
The Biocontrol of Plant Pathogenic Fungi by Selected Lactic Acid Bacteria: From Laboratory to Field Study
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Isolation of Lactic Acid Bacteria
2.3. Preparation of Cell-Free Supernatant from Bacterial Cultures
2.4. Antifungal Activity of Lactic Acid Bacteria
2.5. Identification of Lactic Acid Bacteria
2.6. Culture Preparation for Seed Treatment
2.7. Seed Treatment
2.8. Mini-Plot Experiments
2.9. Field Experiments
2.9.1. Disease Assessment
2.9.2. Plant Emergence and Overwintering Determination
2.9.3. Assessment of the Phytotoxicity Effects of Bacterial Cultures on the Crop
2.9.4. Yield Grain Determination
2.9.5. Weight of 1000 Grains Assessment
2.10. Statistical Analysis
3. Results
3.1. Antifungal Activity of Isolated Lactic Acid Bacteria
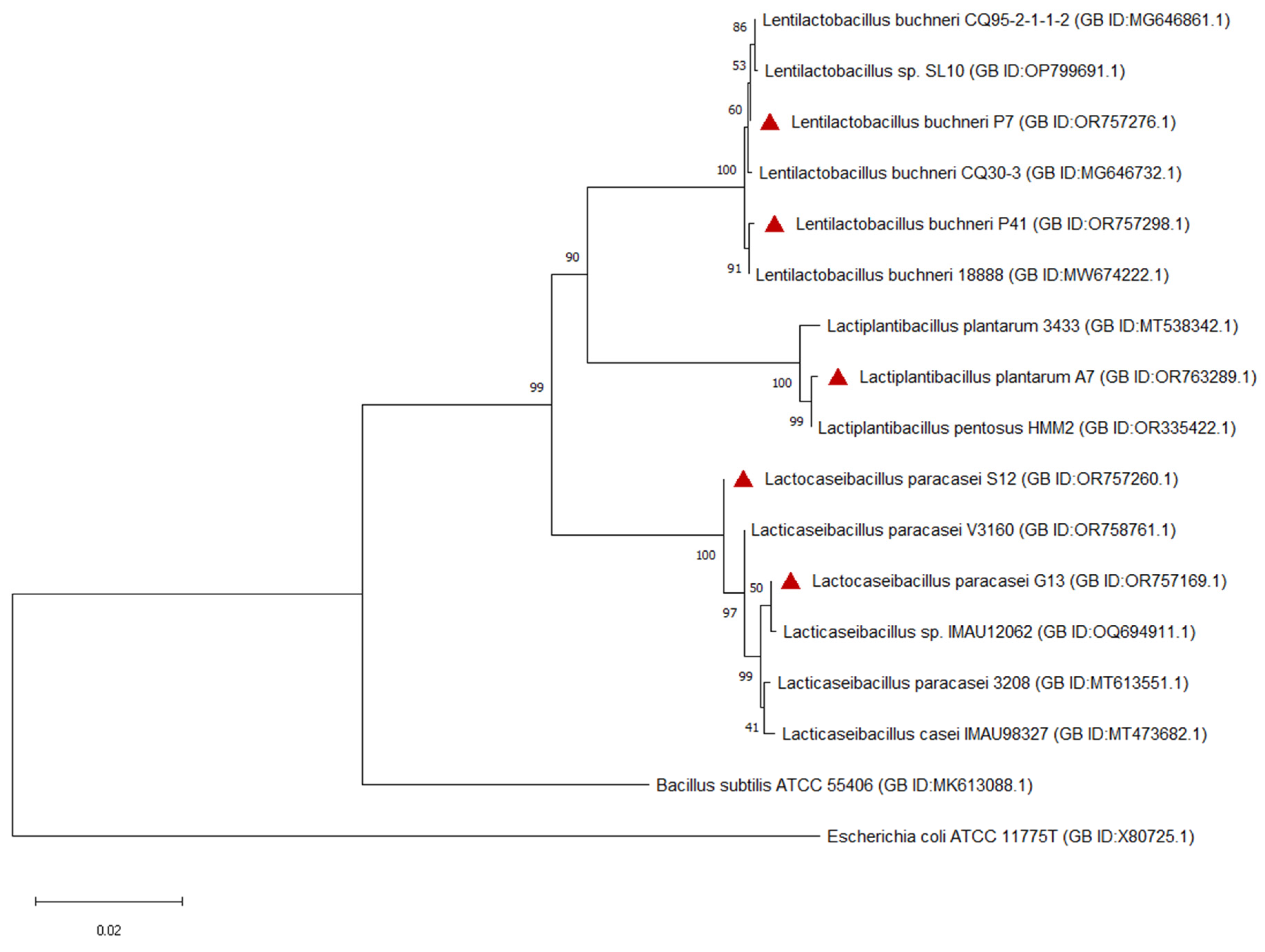
3.2. The Identification of Selected Strains of Lactic Acid Bacteria
3.3. The Efficacy of Tested Bacteria in the Control of Chosen Diseases of Wheat and Rye in Mini-Plot Experiments
3.4. The Efficacy of Tested Bacteria in the Control of Chosen Diseases of Wheat and Rye in Field Experiments
3.4.1. The Effect of Tested Bacteria on the Fungal Disease Severity
3.4.2. The Effect of LAB on Plant Growth, Yield Parameters, and Phytotoxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Directive 2009/128/EC of the European Parliament and of the Council of 21 October 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/PDF/?uri=CELEX:32009L0128&from=pl (accessed on 2 February 2022).
- Annex to the Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions. Available online: https://eur-lex.europa.eu/resource.html?uri=cellar:b828d165-1c22-11ea-8c1f-01aa75ed71a1.0002.02/DOC_2&format=PDF (accessed on 2 February 2022).
- Heimpel, G.E.; Mills, N. Biological Control—Ecology and Applications; Cambridge University Press: Cambridge, UK, 2017. [Google Scholar]
