
Diversity and Traits of Multiple Biotic Stressors Elicit Differential Defense Responses in Legumes
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Systems and Experimental Conditions
2.2. Experimental Setup
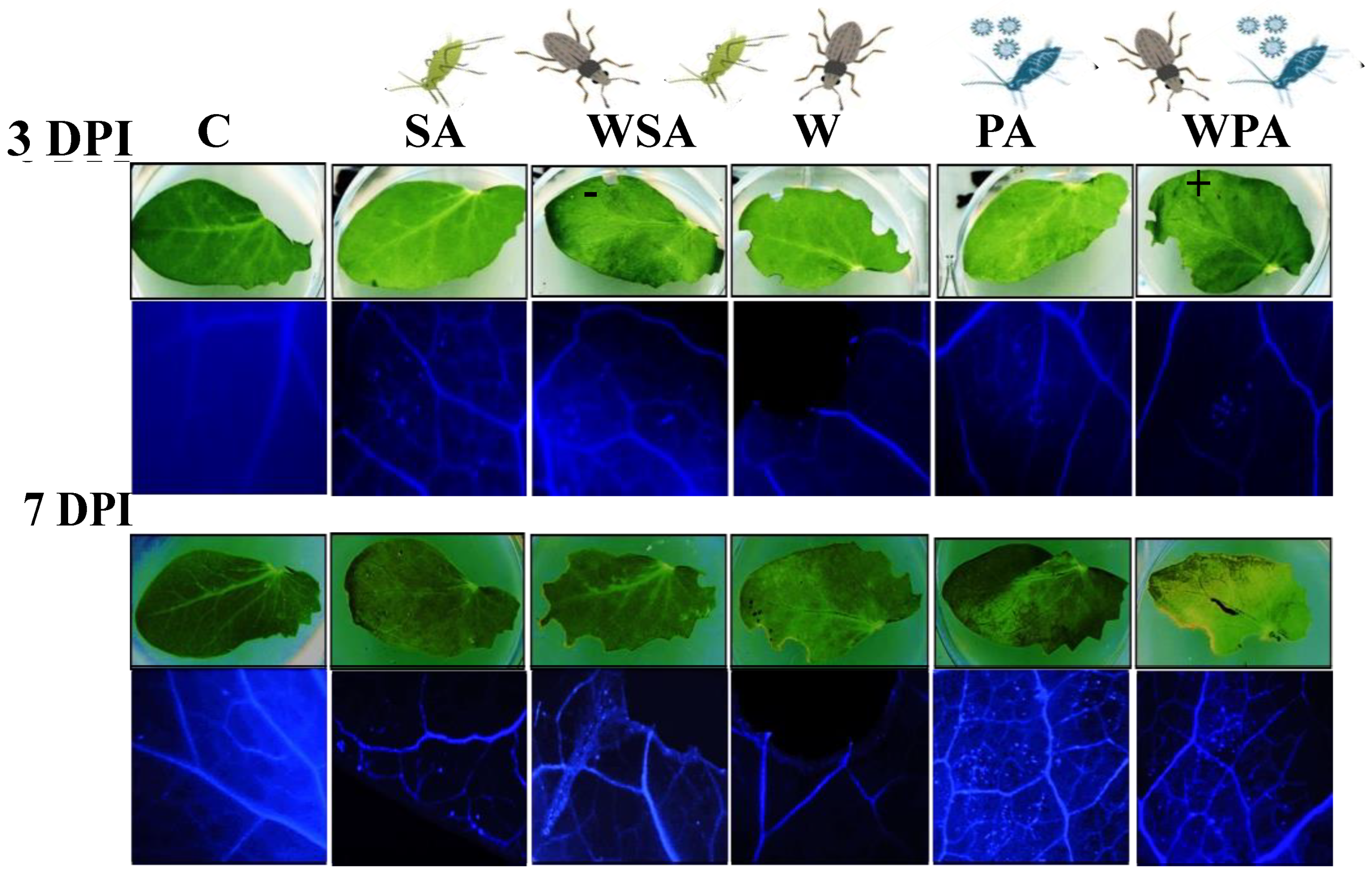
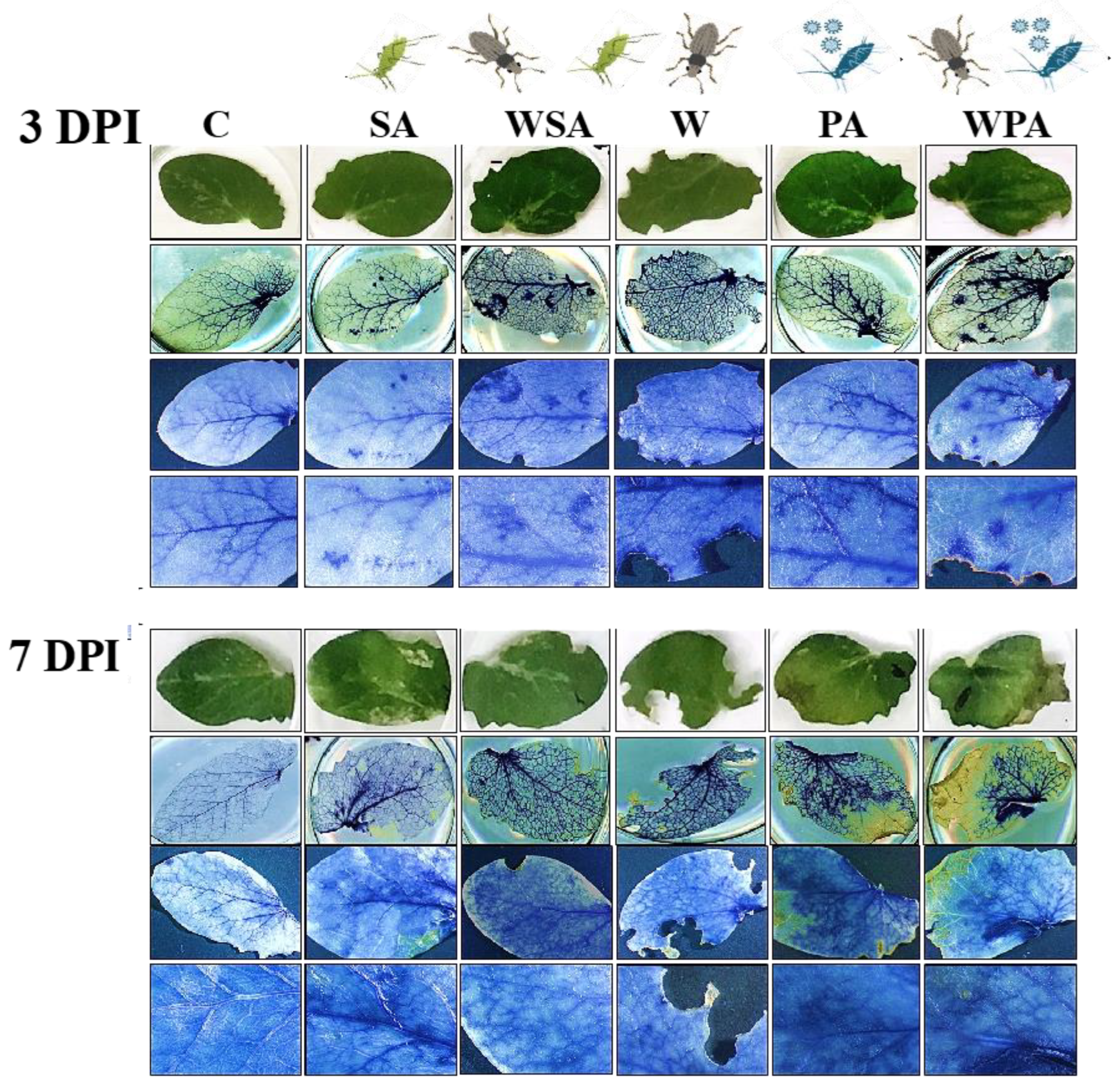
2.3. Detection of Callose Deposition
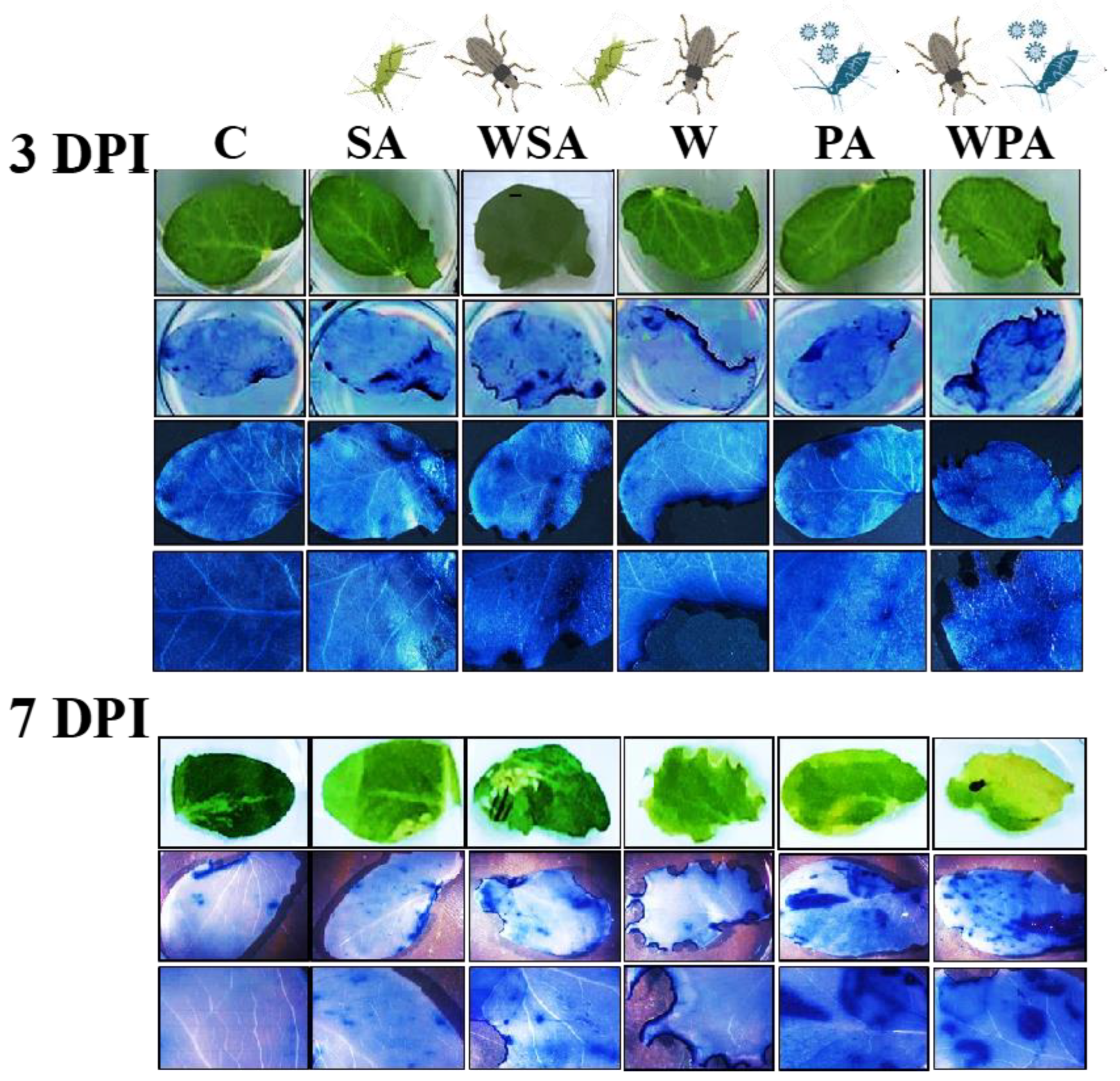
2.4. Detection of Reactive Oxygen Species
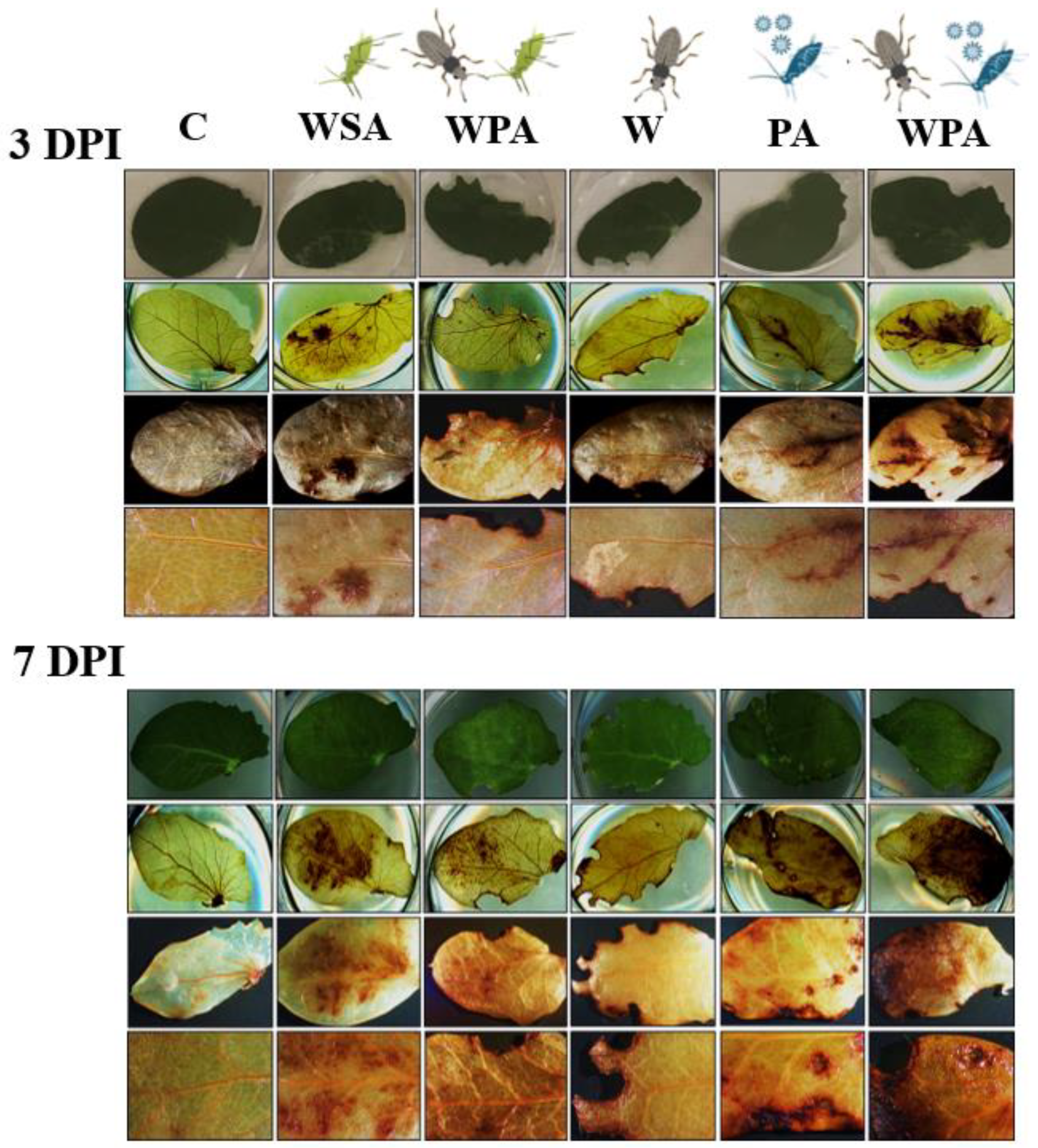
2.5. Detection of Cell Death in Peas
2.6. High-Throughput Phenotyping
3. Results
3.1. Differential Callose and Reactive Oxygen Species Responses of Pea Plants to Biotic Stressors
3.2. Effects of Biotic Stressors on Cell Death in Pea Plants
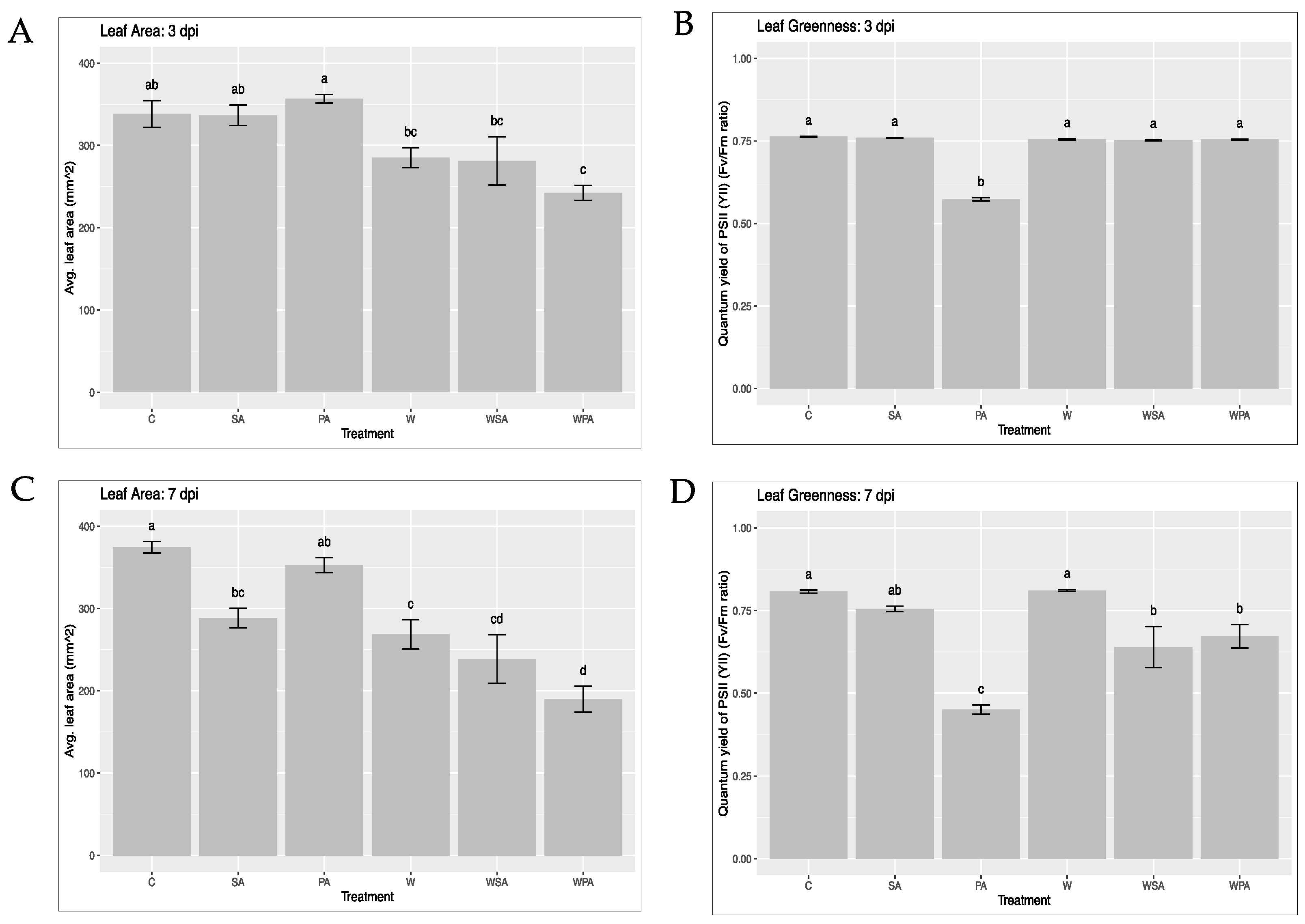
3.3. High-Throughput Phenotyping for Measuring Leaf Area and Chlorophyll Fluorescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pandey, P.; Ramegowda, V.; Senthil-Kumar, M. Shared and unique responses of plants to multiple individual stresses and stress combinations: Physiological and molecular mechanisms. Front. Plant Sci. 2015, 6, 723. [Google Scholar] [CrossRef]
