

Diel Variation in Phytoplankton Biomass Driven by Hydrological Factors at Three Coastal Monitoring Buoy Stations in the Taiwan Strait
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Instruments and Parameters
2.3. Sampling and Measurement
2.4. Statistical Analysis
3. Results
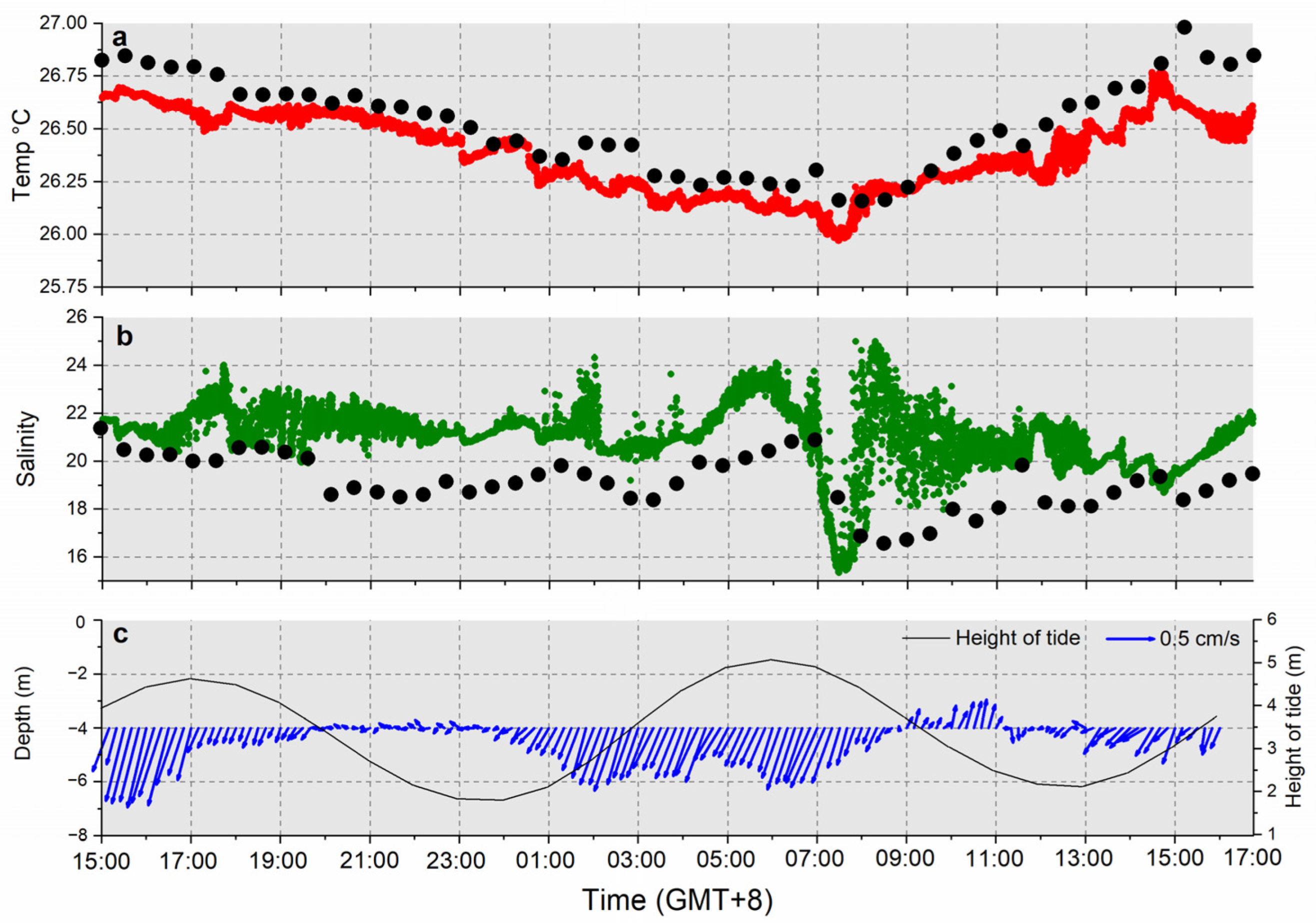
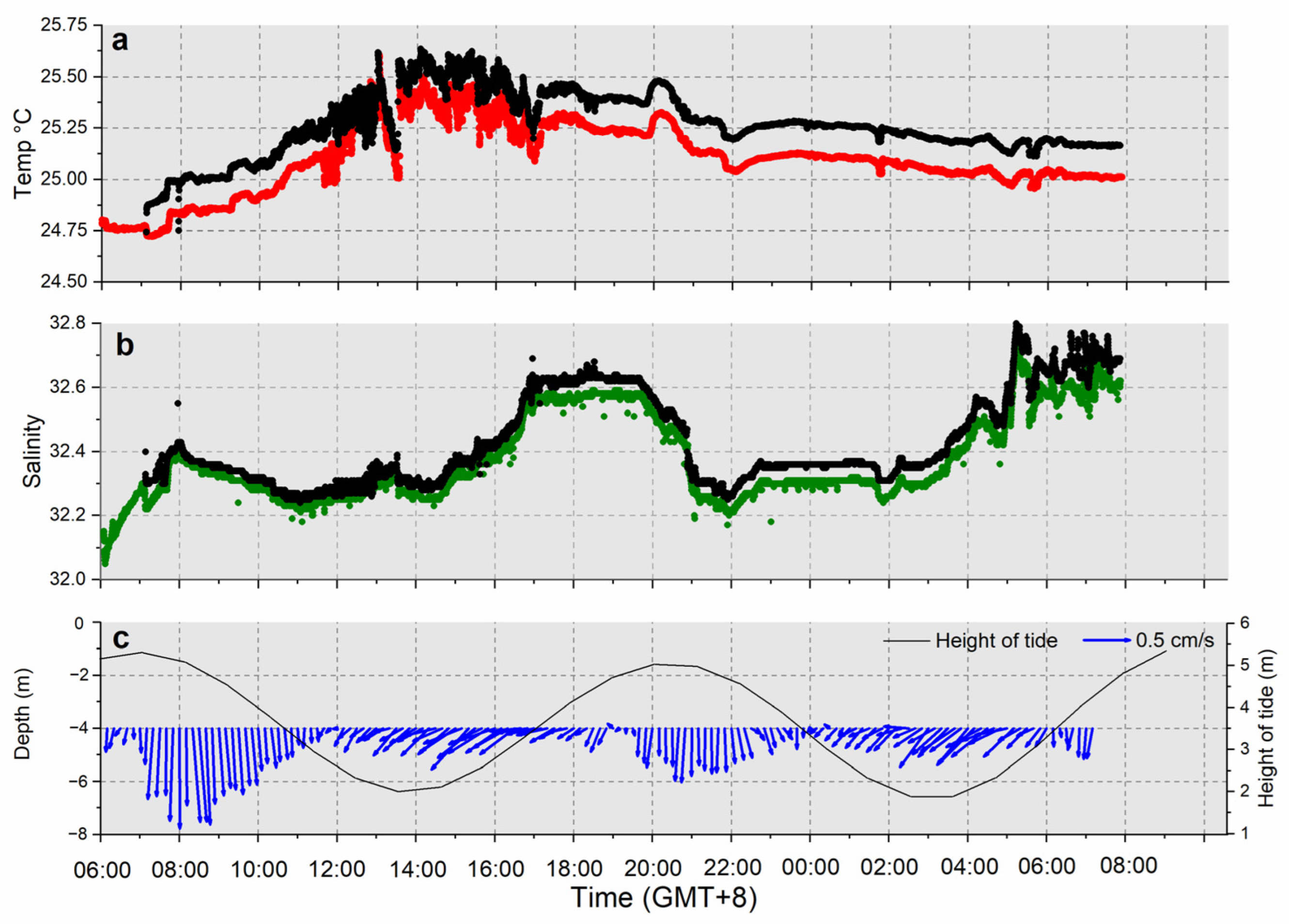
3.1. Hydrological Parameters
3.1.1. Douwei
3.1.2. Minjiang Estuary
3.1.3. Huangqi
3.2. Chemical Parameters
3.2.1. Douwei
3.2.2. Minjiang Estuary
3.2.3. Huangqi
3.3. Chlorophyll and Phycoerythrin Concentration
3.3.1. Douwei
3.3.2. Minjiang Estuary
3.3.3. Huangqi
3.3.4. Chlorophyll a and Phycoerythrin Concentration in Water Samples
4. Discussion
4.1. Phytoplankton Diel Variation in Different Water Masses
4.2. Tidal Influence
5. Conclusions
- There were significant differences in the characteristics of the water masses at the three buoy stations. The Minjiang Estuary Station was mainly influenced by the river plume, while the Huangqi Station was mainly influenced by coastal upwelling. The Douwei Station exhibited typical Taiwan Strait warm-water characteristics. The corresponding phytoplankton chlorophyll a concentration was highest at the Minjiang Estuary, followed by Huangqi, and was the lowest at Douwei.
