Efficacy of N-methyl-N-nitrosourea (MNU) Mutation on Enhancing the Yield and Quality of Rice

 ,
,  ,
,  ,
, 
 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Design
2.2. Plant Materials and Seedling Management
2.3. Measurements of Growth and Yield Attributes
2.4. Measurements of Physicochemical Properties
2.5. Statistical Analysis
3. Results
3.1. Growth Attributes
3.2. Yield and Its Components
3.3. Physicochemical Properties and Appearance
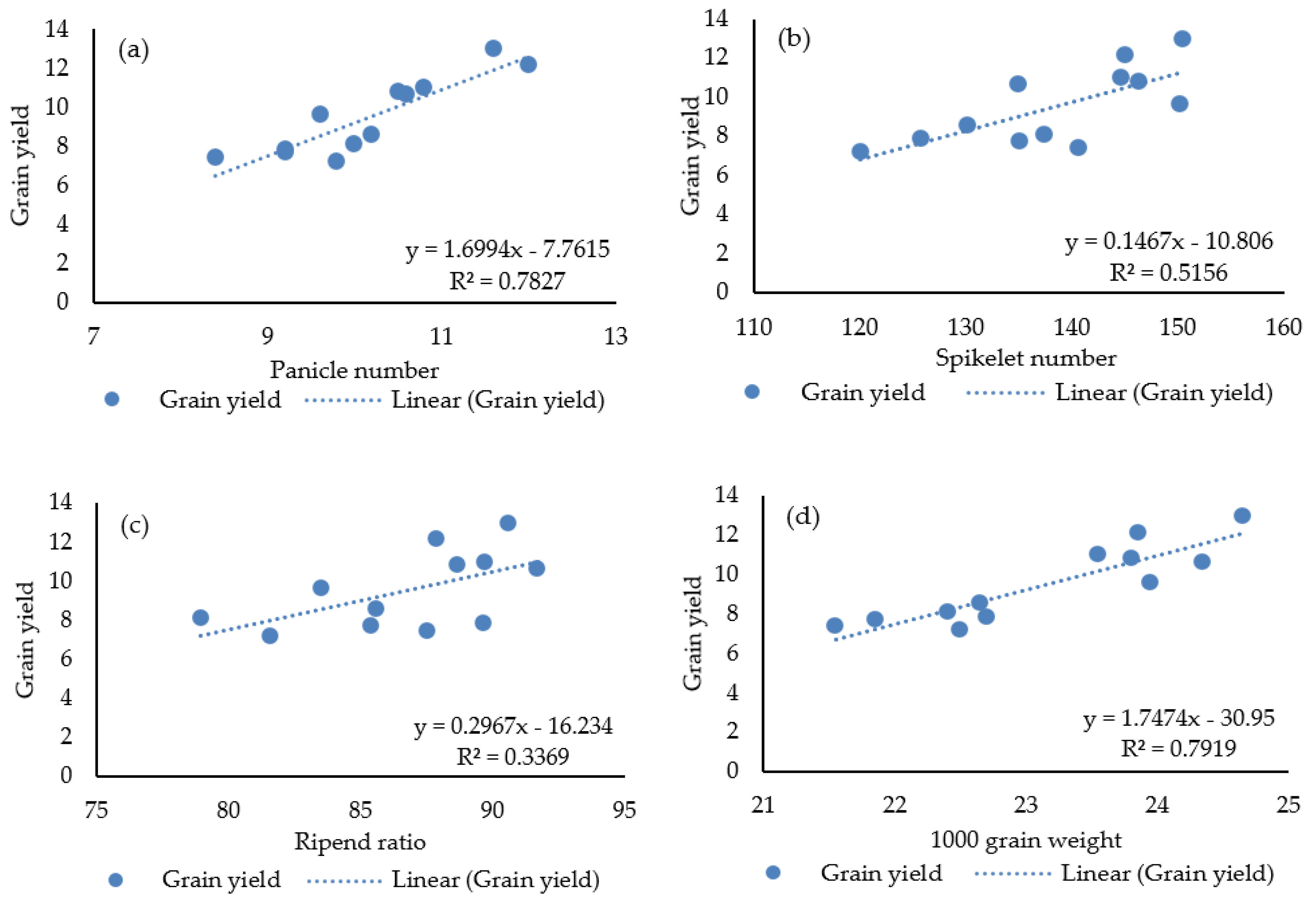
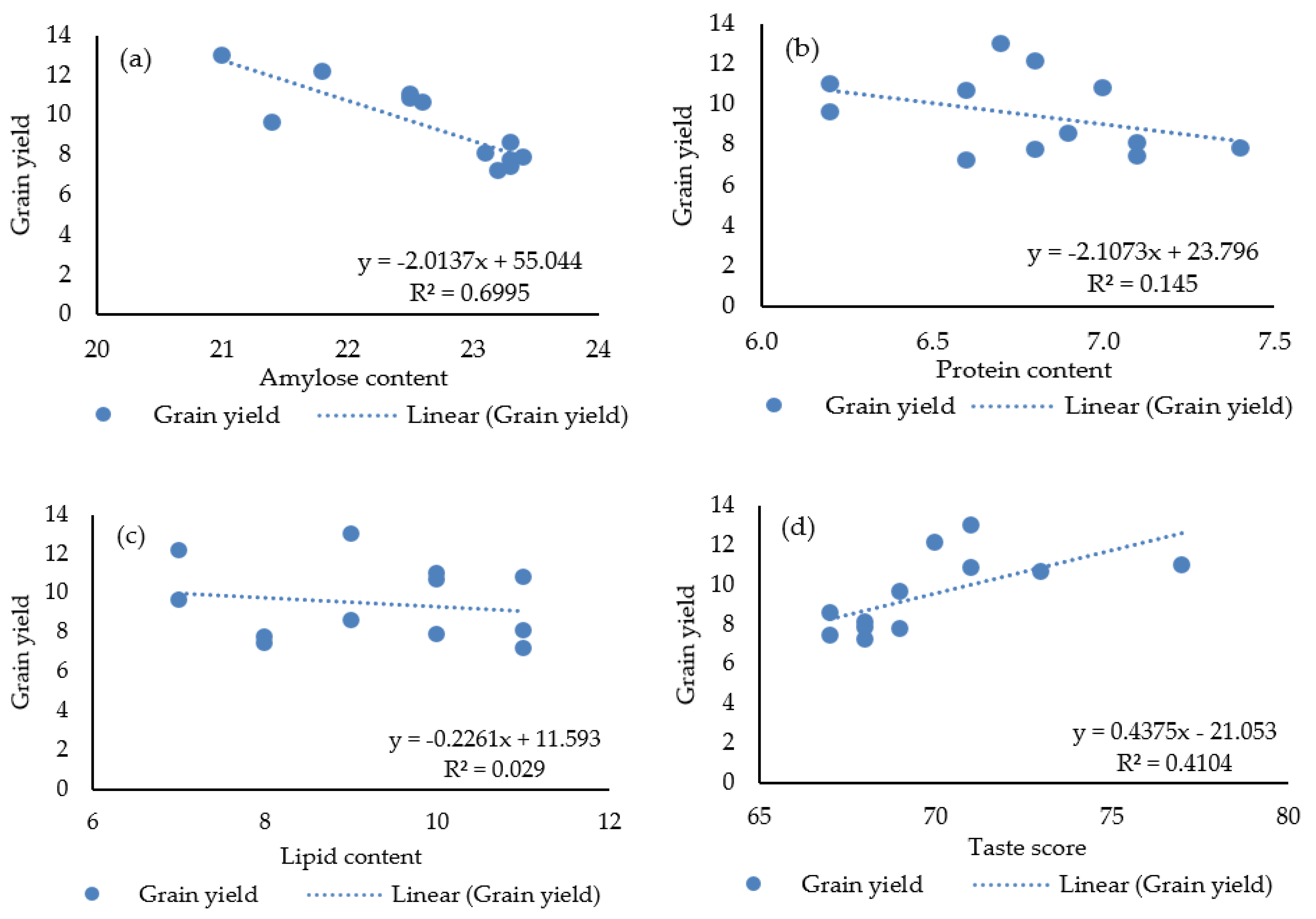
3.4. Relationship of Yield with Other Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kakar, K.; Xuan, T.D.; Haqani, M.I.; Rayee, R.; Wafa, I.K.; Abdiani, S.; Tran, H.D. Current situation and sustainable development for rice cultivation and production in Afghanistan. Agriculture 2019, 9, 49. [Google Scholar] [CrossRef]
- Kakar, K.; Xuan, T.D.; Abdiani, S.; Wafa, I.K.; Noori, Z.; Attai, S.; Khanh, T.D.; Tran, H.-D. Morphological observation and correlation of growth and yield characteristics with grain quality and antioxidant activities in exotic rice varieties of Afghanistan. Agriculture 2019, 9, 167. [Google Scholar] [CrossRef]
- Wang, N.; Long, T.; Yao, W.; Xiong, L.; Zhang, Q.; Wu, C. Mutant resources for the functional analysis of the rice genome. Mol. Plant 2013, 6, 596–604. [Google Scholar] [CrossRef] [PubMed]
- Kakar, K.; Nitta, Y.; Asagi, N.; Komatsuzaki, M.; Shiotau, F.; Kokubo, T.; Xuan, T.D. Morphological analysis on comparison of organic and chemical fertilizers on grain quality of rice at different planting densities. Plant Prod. Sci. 2019, 22, 1–9. [Google Scholar] [CrossRef]
- Anh, T.T.T.; Xuan, T.D.; Huong, C.T.; Dat, T.D. Phenotypic performance of rice (Oryza sativa L.) populations induced by the MNU mutant on the adaptive characteristics. J. Horti. Plant Res. 2019, 5, 13–24. [Google Scholar] [CrossRef]
- Breseghello, F.; Coelho, A.S.G. Traditional and modern plant breeding methods with examples in rice (Oryza sativa L.). J. Agric. Food Chem. 2013, 61, 8277–8286. [Google Scholar] [CrossRef] [PubMed]
