

A High Abundance of Holothuria (Halodeima) atra (Holothuroidea Aspidochirotida) in a Halimeda Dominated Habitat


Abstract
:1. Introduction
2. Materials and Methods
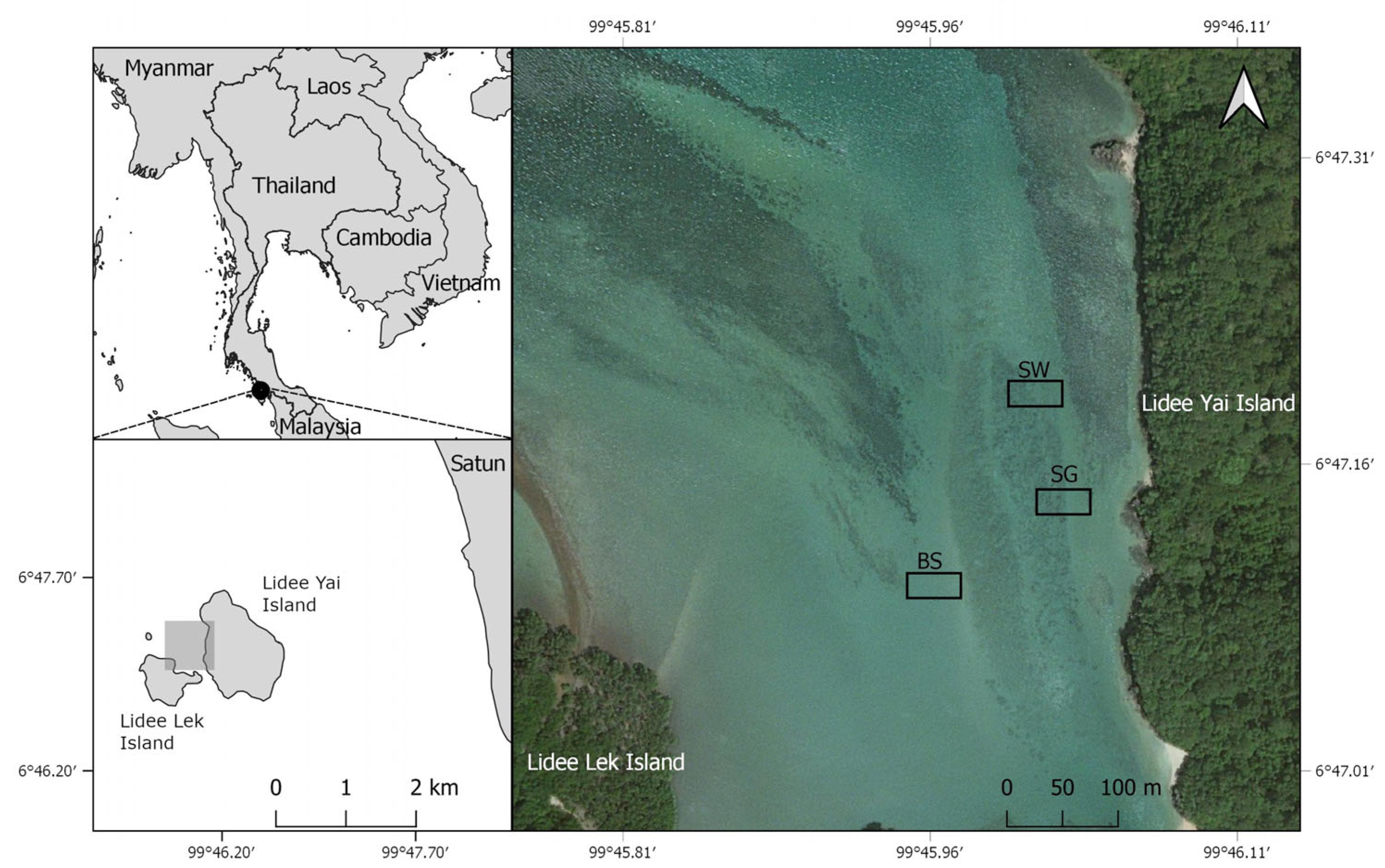
2.1. Study Site
2.2. Field Sampling Periods
2.3. Density, Size, and Recruitment from Asexual Reproduction of H. atra
2.4. Recruitment from Sexual Reproduction of H. atra
2.5. Abundance and Aboveground of Macrophytes
2.6. Properties of Sediment
2.7. Environmental Factors
2.8. Statistical Analyses
3. Results
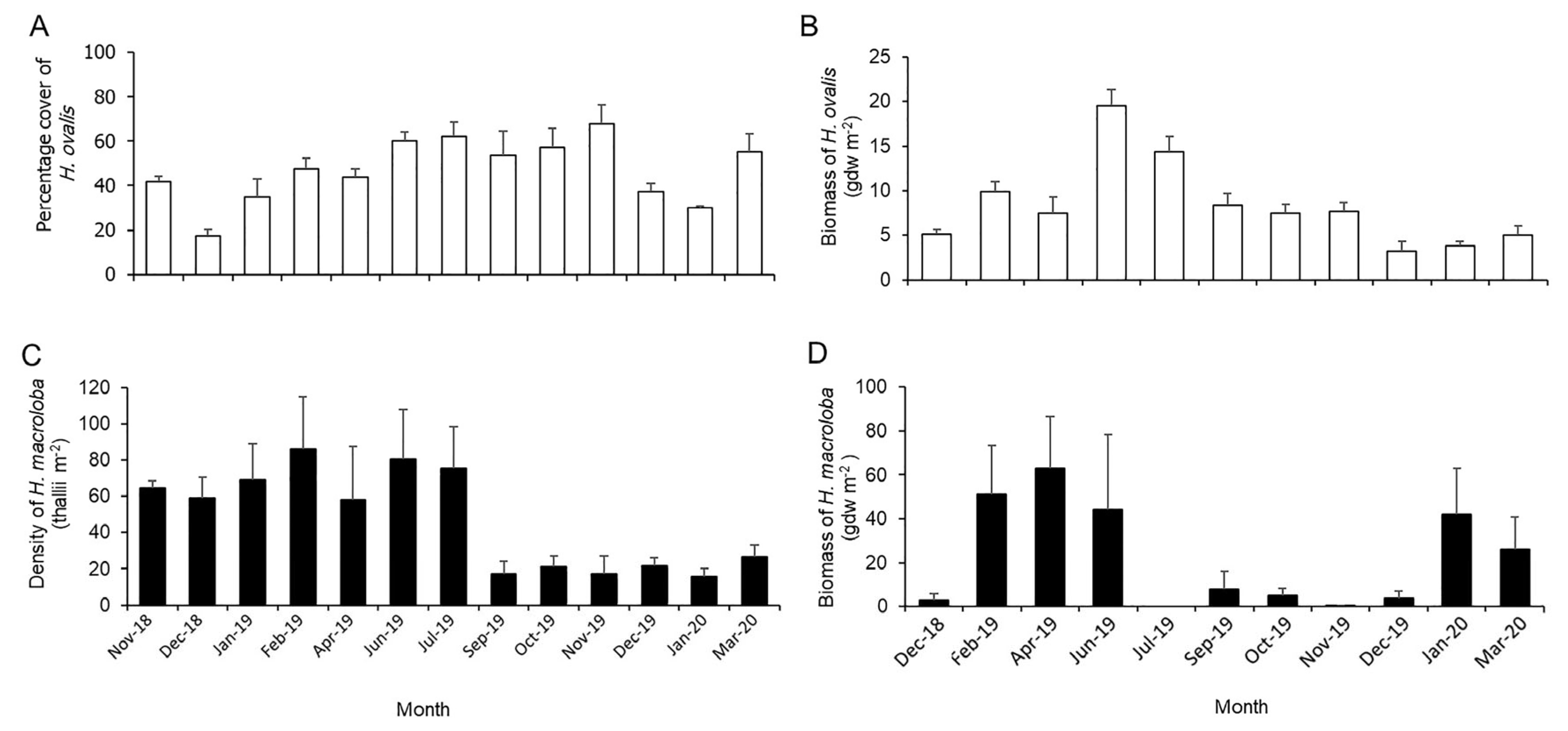
3.1. Abundance and Aboveground Biomass of Macrophyte Vegetation
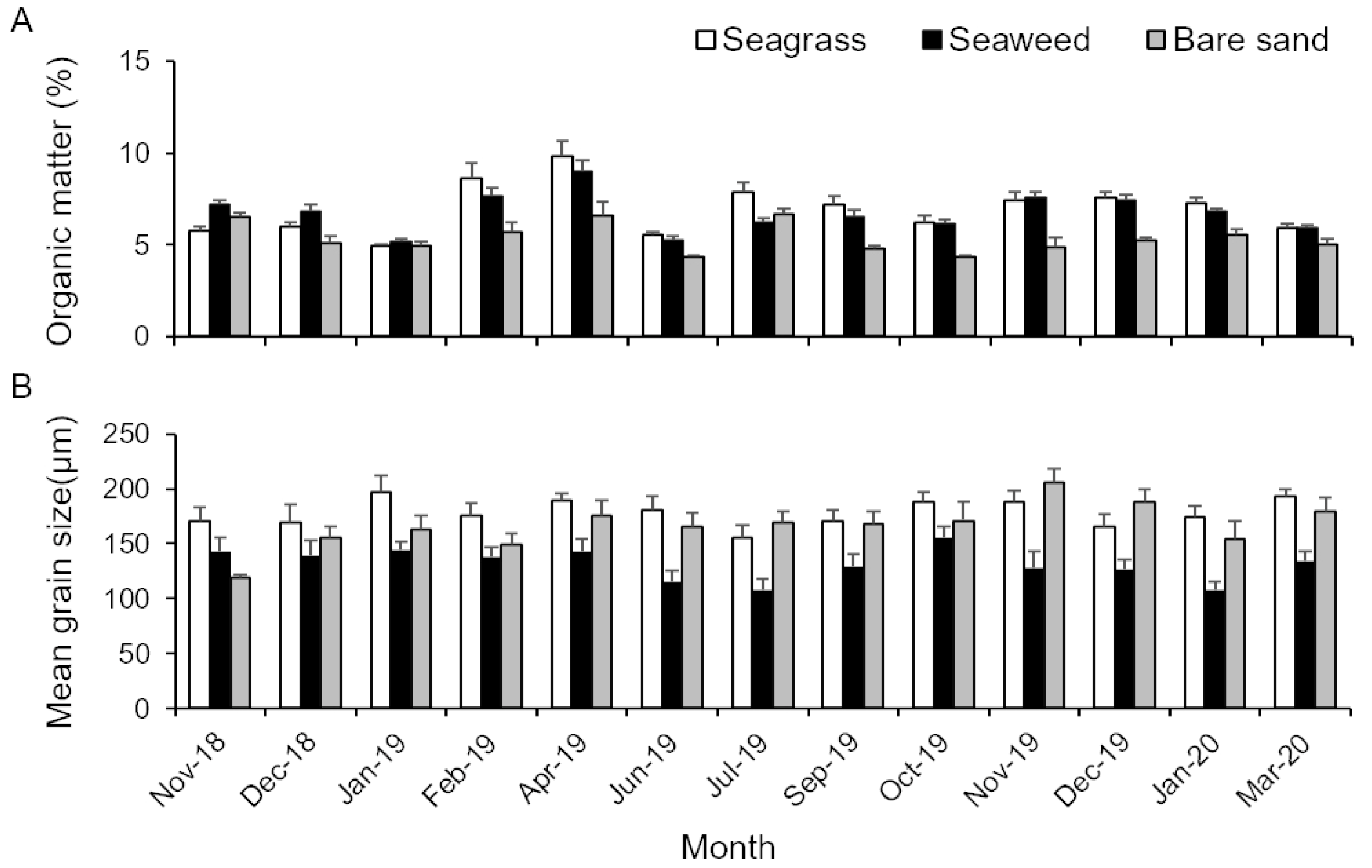
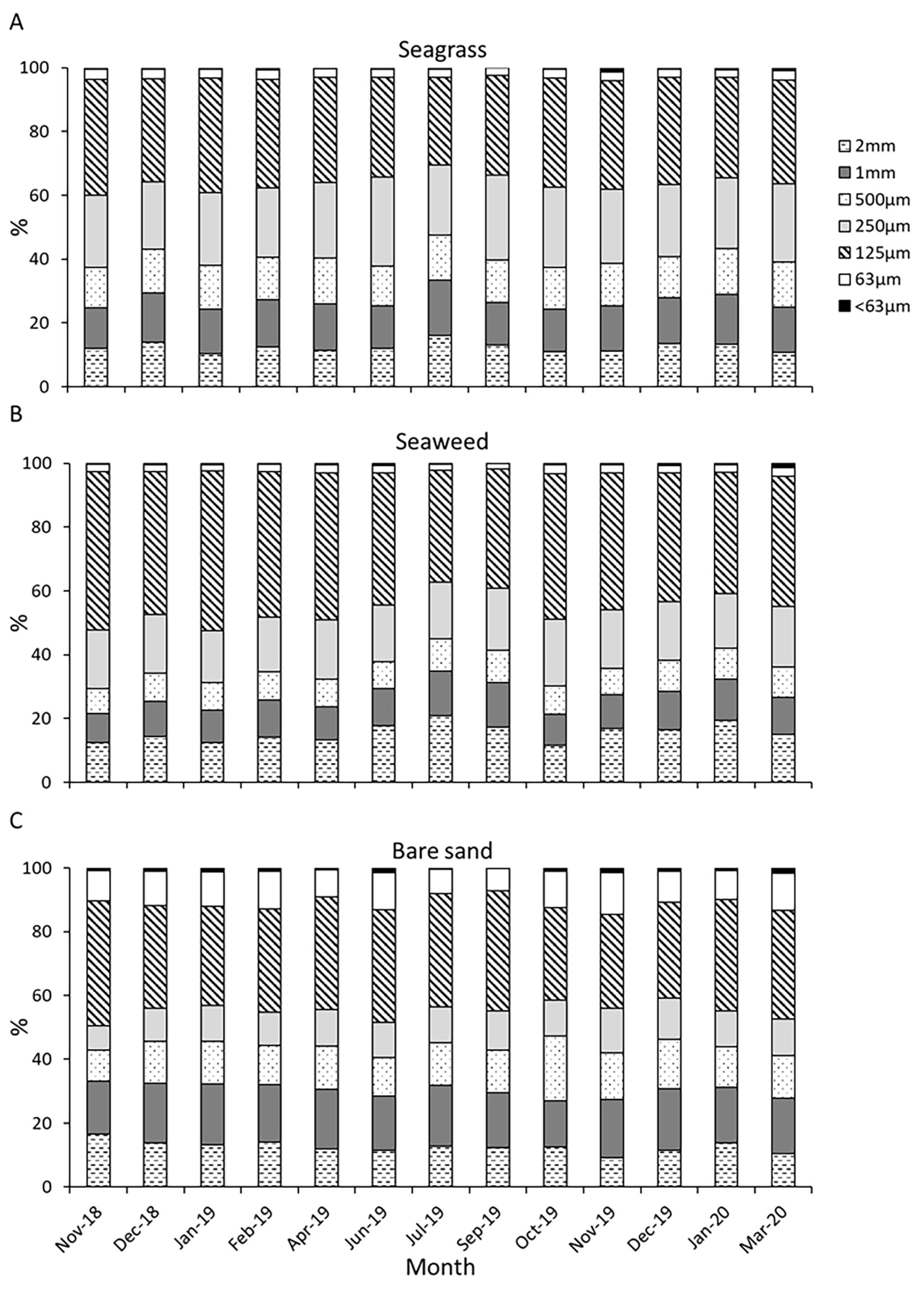
3.2. Properties of Sediment
3.3. Environmental Factors
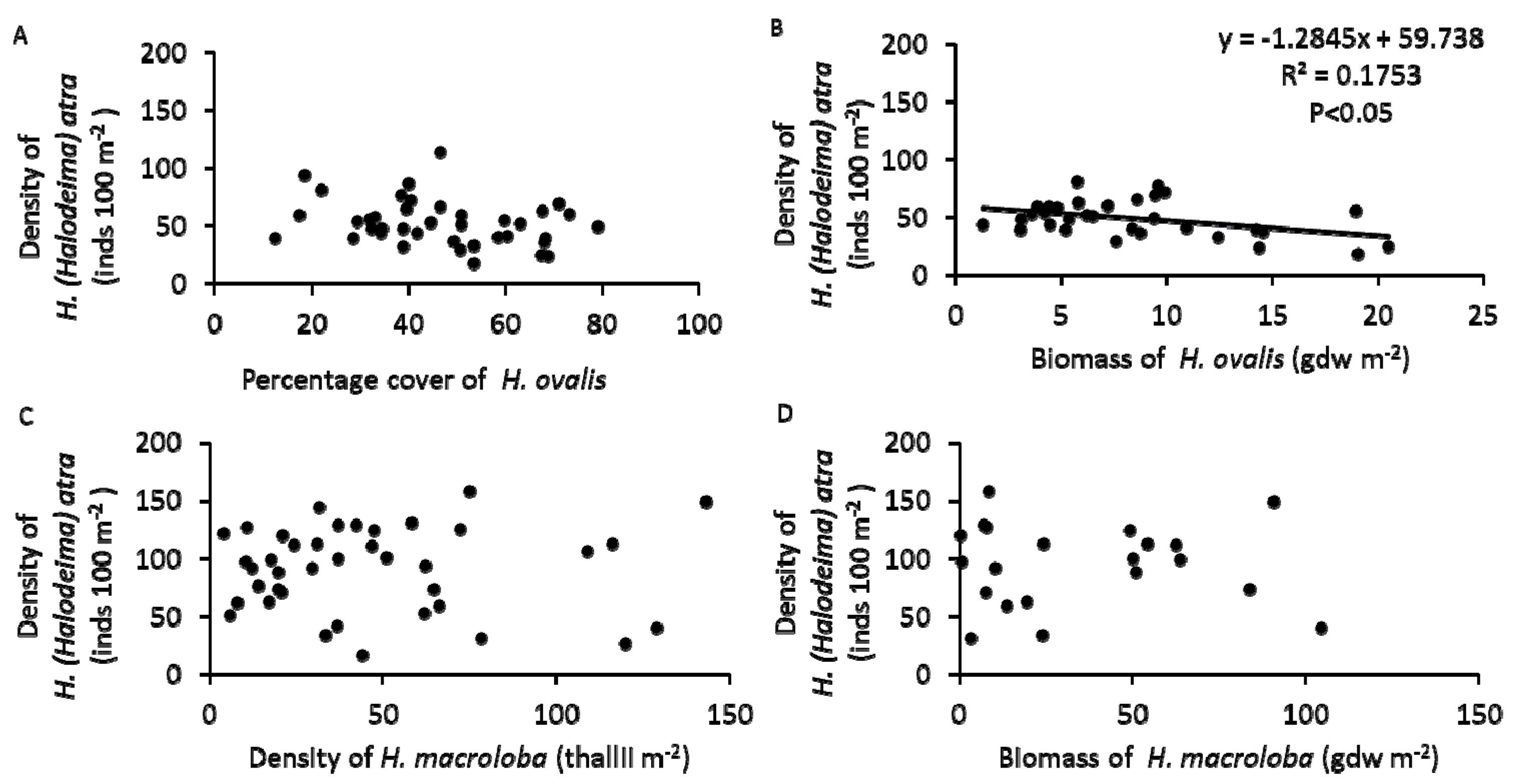
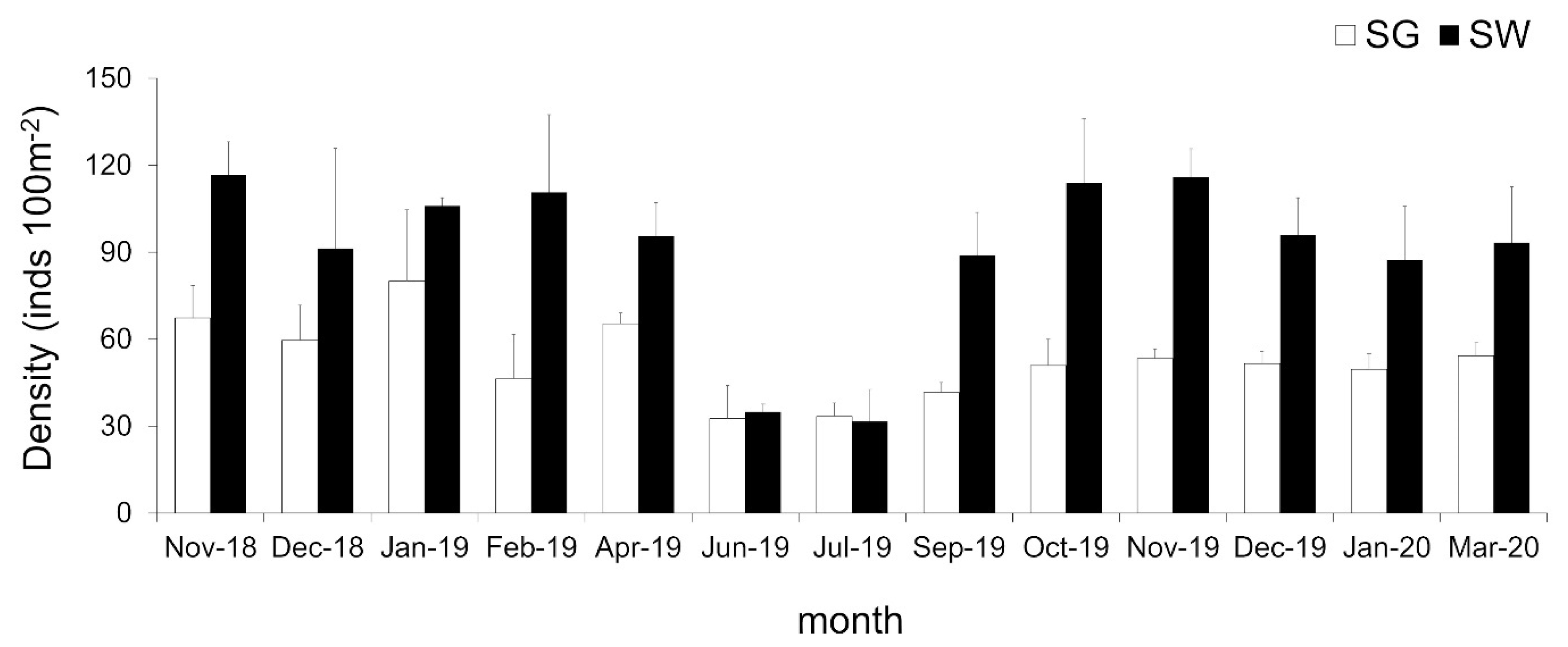
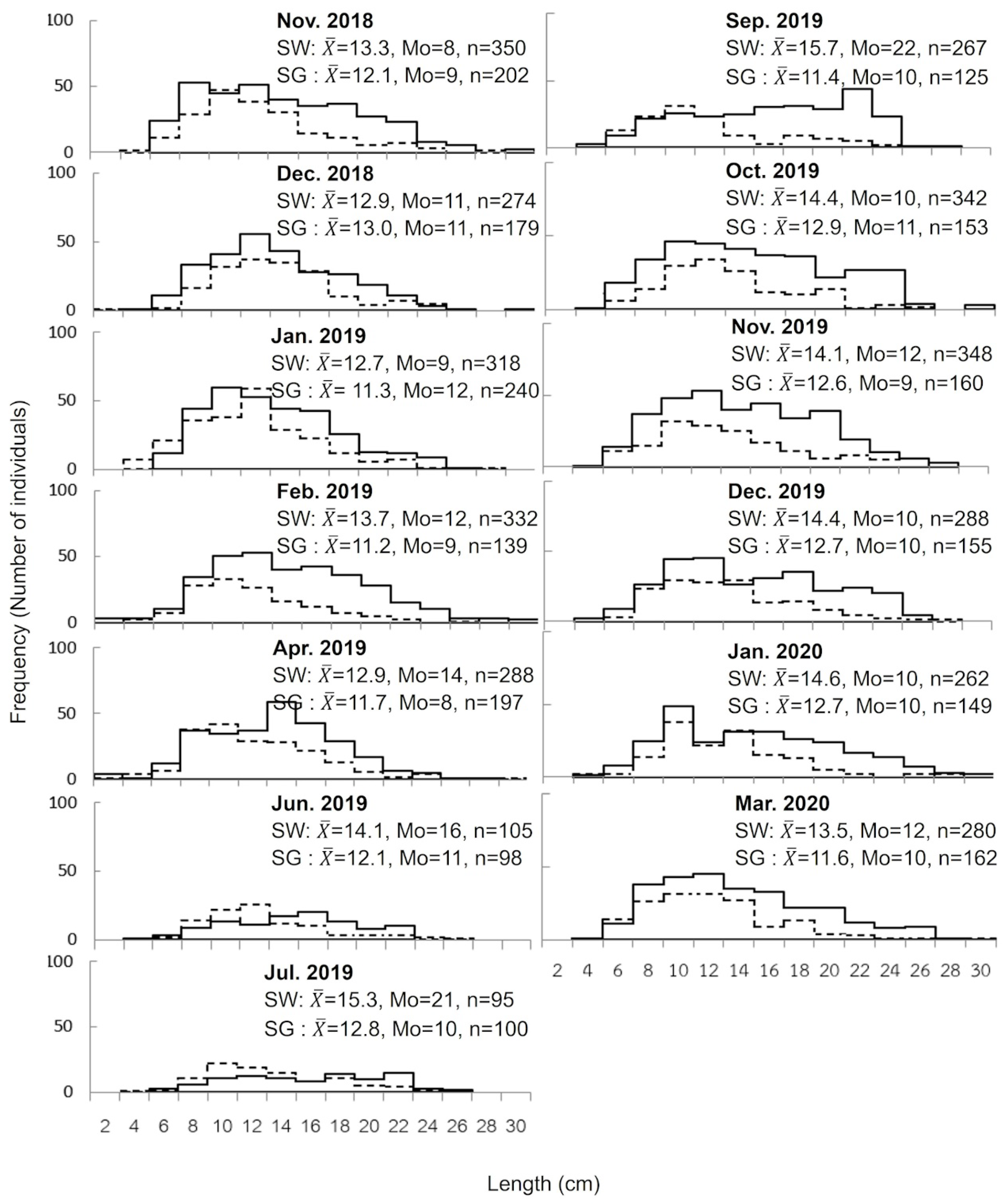
3.4. Population of H. atra
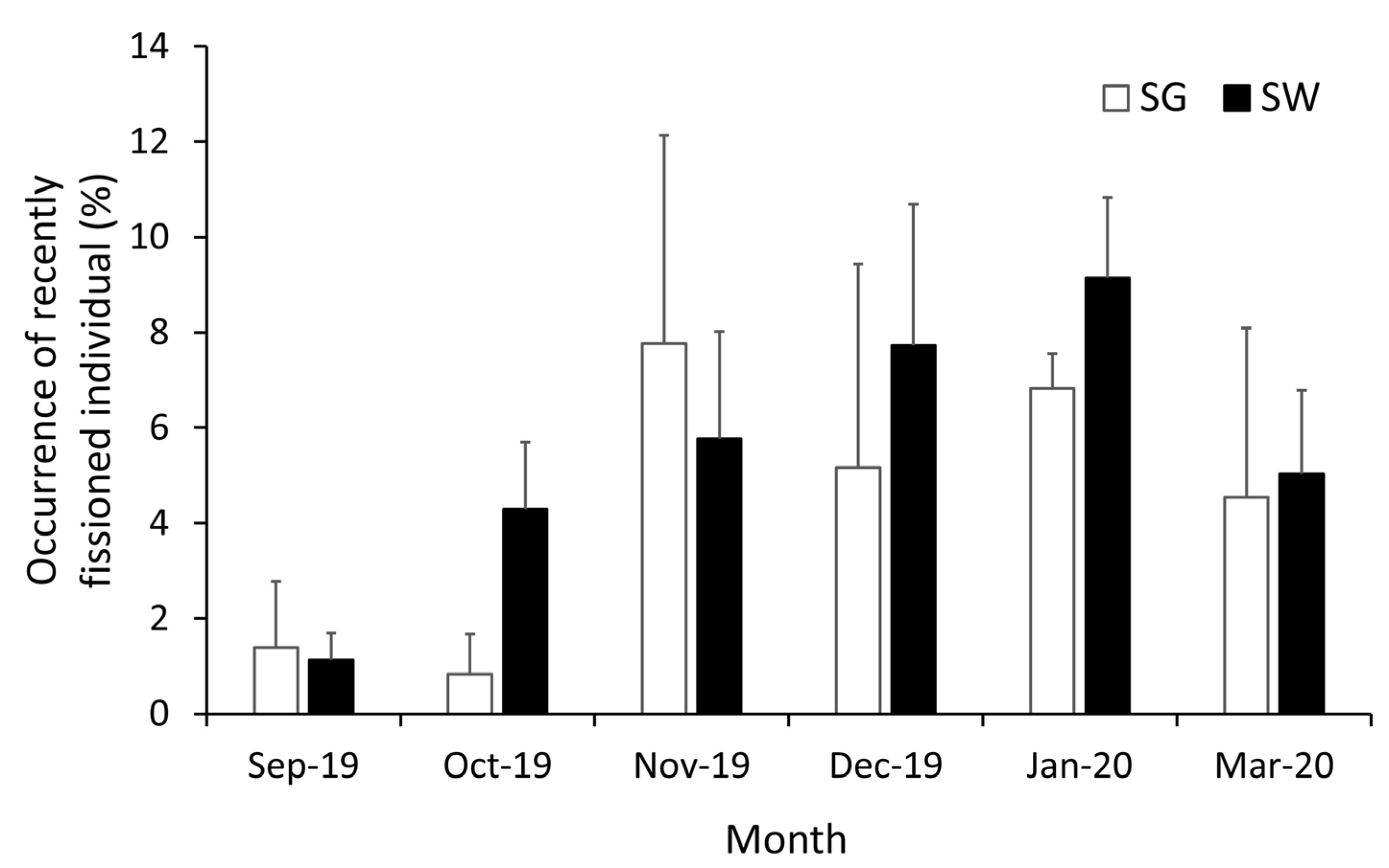
3.5. Recruitment of H. atra
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php?p=browser&id[]=148744&id[]=1393249#focus (accessed on 31 October 2022).
- Toral-Granda, V.; Lovatelli, A.; Vasconcellos, M. Sea Cucumbers: A Global Review of Fisheries and Trade; Food and Agriculture Organization: Rome, Italy, 2008; pp. 1–304. [Google Scholar]
- Mercier, A.; Battaglene, S.C.; Hamel, J.F. Daily burrowing cycle and feeding activity of juvenile sea cucumbers Holothuria scabra in response to environmental factors. J. Exp. Mar. Biol. Ecol. 1999, 239, 125–156. [Google Scholar] [CrossRef]
