

Rational Utilization of Sediment Resources Improves Rice Yield and Nitrogen Use Efficiency under Salt Stress
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Details
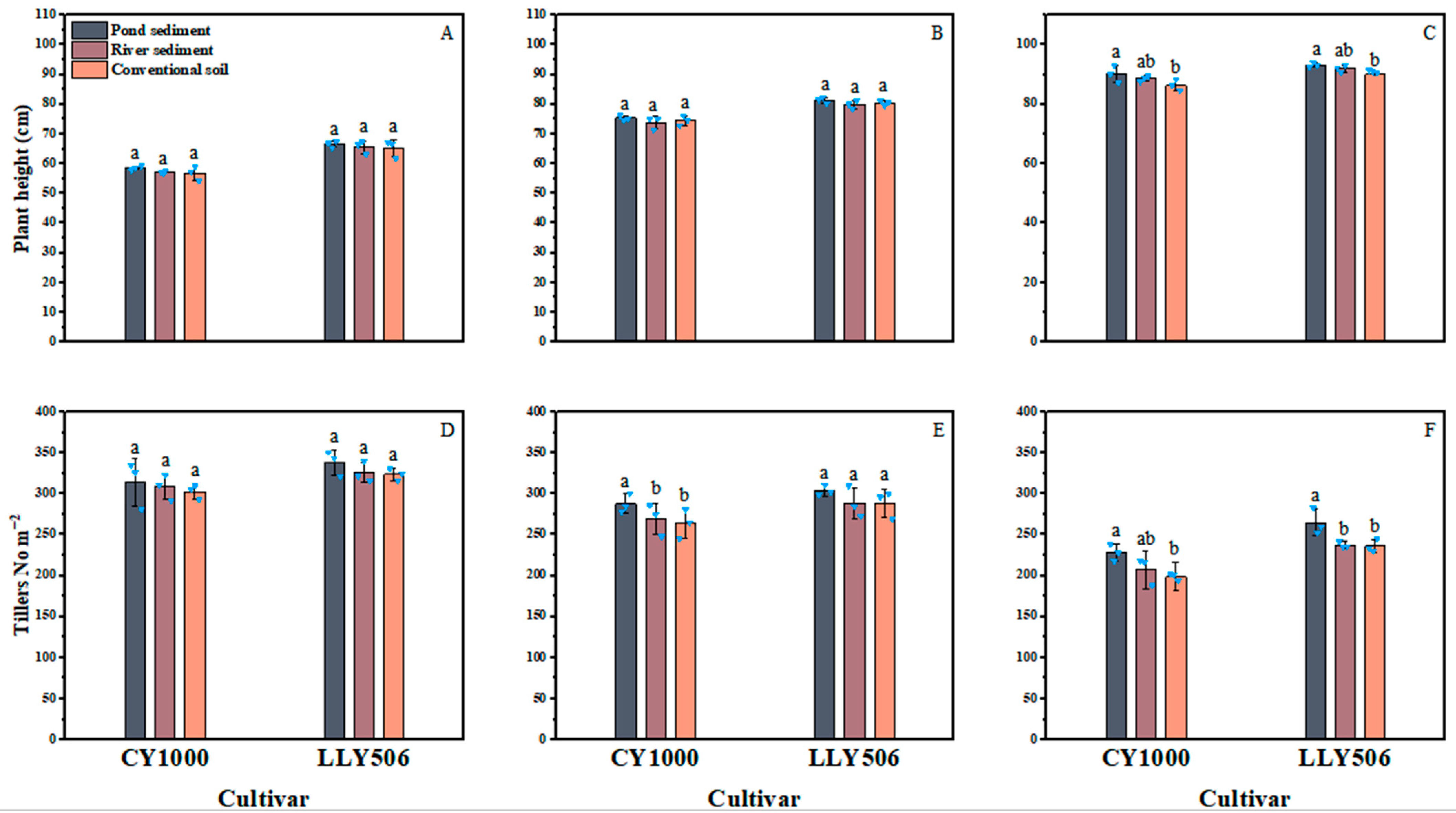
2.2. Agronomic Traits
2.3. Chlorophyll Contents and N-Metabolizing Enzyme Activities
2.4. Na+ and K+ Content, Total N Accumulation (TNA), and N Use Efficiency (NUE)
2.5. Yield and Related Attributes
2.6. Data Analysis
3. Results
3.1. Agronomic Traits
3.2. Yield and Yield Components
3.3. TNA and NUE
3.4. Chlorophyll a + b Contents and N-Metabolizing Enzyme Activities
3.5. Na+ and K+ Ions
3.6. Correlation Analysis of Yield Composition, Root Index, and Nitrogen Use Efficiency of Rice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Zhang, Z.; Tian, H.; Ashraf, U.; Hamoud, Y.A.; Alaa, A.A.; Tang, X.; Duan, M.; Wang, Z.; Pan, S. Nitrogen deep placement combined with straw mulch cultivation enhances physiological traits, grain yield and nitrogen use efficiency in mechanical pot-seedling transplanting rice. Rice Sci. 2022, 29, 89–100. [Google Scholar]
- Peng, S.; Tang, Q.; Zou, Y. Current status and challenges of rice production in China. Plant Prod. Sci. 2009, 12, 3–8. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Lin, Z.; Zhang, Z.; Tian, H.; Ashraf, U.; Alhaj Hamoud, Y.; Duan, M.; Tang, X.; Pan, S. Effects of nitrogen deep placement coupled with straw incorporation on grain quality and root traits from paddy fields. Crop Sci. 2021, 61, 3675–3686. [Google Scholar] [CrossRef]
- Guo, L.Y.; Lu, Y.Y.; Bao, S.Y.; Zhang, Q.; Geng, Y.Q.; Shao, X.W. Carbon and nitrogen metabolism in rice cultivars affected by salt-alkaline stress. Crop Pasture Sci. 2021, 72, 372–382. [Google Scholar] [CrossRef]
- Ashraf, M.; Shahzad, S.M.; Imtiaz, M.; Rizwan, M.S. Salinity effects on nitrogen metabolism in plants—Focusing on the activities of nitrogen metabolizing enzymes: A review. J. Plant Nutr. 2018, 41, 1065–1081. [Google Scholar] [CrossRef]
- Fu, X.; Ma, L.; Gui, R.; Ashraf, U.; Li, Y.; Yang, X.; Zhang, J.; Imran, M.; Tang, X.; Tian, H.; et al. Differential response of fragrant rice cultivars to salinity and hydrogen rich water in relation to growth and antioxidative defense mechanisms. Int. J. Phytoremediation 2021, 23, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Fatima, A.; Hussain, S.; Hussain, S.; Ali, B.; Ashraf, U.; Zulfiqar, U.; Aslam, Z.; Al-Robai, S.A.; Alzahrani, F.O.; Hano, C.; et al. Differential morphophysiological, biochemical, and molecular responses of maize hybrids to salinity and alkalinity stresses. Agronomy 2021, 11, 1150. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, Y.; Hussain, S.; Yang, S.; Li, R.; Liu, S. Study on the effect of salt stress on yield and grain quality among different rice varieties. Front. Plant Sci. 2022, 13, 918460. [Google Scholar] [CrossRef]
