
Transcriptome Analysis of the Marine Nematode Litoditis marina in a Chemically Defined Food Environment with Stearic Acid Supplementation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Worms
2.2. Sea-Salt-CeMM Preparation
2.3. Dietary Stearic Acid Supplementation
2.4. Synchronization of L. marina
2.5. RNA-Seq Analysis
2.6. Quantitative Real-Time PCR (qPCR) Analysis
3. Results
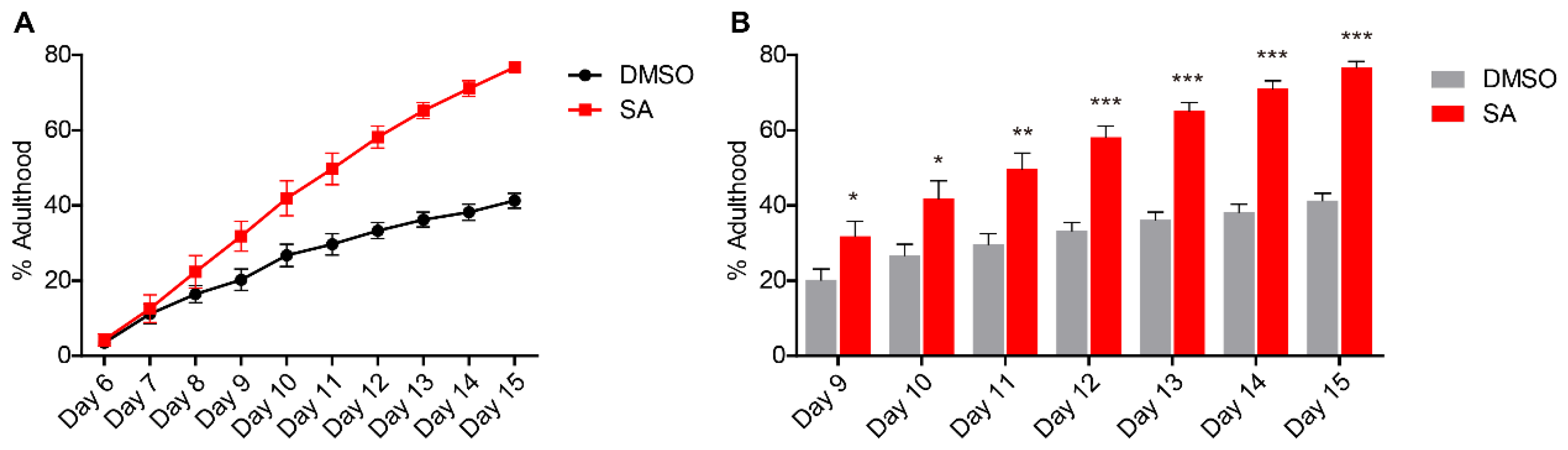
3.1. Atearic Acid Promotes Development of L. marina on CeMM
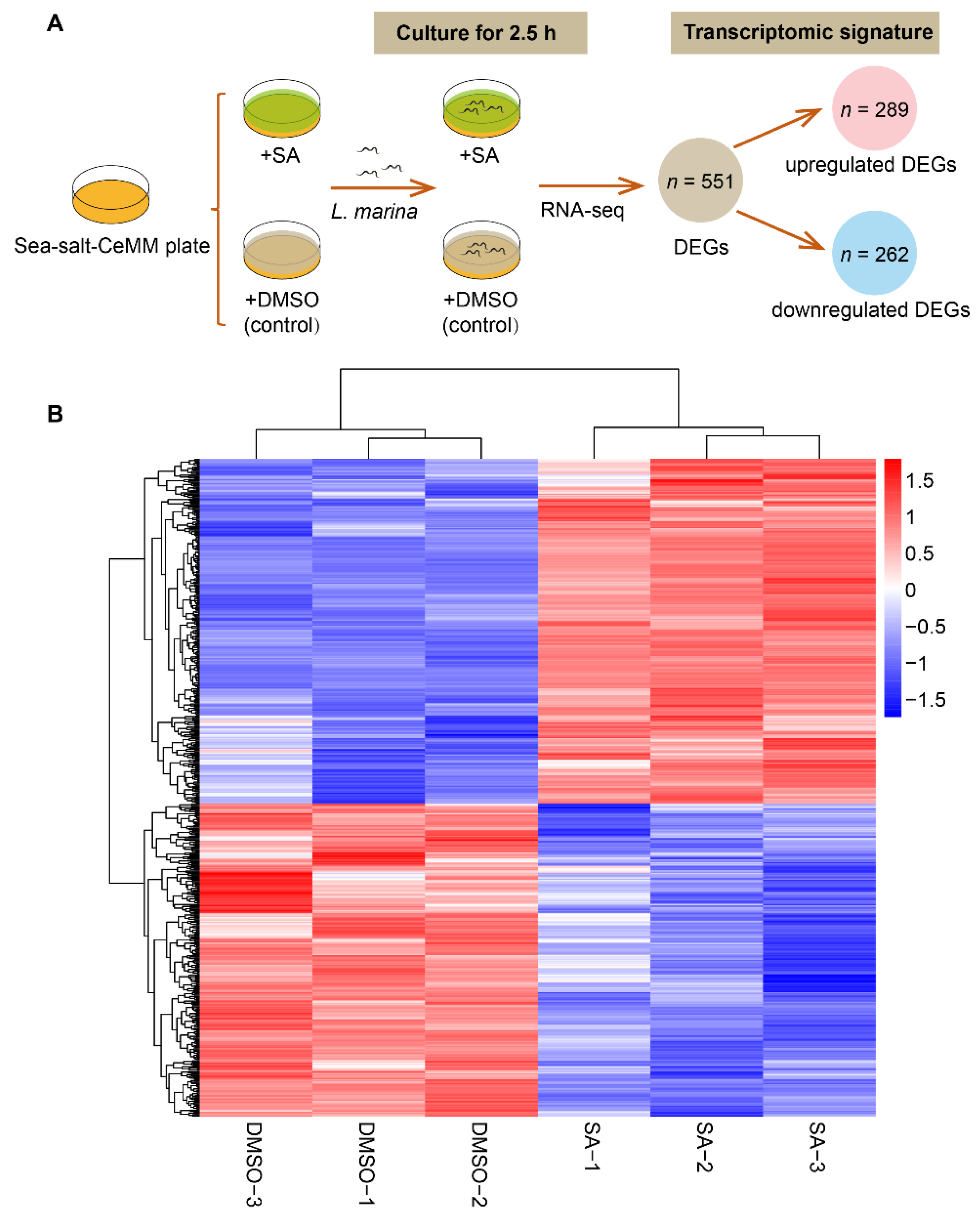
3.2. RNA-Seq Analysis in L. marina on Sea-Salt-CeMM with Stearic Acid Supplementation
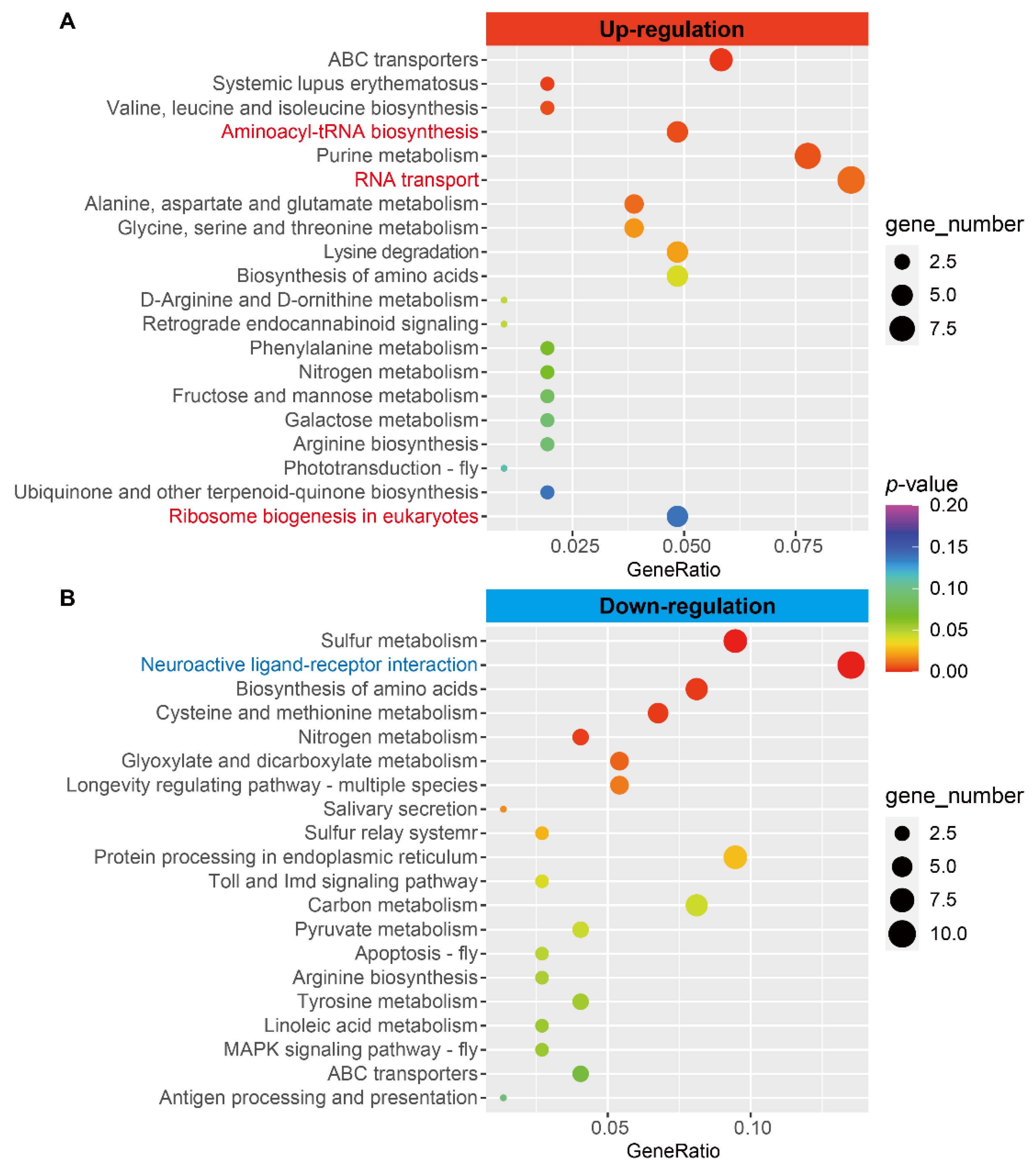
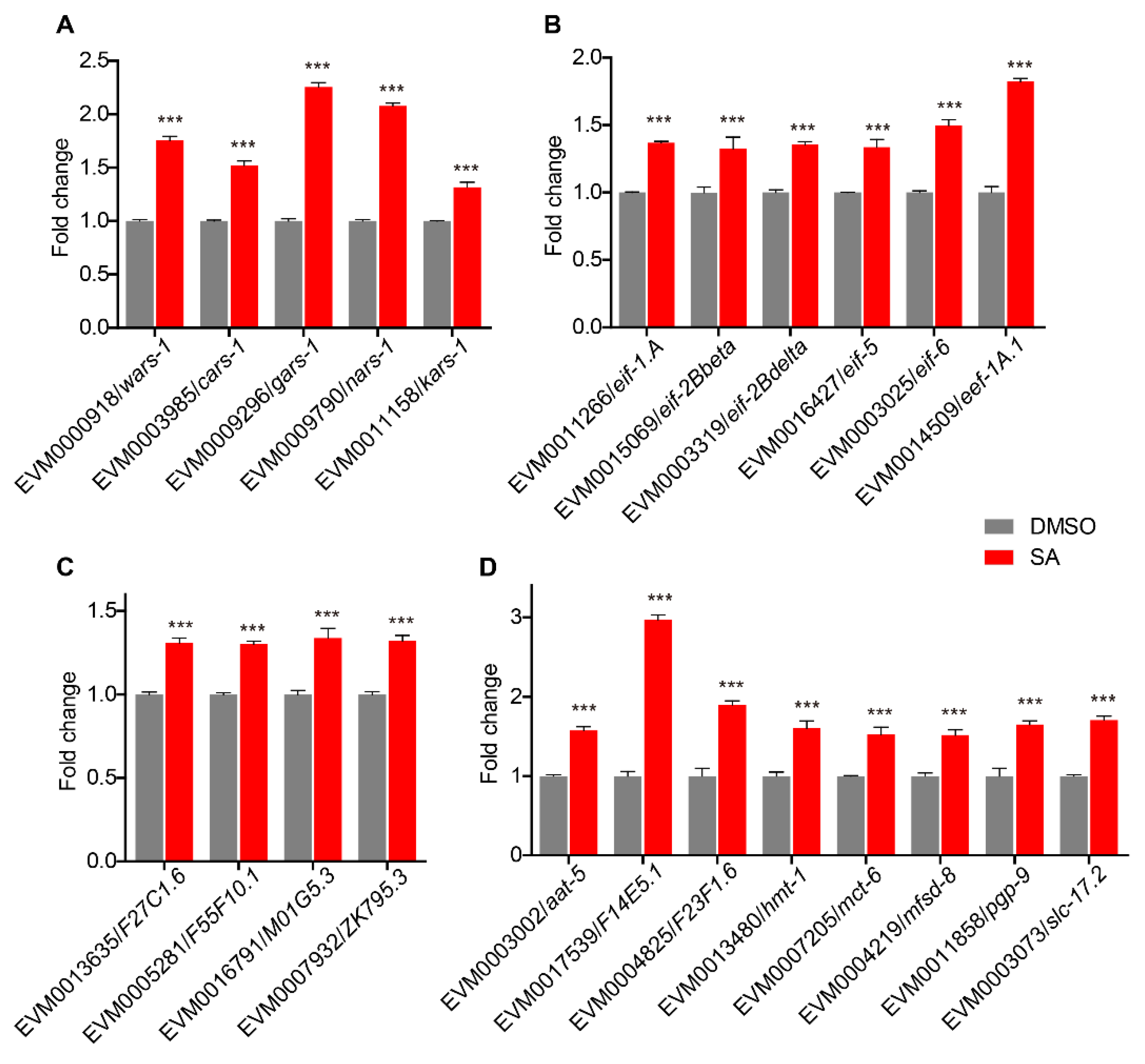
3.3. Upregulation of Aminoacyl-tRNA Biosynthesis Pathway Genes
3.4. Upregulated Expression of Translation Initiation, Elongation, and Ribosome Biogenesis-Related Genes
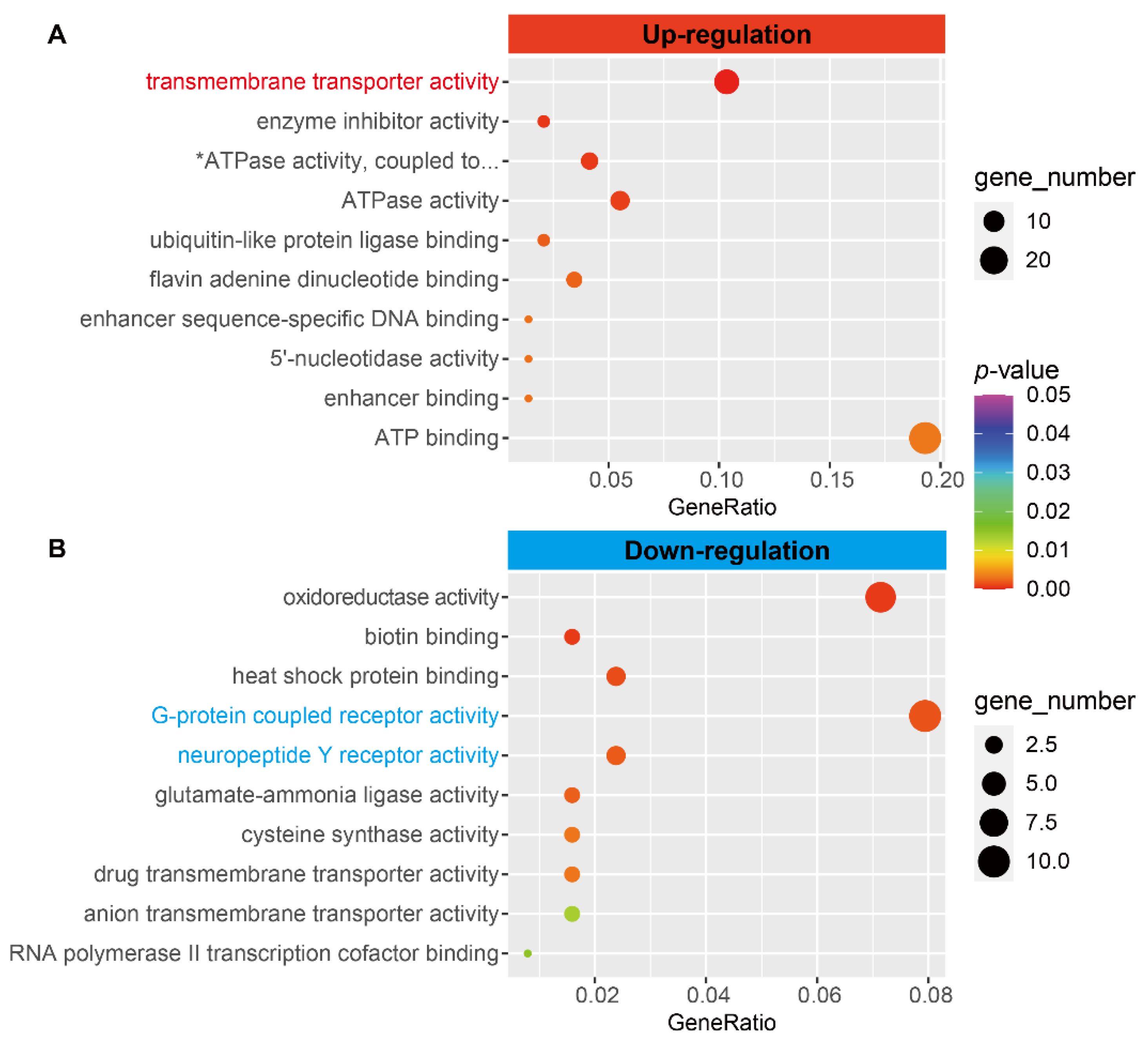
3.5. The Expression of Transmembrane Transporters Related Genes Was Upregulated
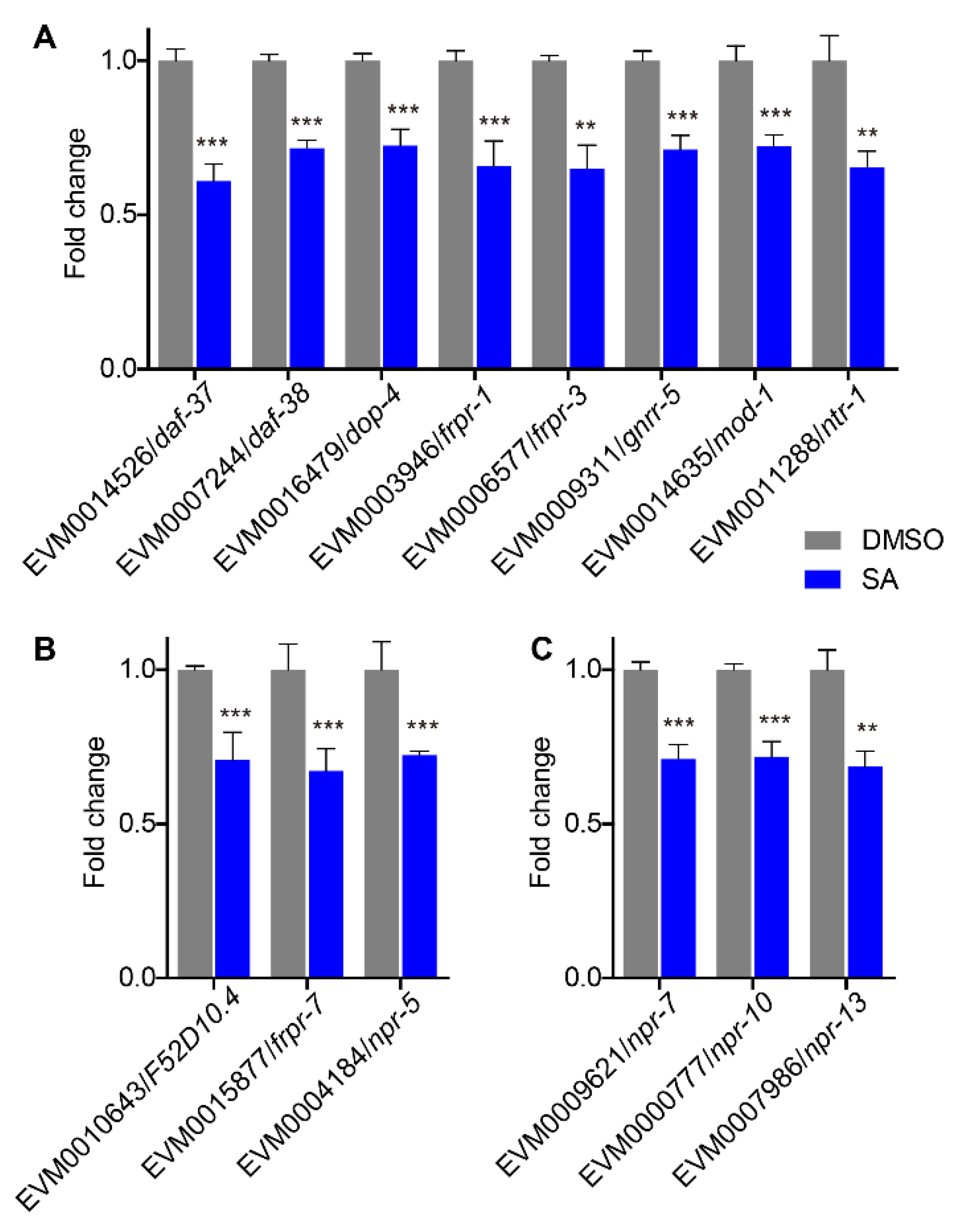
3.6. Downregulation of Neuronal Signaling Genes
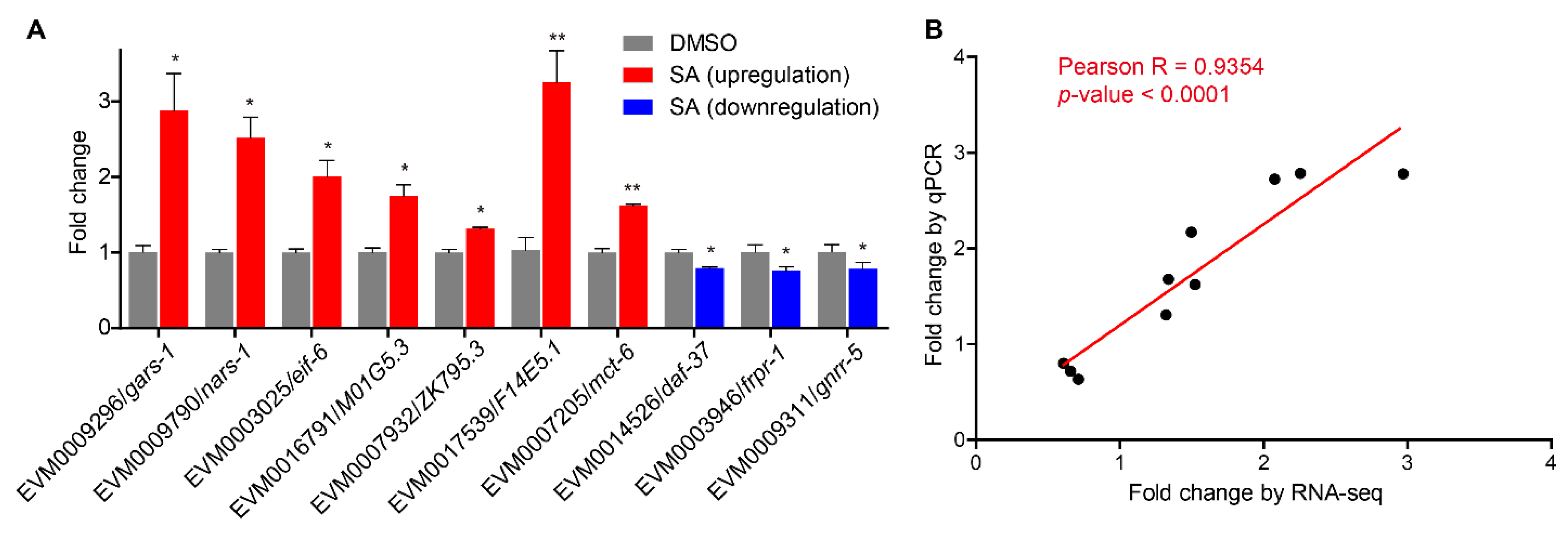
3.7. Quantitative Real-Time PCR Validation
4. Discussion
4.1. Stearic Acid Might Promote L. marina Development through Upregulation of Protein Synthesis Related Genes
4.2. Stearic Acid Might Promote L. marina Development via Upregulation of Transmembrane Transporter Genes
4.3. Stearic Acid Might Promote Development of L. marina through Reduced Neuronal Signaling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sampath, H.; Ntambi, J.M. The fate and intermediary metabolism of stearic acid. Lipids 2005, 40, 1187–1191. [Google Scholar] [CrossRef] [PubMed]
- Spady, D.; Woollett, L.; Dietschy, J. Regulation of plasma LDL-cholesterol levels by dietary cholesterol and fatty acids. Annu. Rev. Nutr. 1993, 13, 355–381. [Google Scholar] [CrossRef] [PubMed]
- Siri-Tarino, P.W.; Sun, Q.; Hu, F.B.; Krauss, R.M. Saturated Fatty Acids and Risk of Coronary Heart Disease: Modulation by Replacement Nutrients. Curr. Atheroscler. Rep. 2010, 12, 384–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, J.E.; Zhang, J.; Kris-Etherton, P.M. Cardiovascular disease risk of dietary stearic acid compared with trans, other saturated, and unsaturated fatty acids: A systematic review. Am. J. Clin. Nutr. 2010, 91, 46–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonanome, A.; Grundy, S.M. Effect of Dietary Stearic Acid on Plasma Cholesterol and Lipoprotein Levels. N. Engl. J. Med. 1988, 318, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Grande, F.; Anderson, J.T.; Keys, A. Comparison of Effects of Palmitic and Stearic Acids in the Diet on Serum Cholesterol in Man. Am. J. Clin. Nutr. 1970, 23, 1184–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, J.A.; Fong, J.; Bernert, J.T., Jr. Serum fatty acids and blood pressure. Hypertension 1996, 27, 303–307. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Griel, A.E.; Psota, T.L.; Gebauer, S.K.; Zhang, J.; Etherton, T.D. Dietary stearic acid and risk of cardiovascular disease: Intake, sources, digestion, and absorption. Lipids 2005, 40, 1193–1200. [Google Scholar] [CrossRef]