- Schneider, K.; Silverman, J.; Woolsey, E.; Eriksson, H.; Byrne, M.; Caldeira, K. Potential influence of sea cucumbers on coral reef CaCO3 budget: A case study at One Tree Reef. J. Geophys. Res. 2011, 116, G04032. [Google Scholar]
- MacTavish, T.; Stenton-Dozey, J.; Vopel, K.; Savage, C. Deposit-Feeding Sea Cucumbers Enhance Mineralization and Nutrient Cycling in Organically Enriched Coastal Sediments. PLoS ONE 2012, 7, e50031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domínguez-Godino, J.A.; González-Wangüemert, M. Habitat associations and seasonal abundance patterns of the sea cucumber Holothuria arguinensis at Ria Formosa coastal lagoon (South Portugal). Aquat. Ecol. 2020, 54, 337–354. [Google Scholar] [CrossRef]
- Purcell, S.W. Value, Market Preferences and Trade of Bêche-de-mer from Pacific Island Sea Cucumbers. PLoS ONE 2014, 9, e95075. [Google Scholar] [CrossRef]
- Xia, S.D.; Wang, X.Y. Nutritional and medicinal value. In The Sea Cucumber Apostichopus japonicus: History, Biology and Aquaculture; Yang, H.S., Mercler, A., Hamel, J.F., Eds.; Academic Press: New York, NY, USA, 2015; Volume 39, pp. 353–366. [Google Scholar]
- Purcell, S.W.; Williamson, D.H.; Ngaluafe, P. Chinese market prices of Bêche-de-mer: Implications for fisheries and aquaculture. Mar. Policy 2018, 91, 58–65. [Google Scholar] [CrossRef]