- Ganapati, R.K.; Naveed, S.A.; Zafar, S.; Wensheng, W.; Jianlong, X.U. Saline-Alkali tolerance in rice: Physiological response, molecular mechanism, and QTL identification and application to breeding. Rice Sci. 2022, 29, 412–434. [Google Scholar] [CrossRef]
- Hussain, S.; Mehmood, U.; Ashraf, U.; Naseer, M.A. Combined salinity and waterlogging stress in plants: Limitations and tolerance mechanisms. In Climate Change and Crop Stress; Academic Press: Cambridge, MA, USA, 2022; pp. 95–112. [Google Scholar]
- Dluzniewska, P.; Gessler, A.; Dietrich, H.; Schnitzler, J.P.; Rennenberg, H. Nitrogen uptake and metabolism in populus canescens as affected by salinity. New Phytol. 2007, 173, 279–293. [Google Scholar] [CrossRef]
- Zhu, G.; Wang, Y.; Shi, X.; Lu, H.; Ren, Z.; Shi, Y.; Jiao, X.; Ibrahim, M.E.H.; Irshad, A.; Zhu, W.; et al. Optimum nitrogen management enhances growth, antioxidant ability and yield performance of rice in saline soil of coastal area of China. Chil. J. Agric. Res. 2020, 80, 629–639. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Qin, C.; Begum, N.; Maodong, Q.; Dong, X.X.; El-Esawi, M. Nitrogen availability prevents oxidative effects of salinity on wheat growth and photosynthesis by up-regulating the antioxidants and osmolytes metabolism, and secondary metabolite accumulation. BMC Plant Biol. 2019, 19, 479. [Google Scholar] [CrossRef] [PubMed]
- Lyu, J.; Wang, X.; Hou, S.; Zeb, A.; Zhu, H.; Xu, Y. Content variation and potential runoff loss risk of nutrients in surface water of saline-alkali paddy in response to the application of different nitrogen fertilizer types. Sustainability 2023, 15, 7040. [Google Scholar] [CrossRef]
- Li, L.; Wu, T.; Li, Y.; Hu, X.; Wang, Z.; Liu, J.; Qin, W.; Ashraf, U. Deep fertilization improves rice productivity and reduces ammonia emissions from rice fields in China; a meta-analysis. Field Crops Res. 2022, 289, 108704. [Google Scholar] [CrossRef]
- Li, Y.; Gao, Y.; Ding, L.; Shen, Q.; Guo, S. Ammonium enhances the tolerance of rice seedlings (Oryza sativa L.) to drought condition. Agric. Water Manag. 2009, 96, 12. [Google Scholar] [CrossRef]
- Zhong, C.; Bai, Z.G.; Zhu, L.F.; Zhang, J.H.; Zhu, C.Q.; Huang, J.L.; Jin, Q.Y.; Cao, X.C. Nitrogen mediated alleviation of photosynthetic inhibition under moderate water deficit stress in rice (Oryza sativa L). Environ. Exp. Bot. 2019, 157, 269–282. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.S.; Guo, R.; Shi, D.C.; Liu, B.; Lin, X.Y.; Yang, C.W. Effects of salt stress on ion balance and nitrogen metabolism of old and young leaves in rice (Oryza sativa L.). BMC Plant Biol. 2012, 194, 1471–2229. [Google Scholar] [CrossRef]