- Kühn, T.; Floegel, A.; Sookthai, D.; Johnson, T.; Rolle-Kampczyk, U.; Otto, W.; Von Bergen, M.; Boeing, H.; Kaaks, R. Higher plasma levels of lysophosphatidylcholine 18:0 are related to a lower risk of common cancers in a prospective metabolomics study. BMC Med. 2016, 14, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondul, A.M.; Moore, S.C.; Weinstein, S.J.; Karoly, E.D.; Sampson, J.N.; Albanes, D. Metabolomic analysis of prostate cancer risk in a prospective cohort: The alpha-tocopherol, beta-carotene cancer prevention (ATBC) study. Int. J. Cancer 2015, 137, 2124–2132. [Google Scholar] [CrossRef]
- Cross, A.J.; Moore, S.; Boca, S.; Huang, W.-Y.; Xiong, X.; Stolzenberg-Solomon, R.; Sinha, R.; Sampson, J.N. A prospective study of serum metabolites and colorectal cancer risk. Cancer 2014, 120, 3049–3057. [Google Scholar] [CrossRef] [PubMed]
- Van Kappel, A.-L.; Winkvist, A.; Kaaks, R.; Lenner, P.; Riboli, E. Fatty-acid composition in serum phospholipids and risk of breast cancer: An incident case-control study in Sweden. Int. J. Cancer 1999, 83, 585–590. [Google Scholar] [CrossRef]
- Nůsková, H.; Serebryakova, M.V.; Ferrer-Caelles, A.; Sachsenheimer, T.; Lüchtenborg, C.; Miller, A.K.; Brügger, B.; Kordyukova, L.V.; Teleman, A.A. Stearic acid blunts growth-factor signaling via oleoylation of GNAI proteins. Nat. Commun. 2021, 12, 1–15. [Google Scholar] [CrossRef]
- Senyilmaz, D.; Virtue, S.; Xu, X.; Tan, C.Y.; Griffin, J.L.; Miller, A.; Vidal-Puig, A.; Teleman, A.A. Regulation of mitochondrial morphology and function by stearoylation of TFR1. Nature 2015, 525, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senyilmaz-Tiebe, D.; Pfaff, D.H.; Virtue, S.; Schwarz, K.V.; Fleming, T.; Altamura, S.; Muckenthaler, M.U.; Okun, J.G.; Vidal-Puig, A.; Nawroth, P. Dietary stearic acid regulates mitochondria in vivo in humans. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gualberto, D.G.; Guo, X.; Correa, P.; Jee, C.; Garcia, L.R. TMC-1 attenuates C. elegans development and sexual behaviour in a chemically defined food environment. Nat. Commun. 2015, 6, 6345. [Google Scholar] [CrossRef] [Green Version]
- Cong, Y.; Yang, H.; Zhang, P.; Xie, Y.; Cao, X.; Zhang, L. Transcriptome Analysis of the Nematode Caenorhabditis elegans in Acidic Stress Environments. Front. Physiol. 2020, 11, 1107. [Google Scholar] [CrossRef]
- Goetsch, K.; Lu, N. Carbohydrate Requirement of Caenorhabditis Elegans and the Final Development of a Chemically Defined Medium. Nematologica 1993, 39, 303–311. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, N.J.; Kozak, E.; Conley, A.C. Chemically defined medium and Caenorhabditis elegans. BMC Biotechnol. 2003, 3, 19. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, N.J.; Udranszky, I.A.; Kozak, E.; Sunga, J.; Kim, S.K.; Jacobson, L.A.; Conley, C.A. Delayed development and lifespan extension as features of metabolic lifestyle alteration in C. elegans under dietary restriction. J. Exp. Biol. 2006, 209, 4129–4139. [Google Scholar] [CrossRef] [Green Version]