- Lovatelli, A.; Conand, C.; Purcell, S.; Uthicke, S.; Hamel, J.F.; Mercier, A. Advances in Sea Cucumber Aquaculture and Management; Food and Agriculture Organization: Rome, Italy, 2004; pp. 1–418. [Google Scholar]
- Hasan, M.H. Destruction of sea cucumber populations due to overfishing at Abu Ghosoun area, Red Sea. J. Basic. Appl. Zool. 2019, 80, 5. [Google Scholar] [CrossRef] [Green Version]
- Anderson, S.C.; Flemming, J.M.; Watson, R.; Lotze, H.K. Serial exploitation of global sea cucumber fisheries. Fish. 2010, 12, 317–339. [Google Scholar] [CrossRef]
- Purcell, S.W.; Samyn, Y.; Conand, C. A Commercially Important Sea Cucumber of the World; Food and Agriculture Organization: Rome, Italy, 2012; pp. 80–81. [Google Scholar]
- Choo, P.S. Population status, fisheries, and trade of sea cucumbers in Asia. In Sea Cucumbers: A Global Review of Fisheries and Trade; Toral-Granda, V., Lovatelli, A., Vasconcellos, M., Eds.; Food and Agriculture Organization: Rome, Italy, 2008; Volume 516, pp. 81–118. [Google Scholar]
- Li, X. Fishery and resource management of tropical sea cucumbers in the islands of the South China Sea. In Advances in Sea Cucumber Aquaculture and Management; Lovatelli, A., Connand, C., Purcell, S., Uthicke, S., Hamel, J.H., Mercier, A., Eds.; Food and Agriculture Organization: Rome, Italy, 2004; Volume 463, pp. 261–265. [Google Scholar]
