University Press: Cambridge, UK, 1985; pp. 427–554. [Google Scholar]
- Bown, P. (Ed.) Calcareous Nannofossil Biostratigraphy; Kluwer Academic Publisher: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1998; pp. 1–315. [Google Scholar]
- Young, J.R.; Geisen, M.; Cros, L.; Kleijne, A.; Sprengel, C.; Probert, I.; Østergaard, J. A Guide to Extant Coccolithophore Taxonomy. J. Nannoplankton Res. 2003, 1–132. [Google Scholar]
- Jordan, R.W.; Cros, L.; Young, J.R. A Revised Classification Scheme for Living Haptophytes. Micropaleontology 2004, 50 (Suppl. 1), 55–79. [Google Scholar] [CrossRef]
- Young, J.R.; Bown, P.R.; Lees, J.A. Nannotax3 Website. 2022. Available online: https://www.mikrotax.org/Nannotax3 (accessed on 21 April 2022).
- Backman, J.; Shackleton, N.J. Quantitative Biochronology of Pliocene and Early Pleistocene Calcareous Nannofossils from the Atlantic, Indian and Pacific Oceans. Mar. Micropaleontol. 1983, 8, 141–170. [Google Scholar] [CrossRef]
- Rio, D.; Fornaciari, E.; Raffi, I. Late Oligocene through Early Pleistocene Calcareous Nannofossil from Eastern Equatorial Indian Ocean. Proc. Ocean Drill. Program Sci. Res. 1990, 115, 175–221. [Google Scholar]
- Fornaciari, E.; Rio, D. Latest Oligocene to Early Middle Miocene Quantitative Calcareous Nannofossil Biostratigraphy in the Mediterranean Region. Micropaleontology 1996, 42, 1–36. [Google Scholar] [CrossRef]
- Backman, J.; Raffi, I.; Rio, D.; Fornaciari, E.; Pälike, H. Biozonation and Biochronology of Miocene through Pleistocene Calcareous Nannofossils from Low and Middle Latitudes. Newsl. Stratigr. 2012, 45, 221–244. [Google Scholar] [CrossRef]
- Pillans, B.; Gibbard, P. The Quaternary Period. In The Geologic Time Scale 2012; Gradstein, F.M., Ogg, J.G., Schmitz, M., Ogg, G., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2012; Volume 2, pp. 979–1010. [Google Scholar]
- Raine, J.I.; Beu, A.G.; Boyes, A.F.; Campbell, H.J.; Cooper, R.A.; Crampton, J.S.; Crundwell, M.P.; Hollis, C.J.; Morgans, H.E.G.; Mortimer, N. New Zealand Geological Timescale NZGT 2015/1. N. Z. J. Geol. Geophys. 2015, 58, 398–403. [Google Scholar] [CrossRef]
- Martini, E. Standard Tertiary and Quaternary Calcareous Nannoplankton Zonation. In Proceedings of the Proc. 2nd International Conference on Planktonic Microfossils; Farinacci, A., Ed.; Edizioni Tecnoscienza: Rome, Italy, 1971; pp. 739–785. [Google Scholar]
- Okada, H.; Bukry, D. Supplementary Modification and Introduction of Code Numbers to the Low-Latitude Coccolith Biostratigraphy. Mar. Micropaleontol. 1980, 5, 321–325. [Google Scholar] [CrossRef]
- Bramlette, M.N.; Riedel, W.R. Stratigraphic Value of Discoasters and Some Other Microfossils Related to Recent Coccolithophores. J. Paleontol. 1954, 28, 385–403. [Google Scholar]
- Bramlette, M.N.; Wilcoxon, J.A. Middle Tertiary Calcareous Nannoplankton of the Cipero Section, Trinidad, WI. Tulane Stud. Geol. Paleontol. 1967, 5, 421–485. [Google Scholar]