- Qiao, C.; Duan, Y.; Zhang, M.; Hagemann, M.; Luo, Q.; Lu, X. Effects of reduced and enhanced glycogen pools on salt-induced sucrose production in a sucrose-secreting strain of Synechococcus elongatus PCC 7942. Appl. Environ. Microbiol. 2018, 84, e02023-17. [Google Scholar] [CrossRef]
- Yang, Z.; Yao, Y.; Sun, M.; Li, G.; Zhu, J. Metagenomics reveal microbial effects of lotus root–fish co-culture on nitrogen cycling in aquaculture pond sediments. Microorganisms 2022, 10, 1740. [Google Scholar] [CrossRef]
- Zuo, W.; Bai, Y.; Lv, M. Sustained effects of one-time sewage sludge addition on rice yield and heavy metals accumulation in salt-affected mudflat soil. Environ. Sci. Pollut. Res. 2021, 28, 7476–7490. [Google Scholar] [CrossRef]
- Shan, S.P.; Wei, Z.W.; Cheng, W.; Du, D.X.; Zheng, D.F.; Ma, G.H. Biofertilizer based on halotolerant microorganisms promotes the growth of rice plants and alleviates the effects of saline stress. Front. Microbiol. 2023, 14, 1165631. [Google Scholar] [CrossRef]
- Singh, R.P.; Agrawal, M. Effect of different sewage sludge applications on growth and yield of Vigina radiate L. Field crop: Metal uptake by plant. Ecol. Eng. 2010, 36, 969–972. [Google Scholar] [CrossRef]
- Wu, J.R.; Jin, X.C.; Zhang, L.J. Preliminary report on the test of river sludge return to field. Zhejiang Agric. Sci. 2018, 59, 1617–1618. [Google Scholar]
- Drozdz, D.; Malinska, K.; Mazurkiewicz, J.; Kacprzak, M.; Mrowiec, M.; Szczypiór, A.; Stachowiak, T. Fish pond sediment from aquaculture production-Current practices and the potential for nutrient recovery: A Review. Int. Agrophysics 2020, 34, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Pan, S.; Wen, X.; Wang, Z.; Ashraf, U.; Tian, H.; Duan, M.; Mo, Z.; Fan, P.; Tang, X. Benefits of mechanized deep placement of nitrogen fertilizer in direct-seeded rice in South China. Field Crops Res. 2017, 203, 139–149. [Google Scholar] [CrossRef]
- Li, Y.S.; Ai, Z.Y.; Mu, Y.X.; Zhao, T.C.; Zhang, Y.C.; Li, L.; Huang, Z.; Nie, L.X.; Khan, M.N. Rice yield penalty and quality deterioration is associated with failure of nitrogen uptake from regreening to panicle initiation stage under salinity. Front. Plant Sci. 2023, 14, 1120755. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, S.; Zhang, Y.; Li, T.; Ge, H.; Xia, S. Rice root morphological and physiological traits interaction with rhizosphere soil and its effect on methane emissions in paddy fields. Soil Biol. Biochem. 2019, 129, 191–200. [Google Scholar] [CrossRef]