- Miller, R.N.G.; Costa Alves, G.S.; Van Sluys, M.A. Plant immunity: Unravelling the complexity of plant responses to biotic stresses. Ann. Bot. 2017, 119, 681–687. [Google Scholar] [CrossRef]
- Basu, S.; Kushwaha, N.K.; Singh, A.K.; Sahu, P.P.; Kumar, R.V.; Chakraborty, S. Dynamics of a geminivirus-encoded pre-coat protein and host RNA-dependent RNA polymerase 1 in regulating symptom recovery in tobacco. J. Exp. Bot. 2018, 69, 2085–2102. [Google Scholar] [CrossRef]
- Basu, S.; Varsani, S.; Louis, J. Altering plant defenses: Herbivore-associated molecular patterns and effector arsenal of chewing herbivores. Mol. Plant-Microbe Interact. 2018, 31, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Nejat, N.; Mantri, N. Plant immune system: Crosstalk between responses to biotic and abiotic stresses the missing link in understanding plant defence. Curr. Issues Mol. Biol. 2017, 23, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Clark, R.E.; Bera, S.; Casteel, C.L.; Crowder, D.W. Responses of pea plants to multiple antagonists are mediated by order of attack and phytohormone crosstalk. Mol. Ecol. 2021, 30, 4939–4948. [Google Scholar] [CrossRef]