- Szewczyk, N.J.; Mancinelli, R.L.; McLamb, W.; Reed, D.; Blumberg, B.S.; Conley, C.A. Caenorhabditis elegans Survives Atmospheric Breakup of STS-107, Space Shuttle Columbia. Astrobiology 2005, 5, 690–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewczyk, N.; Tillman, J.; Conley, C.; Granger, L.; Segalat, L.; Higashitani, A.; Honda, S.; Honda, Y.; Kagawa, H.; Adachi, R.; et al. Description of International Caenorhabditis elegans Experiment first flight (ICE-FIRST). Adv. Space Res. 2008, 42, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Xie, Y.; Xue, B.; Yang, H.; Garcia, L.R.; Zhang, L. Fatty acid metabolic reprogramming promotes C. elegans Development. Biorxiv 2021. [Google Scholar] [CrossRef]
- Derycke, S.; De Meester, N.; Rigaux, A.; Creer, S.; Bik, H.; Thomas, W.; Moens, T. Coexisting cryptic species of the Litoditis marina complex (Nematoda) show differential resource use and have distinct microbiomes with high intraspecific variability. Mol. Ecol. 2016, 25, 2093–2110. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Zhang, P.; Xue, B.; Cao, X.; Ren, X.; Wang, L.; Sun, Y.; Yang, H.; Zhang, L. Establishment of a marine nematode model for animal functional genomics, environmental adaptation and developmental evolution. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, F.; Gao, S.; Liang, Y.; Long, H.; Lv, Z.; Su, Y.; Ye, N.; Zhang, L.; Zhao, C.; et al. Biodiversity-based development and evolution: The emerging research systems in model and non-model organisms. Sci. China Life Sci. 2021, 64, 1236–1280. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, P.; Zhang, L. Genome-Wide Transcriptional Responses of Marine Nematode Litoditis marina to Hyposaline and Hypersaline Stresses. Front. Physiol. 2021, 12, 672099. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Ibba, M.; Söll, D. Aminoacyl-tRNA synthesis. Annu. Rev. Biochem. 2000, 69, 617–650. [Google Scholar] [CrossRef] [PubMed]
- McLaskey, A.K.; Keister, J.E.; Yebra, L. Individual growth rate (IGR) and aminoacyl-tRNA synthetases (AARS) activity as individual-based indicators of growth rate of North Pacific krill, Euphausia pacifica. J. Exp. Mar. Biol. Ecol. 2020, 527, 151360. [Google Scholar] [CrossRef]
- Yebra, L.; Hernández-León, S. Aminoacyl-tRNA synthetases activity as a growth index in zooplankton. J. Plankton Res. 2004, 26, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Kwon, N.H.; Fox, P.L.; Kim, S. Aminoacyl-tRNA synthetases as therapeutic targets. Nat. Rev. Drug Discov. 2019, 18, 629–650. [Google Scholar] [CrossRef]
- Nakielny, S.; Fischer, U.; Michael, W.M.; Dreyfuss, G. RNA transport. Annu. Rev. Neurosci. 1997, 20, 269–301. [Google Scholar] [CrossRef] [Green Version]