- Derkx, A.P.; Orford, S.; Griffiths, S.; Foulkes, M.J.; Hawkesford, M.J. Identification of differentially senescing mutants of wheat and impacts on yield, biomass and nitrogen partitioning. J. Integr. Plant Biol. 2012, 54, 555–566. [Google Scholar] [CrossRef]
- Pandey, M.; Paladi, R.K.; Srivastava, A.K.; Suprasanna, P. Thiourea and hydrogen peroxide priming improved K+ retention and source-sink relationship for mitigating salt stress in rice. Sci. Rep. 2021, 11, 3000. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al, Y.R.; Yaish, M.W. The role of na+ and k+ transporters in salt stress adaptationin glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef]
- Shi, X.L.; Zhou, D.Y.; Guo, P.; Zhang, H.; Dong, J.L.; Ren, J.Y.; Jiang, C.J.; Zhong, C.; Zhao, X.H.; Yu, H.Q. External potassium mediates the response and tolerance to salt stress in peanut at the flowering and needling stages. Photosynthetica 2020, 58, 1141–1149. [Google Scholar] [CrossRef]
- Yan, F.; Wei, H.; Li, W.; Liu, Z.; Tang, S.; Chen, L.; Ding, C.; Jiang, Y.; Ding, Y.; Li, G. Melatonin improves K+ and Na+ homeostasis in rice under salt stress by mediated nitric oxide. Ecotoxicol. Environ. Saf. 2020, 206, 111358. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Yuan, Y.Z.; Ou, J.Q.; Lin, Q.H.; Zhang, C.F. Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat (Triticum aestivum) seedlings exposed to different salinity. J. Plant Physiol. 2007, 164, 695–701. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Fan, X.R.; Sun, S.B.; Xu, G.H.; Hu, J.; Shen, Q.R. Effect of nitrate on activities and transcript levels of nitrate reductase and glutamine synthetase in rice. Pedosphere 2008, 18, 664–673. [Google Scholar] [CrossRef]
- Gu, J.J.; Zhao, H.W.; Jia, Y.; Hu, B.W.; Wang, Z.Q.; Qu, Z.J. Effect of salt stress on nitrogen assimilation of functional leaves and root system of rice in cold region. J. Northeast Agric. Univ. 2020, 2, 9–16. [Google Scholar]
- Duan, Y.H.; Shi, X.J.; Li, S.L.; Sun, X.F.; He, X.H. Nitrogen Use Efficiency as Affected by Phosphorus and Potassium in Long-Term Rice and Wheat Experiments. J. Integr. Agric. 2014, 13, 588–596. [Google Scholar] [CrossRef]
- Phan, N.; Heymans, A.; Bonnave, M.; Lutts, S.; Pham, C.V.; Bertin, P. Nitrogen use efficiency of rice cultivars (Oryza sativa L.) under salt stress and low nitrogen conditions. J. Plant Growth Regul. 2023, 42, 1789–1803. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Location | pH | SOM g kg−1 | TN g kg−1 | AN mg kg−1 | AP mg kg−1 | AK mg kg−1 |
|---|---|---|---|---|---|---|---|
| Conventional soil | 109.14′ E 18.35′ N | 6.96 | 8.844 | 0.44 | 74.34 | 18.886 | 63.21 |
