
Functional Characterization of RNA Silencing Suppressor Encoded by Cotton Leafroll Dwarf Virus

Abstract
:1. Introduction
2. Materials and Methods
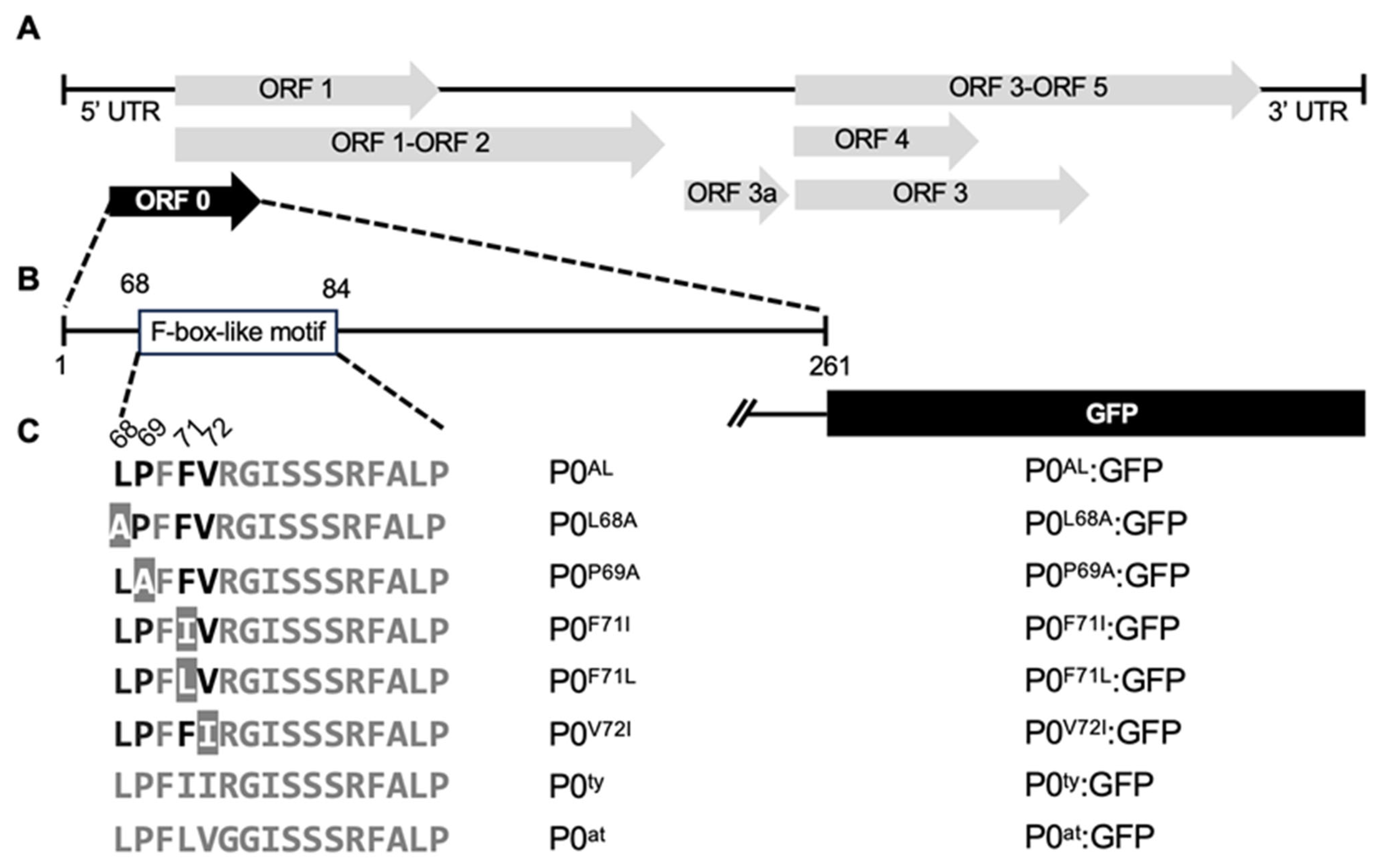
2.1. CLRDV P0 Constructs and Generation of P0AL Mutant Constructs
2.2. Generation of the Green Fluorescent Protein-Tagged Constructs
2.3. Agroinfiltration
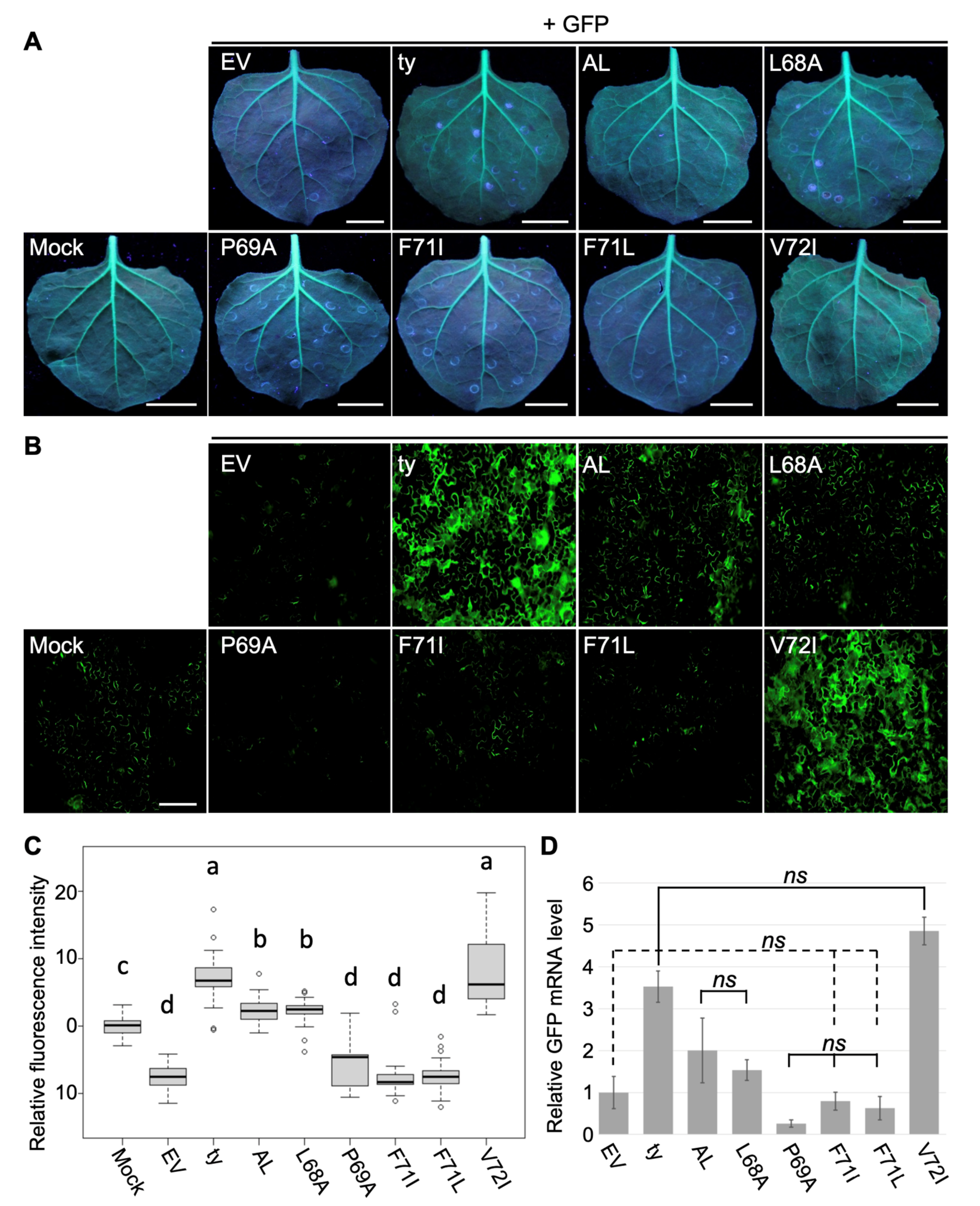
2.4. Examination of Fluorescence in Plants
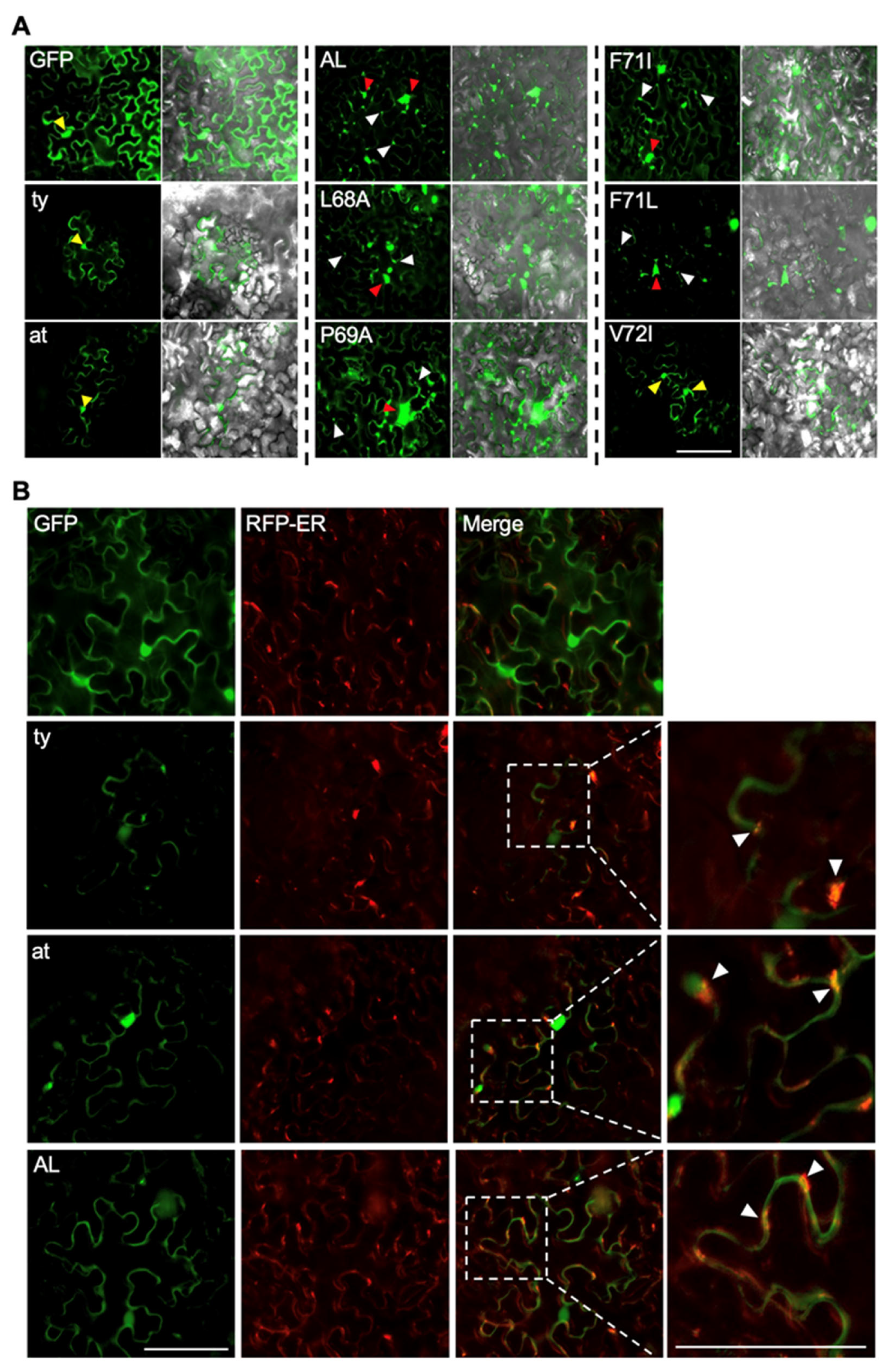
2.5. Intracellular Localization
2.6. Relative Expression Levels of GFP mRNA
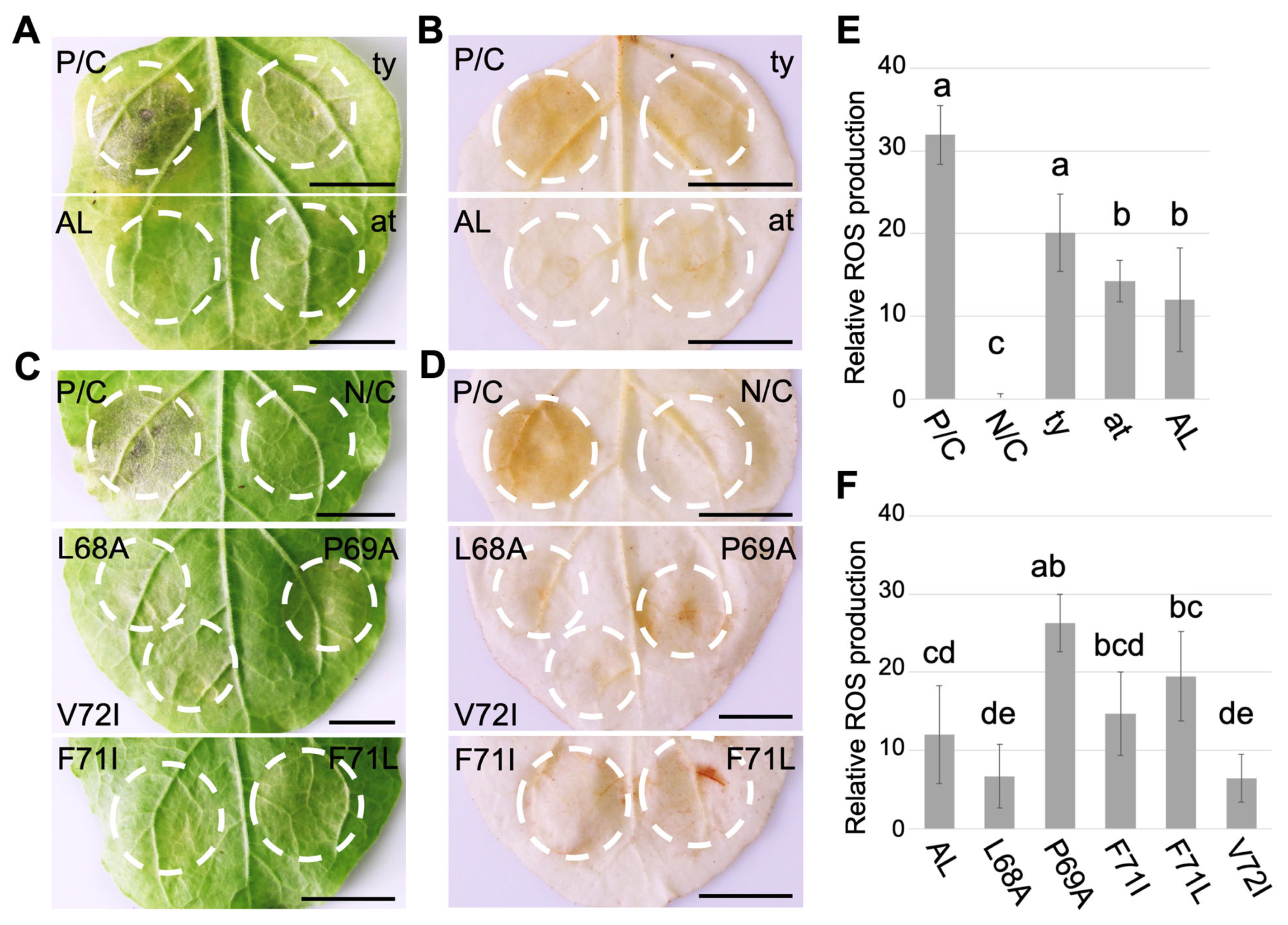
2.7. Histochemical Staining
3. Results
3.1. A Single Amino Acid Substitution Enhances RNA Silencing Suppression Potency of CLRDV P0AL
3.2. F-Box-Like Motif Modulates HR-Like Response Triggered by CLRDV P0
3.3. F-Box-Like Motif Plays a Role in the Intracellular Localization of CLRDV P0
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agrofoglio, Y.C.; Delfosse, V.C.; Casse, M.F.; Hopp, H.E.; Kresic, I.B.; Distéfano, A.J. Identification of a New Cotton Disease Caused by an Atypical Cotton Leafroll Dwarf Virus in Argentina. Phytopathology 2017, 107, 369–376. [Google Scholar] [CrossRef]
- Avelar, S.; Schrimsher, D.W.; Lawrence, K.S.; Brown, J.K. First Report of Cotton leafroll dwarf virus Associated with Cotton Blue Disease Symptoms in Alabama. Plant Dis. 2019, 103, 592. [Google Scholar] [CrossRef]
- Bag, S.; Roberts, P.M.; Kemerait, R.C. Cotton Leafroll Dwarf Disease: An Emerging Virus Disease on Cotton in the U.S. Crop. Soils 2021, 54, 18–22. [Google Scholar] [CrossRef]
- Distéfano, A.J.; Kresic, I.B.; Hopp, H.E. The complete genome sequence of a virus associated with cotton blue disease, cotton leafroll dwarf virus, confirms that it is a new member of the genus Polerovirus. Arch. Virol. 2010, 155, 1849–1854. [Google Scholar] [CrossRef]
- Edula, S.R.; Bag, S.; Milner, H.; Kumar, M.; Suassuna, N.D.; Chee, P.W.; Kemerait, R.C.; Hand, L.C.; Snider, J.L.; Srinivasan, R.; et al. Cotton leafroll dwarf disease: An enigmatic viral disease in cotton. Mol. Plant Pathol. 2023, 24, 513–526. [Google Scholar] [CrossRef]
- Delfosse, V.C.; Barón, M.P.B.; Distéfano, A.J. What we know about poleroviruses: Advances in understanding the functions of polerovirus proteins. Plant Pathol. 2021, 70, 1047–1061. [Google Scholar] [CrossRef]