- Basyuk, E.; Rage, F.; Bertrand, E. RNA transport from transcription to localized translation: A single molecule perspective. RNA Biol. 2021, 18, 1–17. [Google Scholar] [CrossRef]
- Fernandopulle, M.S.; Lippincott-Schwartz, J.; Ward, M.E. RNA transport and local translation in neurodevelopmental and neurodegenerative disease. Nat. Neurosci. 2021, 24, 622–632. [Google Scholar] [CrossRef]
- Thomson, E.; Ferreira-Cerca, S.; Hurt, E. Eukaryotic ribosome biogenesis at a glance. J. Cell Sci. 2013, 126, 4815–4821. [Google Scholar] [CrossRef] [Green Version]
- Lempiäinen, H.; Shore, D. Growth control and ribosome biogenesis. Curr. Opin. Cell Biol. 2009, 21, 855–863. [Google Scholar] [CrossRef] [PubMed]
- Ritter, S.L.; Hall, R. Fine-tuning of GPCR activity by receptor-interacting proteins. Nat. Rev. Mol. Cell Biol. 2009, 10, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michel, M.C.; Beck-Sickinger, A.; Cox, H.; Doods, H.N.; Herzog, H.; Larhammar, D.; Quirion, R.; Schwartz, T.; Westfall, T. XVI. International Union of Pharmacology recommendations for the nomenclature of neuropeptide Y, peptide YY, and pancreatic polypeptide receptors. Pharmacol. Rev. 1998, 50, 143–150. [Google Scholar] [PubMed]
- Meng, Y.; Yuan, C.; Zhang, J.; Zhang, F.; Fu, Q.; Zhu, X.; Shu, G.; Wang, L.; Gao, P.; Xi, Q.; et al. Stearic acid suppresses mammary gland development by inhibiting PI3K/Akt signaling pathway through GPR120 in pubertal mice. Biochem. Biophys. Res. Commun. 2017, 491, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Habib, N.A.; Wood, C.B.; Apostolov, K.; Barker, W.; Hershman, M.J.; Aslam, M.; Heinemann, D.; Fermor, B.; Williamson, R.C.; Jenkins, W.E. Stearic acid and carcinogenesis. Br. J. Cancer 1987, 56, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.S. Effect of dietary stearic acid on the genesis of spontaneous mammary adenocarcinomas in strain A/ST mice. Int. J. Cancer 1984, 34, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Beesley, J.S.; Soutter, W.P.; White, J.O. Effect of Stearic Acid on Human Cervical Cancer Cell Growth. In Eicosanoids and Other Bioactive Lipids in Cancer, Inflammation and Radiation Injury; Springer: Boston, MA, USA, 1993; pp. 573–575. [Google Scholar]
- Li, Y.; Wu, H.; Zhang, R.; Shu, G.; Wang, S.; Gao, P.; Zhu, X.; Jiang, Q.; Wang, L. Diet containing stearic acid increases food reward-related behaviors in mice compared with oleic acid. Brain Res. Bull. 2020, 164, 45–54. [Google Scholar] [CrossRef] [PubMed]
- Bajracharya, R.; Bustamante, S.; Ballard, O.J.W. Stearic Acid Supplementation in High Protein to Carbohydrate (P:C) Ratio Diet Improves Physiological and Mitochondrial Functions of Drosophila melanogaster parkin Null Mutants. J. Gerontol. Ser. A 2019, 74, 1564–1572. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, L.; Zhang, L.; Wang, W.; Wei, S.; Wang, J.; Che, H.; Zhang, Y. Effects of excess sugars and lipids on the growth and development of Caenorhabditis elegans. Genes Nutr. 2020, 15, 1–18. [Google Scholar] [CrossRef]