- Bonham, K.; Held, E. Ecological Observations on the Sea Cucumbers Holothuria atra and H. eucospilota at Rongelap Atoll, Marshall Islands. Pac. Sci. 1963, 17, 305–314. [Google Scholar]
- Chao, S.M.; Chen, C.P.; Alexander, P.S. Fission and its effect on population structure of Holothuria atra (Echinodermata: Holothuroidea) in Taiwan. Mar. Biol. 1993, 116, 109–115. [Google Scholar] [CrossRef]
- Conand, C. Asexual reproduction by fission in Holothuria atra: Variability Asexual Reproduction Regeneration Holothurian Strategy Indo-Pacific of some parameters in populations from the tropical Indo Pacific. Oceanol. Acta 1996, 19, 209–216. [Google Scholar]
- Conand, C. Holothurians (sea cucumbers, Class Holothuroidea). In The Living Marine Resources of the Western Central Pacific; Carpenter, K.E., Niem, V.H., Eds.; Food and Agriculture Organization: Rome, Italy, 1998; Volume 2, pp. 1158–1164. [Google Scholar]
- Thorne, B.V.; Eriksson, H.; Byrne, M. Long term trends in population dynamics and reproduction in Holothuria atra (Aspidochirotida) in the southern Great Barrier Reef; the importance of asexual and sexual reproduction. J. Mar. Biol. Asso. UK 2013, 93, 1067–1072. [Google Scholar] [CrossRef]
- Bussarawit, S.; Thongtham, N. Sea Cucumber Fisheries and Trade in Thailand. In Proceedings of the Conservation of Sea Cucumber in Malaysia, Their Taxonomy, Ecology and Trade, Kuala Lumpur, Malaysia, 25 February 1999; pp. 26–36. [Google Scholar]