- Avelar, S.; Ramos-Sobrinho, R.; Conner, K.; Nichols, R.L.; Lawrence, K.S.; Brown, J.K. Characterization of the Complete Genome and P0 Protein for a Previously Unreported Genotype of Cotton Leafroll Dwarf Virus, an Introduced Polerovirus in the United States. Plant Dis. 2020, 104, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, R.L.; Silva, T.F.; Simões-Araújo, J.L.; Barroso, P.A.V.; Vidal, M.S.; Vaslin, M.F.S. Molecular characterization of a virus from the family Luteoviridae associated with cotton blue disease. Arch. Virol. 2005, 150, 1357–1367. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Adams, M.J.; Carstens, E.B.; Lefkowitz, E.J. (Eds.) Family-Luteoviridae. In Virus Taxonomy; Elsevier: San Diego, CA, USA, 2012; pp. 1045–1053. [Google Scholar] [CrossRef]
- Smirnova, E.; Firth, A.E.; Miller, W.A.; Scheidecker, D.; Brault, V.; Reinbold, C.; Rakotondrafara, A.M.; Chung, B.Y.-W.; Ziegler-Graff, V. Discovery of a Small Non-AUG-Initiated ORF in Poleroviruses and Luteoviruses That Is Required for Long-Distance Movement. PLoS Pathog. 2015, 11, e1004868. [Google Scholar] [CrossRef] [PubMed]
- Sõmera, M.; Fargette, D.; Hébrard, E.; Sarmiento, C. ICTV virus taxonomy profile: Solemoviridae 2021. J. Gen. Virol. 2021, 102, 001707. [Google Scholar] [CrossRef]
- Agrofoglio, Y.C.; Delfosse, V.C.; Casse, M.F.; Hopp, H.E.; Kresic, I.B.; Ziegler-Graff, V.; Distéfano, A.J. P0 protein of cotton leafroll dwarf virus-atypical isolate is a weak RNA silencing suppressor and the avirulence determinant that breaks the cotton Cbd gene-based resistance. Plant Pathol. 2019, 68, 1059–1071. [Google Scholar] [CrossRef]
- Almasi, R.; Miller, W.A.; Ziegler-Graff, V. Mild and severe cereal yellow dwarf viruses differ in silencing suppressor efficiency of the P0 protein. Virus Res. 2015, 208, 199–206. [Google Scholar] [CrossRef]