- Hussain, T.; Yogavel, M.; Sharma, A. Inhibition of protein synthesis and malaria parasite development by drug targeting of methionyl-tRNA synthetases. Antimicrob. Agents Chemother. 2015, 59, 1856–1867. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Bergert, M.; Walther, A.; Suter, B. Double-sieving-defective aminoacyl-tRNA synthetase causes protein mistranslation and affects cellular physiology and development. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaminska M, Havrylenko S, Decottignies P, Gillet S, Le Maréchal P, Negrutskii B, Mirande M: Dissection of the structural organization of the aminoacyl-tRNA synthetase complex. J. Biol. Chem. 2009, 284, 6053–6060. [CrossRef] [Green Version]
- Havrylenko, S.; Legouis, R.; Negrutskii, B.; Mirande, M. Caenorhabditis elegans evolves a new architecture for the multi-aminoacyl-tRNA synthetase complex. J. Biol. Chem. 2011, 286, 28476–28487. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, S.; Borgo, B.; Pazdernik, N.; Fox, P.; Mardis, E.R.; Kohara, Y.; Havranek, J.; Schedl, T. Caenorhabditis elegans glp-4 Encodes a Valyl Aminoacyl tRNA Synthetase. G3 Genes Genomes Genet. 2015, 5, 2719–2728. [Google Scholar] [CrossRef] [Green Version]
- Geillinger, K.E.; Kuhlmann, K.; Eisenacher, M.; Meyer, H.E.; Daniel, H.; Spanier, B. Dynamic Changes of the Caenorhabditis elegans Proteome during Ontogenesis Assessed by Quantitative Analysis with 15N Metabolic Labeling. J. Proteome Res. 2012, 11, 4594–4604. [Google Scholar] [CrossRef]
- Jackson, R.J.; Hellen, C.U.T.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Aitken, C.E.; Lorsch, J.R. A mechanistic overview of translation initiation in eukaryotes. Nat. Struct. Mol. Biol. 2012, 19, 568–576. [Google Scholar] [CrossRef]
- Hanazawa, M.; Kawasaki, I.; Kunitomo, H.; Gengyo-Ando, K.; Bennett, K.L.; Mitani, S.; Iino, Y. The Caenorhabditis elegans eukaryotic initiation factor 5A homologue, IFF-1, is required for germ cell proliferation, gametogenesis and localization of the P-granule component PGL-1. Mech. Dev. 2004, 121, 213–224. [Google Scholar] [CrossRef]
- Yu, X.; Vought, V.E.; Conradt, B.; Maine, E.M. Eukaryotic translation initiation factor 5B activity regulates larval growth rate and germline development in Caenorhabditis elegans. Genesis 2006, 44, 412–418. [Google Scholar] [CrossRef]
- Andersen, G.R.; Nissen, P.; Nyborg, J. Elongation factors in protein biosynthesis. Trends Biochem. Sci. 2003, 28, 434–441. [Google Scholar] [CrossRef]