- Koornneef, A.; Pieterse, C.M.J. Cross talk in defense signaling. Plant Physiol. 2008, 146, 839–844. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signaling. Nat. Plants 2021, 7, 403–412. [Google Scholar] [CrossRef]
- Wang, Y.; Li, X.; Fan, B.; Zhu, C.; Chen, Z. Regulation and function of defense-related callose deposition in plants. Int. J. Mol. Sci. 2021, 22, 2393. [Google Scholar] [CrossRef]
- Wang, B.; Andargie, M.; Fang, R. The function and biosynthesis of callose in high plants. Heliyon 2022, 8, e09248. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Liu, C.; Wang, Y.; Chen, R.; Tang, M.; Du, B.; Zhu, L.; He, G. Herbivore-induced callose deposition on the sieve plates of rice: An important mechanism for host resistance. Plant Physiol. 2008, 146, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Reyes-Caldas, P.; Mann, M.; Seifbarghi, S.; Kahn, A.; Almeida, R.P.P.; Béven, L.; Heck, M.; Hogenhout, S.A.; Coaker, G. Bacterial vector-borne plant diseases: Unanswered questions and future directions. Mol. Plant 2020, 13, 1379–1393. [Google Scholar] [CrossRef] [PubMed]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.; Schwarze, J.; Sherwood, O.L.; Jnaid, Y.; McCabe, P.F.; Kacprzyk, J. Stressed to Death: The role of transcription factors in plant programmed cell death induced by abiotic and biotic stimuli. Front. Plant Sci. 2020, 11, 1235. [Google Scholar] [CrossRef]
- Kumudini, S.; Savita, V.P. Antioxidant-mediated defense in triggering resistance against biotic stress in plants. In Biocontrol Agents and Secondary Metabolites, 1st ed.; Woodhead Publishing: Sawston, UK, 2021; pp. 383–399. [Google Scholar]
- Marcec, M.J.; Gilroy, S.; Poovaiah, B.W.; Tanaka, K. Mutual interplay of Ca2+ and ROS signaling in plant immune response. Plant Sci. 2019, 283, 343–354. [Google Scholar] [CrossRef]
- Pérez-Bueno, M.L.; Pineda, M.; Barón, M. Phenotyping plant responses to biotic stress by chlorophyll fluorescence imaging. Front. Plant Sci. 2019, 10, 1135. [Google Scholar] [CrossRef]