- Delfosse, V.C.; Agrofoglio, Y.C.; Casse, M.F.; Kresic, I.B.; Hopp, H.E.; Ziegler-Graff, V.; Distéfano, A.J. The P0 protein encoded by cotton leafroll dwarf virus (CLRDV) inhibits local but not systemic RNA silencing. Virus Res. 2014, 180, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Zhuo, T.; Zhao, T.Y.; Zhou, C.J.; Li, Y.Y.; Wang, Y.; Li, D.W.; Yu, J.L.; Han, C.G. Functional Characterization of RNA Silencing Suppressor P0 from Pea Mild Chlorosis Virus. Int. J. Mol. Sci. 2020, 21, 7136. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-D.; Dughbaj, M.A.; Nguyen, T.T.V.; Nguyen, T.Q.Y.; Oza, S.; Valdez, K.; Anda, P.; Waltz, J.; Sacco, M.A. Systematic mutagenesis of Polerovirus protein P0 reveals distinct and overlapping amino acid functions in Nicotiana glutinosa. Virology 2023, 578, 24–34. [Google Scholar] [CrossRef]
- Wang, L.; Tian, P.; Yang, X.; Zhou, X.; Zhang, S.; Li, C.; Yang, X.; Liu, Y. Key Amino Acids for Pepper Vein Yellows Virus P0 Protein Pathogenicity, Gene Silencing, and Subcellular Localization. Front. Microbiol. 2021, 12, 1653. [Google Scholar] [CrossRef] [PubMed]
- Hammond, S.M. Dicing and slicing: The core machinery of the RNA interference pathway. FEBS Lett. 2005, 579, 5822–5829. [Google Scholar] [CrossRef]
- Li, F.; Wang, A. RNA-Targeted Antiviral Immunity: More Than Just RNA Silencing. Trends Microbiol. 2019, 27, 792–805. [Google Scholar] [CrossRef]
- Voinnet, O. Induction and suppression of RNA silencing: Insights from viral infections. Nat. Rev. Genet. 2005, 6, 206–220. [Google Scholar] [CrossRef]
- Blevins, T.; Rajeswaran, R.; Shivaprasad, P.V.; Beknazariants, D.; Si-Ammour, A.; Park, H.-S.; Vazquez, F.; Robertson, D.; Meins, F.; Hohn, T.; et al. Four plant Dicers mediate viral small RNA biogenesis and DNA virus induced silencing. Nucleic Acids Res. 2006, 34, 6233–6246. [Google Scholar] [CrossRef]
- Derrien, B.; Clavel, M.; Baumberger, N.; Iki, T.; Sarazin, A.; Hacquard, T.; Ponce, M.R.; Ziegler-Graff, V.; Vaucheret, H.; Micol, J.L.; et al. A Suppressor Screen for AGO1 Degradation by the Viral F-Box P0 Protein Uncovers a Role for AGO DUF1785 in sRNA Duplex Unwinding. Plant Cell 2018, 30, 1353–1374. [Google Scholar] [CrossRef]
- Pazhouhandeh, M.; Dieterle, M.; Marrocco, K.; Lechner, E.; Berry, B.; Brault, V.; Hemmer, O.; Kretsch, T.; Richards, K.E.; Genschik, P.; et al. F-box-like domain in the polerovirus protein P0 is required for silencing suppressor function. Proc. Natl. Acad. Sci. USA 2006, 103, 1994–1999. [Google Scholar] [CrossRef]