- Sikora, P.; Chawade, A.; Larsson, M.; Olsson, J.; Olsson, O. Mutagenesis as a tool in plant genetics, functional genomics, and breeding. Int. J. Plant Genom. 2011, 2011, 314829. [Google Scholar] [CrossRef]
- Bentota, A.P. Mutation improvement of rice variety BW-267-3 for red pericarp grains and lodging resistance. Plant Mutat. Rep. 2006, 1, 42–43. [Google Scholar]
- El-Degwy, I.S. Mutation induced genetic variability in rice (Oryza sativa L.). Int. J. Agric. Crop Sci. 2013, 5, 2789–2794. [Google Scholar]
- Shadakshari, Y.G.; Chandrappa, H.M.; Kulkarni, R.S.; Shashidhar, H.E. Induction of beneficial mutants in rice (Oryza sativa L.). Indian J. Genet. Plant Breed. 2001, 61, 274–276. [Google Scholar]
- Riaz, A.; Gul, A. Plant mutagenesis and crop improvement. In Crop Production and Global Environment Issues; Hakeem, K., Ed.; Springer: Cham, Switzerland, 2015; pp. 181–209. [Google Scholar]
- Ahloowalia, B.S.; Maluszynski, M.; Nichterlein, K. Global impact of mutation-derived varieties. Euphytica 2004, 135, 187–204. [Google Scholar] [CrossRef]
- Xuan, T.D.; Bach, D.T.; Dat, T.D. Involvement of phenolics, flavonoids, and phenolic acids in high yield characteristics of rice (Oryza sativa L.). Int. Lett. Nat. Sci. 2018, 68, 19–26. [Google Scholar] [CrossRef]
- Fang, J.; Zhou, Y.; Zou, D.; Ning, J. Breeding of high yield and high protein rice varieties by space mutation. Acta Agric. Nucleatae Sin. 2004, 18, 280–283. [Google Scholar]
- Penna, S.; Jain, S.M. Mutant resources and mutagenomics in crop plants. Emir. J. Food Agric. 2017, 29, 651–657. [Google Scholar]
- Lo, S.F.; Fan, M.J.; Hsing, Y.I.; Chen, L.J.; Chen, S.; Wen, I.C.; Liu, Y.L.; Chen, K.T.; Jiang, M.J.; Lin, M.K.; et al. Genetic resources offer efficient tools for rice functional genomics research. Plant Cell Environ. 2016, 39, 998–1013. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.D.; Anh, T.T.T.; Tran, H.D.; Khanh, T.D.; Dat, T.D. Mutation breeding of a N-methyl-N-nitrosourea (MNU)-induced rice (Oryza sativa L. ssp. Indica) population for the yield attributing traits. Sustainability 2019, 11, 1062. [Google Scholar] [CrossRef]
- Anh, T.T.T.; Khanh, T.D.; Dat, T.D.; Xuan, T.D. Identification of phenotypic variation and genetic diversity in rice (Oryza sativa L.) mutants. Agriculture 2018, 8, 30. [Google Scholar] [CrossRef]