- The diurnal variation in phytoplankton was jointly regulated by water masses, tides, and light. At the three stations, three different response patterns were observed. The station at Minjiang Estuary, with the highest biomass, was notably influenced by tidal action. On the other hand, the Huangqi Station, being affected by upwelling, exhibited two peaks in chlorophyll a concentration, which were attributed to tides and light, respectively. In contrast, the lowest concentration of phytoplankton chlorophyll a was found at Douwei Station, which was mainly attributed to the stable water mass found here compared with ME and HQ, which was not affected by physical processes.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Falkowski, P.G.; Barber, R.T.; Smetacek, V. Biogeochemical Controls and Feedbacks on Ocean Primary Production. Science 1998, 281, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Manizza, M.; Quéré, C.L.; Watson, A.J.; Buitenhuis, E.T. Bio-optical feedbacks among phytoplankton, upper ocean physics and sea-ice in a global model. Geophys. Res. Lett. 2005, 32, L05603. [Google Scholar] [CrossRef]
- Smayda, T.J.; Reynolds, C.S. Community Assembly in Marine Phytoplankton: Application of Recent Models to Harmful Dinoflagellate Blooms. J. Plankton Res. 2001, 23, 447–461. [Google Scholar] [CrossRef]
- Jan, S.; Sheu, D.D.; Kuo, H.-M. Water mass and throughflow transport variability in the Taiwan Strait. J. Geophys. Res. Ocean. 2006, 111, C12012. [Google Scholar] [CrossRef]
- Jan, S.; Wang, J.; Chern, C.-S.; Chao, S.-Y. Seasonal variation of the circulation in the Taiwan Strait. J. Mar. Syst. 2022, 35, 249–268. [Google Scholar] [CrossRef]
- Hsu, P.-C.; Lu, C.-Y.; Hsu, T.-W.; Ho, C.-R. Diurnal to Seasonal Variations in Ocean Chlorophyll and Ocean Currents in the North of Taiwan Observed by Geostationary Ocean Color Imager and Coastal Radar. Remote Sens. 2020, 12, 2853. [Google Scholar] [CrossRef]
- Gattuso, J.P.; Frankignoulle, M.; Wollast, R. Carbon and Carbonate Metabolism in Coastal Aquatic Ecosystems. Annu. Rev. Econ. 1998, 29, 405–434. [Google Scholar] [CrossRef]
- Simpson, J.H.; Brown, J.; Matthews, J.; Allen, G. Tidal straining, density currents, and stirring in the control of estuarine stratification. Estuaries 1990, 13, 125–132. [Google Scholar] [CrossRef]
- Thyssen, M.; Mathieu, D.; Garcia, N.; Denis, M. Short-term variation of phytoplankton assemblages in Mediterranean coastal waters recorded with an automated submerged flow cytometer. J. Plankton Res. 2008, 30, 1027–1040. [Google Scholar] [CrossRef]
- Tseng, H.-C.; You, W.-L.; Huang, W.; Chung, C.-C.; Tsai, A.-Y.; Chen, T.-Y.; Lan, K.-W.; Gong, G.-C. Seasonal Variations of Marine Environment and Primary Production in the Taiwan Strait. Front. Mar. Sci. 2020, 7, 38. [Google Scholar] [CrossRef]
- Zhong, Y.; Liu, X.; Xiao, W.; Laws, E.A.; Chen, J.; Wang, L.; Liu, S.; Zhang, F.; Huang, B. Phytoplankton community patterns in the Taiwan Strait match the characteristics of their realized niches. Prog. Oceanogr. 2020, 186, 102366. [Google Scholar] [CrossRef]