- Rio, D. The Fossil Distribution of Coccolithophore Genus Gephyrocapsa Kamptner and Related Plio-Pleistocene Chronostratigraphic Problems. Deep Sea Drill. Proj. Initial Rep. 1982, 68, 325–343. [Google Scholar]
- Raffi, I.; Backman, J.; Rio, D.; Shackleton, N.J. Plio-Pleistocene Nannofossil Biostratigraphy and Calibration to Oxygen Isotope Stratigraphies from Deep Sea Drilling Project Site 607 and Ocean Drilling Program Site 677. Paleoceanography 1993, 8, 387–408. [Google Scholar] [CrossRef]
- Naish, T.R.; Abbott, S.T.; Alloway, B.V.; Beu, A.G.; Carter, M.R.; Edwards, A.R.; Journeaux, D.T.; Kamp, P.J.J.; Pillans, B.J.; Saul, G.; et al. Astronomical Calibration of a Southern Hemisphere Plio-Pleistocene Reference Section, Wanganui Basin, New Zealand. Quat. Sci. Rev. 1998, 17, 695–710. [Google Scholar] [CrossRef]
- Incarbona, A.; Sprovieri, M.; Di Stefano, A.; Di Stefano, E.; Manta, D.S.; Pelosi, N.; Ribera D’Alcalà, M.; Sprovieri, R.; Ziveri, P. Productivity Modes in the Mediterranean Sea during Dansgaard-Oeschger (20,000-70,000yr Ago) Oscillations. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 392, 128–137. [Google Scholar] [CrossRef]
- Sprovieri, R.; Di Stefano, E.; Howell, M.; Sakamoto, T.; Di Stefano, A.; Marino, M. Integrated Calcareous Plankton Biostratigraphy and Cyclostratigraphy at Site 964. Proc. Ocean Drill. Program Sci. Res. 1998, 160, 155–166. [Google Scholar]
- Suganuma, Y.; Okada, M.; Head, M.J.; Kameo, K.; Haneda, Y.; Hayashi, H.; Irizuki, T.; Itaki, T.; Izumi, K.; Kubota, Y.; et al. Formal Ratification of the Global Boundary Stratotype Section and Point (GSSP) for the Chibanian Stage and Middle Pleistocene Subseries of the Quaternary System: The Chiba Section, Japan. Epis. J. Int. Geosci. 2021, 44, 317–347. [Google Scholar] [CrossRef]
- Raffi, I. Revision of the Early-Middle Pleistocene Calcareous Nannofossil Biochronology (1.75–0.85 Ma). Mar. Micropaleontol. 2002, 45, 25–55. [Google Scholar] [CrossRef]
- Lourens, L.J.; Antonarakou, A.; Hilgen, F.J.; Van Hoof, A.A.M.; Vergnaud-Grazzini, C.; Zachariasse, W.J. Evaluation of the Plio-Pleistocene Astronomical Timescale. Paleoceanography 1996, 11, 391–413. [Google Scholar] [CrossRef]
- Raffi, I.; Rio, D.; D’Atri, A.; Fornaciari, E.; Rocchetti, S. Quantitative distribution patterns and biomagnetostratigraphy of Middle and Late Miocene calcareous nannofossils from equatorial Indian and Pacific Oceans (legs 115, 130 and 138). Proc. Ocean Drill. Program Sci. Res. 1995, 138, 479–502. [Google Scholar]
- Cita, M.B.; Gibbard, P.L.; Head, M.J. Formal Ratification of the GSSP for the Base of the Calabrian Stage (Second Stage of the Pleistocene Series, Quaternary System). Epis. J. Int. Geosci. 2012, 35, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Lourens, L.; Hilgen, F.; Shackleton, N.J.; Laskar, J.; Wilson, D. The Neogene Period. In A Geologic Time Scale 2004; Smith, A.G., Gradstein, F.M., Ogg, J.G., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 409–440. ISBN 9780521786737. [Google Scholar]