- Hussain, S.; Fujii, T.; McGoey, S.; Yamada, M.; Ramzan, M.; Akmal, M. Evaluation of different rice varieties for growth and yield characteristics. J. Anim. Plant Sci. 2014, 24, 1504–1510. [Google Scholar]
- Singh, N.; Sodhi, N.S.; Kaur, M.; Saxena, S.K. Physico-chemical, morphological, thermal, cooking and textural properties of chalky and translucent rice kernels. Food Chem. 2003, 82, 433–439. [Google Scholar] [CrossRef]
- Ashrafuzzaman, M.; Islam, M.R.; Ismail, M.R.; Shahidullah, S.M.; Hanafi, M.M. Evaluation of six aromatic rice varieties for yield and yield contributing characters. Int. J. Agric. Biol. 2009, 11, 616–620. [Google Scholar]
- Sultana, S.; Ali, M.Y. Effect of different nitrogen levels on the yield performance of aromatic rice varieties. Bull. Inst. Trop. Agric. Kyushu Univ. 2014, 37, 47–56. [Google Scholar]
- Kanegana, M.Z.; Kargbo, A.M. Effect of Nitrogen Topdressing at Different Growth Stages on Rice Plant Growth, Yield and Yield Components; Report on Experiments in Rice Cultivation Techniques Development Course; Japan Int. Coop. Agency: Tsukuba, Japan, 2011; pp. 43–45. [Google Scholar]
- Feng, L.Y.; Raza, M.A.; Li, Z.C.; Chen, Y.; Khalid, M.H.B.; Du, J.; Liu, W.; Wu, X.; Song, C.; Yu, L.; et al. The influence of light intensity and leaf movement on photosynthesis characteristics and carbon balance of soybean. Front. Plant Sci. 2019, 9, 1952. [Google Scholar] [CrossRef] [PubMed]
- Badshah, M.A.; Naimei, T.; Zou, Y.; Ibrahim, M.; Wang, K. Yield and tillering response of super hybrid rice Liangyoupeijiu to tillage and establishment methods. Crop J. 2014, 2, 79–86. [Google Scholar] [CrossRef]
- Champagne, E.T. Rice: Chemistry and Technology, 3rd ed.; American Association of Cereal Chemists: Eagan, MN, USA, 2004; p. 640. [Google Scholar]
- Laza, M.R.C.; Peng, S.; Akita, S.; Saka, H. Effect of panicle size on grain yield of IRRI-released indica rice cultivars in the wet season. Plant Prod. Sci. 2004, 7, 271–276. [Google Scholar] [CrossRef]
- Kasim, N.; Sjahril, R.; Riadi, M.; Arbie, F. The phenotype performance of M3 red rice mutant (Oryza sativa L.). IOP Conf. Ser. Earth Environ. Sci. 2018, 157, 012047. [Google Scholar] [CrossRef]