- Zhong, Y.; Hu, J.; Laws, E.A.; Liu, X.; Chen, J.; Huang, B. Plankton community responses to pulsed upwelling events in the southern Taiwan Strait. ICES J. Mar. Sci. 2019, 76, 2374–2388. [Google Scholar] [CrossRef]
- Wang, Y.; Kang, J.-H.; Ye, Y.-Y.; Lin, G.-M.; Yang, Q.-L.; Lin, M. Phytoplankton community and environmental correlates in a coastal upwelling zone along western Taiwan Strait. J. Mar. Syst. 2016, 154, 252–263. [Google Scholar] [CrossRef]
- Shang, S.L.; Zhang, C.Y.; Hong, H.S.; Shang, S.P.; Chai, F. Short-term variability of chlorophyll associated with upwelling events in the Taiwan Strait during the southwest monsoon of 1998. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2004, 51, 1113–1127. [Google Scholar] [CrossRef]
- Zhang, Y.; Su, J.-Z.; Su, Y.-P.; Lin, H.; Xu, Y.-C.; Barathan, B.P.; Zheng, W.-N.; Schulz, K.G. Spatial Distribution of Phytoplankton Community Composition and Their Correlations with Environmental Drivers in Taiwan Strait of Southeast China. Diversity 2020, 12, 433. [Google Scholar] [CrossRef]
- Parsons, T.; Maita, Y.; Lalli, C.M. Amanual of chemical and biological methods for seawater analysis. In Biological Oceanographic Processes; Parsons, T., Ed.; Pergamon Press: New York, NY, USA, 1984; p. 173. [Google Scholar]
- Kasinak, J.-M.E.; Holt, B.M.; Chislock, M.F.; Wilson, A.E. Benchtop fluorometry of phycocyanin as a rapid approach for estimating cyanobacterial biovolume. J. Plankton Res. 2015, 37, 248–257. [Google Scholar] [CrossRef]
- Hong, H.; Chai, F.; Zhang, C.; Huang, B.; Jiang, Y.; Hu, J. An overview of physical and biogeochemical processes and ecosystem dynamics in the Taiwan Strait. Cont. Shelf Res. 2011, 31, S3–S12. [Google Scholar] [CrossRef]
- Tang, D.; Kester, D.R.; Ni, I.-H.; Kawamura, H.; Hong, H. Upwelling in the Taiwan Strait during the summer monsoon detected by satellite and shipboard measurements. Remote Sens. Environ. 2002, 83, 457–471. [Google Scholar] [CrossRef]
- Chen, Z.; Pan, J.; Jiang, Y.; Lin, H. Far-reaching transport of Pearl River plume water by upwelling jet in the northeastern South China Sea. J. Mar. Syst. 2017, 173, 60–69. [Google Scholar] [CrossRef]
- Brunet, C.; Casotti, R.; Vantrepotte, V. Phytoplankton diel and vertical variability in photobiological responses at a coastal station in the Mediterranean Sea. J. Plankton Res. 2008, 30, 645–654. [Google Scholar] [CrossRef]
- O’Boyle, S.; Silke, J. A review of phytoplankton ecology in estuarine and coastal waters around Ireland. J. Plankton Res. 2010, 32, 99–118. [Google Scholar] [CrossRef]
- Carberry, L.; Roesler, C.; Drapeau, S. Correcting in situ chlorophyll fluorescence time-series observations for nonphotochemical quenching and tidal variability reveals nonconservative phytoplankton variability in coastal waters. Limnol. Oceanogr. Methods 2019, 17, 462–473. [Google Scholar] [CrossRef] [PubMed]