- Asha, P.S.; Diwakar, K.; Santhanavalli, G.; Manissery, M.K. Comparative distribution and habitat preference of the sea cucumber Holothuria atra Jaeger at protected and unprotected sites in Thoothukudi region of Gulf of Mannar, south-east coast of India. Indian J. Fish. 2015, 62, 52–57. [Google Scholar]
- Short, F.T.; Wyllie-Echeverria, S. Natural and human-induced disturbance of seagrasses. Environ. Conserv. 1996, 23, 17–27. [Google Scholar] [CrossRef]
- Edmunds, P.J.; Carpenter, R.C. Recovery of Diadema antillarum reduces macroalgal cover and increases abundance of juvenile corals on a Caribbean reef. Proc. Natl. Acad. Sci. USA 2001, 98, 5067–5071. [Google Scholar] [CrossRef] [Green Version]
- Cheevaporn, V.; Menasveta, P. Water pollution and habitat degradation in the Gulf of Thailand. Mar. Pollut. Bull. 2003, 47, 43–51. [Google Scholar] [CrossRef]
- Choo, P.S. The Philippines: A hotspot of sea cucumber fisheries in Asia. In Sea Cucumbers: A Global Review of Fisheries and Trade; Toral-Granda, V., Lovatelli, A., Vasconcellos, M., Eds.; Food and Agriculture Organization: Rome, Italy, 2008; Volume 516, pp. 119–140. [Google Scholar]
- Mercier, A.; Battaglene, S.C.; Hamel, J.F. Periodic movement, recruitment and size-related distribution of the sea cucumber Holothuria scabra in Solomon Islands. Hydrobiologia 2000, 440, 81–100. [Google Scholar] [CrossRef]
- Coppejans, E.; Prathep, A.; Lewmanomont, K.; Hayashizaki, K.; De Clerck, O.; Leliaert, F.; Terada, R. Seaweeds and Seagrasses of the Southern Andaman Sea Coast of Thailand; Kagoshima University Museum: Kagoshima, Japan, 2017; pp. 13–245. [Google Scholar]