- Hornibrook, N.D.B. Correlation of Pliocene Biostratigraphy, Magnetostratigraphy and O18 fluctuations in New Zealand and DSDP Site 284. Newsl. Stratigr. 1980, 9, 114–120. [Google Scholar] [CrossRef]
- Rio, D.; Sprovieri, R.; Di Stefano, E. The Gelasian Stage: A Proposal of a New Chronostratigraphic Unit of the Pliocene Series. Riv. Ital. Paleontol. Stratigr. 1994, 100, 103–124. [Google Scholar]
- Rio, D.; Sprovieri, R.; Castradori, D.; Di Stefano, E. The Gelasian Stage (Upper Pliocene): A New Unit of the Global Standard Chronostratigraphic Scale. Epis. J. Int. Geosci. 1998, 21, 82–87. [Google Scholar] [CrossRef]
- Sprovieri, R. Pliocene-Early Pleistocene Astronomically Forced Planktonic Foraminifera Abundance Fluctuations and Chronology of Mediterranean Calcareous Plankton Bio-Events. Riv. Ital. Paleontol. Stratigr. 1993, 99, 371–414. [Google Scholar]
- Castradori, D.; Rio, D.; Hilgen, F.J.; Lourens, L.J. The Global Standard Stratotype-Section and Point (GSSP) of the Piacenzian Stage (Middle Pliocene). Epis. J. Int. Geosci. 1998, 21, 88–93. [Google Scholar] [CrossRef]
- Gartner, S. Neogene Calcareous Nannofossil Biostratigraphy, Leg 116 (Central Indian Ocean). Proc. Ocean Drill. Program Sci. Res. 1990, 116, 165–187. [Google Scholar]
- Di Stefano, A.; Sturiale, G. Refinements of Calcareous Nannofossil Biostratigraphy at the Miocene/Pliocene Boundary in the Mediterranean Region. Geobios 2010, 449, 5–20. [Google Scholar] [CrossRef]
- Van Couvering, J.A.; Castradori, D.; Cita, M.B.; Hilgen, F.J.; Rio, D. The Base of the Zanclean Stage and of the Pliocene Series. Epis. J. Int. Geosci. 2000, 23, 179–187. [Google Scholar] [CrossRef]
- Raffi, I.; Mozzato, C.; Fornaciari, E.; Hilgen, F.J.; Rio, D. Late Miocene Calcareous Nannofossil Biostratigraphy and Astrobiochronology for the Mediterranean Region. Micropaleontology 2003, 49, 1–26. [Google Scholar] [CrossRef]
- Morigi, C.; Negri, A.; Giunta, S.; Kouwenhoven, T.; Krijgsman, W.; Blanc-Valleron, M.-M.; Orszag-Sperber, F.; Rouchy, J.-M. Integrated Quantitative Biostratigraphy of the Latest Tortonian–Early Messinian Pissouri Section (Cyprus): An Evaluation of Calcareous Plankton Bioevents. Geobios 2007, 40, 267–279. [Google Scholar] [CrossRef]
- Hilgen, F.J.; Iaccarino, S.; Krijgsman, W.; Villa, G.; Langereis, C.G.; Zachariasse, W.J. The Global Boundary Stratotype Section and Point (GSSP) of the Messinian Stage (Uppermost Miocene). Epis. J. Int. Geosci. 2000, 23, 172–178. [Google Scholar] [CrossRef]
- Zeeden, C.; Hilgen, F.; Westerhold, T.; Lourens, L.; Röhl, U.; Bickert, T. Revised Miocene Splice, Astronomical Tuning and Calcareous Plankton Biochronology of ODP Site 926 between 5 and 14.4Ma. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 369, 430–451. [Google Scholar] [CrossRef]