- González Guzmán, M.; Cellini, F.; Fotopoulos, V.; Balestrini, R.; Arbona, V. New approaches to improve crop tolerance to biotic and abiotic stresses. Physiol. Plant. 2022, 174, e13547. [Google Scholar] [CrossRef]
- Rousseau, C.; Belin, E.; Bove, E.; Rousseau, D.; Fabre, F.; Berruyer, R.; Guillaumès, J.; Manceau, C.; Jacques, M.A.; Boureau, T. High throughput quantitative phenotyping of plant resistance using chlorophyll fluorescence image analysis. Plant Methods 2013, 9, 17. [Google Scholar] [CrossRef]
- Zhang, C.; Zhou, L.; Xiao, Q.; Bai, X.; Wu, B.; Wu, N.; Zhao, Y.; Wang, J.; Feng, L. End-to-end fusion of hyperspectral and chlorophyll fluorescence imaging to identify rice stresses. Plant Phenomics 2022, 2022, 9851096. [Google Scholar] [CrossRef]
- Clark, R.E.; Basu, S.; Eigenbrode, S.D.; Oeller, L.C.; Crowder, D.W. Risk assessment for non-crop hosts of pea enation mosaic virus and the aphid vector Acyrthosiphon pisum. Agric. For. Entomol. 2013, 25, 427–434. [Google Scholar] [CrossRef]