- Sloan, N.A.; von Bodungen, B. Distribution and Feeding of the Sea Cucumber Isostichopus badionotus in Relation to Shelter and Sediment Criteria of the Bermuda Platform. Mar. Ecol. Prog. Ser. 1980, 2, 257–264. [Google Scholar] [CrossRef]
- Storer, D.A. A simple high sample volume ashing procedure for determining soil organic matter. Commun. Soil Sci. Plant Anal. 1984, 15, 759–772. [Google Scholar] [CrossRef]
- Wentworth, C.K. A Scale of Grade and Class Terms for Clastic Sediments. J. Geol. 1922, 30, 377–392. [Google Scholar] [CrossRef]
- Blott, S.J.; Pye, K. GRADISTAT: A grain size distribution and statistics package for the analysis unconsolidated sediments. Earth Surf. Process. Landf. 2001, 26, 1237–1248. [Google Scholar] [CrossRef]
- Stankovic, M.; Kaewsrikhaw, R.; Rattanachot, E.; Prathep, A. Modeling of suitable habitat for small-scale seagrass restoration in tropical ecosystems. Estuar. Coast. Shelf Sci. 2019, 231, 106465. [Google Scholar] [CrossRef]
- Komatsu, T.; Umezawa, Y.; Nakaoka, M.; Supanwanid, C.; Kanamoto, Z. Water flow and sediment in Enhalus acoroides and other seagrass beds in the Andaman Sea, off Khao Bae Na, Thailand. Coast. Mar. Sci. 2004, 29, 63–68. [Google Scholar]
- R Core Team. The R Project for Statistica. Available online: https://www.R-project.org/ (accessed on 10 November 2021).