- Akinyuwa, M.F.; Price, B.K.; Martin, K.M.; Kang, S.-H. A newly isolated cotton-infecting Polerovirus with cryptic pathogenicity encodes a weak suppressor of RNA silencing. Front. Agron. 2023, 5, 123516. [Google Scholar] [CrossRef]
- Bortolamiol-Bécet, D.; Monsion, B.; Chapuis, S.; Hleibieh, K.; Scheidecker, D.; Alioua, A.; Bogaert, F.; Revers, F.; Brault, V.; Ziegler-Graff, V. Phloem-Triggered Virus-Induced Gene Silencing Using a Recombinant Polerovirus. Front. Microbiol. 2018, 9, 2449. [Google Scholar] [CrossRef]
- Fusaro, A.F.; Correa, R.L.; Nakasugi, K.; Jackson, C.; Kawchuk, L.; Vaslin, M.F.S.; Waterhouse, P.M. The Enamovirus P0 protein is a silencing suppressor which inhibits local and systemic RNA silencing through AGO1 degradation. Virology 2012, 426, 178–187. [Google Scholar] [CrossRef]
- Kozlowska-Makulska, A.; Guilley, H.; Szyndel, M.S.; Beuve, M.; Lemaire, O.; Herrbach, E.; Bouzoubaa, S. P0 proteins of European beet-infecting poleroviruses display variable RNA silencing suppression activity. J. Gen. Virol. 2010, 91, 1082–1091. [Google Scholar] [CrossRef]
- Li, Y.; Sun, Q.; Zhao, T.; Xiang, H.; Zhang, X.; Wu, Z.; Zhou, C.; Zhang, X.; Wang, Y.; Zhang, Y.; et al. Interaction between Brassica yellows virus silencing suppressor P0 and plant SKP1 facilitates stability of P0 in vivo against degradation by proteasome and autophagy pathways. New Phytol. 2019, 222, 1458–1473. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Zhao, X.; Dong, Q.; Zhou, B.L.; Gao, Z.L. Characterization of an RNA silencing suppressor encoded by maize yellow dwarf virus-RMV2. Virus Genes 2018, 54, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Liang, K.-L.; Liu, J.-Y.; Bao, Y.-Y.; Wang, Z.-Y.; Xu, X.-B. Screening and Identification of Host Factors Interacting with the Virulence Factor P0 Encoded by Sugarcane Yellow Leaf Virus by Yeast Two-Hybrid Assay. Genes 2023, 14, 1397. [Google Scholar] [CrossRef]
- Rashid, M.-O.; Zhang, X.-Y.; Wang, Y.; Li, D.-W.; Yu, J.-L.; Han, C.-G. The Three Essential Motifs in P0 for Suppression of RNA Silencing Activity of Potato leafroll virus Are Required for Virus Systemic Infection. Viruses 2019, 11, 170. [Google Scholar] [CrossRef]