- Chisholm, P.J.; Sertsuvalkul, N.; Casteel, C.L.; Crowder, D.W. Reciprocal plant-mediated interactions between a virus and a non-vector herbivore. Ecology 2018, 99, 2139–2144. [Google Scholar] [CrossRef]
- Cárcamo, H.A.; Vankosky, M.A.; Wijerathna, A.; Olfert, O.O.; Meers, S.B.; Evenden, M.L. Progress toward integrated pest management of pea leaf weevil: A review. Ann. Entomol. Soc. Am. 2018, 111, 144–153. [Google Scholar] [CrossRef]
- Chisholm, P.J.; Eigenbrode, S.D.; Clark, R.E.; Basu, S.; Crowder, D.W. Plant-mediated interactions between a vector and a non-vector herbivore promote the spread of a plant virus. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191383. [Google Scholar] [CrossRef] [PubMed]
- Black, A.E.; Strand, E.; Wright, R.G.; Scott, J.M.; Morgan, P.; Watson, C. Land use history at multiple scales: Implications for conservation planning. Landsc. Urban Plan. 1998, 43, 49–63. [Google Scholar] [CrossRef]
- National Library of Medicine, National Center for Biotechnology Information. Basic Local Alignment Search Tool (BLAST). 2023. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 24 September 2023).