- Panyawai, J.; Tuntiprapas, P.; Prathep, A. High macrophyte canopy complexity enhances sediment retention and carbon storage in coastal vegetative meadows at Tangkhen Bay, Phuket, Southern Thailand. Ecol. Res. 2019, 34, 201–212. [Google Scholar] [CrossRef]
- Adams, A.J.; Locascio, J.V.; Robbins, B.D. Microhabitat use by a post-settlement stage estuarine fish: Evidence from relative abundance and predation among habitats. J. Exp. Mar. Biol. Ecol. 2004, 299, 17–33. [Google Scholar] [CrossRef]
- Seeruttun, R. A Study on the Factors Influencing the Growth and Survival of Juvenile Sea Cucumber, Holothuria atra, under Laboratory Conditions. Univ. Maurit. R. J. 2008, 14, 1–15. [Google Scholar]
- Seabra, R.; Wethey, D.S.; Santos, A.M.; Lima, F.P. Side matters: Microhabitat influence on intertidal heat stress over a large geographical scale. J. Exp. Mar. Biol. Ecol. 2011, 400, 200–208. [Google Scholar] [CrossRef]
- Dissanayake, D.C.T.; Stefansson, G. Habitat preference of sea cucumbers: Holothuria atra and Holothuria edulis in the coastal waters of Sri Lanka. J. Mar. Biol. Assoc. UK 2012, 92, 581–590. [Google Scholar] [CrossRef]
- Lampe, K. Holothurian density, distribution and diversity comparing sites with different degrees of exploitation in the shallow lagoons of Mauritius. SPC Bêche-de-mer Inf. Bull. 2013, 33, 23–29. [Google Scholar]
- Cabansag, J.B.P.; Margarita, T.; Gajelan-Samson, M.B.P.; Diaz, F.A. Abundance and density of Holothuria atra and Bohadschia marmorata in Marine Protected Areas (MPAs) of Guiuan Eastern Samar and Palompon Leyte, Philippines. Int. J. Agric. Technol. 2014, 10, 119–131. [Google Scholar]
- Setyastuti, A. Echinodermata, Holothuria atra, in an intertidal seagrass bed off The Bama beach, Baluran National Park, East Java, Indonesia. J. Ilmu Teknol. Kelaut. Trop. 2014, 6, 31–39. [Google Scholar]
- Darnell, M.Z.; Backwell, P.R.Y.; Stockbridge, J.; Dyson, M.L.; Munguia, P. Thermal and desiccation constraints drive territory preference in fiddler crabs. J. Exp. Mar. Biol. Ecol. 2019, 518, 151173. [Google Scholar] [CrossRef]
- Hendriks, I.E.; Bouma, T.J.; Morris, E.P.; Duarte, C.M. Effects of seagrasses and algae of the Caulerpa family on hydrodynamics and particle-trapping rates. Mar. Biol. 2010, 157, 473–481. [Google Scholar] [CrossRef]
- Honkoop, P.J.C.; Berghuis, E.M.; Holthuijsen, S.; Lavaleye, M.S.S.; Piersma, T. Molluscan assemblages of seagrass-covered and bare intertidal flats on the Banc d’Arguin, Mauritania, in relation to characteristics of sediment and organic matter. J. Sea Res. 2008, 60, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Tuntiprapas, P.; Hayashizaki, K.; Ogawa, H.; Panyawai, J.; Tamada, S.; Stankovic, M.; Prathep, A. The contributions of allochthonous and autochthonous materials to organic carbon in coastal sediment: A case study from Tangkhen Bay, Phuket, Thailand. Ecol. Res. 2019, 34, 718–729. [Google Scholar] [CrossRef]
- Kaewsrikhaw, R.; Ritchie, R.J.; Prathep, A. Variations of tidal exposures and seasons on growth, morphology, anatomy and physiology of the seagrass Halophila ovalis (R.Br.). Hook. f. in a seagrass bed in Trang Province, Southern Thailand. Aquat. Bot. 2016, 130, 11–20. [Google Scholar] [CrossRef]
- Vermaat, J.E.; Agawin, N.S.R.; Duarte, C.M.; Fortes, M.D.; Marbà, N.; Uri, J.S. Meadow maintenance, growth and productivity of a mixed Philippine seagrass bed. Mar. Eco. Rog. Ser. 1995, 124, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Mayakun, J.; Prathep, A. Calcium carbonate productivity by Halimeda macroloba in the tropical intertidal ecosystem: The significant contributor to global carbonate budgets. Phycol. Res. 2019, 67, 94–101. [Google Scholar] [CrossRef]
- Moriarty, D.J.W. Feeding of Holothuria atra and Stichopus chloronotus on Bacteria, organic carbon and organic nitrogen in sediments of the Great Barrier Reef. Aust. J. Mar. Fresh. Res. 1982, 33, 255–263. [Google Scholar] [CrossRef]
- Uthicke, S.; Karez, R. Sediment patch selectivity in tropical sea cucumbers (Holothurioidea: Aspidochirotida) analyzed with multiple choice experiments. J. Exp. Mar. Bio. Ecol. 1999, 236, 69–87. [Google Scholar] [CrossRef]
- Viyakarn, V.; Chavanich, S.; Heery, E.; Raksasab, C. Distribution of sea cucumbers, Holothuria atra, on reefs in the upper Gulf of Thailand and the effect of their population densities on sediment microalgal productivity. Estuar. Coast. Shelf Sci. 2020, 235, 106514. [Google Scholar] [CrossRef]
- Ralph, P.J.; Burchett, M.D. Photosynthetic responses of the seagrass Halophila ovalis (R. Br.) Hook. f. to high irradiance stress, using chlorophyll a fluorescence. Aquat. Bot. 1995, 51, 55–66. [Google Scholar] [CrossRef]
- Hartati, R.; Zainuri, M.; Ambariyanto, A.; Widianingsih, W. Feeding selectivity of Holothuria atra in different microhabitat in Panjang Island, Jepara (Java, Indonesia). Biodiversitas 2020, 21, 2233–2239. [Google Scholar] [CrossRef]
- Ebert, T.A. Growth and Size of the Tropical Sea Cucumber Holothuria (Halodeima) atra Jager at Enewetak Atoll, Marshall Islands. Pac. Sci. 1978, 32, 183–191. [Google Scholar]
- Jaquemet, S.; Rousset, V.; Conand, C. Asexual reproduction parameters and the influence of fission on a Holothuria atra sea cucumber population from a fringing reef on Reunion Island (Indian Ocean). SPC Bêche-de-mer Inf. Bull. 1999, 11, 12–18. [Google Scholar]
- Conand, C. Monitoring a fissiparous population of Holothuria atra on a fringing reef on Reunion Island (Indian Ocean). SPC Bêche-de-mer Inf. Bull. 2004, 20, 22–25. [Google Scholar]