| River sediment | 108.90′ E 18.43′ N | 5.62 | 18.44 | 1.33 | 313.38 | 47.94 | 187.12 |
| Pond sediment | 108.82′ E 18.47′ N | 6.85 | 30.99 | 2.51 | 470.25 | 122.29 | 220.22 |
| Cultivar | Treatments | Productive Panicle (m−2) | Spikelets Per Panicle | Grain Filling (%) | 1000-grain Weight (g) | Grain Yield (t ha−1) |
|---|---|---|---|---|---|---|
| CY1000 | PS | 216.67 ± 11.81 a | 186.33 ± 9.07 a | 84.47 ± 0.81 a | 24.57 ± 0.87 a | 5.27 ± 0.25 a |
| RS | 197.92 ± 8.13 ab | 173.33 ± 11.06 a | 81.87 ± 0.40 a | 24.23 ± 1.83 a | 4.93 ± 0.25 a | |
| CS | 192.92 ± 10.48 b | 157.67 ± 6.43 b | 80.37 ± 1.79 a | 21.40 ± 0.66 b | 3.20 ± 0.26 b | |
| LLY506 | PS | 255.63 ± 12.91 a | 182.33 ± 5.13 a | 83.97 ± 3.80 a | 22.50 ± 0.70 a | 5.17 ± 0.31 a |
| RS | 233.33 ± 8.32 b | 158.33 ± 8.50 b | 77.07 ± 4.48 b | 20.87 ± 1.29 a | 4.80 ± 0.26 a | |
| CS | 231.88 ± 14.13 b | 135.00 ± 12.77 c | 82.6 a ± 3.48 ab | 18.67 ± 2.87 b | 3.60 ± 0.10 b | |
| Cultivars (C) | ** | * | ns | ** | ns | |
| Sediment Resources (S) | ** | ** | * | ** | * | |
| C × S | ns | ns | ns | ns | ns | |
| Cultivar | Treatments | TNA (kg ha−1) | NGPE (kg kg−1) | PFPN (kg kg−1) | NHI |
|---|---|---|---|---|---|
| CY1000 | PS | 147.94 ± 1.93 a | 34.25 ± 1.76 a | 28.15 ± 1.39 a | 0.49 ± 0.04 a |
| RS | 139.92 ± 7.34 a | 35.26 ± 0.31 a | 27.41 ± 1.40 a | 0.46 ± 0.02 ab | |
| CS | 112.42 ± 2.41 b | 28.50 ± 2.92 b | 17.78 ± 1.47 b | 0.41 ± 0.01 b | |
| LLY506 | PS | 141.92 ± 7.96 a | 36.52 ± 3.62 a | 28.70 ± 1.70 a | 0.50 ± 0.03 a |
| RS | 127.34 ± 5.23 b | 37.68 ± 0.68 a | 26.67 ± 1.47 a | 0.45 ± 0.05 b | |
| CS | 112.37 ± 7.83 c | 32.10 ± 1.47 b | 20.00 ± 0.56 b | 0.44 ± 0.05 b | |
| Cultivars (C) | ns | * | ns | ns | |
| Sediment Resources (S) | ** | ** | ** | * | |
| C × S | ns | ns | ns | ns | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, S.; He, A.; Guo, X.; Li, R.; Wang, J.; Qi, Y.; Zhang, P.; Tao, W.; Ji, J.; Zhao, T.; et al. Rational Utilization of Sediment Resources Improves Rice Yield and Nitrogen Use Efficiency under Salt Stress. Agriculture 2024, 14, 283. https://doi.org/10.3390/agriculture14020283
Cheng S, He A, Guo X, Li R, Wang J, Qi Y, Zhang P, Tao W, Ji J, Zhao T, et al. Rational Utilization of Sediment Resources Improves Rice Yield and Nitrogen Use Efficiency under Salt Stress. Agriculture. 2024; 14(2):283. https://doi.org/10.3390/agriculture14020283
Chicago/Turabian StyleCheng, Shenghai, Aibin He, Xiayu Guo, Rongyi Li, Jianbo Wang, Yucheng Qi, Pengfei Zhang, Weixu Tao, Junchao Ji, Tingcheng Zhao, and et al. 2024. "Rational Utilization of Sediment Resources Improves Rice Yield and Nitrogen Use Efficiency under Salt Stress" Agriculture 14, no. 2: 283. https://doi.org/10.3390/agriculture14020283