- Hilgen, F.J.; Krijgsman, W.; Raffi, I.; Turco, E.; Zachariasse, W.J. Integrated Stratigraphy and Astronomical Calibration of the Serravallian/Tortonian Boundary Section at Monte Gibliscemi (Sicily, Italy). Mar. Micropaleontol. 2000, 38, 181–211. [Google Scholar] [CrossRef]
- Hilgen, F.; Aziz, H.A.; Bice, D.; Iaccarino, S.; Krijgsman, W.; Kuiper, K.; Montanari, A.; Raffi, I.; Turco, E.; Zachariasse, W.-J. The Global Boundary Stratotype Section and Point (GSSP) of the Tortonian Stage (Upper Miocene) at Monte Dei Corvi. Epis. J. Int. Geosci. 2005, 28, 6–17. [Google Scholar] [CrossRef]
- Lirer, F.; Caruso, A.; Foresi, L.M.; Sprovieri, M.; Bonomo, S.; Di Stefano, A.; Di Stefano, E.; Iaccarino, S.M.; Salvatorini, G.; Sprovieri, R.; et al. Astrochronological Calibration of the Upper Serravallian/Lower Tortonian Sedimentary Sequence at Tremiti Islands (Adriatic Sea, Southern Italy). Riv. Ital. Paleontol. Stratigr. 2002, 108, 241–256. [Google Scholar]
- Abels, H.A.; Hilgen, F.J.; Krijgsman, W.; Kruk, R.W.; Raffi, I.; Turco, E.; Zachariasse, W.J. Long-Period Orbital Control on Middle Miocene Global Cooling: Integrated Stratigraphy and Astronomical Tuning of the Blue Clay Formation on Malta. Paleoceanography 2005, 20, 1–20. [Google Scholar] [CrossRef]
- Hilgen, F.J.; Abels, H.A.; Iaccarino, S.; Krijgsman, W.; Raffi, I.; Sprovieri, R.; Turco, E.; Zachariasse, W.J. The Global Stratotype Section and Point (GSSP) of the Serravallian Stage (Middle Miocene). Epis. J. Int. Geosci. 2009, 32, 152–166. [Google Scholar] [CrossRef] [Green Version]
- Turco, E.; Hilgen, F.; Raffi, I.; Di Stefano, A.; Foresi, L.M.; Holbourn, A.; Iaccarino, S.; Lirer, F. The Global Stratotype Section and Point (GSSP) of the Langhian Stage (Middle Miocene): A Proposal. Submitt. IUG-ICS-Subcomm. Neogene Stratigr. 2023, 1–37. [Google Scholar]
- Foresi, L.M.; Baldassini, N.; Sagnotti, L.; Lirer, F.; Di Stefano, A.; Caricchi, C.; Verducci, M.; Salvatorini, G.; Mazzei, R. Integrated Stratigraphy of the St. Thomas Section (Malta Island): A Reference Section for the Lower Burdigalian of the Mediterranean Region. Mar. Micropaleontol. 2014, 111, 66–89. [Google Scholar] [CrossRef]
- Haq, B.U.; Hardenbol, J.; Vail, P.R. Chronology of Fluctuating Sea Levels since the Triassic. Science 1987, 235, 1156–1167. [Google Scholar] [CrossRef]
- Cande, S.C.; Kent, D. Revised Calibration of the Geomagnetic Polarity Timescale for the Late Cretaceous and Cenozoic. J. Geophys. Res. 1995, 100, 6093–6095. [Google Scholar] [CrossRef]
- Capraro, L.; Bonomo, S.; Di Stefano, A.; Ferretti, P.; Fornaciari, E.; Galeotti, S.; Incarbona, A.; Macrì, P.; Raffi, I.; Sabatino, N.; et al. The Monte San Nicola Section (Sicily) Revisited: A Potential Unit-Stratotype of the Gelasian Stage. Quat. Sci. Rev. 2022, 278, 107367. [Google Scholar] [CrossRef]
- Gamberi, F.; Della Valle, G.; Marani, M.; Mercorella, A.; Distefano, S.; Di Stefano, A. Tectonic Controls on Sedimentary System along the Continental Slope of the Central and Southeastern Tyrrhenian Sea. Ital. J. Geosci. 2019, 138, 317–332. [Google Scholar] [CrossRef]