- Carr-Schmid, A.; Valente, L.; Loik, V.I.; Williams, T.; Starita, L.M.; Kinzy, T.G. Mutations in elongation factor 1β, a guanine nucleotide exchange factor, enhance translational fidelity. Mol. Cell. Biol. 1999, 19, 5257–5266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Gao, Y.; Li, Z.; Chen, L.; Xia, Z.; Xu, T.; Qin, Y. Mitochondrial EF4 links respiratory dysfunction and cytoplasmic translation in Caenorhabditis elegans. Biochim. Biophys. Acta 2014, 1837, 1674–1683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamath, R.S.; Fraser, A.G.; Dong, Y.; Poulin, G.; Durbin, R.; Gotta, M.; Kanapin, A.; Le Bot, N.; Moreno, S.; Sohrmann, M.; et al. Systematic functional analysis of the Caenorhabditis elegans genome using RNAi. Nature 2003, 421, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Melo, J.A.; Ruvkun, G. Inactivation of Conserved C. elegans Genes Engages Pathogen- and Xenobiotic-Associated Defenses. Cell 2012, 149, 452–466. [Google Scholar] [CrossRef] [Green Version]
- Fraser, A.G.; Kamath, R.S.; Zipperlen, P.; Martinez-Campos, M.; Sohrmann, M.; Ahringer, J. Functional genomic analysis of C. elegans chromosome I by systematic RNA interference. Nature 2000, 408, 325–330. [Google Scholar] [CrossRef]
- Saijou, E.; Fujiwara, T.; Suzaki, T.; Inoue, K.; Sakamoto, H. RBD-1, a nucleolar RNA-binding protein, is essential for Caenorhabditis elegans early development through 18S ribosomal RNA processing. Nucleic Acids Res. 2004, 32, 1028–1036. [Google Scholar] [CrossRef] [Green Version]
- Kudron, M.M.; Reinke, V. C. elegans Nucleostemin Is Required for Larval Growth and Germline Stem Cell Division. PLoS Genet. 2008, 4, e1000181. [Google Scholar] [CrossRef]
- Miguel-Aliaga, I. Nerveless and gutsy: Intestinal nutrient sensing from invertebrates to humans. Semin. Cell Dev. Biol. 2012, 23, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Palm, W.; Thompson, C.B. Nutrient acquisition strategies of mammalian cells. Nature 2017, 546, 234–242. [Google Scholar] [CrossRef]
- Thwaites, D.T.; Anderson, C. H+-coupled nutrient, micronutrient and drug transporters in the mammalian small intestine. Exp. Physiol. 2007, 92, 603–619. [Google Scholar] [CrossRef] [Green Version]
- Prudente, S.; Flex, E.; Morini, E.; Turchi, F.; Capponi, D.; De Cosmo, S.; Tassi, V.; Guida, V.; Avogaro, A.; Folli, F.; et al. A Functional Variant of the Adipocyte Glycerol Channel Aquaporin 7 Gene Is Associated with Obesity and Related Metabolic Abnormalities. Diabetes 2007, 56, 1468–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, R.; Sarioglu, S.; Sökmen, S.; Füzün, M.; Küpelioğlu, A.; Valentine, H.; Görken, I.B.; Airley, R.; West, C. Glucose transporter-1 (GLUT-1): A potential marker of prognosis in rectal carcinoma? Br. J. Cancer 2003, 89, 870–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macheda, M.L.; Rogers, S.; Best, J.D. Molecular and cellular regulation of glucose transporter (GLUT) proteins in cancer. J. Cell. Physiol. 2005, 202, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Benner, J. Amino acid homeostasis in Caenorhabditis elegans lacking the intestinal peptide transporter PEPT-1 and identification of PEPT-1 modulator proteins. Ph.D. Thesis, Technical University of Munich, Freising, Germany, 2011. [Google Scholar]