- Schenk, S.T.; Schikora, A. Staining of callose depositions in root and leaf tissues. Bio-Protocol 2015, 5, e1429. [Google Scholar] [CrossRef]
- Yang, C.Y.; Hong, C.P. The NADPH oxidase Rboh D is involved in primary hypoxia signalling and modulates expression of hypoxia-inducible genes under hypoxic stress. Environ. Exp. Bot. 2015, 115, 63–72. [Google Scholar] [CrossRef]
- Mason, G.A.; Lemus, T.; Queitsch, C. The mechanistic underpinnings of an ago1-mediated, environmentally dependent, and stochastic phenotype. Plant Physiol. 2016, 170, 2420–2431. [Google Scholar] [CrossRef]
- van Geem, M.; Gols, R.; Raaijmakers, C.E.; Harvey, J.A. Effects of population-related variation in plant primary and secondary metabolites on aboveground and belowground multitrophic interactions. Chemoecology 2016, 26, 219–233. [Google Scholar] [CrossRef]
- Shikano, I. Evolutionary ecology of multitrophic interactions between plants, insect herbivores and entomopathogens. J. Chem. Ecol. 2017, 43, 586–598. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant-Microbe Interact. 2011, 24, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Kim, J.Y. Callose synthesis in higher plants. Plant Signal. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Sun, X.; Li, F.; Ma, N.; Wang, M.; Chen, Y.; Liu, N.; Jin, Y.; Zhang, J.; Hou, C.; et al. H2O2 mediates transcriptome reprogramming during soybean mosaic virus-induced callose deposition in soybean. Crop J. 2022, 10, 262–272. [Google Scholar] [CrossRef]
- Mbiza, N.I.T.; Hu, Z.; Zhang, H.; Zhang, Y.; Luo, X.; Wang, Y.; Wang, Y.; Liu, T.; Li, J.; Wang, X.; et al. GhCalS5 is involved in cotton response to aphid attack through mediating callose formation. Front. Plant Sci. 2022, 13, 892630. [Google Scholar] [CrossRef]
- Erb, M.; Reymond, P. Molecular interactions between plants and insect herbivores. Annu. Rev. Plant Biol. 2019, 29, 527–557. [Google Scholar] [CrossRef]
- Ye, M.; Glauser, G.; Lou, Y.; Erb, M.; Hu, L. Molecular dissection of early defense signaling underlying volatile-mediated defense regulation and herbivore resistance in rice. Plant Cell 2019, 31, 687–698. [Google Scholar] [CrossRef]
- Li, J.; Liu, X.; Wang, Q.; Huangfu, J.; Schuman, M.C.; Lou, Y. A group d MAPK protects plants from autotoxicity by suppressing herbivore-induced defense signaling. Plant Physiol. 2019, 179, 1386–1401. [Google Scholar] [CrossRef]
- Yi, G.; Wu, W.; Wei, T. Delivery of Rice Gall Dwarf Virus into plant phloem by its leafhopper vectors activates callose deposition to enhance viral transmission. Front. Microbiol. 2021, 12, 662577. [Google Scholar] [CrossRef]
- Chowdhury, R.N.; Lasky, D.; Karki, H.; Zhang, Z.; Goyer, A.; Halterman, D.; Rakotondrafara, A.M. HCPro suppression of callose deposition contributes to strain-specific resistance against potato virus Y. Phytopathology 2020, 110, 164–173. [Google Scholar] [CrossRef]
- Ellinger, D.; Naumann, M.; Falter, C.; Zwikowics, C.; Jamrow, T.; Manisseri, C.; Somerville, S.C.; Voigt, C.A. Elevated early callose deposition results in complete penetration resistance to powdery mildew in Arabidopsis. Plant Physiol. 2013, 161, 1433–1444. [Google Scholar] [CrossRef]
- Bittner, N.; Hundacker, J.; Achotegui-Castells, A.; Anderbrant, O.; Hilker, M. Defense of Scots pine against sawfly eggs (Diprion pini) is primed by exposure to sawfly sex pheromones. Proc. Natl. Acad. Sci. USA 2019, 116, 24668–24675. [Google Scholar] [CrossRef] [PubMed]
- Goggin, F.L.; Fischer, H.D. Reactive oxygen species in plant interactions with aphids. Front. Plant Sci. 2022, 12, 811105. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.A.; Gullner, G.; Clemente-Moreno, M.J.; Künstler, A.; Juhász, C.; Díaz-Vivancos, P.; Király, L. Oxidative stress and antioxidative responses in plant–virus interactions. Physiol. Mol. Plant Pathol. 2016, 94, 134–148. [Google Scholar] [CrossRef]
- Guo, H.; Gu, L.; Liu, F.; Chen, F.; Ge, F.; Sun, Y. Aphid-borne viral spread Is enhanced by virus-induced accumulation of plant reactive oxygen species. Plant Physiol. 2019, 179, 143–155. [Google Scholar] [CrossRef]