- Maniscalco, R.; Casciano, C.I.; Distefano, S.; Grossi, F.; Di Stefano, A. Facies Analysis in the Second Cycle Messinian Evaporites Predating the Early Pliocene Reflooding: The Balza Soletta Section (Corvillo Basin, Central Sicily). Ital. J. Geosci. 2019, 138, 301–316. [Google Scholar] [CrossRef]
- Fabbrini, A.; Baldassini, N.; Caricchi, C.; Foresi, L.M.; Sagnotti, L.; Dinarés-Turell, J.; Di Stefano, A.; Lirer, F.; Menichetti, M.; Winkler, A.; et al. In Search of the Burdigalian GSSP: New Evidence from the Contessa Section (Italy). Ital. J. Geosci. 2019, 138, 274–295. [Google Scholar] [CrossRef]
- Fabbrini, A.; Baldassini, N.; Caricchi, C.; Di Stefano, A.; Dinarès-Turell, J.; Foresi, L.M.; Lirer, F.; Patricolo, S.; Sagnotti, L.; Winkler, A. Integrated Quantitative Calcareous Plankton Bio-Magnetostratigraphy of the Early Miocene from IODP Leg 342, Hole U1406A, Newfoundland Ridge, NW Atlantic Ocean. Stratigr. Geol. Correl. 2019, 27, 259–276. [Google Scholar] [CrossRef]
- Bukry, D. Low-latitude coccolith biostratigraphic zonation. Deep Sea Drill. Proj. Rep. Leg 15 1973, 15, 685–703. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Epoch | Bioevent | Marker Species | 1 | 2 | 3 | Reliability Degree | Chronostratigraphic Remarks | ||
|---|---|---|---|---|---|---|---|---|---|
| Pleistocene-Holocene | 1 | B | E. huxleyi | 0.26 | 0.29 [27] | 0.24 [38] | 0.26 [39] | good | Base Late Pleistoc. 0.13 [30] |
| 2 | T | P. lacunosa | 0.44 | 0.43 [29] | 0.46 [40] | good | Chibanian GSSP-0.77 [41] | ||
| 3 | PT | m Gephyrocapsa | 1.00 | 1.06 [42] | good | ||||
| 4 | B | G. omega | 1.00 | 0.96 [40] | good | ||||
| 5 | T | H. sellii | 1.13 | 1.24 [37] | 1.26 [40] | moderate | |||
| 6 | T | C. macintyrei | 1.13 | 1.60 [29] | 1.67 [43] | unreliable | |||
| 7 | PB | m Gephyrocapsa | 1.31 | good | |||||
| 8 | T | l Gephyrocapsa | 1.31 | 1.25 [37] | 1.24 [43] | good | |||
| 9 | B | l Gephyrocapsa | 1.72 | 1.59 [42] | 1.61 [43] | good | |||
| 10 | B | m Gephyrocapsa | 1.85 | 1.71 [42] | 1.69 [44] | 1.71 [43] | good | Calabrian GSSP-1.80 [45] | |
| 11 | T | D. brouweri | 1.96 | 1.93 [46] | 2.06 [44] ~1.93 [47] | 1.95 [43] | good | ||
| 12 | T | D. pentaradiatus | 2.61 | 2.39 [46] | 2.51 [48] | good | Gelasian GSSP-2.58 [49] | ||
| Late Miocene-Pliocene | 13 | T | D. surculus | 2.64 | 2.53 [46] | ~2.37 [47] | 2.55 [48] | good | |
| 14 | T | D. tamalis | 2.84 | 2.76 [46] | 2.87 [44] | 2.82 [40] | good | ||
| 15 | T | R. pseudoumbilicus | 3.74 | 3.82 [46] | ~3.75 [47] | 3.85 [50] | good | Piacenzian GSSP-3.60 [51] | |
| 16 | Bc | D. asymmetricus | 4.14 | 4.04 [29] | 4.11 [50] | good | |||
| 17 | B | P. lacunosa | 4.14 | 4.00 [52] | good | ||||
| 18 | Bc | H. sellii | 4.41 | 4.62 [53] | moderate | ||||
| 19 | T | D. quinqueramus | 5.61 | 5.53 [29] | good | Zanclean GSSP-5.33 [54] | |||
| 20 | T | N. amplificus | 6.07 | 5.98 [29] | 5.85 [55] | unreliable | |||