- Ali, K.; Arif, M.; Shah, S.; Hussain, Z.; Ali, A.; Munir, S.; Sher, H. Effect of organic and inorganic nutrients sources on phenology and growth of wheat. Pak. J. Bot. 2015, 47, 2215–2222. [Google Scholar]
- Vergara, B.S.; Lilis, R.; Tanaka, A. Relationship between length of growing period and yield of rice plants under a limited nitrogen supply. Soil Sci. Plant Nutr. 1964, 10, 15–21. [Google Scholar] [CrossRef]
- Abacar, J.D.; Lin, Z.; Zhang, X.; Ding, C.; She, T.; Liu, Z.; Wang, S.; Ding, Y. Variation in yield and physicochemical quality traits among mutants of Japonica rice cultivar Wuyujing 3. Rice Sci. 2016, 23, 33–41. [Google Scholar] [CrossRef]
- Duan, L.; Huang, C.; Chen, G.; Xiong, L.; Liu, Q.; Yang, W. Determination of rice panicle numbers during heading by multi-angle imaging. Crop J. 2015, 3, 211–219. [Google Scholar] [CrossRef] [Green Version]
- Noori, Z.; Mujadidi, M.W.; Amin, M.W. Physicochemical properties and morphological observations of selected local rice varieties in northern Afghanistan. Int. J. Agric. Environ. Food Sci. 2018, 2, 99–103. [Google Scholar] [CrossRef]
- Kaur, P.; Pal, P.; Virdi, A.S.; Kaur, A.; Singh, N.; Mahajan, G. Protein and starch characteristics of milled rice from different cultivars affected by transplantation date. J. Food Sci. Technol. 2016, 53, 3186–3196. [Google Scholar] [CrossRef] [Green Version]
- Juliano, B.O.; Villareal, C.P. Grain Quality Evaluation of World Rices; International Rice Research Institute: Manila, Philippines, 1993. [Google Scholar]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Composition and functional properties of rice. Int. J. Food Sci. Technol. 2002, 37, 849–868. [Google Scholar] [CrossRef]
- Qiao, J.; Liu, Z.; Deng, S.; Ning, H.; Yang, X.; Lin, Z.; Li, G.; Wang, Q.; Wang, S.; Ding, Y. Occurrence of perfect and imperfect grains of six japonica rice cultivars as affected by nitrogen fertilization. Plant Soil 2011, 349, 191–202. [Google Scholar] [CrossRef]