- Uthicke, S. Influence of asexual reproduction on the structure and dynamics of Holothuria (Halodeima) atra and Stichopus chloronotus populations of the Great Barrier Reef. Mar. Fresh. Res. 2001, 52, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Lane, D. Sea cucumber diversity and resources in Brunei, Borneo Island. In Proceedings of the 11th International Echinoderm Conference, Munich, Germany, 6-10 October 2004; pp. 231–237. [Google Scholar]
- Lee, J.; Byrne, M.; Uthicke, S. The influence of population density on fission and growth of Holothuria atra in natural mesocosms. J. Exp. Mar. Biol. Ecol. 2008, 365, 126–135. [Google Scholar] [CrossRef]
- Ebert, T.A. Dynamics of Holothuria atra at Enewetak Atoll, Republic of the Marshall Islands, based on tetracycline tagging. In Echinoderms: Durham: Proceedings of the 12th International Echinoderm Conference, Durham, UK, 7-11 August 2006; Taylor & Francis: Washington DC, USA, 2009; pp. 609–614. [Google Scholar]
- Ongkers, O.T.S.; Pattinasarany, M.; Mamesah, J.A.B.; Uneputty, P.R.A.; Pattikawa, J.A. Size distribution and growth pattern of Holothuria atra and Holothuria scabra in the coastal waters of Morella, Central Maluku Indonesia. Int. J. Fish. Aquat. Stud. 2018, 6, 301–305. [Google Scholar]
- Harriott, V.J. Reproductive biology of three congeneric sea cucumber species, Holothuria atra, H. Impatiens and H. edulis, at Heron Reef, Great Barrier Reef. Mar. Freshw. Res. 1985, 36, 51–57. [Google Scholar] [CrossRef]
- Uthicke, S. Seasonality of asexual reproduction in Holothuria (Halodeima) atra, H. (H.) edulis and Stichopus chloronotus (Holothuroidea: Aspidochirotida) on the Great Barrier Reef. Mar. Biol. 1997, 129, 435–441. [Google Scholar] [CrossRef]
- Chao, S.M.; Chen, C.P.; Alexander, P.S. Reproduction and growth of Holothuria atra (Echinodermata: Holothuroidea) at two contrasting sites in southern Taiwan. Mar. Biol. 1994, 119, 565–570. [Google Scholar] [CrossRef]
- Indriana, L.F.; Firdaus, M.; Soffa, F.B. Fissiparous sea cucumber (Holothuria atra) population in north lombok, Indonesia. Mar. Res. Indones. 2018, 43, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Dissanayake, D.C.T.; Stefansson, G. Reproductive biology of the commercial sea cucumber Holothuria atra (Holothuroidea: Aspidochirotida) in the northwestern coastal waters of Sri Lanka. Invertebr. Reprod. Dev. 2010, 54, 65–76. [Google Scholar] [CrossRef]
- Hamel, J.; Mercier, A. Early development, settlement, growth, and spatial distribution of the sea cucumber Cucumaria frondose (Echinodermata: Holothuroidea). J. Fish. Aquat. Sci. 1996, 53, 253–271. [Google Scholar] [CrossRef]
- Balch, T.; Scheibling, R.E. Larval supply, settlement, and recruitment in echinoderms. In Echinoderm Studies; Jangoux, M., Lawrence, J.M., Eds.; A. A Balkema: Rotterdam; The Netherland; Brookfield, VT, USA, 2001; Volume 6, pp. 1–83. [Google Scholar]
- Asha, P.S.; Muthiah, P. Effects of temperature, salinity and pH on larval growth, survival and development of the sea cucumber Holothuria spinifera Theel. Aquaculture 2005, 250, 823–829. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Total Length of H. atra | Organic Matter | ||||
|---|---|---|---|---|---|---|
| df | SS | H | Df | SS | H | |
| Habitat type | 1 | 4,006,900 | 153.930 *** | 2 | 851,280 | 84.595 *** |
| Month | 12 | 1,921,400 | 73.815 *** | 12 | 1,046,093 | 103.954 *** |
| Habitat type × month | 12 | 1,304,300 | 50.106 *** | 24 | 427,825 | 42.515 * |
| Residuals | 5573 | 13,849,000 | 308 | |||
| Source of Variation | Density | Occurrence of Recently Fissioned Individual | Mean Grain Size | ||||||
|---|---|---|---|---|---|---|---|---|---|
| df | MS | F | df | MS | F | df | MS | F | |
| Habitat type | 1 | 28,425 | 45.187 *** | 1 | 10.81 | 0.575 ns | 2 | 68,820 | 61.713 *** |
| Month | 12 | 2159 | 3.433 *** | 5 | 40.89 | 2.176 ns | 12 | 3039 | 2.725.** |
| Habitat type × month | 12 | 659 | 1.048 ns | 5 | 6.27 | 0.334 ns | 24 | 1967 | 1.764 * |
| Residuals | 52 | 629 | 24 | 18.79 | 308 | 1115 | |||
| Variable | Coefficient | SE | t-Value | VIF |
|---|---|---|---|---|
| Intercept | 351.171 | 194.561 | 1.805 ns | |
| Habitat type | 67.647 | 11.631 | 5.816 *** | 2.84 |
| Organic matter | 4.834 | 3.458 | 1.398 ns | 1.54 |
| Mean grain size | 0.571 | 0.207 | 2.754 ** | 2.71 |
| Light | 0.024 | 0.025 | 0.962 ns | 1.25 |
| Temperature | −14.749 | 5.837 | −2.527 * | 1.41 |
| Dissolution rate of gypsum ball | −7.348 | 43.969 | −0.167 ns | 1.65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kongsap, V.; Rattanachot, E.; Prathep, A.; Buaphol, W.; Mayakun, J. A High Abundance of Holothuria (Halodeima) atra (Holothuroidea Aspidochirotida) in a Halimeda Dominated Habitat. J. Mar. Sci. Eng. 2023, 11, 451. https://doi.org/10.3390/jmse11020451
Kongsap V, Rattanachot E, Prathep A, Buaphol W, Mayakun J. A High Abundance of Holothuria (Halodeima) atra (Holothuroidea Aspidochirotida) in a Halimeda Dominated Habitat. Journal of Marine Science and Engineering. 2023; 11(2):451. https://doi.org/10.3390/jmse11020451
Chicago/Turabian StyleKongsap, Vanyarat, Ekkalak Rattanachot, Anchana Prathep, Witthaya Buaphol, and Jaruwan Mayakun. 2023. "A High Abundance of Holothuria (Halodeima) atra (Holothuroidea Aspidochirotida) in a Halimeda Dominated Habitat" Journal of Marine Science and Engineering 11, no. 2: 451. https://doi.org/10.3390/jmse11020451