- Wang, K.; Empleo, R.; Nguyen, T.T.V.; Moffett, P.; Sacco, M.A. Elicitation of hypersensitive responses in Nicotiana glutinosa by the suppressor of RNA silencing protein P0 from poleroviruses. Mol. Plant Pathol. 2016, 16, 435–448. [Google Scholar] [CrossRef]
- Baumberger, N.; Tsai, C.-H.; Lie, M.; Havecker, E.; Baulcombe, D.C. The Polerovirus Silencing Suppressor P0 Targets ARGONAUTE Proteins for Degradation. Curr. Biol. 2007, 17, 1609–1614. [Google Scholar] [CrossRef]
- Cascardo, R.-. S, Arantes I.-L, Silva T.-F, Sachetto-Martins G, Vaslin M.-F, Corrêa R.-L. Function and diversity of P0 proteins among cotton leafroll dwarf virus isolates. Virol J. 2015, 12, 123. [Google Scholar] [CrossRef]
- LaTourrette, K.; Holste, N.M.; Garcia-Ruiz, H. Polerovirus genomic variation. Virus Evol. 2021, 7, veab102. [Google Scholar] [CrossRef]
- Bortolamiol, D.; Pazhouhandeh, M.; Marrocco, K.; Genschik, P.; Ziegler-Graff, V. The Polerovirus F Box Protein P0 Targets ARGONAUTE1 to Suppress RNA Silencing. Curr. Biol. 2007, 17, 1615–1621. [Google Scholar] [CrossRef]
- Correa, R.L.; Bruckner, F.P.; Cascardo, R.D.S.; Alfenas-Zerbini, P. The Role of F-Box Proteins during Viral Infection. Int. J. Mol. Sci. 2013, 14, 4030–4049. [Google Scholar] [CrossRef] [PubMed]
- Derrien, B.; Baumberger, N.; Schepetilnikov, M.; Viotti, C.; De Cillia, J.; Ziegler-Graff, V.; Isono, E.; Schumacher, K.; Genschik, P. Degradation of the antiviral component ARGONAUTE1 by the autophagy pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 15942–15946. [Google Scholar] [CrossRef]
- Sun, Q.; Li, Y.-Y.; Wang, Y.; Zhao, H.-H.; Zhao, T.-Y.; Zhang, Z.-Y.; Li, D.-W.; Yu, J.-L.; Wang, X.-B.; Zhang, Y.-L.; et al. Brassica yellows virus P0 protein impairs the antiviral activity of NbRAF2 in Nicotiana benthamiana. J. Exp. Bot. 2018, 69, 3127–3139. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.K.F.; Romanel, E.; Silva, T.D.F.; Castilhos, Y.; Schrago, C.G.; Galbieri, R.; Bélot, J.-L.; Vaslin, M.F.S. Complete genome sequences of two new virus isolates associated with cotton blue disease resistance breaking in Brazil. Arch. Virol. 2015, 160, 1371–1374. [Google Scholar] [CrossRef]