- Noori, Z.; Kakar, K.; Fujii, T.; Ji, B. Growth and yield characteristics of upland rice cultivar NERICA-4 grown under paddy field condition. Int. J. Agron. Agric. Res. 2017, 10, 59–68. [Google Scholar]
- Xu, Z.; Chen, W.; Ma, D.; Lu, Y.; Zhou, S.; Liu, L. Correlations between rice grain shapes and main qualitative characteristics. Zuo Wu Xue Bao 2004, 30, 894–900. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Code | Origin | Descriptions | Status |
|---|---|---|---|
| K1 | DT84 | A traditional sticky rice with good quality in the north of Vietnam | Cultivar |
| K2 | Mutated DT84 | F2 (self-pollination from the mutated DT84 F1) | Mutant line |
| K3 | Q5 | A commercial rice cultivar with good quality in the north of Vietnam | Cultivar |
| K4 | Mutated Q5 | F2 (self-pollination from the mutated Q5 F1) | Mutant line |
| Code | Plant Length (cm) | Tiller Number per Hill | Days to Heading | Days to Maturity | Panicle Length (cm) |
|---|---|---|---|---|---|
| K1 | 113.4 ± 3.6 a | 12.1 ± 0.4 ab | 106.3 ± 0.6 b | 136.7 ± 1.1 b | 23.1 ± 0.5 c |
| K2 | 114.1 ± 5.6 a | 13.7 ± 0.4 a | 107.6 ± 0.7 b | 138.2 ± 0.7 b | 26.4 ± 0.3 b |
| K3 | 108.5 ± 3.1 a | 11.1 ± 1.1 b | 104.0 ± 2.6 b | 132.7 ± 2.3 c | 26.3 ± 0.2 b |
| K4 | 109.5 ± 2.2 a | 12.5 ± 1.3 ab | 122.3 ± 1.1 a | 151.0 ± 1.0 a | 29.2 ± 0.6 a |
| Code | Panicle Number per Hill | Spikelet Number per Panicle | Ripened Ratio (%) | 1000 Grain Weight (g) | 1000 Brown Grain Weight (g) | 1000 Husk Weight (g) | Grain Yield (t/ha) |
|---|---|---|---|---|---|---|---|
| K1 | 9.7 ± 0.4 ab | 127.7 ± 8.8 b | 83.4 ± 5.6 a | 22.5 ± 0.1 b | 19.2 ± 0.3 b | 3.3 ± 0.1 a | 7.7 ± 0.5 b |
| K2 | 10.6 ± 0.1 a | 142.0 ± 6.1 ab | 90.0 ± 1.5 a | 23.9 ± 0.4 a | 21.3 ± 0.2 a | 2.6 ± 0.2 a | 10.8 ± 0.2 a |
| K3 | 9.3 ± 0.9 b | 135.3 ± 5.2 ab | 86.2 ± 1.2 a | 22.0 ± 0.6 b | 18.0 ± 0.4 c | 4.0 ± 1.0 a | 7.9 ± 0.6 b |
| K4 | 11.1 ± 1.3 a | 148.6 ± 3.1 a | 87.3 ± 3.6 a | 24.1 ± 0.4 a | 21.1 ± 0.3 a | 3.0 ± 0.5 a | 11.6 ± 0.2 a |
| Code | Amylose (%) | Protein (%) | Lipid (%) | Taste Score | Grain Length (mm) | Grain Width (mm) | Grain Length/Width Ratio |
|---|---|---|---|---|---|---|---|
| K1 | 23.2 ± 0.1 a | 7.0 ± 0.4 a | 10.7 ± 0.6 a | 68.0 ± 0.0 b | 6.2 ± 0.3 b | 2.5 ± 0.1 a | 2.5 ± 0.0 b |