| 21 | B | N. amplificus | 6.27 | 6.82 [29] | 6.69 [55] 6.21 [56] | unreliable | |||
| 22 | B | A. delicatus | 7.15 | 7.13 [55] | good | Messinian GSSP-7.24 [57] | |||
| 23 | B | A. primus | 7.47 | 7.45 [58] | 7.39 [29] | 7.42 [55] | good | ||
| 24 | T | M. convallis | 7.68 | 7.78 [59] | 7.78 [55] | moderate | |||
| 25 | B | D. quinqueramus | 8.12 | 8.10 [44] | good | ||||
| 26 | Bc | D. pentaradiatus | 9.45 | 9.37 [55] | moderate | ||||
| 27 | T | C. coalitus | 9.84 | 9.62 [58] | 9.65 [29] | unreliable | |||
| 28 | B | M. convallis | 9.71 | 9.75 [29] | 9.23 [55] | moderate | |||
| 29 | Bc | D. bellus | 10.47 | 10.64 [58] | 10.38 [59] | moderate | |||
| 30 | B | C. coalitus | 10.77 | 10.89 [58] | 10.79 [29] | unreliable | |||
| 31 | Tc | C. miopelagicus | 11.15 | 11.04 [58] | 10.61 [29] | 10.90 [59] | moderate | ||
| 32 | Tc | D. kugleri | 11.25 | 11.61 [58] | 11.60 [29] | 11.60 [59] | unreliable | Tortonian GSSP-11.61 [60] | |
| Early-Middle Miocene | 33 | B | D. kugleri | 11.90 | 11.89 [58] | 11.88 [29] | 11.90 [59] | unreliable | |
| 34 | Tc | C. premacintyrei | 12.51 | 12.57 [29] | 12.51 [61] | good | |||
| 35 | T | S. heteromorphus | 13.53 | 13.60 [58] | 13.59 [62] | good | Serravallian GSSP-13.82 [63] | ||
| 36 | T | H. ampliaperta | 13.89 | 14.86 [29] | unreliable | ||||
| 37 | X | D.deflandrei/D.variab. | 15.58 | 15.80 [29] | moderate | Langhian GSSP-15.99 [64] | |||
| 38 | T | S. disbelemnos | 17.76 | 17.69 [65] | to be tested | ||||
| 39 | B | S. heteromorphus | 17.92 | 17.65 Bc [29] | 17.99 [65] | good | |||
| 40 | Tc | S. belemnos | 18.19 | 17.94 T [29] | 18.02 T [65] | moderate | |||
| 41 | Bc | S. belemnos | 18.73 | 19.01 B [29] | 19.12 B [65] | moderate | Base Burdigalian Stage? [66] | ||
| 42 | T | T. carinatus | 19.29 | 19.18 [29] | moderate | ||||
| 43 | AT | C. abisectus | 19.78 | to be tested | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Stefano, A.; D’Andrea, N.M.; Distefano, S.; Urso, S.; Borzì, L.; Baldassini, N.; Barbagallo, V. Calcareous Nannofossil Biostratigraphy and Biochronology at ODP Site 1123 (Offshore New Zealand): A Reference Section for the Last 20 Myr in the Southern Ocean. J. Mar. Sci. Eng. 2023, 11, 408. https://doi.org/10.3390/jmse11020408
Di Stefano A, D’Andrea NM, Distefano S, Urso S, Borzì L, Baldassini N, Barbagallo V. Calcareous Nannofossil Biostratigraphy and Biochronology at ODP Site 1123 (Offshore New Zealand): A Reference Section for the Last 20 Myr in the Southern Ocean. Journal of Marine Science and Engineering. 2023; 11(2):408. https://doi.org/10.3390/jmse11020408
Chicago/Turabian StyleDi Stefano, Agata, Natale Maria D’Andrea, Salvatore Distefano, Salvatore Urso, Laura Borzì, Niccolò Baldassini, and Viviana Barbagallo. 2023. "Calcareous Nannofossil Biostratigraphy and Biochronology at ODP Site 1123 (Offshore New Zealand): A Reference Section for the Last 20 Myr in the Southern Ocean" Journal of Marine Science and Engineering 11, no. 2: 408. https://doi.org/10.3390/jmse11020408