- Salisbury, J.E.; Jönsson, B.F.; Mannino, A.; Kim, W.; Goes, J.I.; Choi, J.-Y.; Concha, J.A. Assessing Net Growth of Phytoplankton Biomass on Hourly to Annual Time Scales Using the Geostationary Ocean Color Instrument. Geophys. Res. Lett. 2021, 48, e2021GL095528. [Google Scholar] [CrossRef]
- Blauw, A.N.; Benincà, E.; Laane, R.W.P.M.; Greenwood, N.; Huisman, J. Dancing with the Tides: Fluctuations of Coastal Phytoplankton Orchestrated by Different Oscillatory Modes of the Tidal Cycle. PLoS ONE 2012, 7, e49319. [Google Scholar] [CrossRef] [PubMed]
- Cereja, R.; Brotas, V.; Cruz, J.P.C.; Rodrigues, M.; Brito, A.C. Tidal and Physicochemical Effects on Phytoplankton Community Variability at Tagus Estuary (Portugal). Front. Mar. Sci. 2021, 8, 675699. [Google Scholar] [CrossRef]
- Baliarsingh, S.K.; Lotliker, A.A.; Srichandan, S.; Roy, R.; Sahu, B.K.; Samanta, A.; Nair, T.M.B.; Acharyya, T.; Parida, C.; Singh, S.; et al. Evaluation of hydro-biological parameters in response to semi-diurnal tides in a tropical estuary. Ecohydrol. Hydrobiol. 2021, 21, 700–717. [Google Scholar] [CrossRef]
- Sin, Y.; Jeong, B.; Park, C. Semidiurnal Dynamics of Phytoplankton Size Structure and Taxonomic Composition in a Macrotidal Temperate Estuary. Estuaries Coasts 2015, 38, 546–557. [Google Scholar] [CrossRef]
- Shanks, A.L.; Morgan, S.G.; Macmahan, J.; Reniers, A.; Jarvis, M.; Brown, J.; Fujimura, A.; Griesemer, C. Onshore transport of plankton by internal tides and upwelling-relaxation events. Mar. Ecol. Prog. Ser. 2014, 502, 39–51. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stations | Longitude (°E) | Latitude 3 (°N) | Depth (m) | Time and Date | Sunrise | Sunset |
|---|---|---|---|---|---|---|
| Douwei | 119°02.70′ | 25°04.07′ | 9 | 8 p.m. 4 June –9 p.m. 5 June | 05:15 | 18:52 |
| Minjiang Estuary | 119°43.50′ | 25°58.52′ | 11 | 3 p.m. 6 June –4 p.m. 7 June | 05:10 | 18:52 |
| Huangqi | 119°55.76′ | 26°23.36′ | 26 | 6 a.m. 8 June –7 a.m. 9 June | 05:09 | 18:53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, C.; Wang, L.; Zhang, Y.; Lin, M.; Wan, Y.; Zhou, X.; Jing, C.; Guo, X. Diel Variation in Phytoplankton Biomass Driven by Hydrological Factors at Three Coastal Monitoring Buoy Stations in the Taiwan Strait. J. Mar. Sci. Eng. 2023, 11, 2252. https://doi.org/10.3390/jmse11122252
Jia C, Wang L, Zhang Y, Lin M, Wan Y, Zhou X, Jing C, Guo X. Diel Variation in Phytoplankton Biomass Driven by Hydrological Factors at Three Coastal Monitoring Buoy Stations in the Taiwan Strait. Journal of Marine Science and Engineering. 2023; 11(12):2252. https://doi.org/10.3390/jmse11122252
Chicago/Turabian StyleJia, Cun, Lei Wang, Youquan Zhang, Meihui Lin, Yan Wan, Xiwu Zhou, Chunsheng Jing, and Xiaogang Guo. 2023. "Diel Variation in Phytoplankton Biomass Driven by Hydrological Factors at Three Coastal Monitoring Buoy Stations in the Taiwan Strait" Journal of Marine Science and Engineering 11, no. 12: 2252. https://doi.org/10.3390/jmse11122252