- Schwartz, M.S.; Benci, J.L.; Selote, D.S.; Sharma, A.K.; Chen, A.G.Y.; Dang, H.; Fares, H.; Vatamaniuk, O.K. Detoxification of Multiple Heavy Metals by a Half-Molecule ABC Transporter, HMT-1, and Coelomocytes of Caenorhabditis elegans. PLoS ONE 2010, 5, e9564. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Sharma, A.K.; Vatamaniuk, O.K. N-terminal extension and C-terminal domains are required for ABCB6/HMT-1 protein interactions, function in cadmium detoxification, and localization to the endosomal-recycling system in Caenorhabditis elegans. Front. Physiol. 2018, 9, 885. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Fang, L.L.; Johnsen, R.; Baillie, D.L. ATP-binding cassette protein E is involved in gene transcription and translation in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2004, 323, 104–111. [Google Scholar] [CrossRef]
- Bagchi, S.; Perland, E.; Hosseini, K.; Lundgren, J.; Al-Walai, N.; Kheder, S.; Fredriksson, R. Probable role for major facilitator superfamily domain containing 6 (MFSD6) in the brain during variable energy consumption. Int. J. Neurosci. 2020, 130, 476–489. [Google Scholar] [CrossRef]
- Berger, J.H.; Charron, M.J.; Silver, D.L. Major Facilitator Superfamily Domain-Containing Protein 2a (MFSD2A) Has Roles in Body Growth, Motor Function, and Lipid Metabolism. PLoS ONE 2012, 7, e50629. [Google Scholar] [CrossRef] [Green Version]
- Lekholm, E.; Perland, E.; Eriksson, M.M.; Hellsten, S.V.; Lindberg, F.; Rostami, J.; Fredriksson, R. Putative Membrane-Bound Transporters MFSD14A and MFSD14B Are Neuronal and Affected by Nutrient Availability. Front. Mol. Neurosci. 2017, 10, 11. [Google Scholar] [CrossRef] [Green Version]
- Perland, E.; Lekholm, E.; Eriksson, M.M.; Bagchi, S.; Arapi, V.; Fredriksson, R. The Putative SLC Transporters Mfsd5 and Mfsd11 Are Abundantly Expressed in the Mouse Brain and Have a Potential Role in Energy Homeostasis. PLoS ONE 2016, 11, e0156912. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.W.; Tang, N.H.; Piggott, C.A.; Andrusiak, M.G.; Park, S.; Zhu, M.; Kurup, N.; Cherra, S.J., III; Wu, Z.; Chisholm, A.D. Expanded genetic screening in Caenorhabditis elegans identifies new regulators and an inhibitory role for NAD+ in axon regeneration. Elife 2018, 7, e39756. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.J.; Church, D.L.; Hatzold, J.; Conradt, B.; Lambie, E.J. Gem-1 encodes an SLC16 monocarboxylate transporter-related protein that functions in parallel to the gon-2 TRPM channel during gonad development in Caenorhabditis elegans. Genetics 2009, 181, 581–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogurusu, T.; Sakata, K. The Caenorhabditis elegans homolog of human mitochondrial pyrimidine nucleotide transporter regulates glucose transport. Biochem. Biophys. Res. Commun. 2021, 557, 117–121. [Google Scholar] [CrossRef]
- Avery, L.; You, Y.-J. C. elegans Feeding (21 May 2012), WormBook, Ed. The C. elegans Research Community, WormBook. Available online: http://www.wormbook.org/chapters/www_feeding/feeding.html (accessed on 4 February 2022).