- Galbieri, R.; Boldt, A.S.; Scoz, L.B.; Rodrigues, S.M.; Rabel, D.O.; Belot, J.L.; Vaslin, M.; Silva, T.d.F.; Kobayasti, L.; Chitarra, L.G. Cotton blue disease in central-west Brazil: Occurrence, vector (Aphis gossypii) control levels and cultivar reaction. Trop. Plant Pathol. 2017, 42, 468–474. [Google Scholar] [CrossRef]
- Zhuo, T.; Li, Y.-Y.; Xiang, H.-Y.; Wu, Z.-Y.; Wang, X.-B.; Wang, Y.; Zhang, Y.-L.; Li, D.-W.; Yu, J.-L.; Han, C.-G. Amino Acid Sequence Motifs Essential for P0-Mediated Suppression of RNA Silencing in an Isolate of Potato leafroll virus from Inner Mongolia. Mol. Plant-Microbe Interact. 2014, 27, 515–527. [Google Scholar] [CrossRef]
- Lin, J.; Guo, J.; Finer, J.; Dorrance, A.E.; Redinbaugh, M.G.; Qu, F. The Bean Pod Mottle Virus RNA2-Encoded 58-Kilodalton Protein P58 Is Required in cis for RNA2 Accumulation. J. Virol. 2014, 88, 3213–3222. [Google Scholar] [CrossRef]
- Luo, C.; Wang, Z.Q.; Liu, X.; Zhao, L.; Zhou, X.; Xie, Y. Identification and Analysis of Potential Genes Regulated by an Alphasatellite (TYLCCNA) that Contribute to Host Resistance against Tomato Yellow Leaf Curl China Virus and Its Betasatellite (TYLCCNV/TYLCCNB) Infection in Nicotiana benthamiana. Viruses 2019, 11, 442. [Google Scholar] [CrossRef]
- Kang, S.-H.; Bak, A.; Kim, O.-K.; Folimonova, S.Y. Membrane association of a nonconserved viral protein confers virus ability to extend its host range. Virology 2015, 482, 208–217. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kang, S.-H.; Sun, Y.-D.; Atallah, O.O.; Huguet-Tapia, J.C.; Noble, J.D.; Folimonova, S.Y. A Long Non-Coding RNA of Citrus tristeza virus: Role in the Virus Interplay with the Host Immunity. Viruses 2019, 11, 436. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.T.; Voinnet, O.; Baulcombe, D.C. Initiation and Maintenance of Virus-Induced Gene Silencing. Plant Cell 1998, 10, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Mangwende, T.; Wang, M.-L.; Borth, W.; Hu, J.; Moore, P.H.; Mirkov, T.E.; Albert, H.H. The P0 gene of Sugarcane yellow leaf virus encodes an RNA silencing suppressor with unique activities. Virology 2009, 384, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Y.; Li, Y.-Y.; Wang, Y.; Li, D.-W.; Yu, J.-L.; Han, C.-G. Comparative Analysis of Biological Characteristics among P0 Proteins from Different Brassica Yellows Virus Genotypes. Biology 2021, 10, 1076. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-H.; Qu, F.; Morris, T.J. A spectrum of HRT-dependent hypersensitive responses elicited by the 52 amino acid N-terminus of turnip crinkle virus capsid protein and its mutants. Virus Res. 2015, 200, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Michaeli, S.; Clavel, M.; Lechner, E.; Viotti, C.; Wu, J.; Dubois, M.; Hacquard, T.; Derrien, B.; Izquierdo, E.; Lecorbeiller, M.; et al. The viral F-box protein P0 induces an ER-derived autophagy degradation pathway for the clearance of membrane-bound AGO1. Proc. Natl. Acad. Sci. USA 2019, 116, 22872–22883. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-F.; Sun, R.; Guo, Q.; Zhang, S.; Meulia, T.; Halfmann, R.; Li, D.; Qu, F. A self-perpetuating repressive state of a viral replication protein blocks superinfection by the same virus. PLoS Pathog. 2017, 13, e1006253. [Google Scholar] [CrossRef] [PubMed]
- Kumar, T.K.S.; Samuel, D.; Jayaraman, G.; Srimathi, T.; Yu, C. The role of proline in the prevention of aggregation during protein folding in vitro. IUBMB Life 1998, 46, 509–517. [Google Scholar] [CrossRef]
- Pemberton, T.A.; Still, B.R.; Christensen, E.M.; Singh, H.; Srivastava, D.; Tanner, J.J. Proline: Mother Nature’s cryoprotectant applied to protein crystallography. Acta Crystallogr. Sect. D Struct. Biol. 2012, 68, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Sobrinho, R.; Adegbola, R.O.; Lawrence, K.; Schrimsher, D.W.; Isakeit, T.; Alabi, O.J.; Brown, J.K. Cotton Leafroll Dwarf Virus US Genomes Comprise Divergent Subpopulations and Harbor Extensive Variability. Viruses 2021, 13, 2230. [Google Scholar] [CrossRef]
- Tabassum, A.; Bag, S.; Suassuna, N.D.; Conner, K.N.; Chee, P.; Kemerait, R.C.; Roberts, P. Genome analysis of cotton leafroll dwarf virus reveals variability in the silencing suppressor protein, genotypes and genomic recombinants in the USA. PLoS ONE 2021, 16, E0252523. [Google Scholar] [CrossRef]
- Csorba, T.; Lózsa, R.; Hutvágner, G.; Burgyán, J. Polerovirus protein P0 prevents the assembly of small RNA-containing RISC complexes and leads to degradation of ARGONAUTE1. Plant J. 2010, 62, 463–472. [Google Scholar] [CrossRef]