| K2 | 22.5 ± 0.1 b | 6.6 ± 0.4 a | 10.3 ± 0.6 a | 73.7 ± 3.0 a | 6.3 ± 0.1 b | 2.4 ± 0.1 a | 2.6 ± 0.1 b |
| K3 | 23.3 ± 0.0 a | 6.9 ± 0.1 a | 8.3 ± 0.6 b | 67.7 ± 1.1 b | 6.2 ± 0.1 b | 2.2 ± 0.2 a | 2.8 ± 0.4 ab |
| K4 | 21.4 ± 0.4 c | 6.6 ± 0.3 a | 7.7 ± 1.1 b | 70.0 ± 1.0 ab | 7.1 ± 0.4 a | 2.1 ± 0.0 a | 3.3 ± 0.2 a |
| TN | PL | PN | SN | RR | GW | GY | AC | PC | LC | TS | GL | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PL | 0.047 | |||||||||||
| PN | 0.726 ** | 0.437 | ||||||||||
| SN | 0.178 | 0.810 *** | 0.422 | |||||||||
| RR | 0.559 * | 0.306 | 0.328 | 0.280 | ||||||||
| GW | 0.658 * | 0.574 * | 0.779 ** | 0.554 | 0.500 | |||||||
| GY | 0.685 ** | 0.671 ** | 0.885 *** | 0.718 ** | 0.580 * | 0.890 *** | ||||||
| AC | −0.330 | −0.807 *** | −0.664 ** | −0.786 ** | −0.274 | −0.833 *** | −0.836 *** | |||||
| PC | −0.242 | −0.553 * | −0.320 | −0.373 | −0.018 | −0.476 | −0.381 | 0.534 | ||||
| LC | 0.322 | −0.734 ** | −0.004 | −0.433 | −0.101 | −0.027 | −0.170 | 0.393 | 0.232 | |||
| TS | 0.803 ** | 0.260 | 0.528 | 0.415 | 0.549 | 0.597 * | 0.641 * | −0.379 | −0.595 * | 0.180 | ||
| GL | −0.003 | 0.707 ** | 0.360 | 0.587 * | 0.115 | 0.520 | 0.497 | −0.723 ** | −0.294 | −0.693 ** | 0.013 | |
| GWD | 0.357 | −0.686 ** | −0.282 | −0.307 | −0.013 | −0.256 | −0.302 | 0.444 | 0.195 | 0.574 * | 0.298 | −0.478 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakar, K.; Xuan, T.D.; Quan, N.V.; Wafa, I.K.; Tran, H.-D.; Khanh, T.D.; Dat, T.D. Efficacy of N-methyl-N-nitrosourea (MNU) Mutation on Enhancing the Yield and Quality of Rice. Agriculture 2019, 9, 212. https://doi.org/10.3390/agriculture9100212
Kakar K, Xuan TD, Quan NV, Wafa IK, Tran H-D, Khanh TD, Dat TD. Efficacy of N-methyl-N-nitrosourea (MNU) Mutation on Enhancing the Yield and Quality of Rice. Agriculture. 2019; 9(10):212. https://doi.org/10.3390/agriculture9100212
Chicago/Turabian StyleKakar, Kifayatullah, Tran Dang Xuan, Nguyen Van Quan, Imran Khan Wafa, Hoang-Dung Tran, Tran Dang Khanh, and Tran Dang Dat. 2019. "Efficacy of N-methyl-N-nitrosourea (MNU) Mutation on Enhancing the Yield and Quality of Rice" Agriculture 9, no. 10: 212. https://doi.org/10.3390/agriculture9100212