- Wagner, D.; Przybyla, D.; op den Camp, R.; Kim, C.; Landgraf, F.; Lee, K.P.; Würsch, M.; Laloi, C.; Nater, M.; Hideg, E.; et al. The genetic basis of singlet oxygen–induced stress responses of Arabidopsis thaliana. Science 2004, 306, 1183–1185. [Google Scholar] [CrossRef]
- Van Breusegem, F.; Dat, J.F. Reactive oxygen species in plant cell death. Plant Physiol. 2006, 141, 384–390. [Google Scholar] [CrossRef]
- Paiva, C.N.; Bozza, M.T. Are reactive oxygen species always detrimental to pathogens? Antioxid. Redox Signal. 2014, 20, 1000–1037. [Google Scholar] [CrossRef]
- Balint-Kurti, P. The plant hypersensitive response: Concepts, control and consequences. Mol. Plant Pathol. 2019, 20, 1163–1178. [Google Scholar] [CrossRef]
- Wang, H.; Lin, J.; Chang, Y.; Jiang, C.Z. Comparative transcriptomic analysis reveals that ethylene/H2O2-mediated hypersensitive response and programmed cell death determine the compatible interaction of sand pear and Alternaria alternata. Front. Plant Sci. 2017, 8, 195. [Google Scholar] [CrossRef]
- Jeblick, T.; Leisen, T.; Steidele, C.E.; Albert, I.; Müller, J.; Kaiser, S.; Mahler, F.; Sommer, F.; Keller, S.; Hückelhoven, R.; et al. Botrytis hypersensitive response inducing protein 1 triggers noncanonical PTI to induce plant cell death. Plant Physiol. 2023, 191, 125–141. [Google Scholar] [CrossRef]
- Salguero-Linares, J.; Coll, N.S. Cell death as a defense strategy against pathogens in plants and animals. PLoS Pathog. 2023, 19, e1011253. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Yoon, J.B.; Park, H.G.; Park, E.W.; Kim, Y.H. Structural modifications and programmed cell death of chili pepper fruit related to resistance responses to Colletotrichum gloeosporioides infection. Phytopathology 2004, 94, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Goggin, F.L. Plant-aphid interactions: Molecular and ecological perspectives. Curr. Opin. Plant Biol. 2007, 10, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 2007, 59, 501–520. [Google Scholar] [CrossRef] [PubMed]
- Goggin, F.L.; Lorence, A.; Topp, C.N. Applying high-throughput phenotyping to plant–insect interactions: Picturing more resistant crops. Curr. Opin. Insect Sci. 2015, 9, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Poque, S.; Valkonen, J.P.T. Phenotyping viral infection in sweet potato using a high-throughput chlorophyll fluorescence and thermal imaging platform. Plant Methods 2019, 15, 116. [Google Scholar] [CrossRef]
- Rahoutei, J.; García-Luque, I.; Barón, M. Inhibition of photosynthesis by viral infection: Effect on PSII structure and function. Physiol. Plant. 2000, 110, 286–292. [Google Scholar] [CrossRef]
- Farooq, T.; Liu, D.; Zhou, X.; Yang, Q. Tomato Yellow Leaf Curl China Virus impairs photosynthesis in the infected Nicotiana benthamiana with βC1 as an aggravating factor. Plant Pathol. J. 2019, 35, 521–529. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Amplicon Size (bp) |
|---|---|---|
| PEMP CP Fp PEMP CP Rp | GTGGTGGCACCCTCTATG GTGTCCACATGGTAGGCTATG | 290 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basu, S.; Moroz, N.; Lee, B.W.; Tanaka, K.; Oeller, L.; Baerlocher, C.W.; Crowder, D.W. Diversity and Traits of Multiple Biotic Stressors Elicit Differential Defense Responses in Legumes. Agriculture 2023, 13, 2093. https://doi.org/10.3390/agriculture13112093
Basu S, Moroz N, Lee BW, Tanaka K, Oeller L, Baerlocher CW, Crowder DW. Diversity and Traits of Multiple Biotic Stressors Elicit Differential Defense Responses in Legumes. Agriculture. 2023; 13(11):2093. https://doi.org/10.3390/agriculture13112093
Chicago/Turabian StyleBasu, Saumik, Natalia Moroz, Benjamin W. Lee, Kiwamu Tanaka, Liesl Oeller, Chase W. Baerlocher, and David W. Crowder. 2023. "Diversity and Traits of Multiple Biotic Stressors Elicit Differential Defense Responses in Legumes" Agriculture 13, no. 11: 2093. https://doi.org/10.3390/agriculture13112093