- Li, S.; Le, B.; Ma, X.; Li, S.; You, C.; Yu, Y.; Zhang, B.; Liu, L.; Gao, L.; Shi, T.; et al. Biogenesis of phased siRNAs on membrane-bound polysomes in Arabidopsis. eLife 2016, 5, e22750. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′ to 3′) | Tm * | References |
|---|---|---|---|
| Q-5 site-directed mutagenesis | |||
| pAI-P0L68A:FW | TCTTTTTCTCGCTCCATTCTTCG | 60.9 | This study, Section 2.2 and Section 2.3 |
| pAI-P0L68A:RV | AGAGAACGAAGGAGAAAAGA | 54.3 | |
| pAI-P0P69A:FW | TTTTCTCCTTGCATTCTTCGTTA | 57.6 | |
| pAI-P0P69A:RV | AGAAGAGAACGAAGGAGAAA | 54.3 | |
| pAI-P0V72I:FW | TCCATTCTTCATTAGGGGAATTT | 57.6 | |
| pAI-P0V72I:RV | AGGAGAAAAAGAAGAGAACG | 54.3 | |
| Non-tagged P0s | |||
| P0AL.FW.ApaI | ACTAGGGCCCAACAATGTTGAATTTGATCATCTGC | 73.1 | This study, Section 2.2 |
| P0AL.RV.XbaI | GGACTCTAGATCAACTGCTTTCTCCTTCAC | 70.8 | |
| P0at.FW.ApaI | ACTAGGGCCCAACAATGTTGAACTTGATTATCTGC | 73.1 | |
| GFP-tagged P0s | |||
| P0:GFP-FW | TATGTGAAGGAGAAAGCAGTATGGCTAGCAAAGGAGAAGA | 76.0 | This study, Section 2.3 |
| P0:GFP-RV | TCTTCTCCTTTGCTAGCCATACTGCTTTCTCCTTCACATA | 76.0 | |
| GFP.RV.XbaI | GTACTCTAGACTATTTGTAGAGCTCATCC | 67.4 | |
| Sequence verification | |||
| pAI SEQ FW | CCTCGAGAATTCTCAACACAAC | 60.1 | [24] |
| pAI SEQ RV | GCTCAACACATGAGCGAAACCC | 64.2 | |
| Quantitative mRNA analysis | |||
| MFA.Gq-PCR:FW | GATGACGGGAACTACAAGAC | 58.4 | [24] |
| MFA.Gq-PCR:RV | CGAGTACAACTATAACTCACAC | 58.4 | |
| NbACTIN2-FW | CAATCCAGACACTGTACTTTCTCTC | 64.1 | [44] |
| NbACTIN2-RV | AAGCTGCAGGTATCCATGAGACTA | 63.6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akinyuwa, M.F.; Kang, S.-H. Functional Characterization of RNA Silencing Suppressor Encoded by Cotton Leafroll Dwarf Virus. Agriculture 2024, 14, 194. https://doi.org/10.3390/agriculture14020194
Akinyuwa MF, Kang S-H. Functional Characterization of RNA Silencing Suppressor Encoded by Cotton Leafroll Dwarf Virus. Agriculture. 2024; 14(2):194. https://doi.org/10.3390/agriculture14020194
Chicago/Turabian StyleAkinyuwa, Mary F., and Sung-Hwan Kang. 2024. "Functional Characterization of RNA Silencing Suppressor Encoded by Cotton Leafroll Dwarf Virus" Agriculture 14, no. 2: 194. https://doi.org/10.3390/agriculture14020194