- Gallagher, T.; Kim, J.; Oldenbroek, M.; Kerr, R.; You, Y.-J. ASI Regulates Satiety Quiescence in C. elegans. J. Neurosci. 2013, 33, 9716–9724. [Google Scholar] [CrossRef] [Green Version]
- Ben Arous, J.; Laffont, S.; Chatenay, D. Molecular and sensory basis of a food related two-state behavior in C. elegans. PLoS ONE 2009, 4, e7584. [Google Scholar] [CrossRef] [Green Version]
- Yapici, N.; Cohn, R.; Schusterreiter, C.; Ruta, V.; Vosshall, L.B. A Taste Circuit that Regulates Ingestion by Integrating Food and Hunger Signals. Cell 2016, 165, 715–729. [Google Scholar] [CrossRef] [Green Version]
- Herman, A.M.; Ortiz-Guzman, J.; Kochukov, M.; Herman, I.; Quast, K.B.; Patel, J.; Tepe, B.; Carlson, J.C.; Ung, K.; Selever, J.; et al. A cholinergic basal forebrain feeding circuit modulates appetite suppression. Nature 2016, 538, 253–256. [Google Scholar] [CrossRef] [Green Version]
- Mineur, Y.S.; Abizaid, A.; Rao, Y.; Salas, R.; DiLeone, R.J.; Gündisch, D.; Diano, S.; De Biasi, M.; Horvath, T.L.; Gao, X.-B.; et al. Nicotine Decreases Food Intake Through Activation of POMC Neurons. Science 2011, 332, 1330–1332. [Google Scholar] [CrossRef] [Green Version]
- Park, D.; O’Doherty, I.; Somvanshi, R.K.; Bethke, A.; Schroeder, F.C.; Kumar, U.; Riddle, D.L. Interaction of structure-specific and promiscuous G-protein–coupled receptors mediates small-molecule signaling in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2012, 109, 9917–9922. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, M.; Fuke, S.; Suo, S.; Sasagawa, N.; Van Tol, H.H.M.; Ishiura, S. Characterization of a novel D2-like dopamine receptor with a truncated splice variant and a D1-like dopamine receptor unique to invertebrates from Caenorhabditis elegans. J. Neurochem. 2005, 94, 1146–1157. [Google Scholar] [CrossRef]
- Nagashima, T.; Oami, E.; Kutsuna, N.; Ishiura, S.; Suo, S. Dopamine regulates body size in Caenorhabditis elegans. Dev. Biol. 2016, 412, 128–138. [Google Scholar] [CrossRef]
- Ezcurra, M.; Tanizawa, Y.; Swoboda, P. Schafer WR: Food sensitizes C. elegans avoidance behaviours through acute dopamine signalling. EMBO J. 2011, 30, 1110–1122. [Google Scholar] [CrossRef] [Green Version]
- Chew, Y.L.; Tanizawa, Y.; Cho, Y.; Zhao, B.; Alex, J.Y.; Ardiel, E.L.; Rabinowitch, I.; Bai, J.; Rankin, C.H.; Lu, H. An Afferent Neuropeptide System Transmits Mechanosensory Signals Triggering Sensitization and Arousal in C. elegans. Neuron 2018, 99, 1233–1246. [Google Scholar] [CrossRef] [Green Version]
- Oranth, A.; Schultheis, C.; Tolstenkov, O.; Erbguth, K.; Nagpal, J.; Hain, D.; Brauner, M.; Wabnig, S.; Costa, W.S.; McWhirter, R.D.; et al. Food Sensation Modulates Locomotion by Dopamine and Neuropeptide Signaling in a Distributed Neuronal Network. Neuron 2018, 100, 1414–1428.e10. [Google Scholar] [CrossRef] [Green Version]
- Ranganathan, R.; Cannon, S.; Horvitz, H.R. MOD-1 is a serotonin-gated chloride channel that modulates locomotory behaviour in C. elegans. Nature 2000, 408, 470–475. [Google Scholar] [CrossRef]
- Srinivasan, S.; Sadegh, L.; Elle, I.C.; Christensen, A.G.; Faergeman, N.J.; Ashrafi, K. Serotonin Regulates C. elegans Fat and Feeding through Independent Molecular Mechanisms. Cell Metab. 2008, 7, 533–544. [Google Scholar] [CrossRef] [Green Version]
- De Bono, M.; Bargmann, C.I. Natural variation in a neuropeptide Y receptor homolog modifies social behavior and food response in C. elegans. Cell 1998, 94, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.; Reale, V.; Olofsson, B.; Knights, A.; Evans, P.; de Bono, M. Coordinated Regulation of Foraging and Metabolism in C. elegans by RFamide Neuropeptide Signaling. Cell Metab. 2009, 9, 375–385. [Google Scholar] [CrossRef] [Green Version]
- Chow, E.S.H. The Caenorhabditis elegans ALA Neuron: Its Transcriptome and Role in Inducing Sleep. Ph.D. Thesis, California Institute of Technology, Pasadena, CA, USA, 2014. [Google Scholar]
- Hapiak, V.; Summers, P.; Ortega, A.; Law, W.J.; Stein, A.; Komuniecki, R. Neuropeptides Amplify and Focus the Monoaminergic Inhibition of Nociception in Caenorhabditis elegans. J. Neurosci. 2013, 33, 14107–14116. [Google Scholar] [CrossRef]
- Garrison, J.L.; Macosko, E.Z.; Bernstein, S.; Pokala, N.; Albrecht, D.R.; Bargmann, C.I. Oxytocin/Vasopressin-Related Peptides Have an Ancient Role in Reproductive Behavior. Science 2012, 338, 540–543. [Google Scholar] [CrossRef] [Green Version]
- Beets, I.; Janssen, T.; Meelkop, E.; Temmerman, L.; Suetens, N.; Rademakers, S.; Jansen, G.; Schoofs, L. Vasopressin/Oxytocin-Related Signaling Regulates Gustatory Associative Learning in C. elegans. Science 2012, 338, 543–545. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, X.; Sun, P.; Zhang, L. Transcriptome Analysis of the Marine Nematode Litoditis marina in a Chemically Defined Food Environment with Stearic Acid Supplementation. J. Mar. Sci. Eng. 2022, 10, 428. https://doi.org/10.3390/jmse10030428
Cao X, Sun P, Zhang L. Transcriptome Analysis of the Marine Nematode Litoditis marina in a Chemically Defined Food Environment with Stearic Acid Supplementation. Journal of Marine Science and Engineering. 2022; 10(3):428. https://doi.org/10.3390/jmse10030428
Chicago/Turabian StyleCao, Xuwen, Peiqi Sun, and Liusuo Zhang. 2022. "Transcriptome Analysis of the Marine Nematode Litoditis marina in a Chemically Defined Food Environment with Stearic Acid Supplementation" Journal of Marine Science and Engineering 10, no. 3: 428. https://doi.org/10.3390/jmse10030428