- World Health Organization. Public Health Impact of Pesticides Used in Agriculture; World Health Organization: Geneva, Switzerland, 1990. [Google Scholar]
- Stoytcheva, M. (Ed.) Pesticides in the Modern World: Effects of Pesticides Exposure; BoD—Books on Demand: Norderstedt, Germany, 2011. [Google Scholar]
- Nicolopoulou-Stamati, P.; Maipas, S.; Kotampasi, C.; Stamatis, P.; Hens, L. Chemical pesticides and human health: The urgent need for a new concept in agriculture. Front. Public Health 2016, 4, 148. [Google Scholar] [CrossRef] [PubMed]
- Isenring, R. Pesticides reduce biodiversity. Pestic. News 2010, 88, 4–7. [Google Scholar]
- Leahy, J.; Mendelsohn, M.; Kough, J.; Jones, R.; Berckes, N. Biopesticide oversight and registration at the US Environmental Protection Agency. In Biopesticides: State of the Art and Future Opportunities; ACS Publications: Washington, DC, USA, 2014; pp. 3–18. [Google Scholar]
- Benhamou, N.; Le Floch, G.; Vallance, J.; Gerbore, J.; Grizard, D.; Rey, P. Pythium oligandrum: An example of opportunistic success. Microbiology 2012, 158, 2679–2694. [Google Scholar] [CrossRef] [PubMed]
- Klimek-Kopyra, A.; Dłużniewska, J.; Sikora, A. Influence of Biofungicides Containing Microorganisms Such as Pythium oligandrum and Bacillus subtilis on Yield, Morphological Parameters, and Pathogen Suppression in Six Winter Pea Cultivars. Agriculture 2023, 13, 1170. [Google Scholar] [CrossRef]
- Ibáñez, A.; Garrido-Chamorro, S.; Vasco-Cárdenas, M.F.; Barreiro, C. From Lab to Field: Biofertilizers in the 21st Century. Horticulturae 2023, 9, 1306. [Google Scholar] [CrossRef]
- Corsetti, A.; Gobbetti, M.; Rossi, J.; Damiani, P. Antimould activity of sourdough lactic acid bacteria: Identification of a mixture of organic acids produced by Lactobacillus sanfrancisco CB1. Appl. Microbiol. Biotechnol. 1998, 50, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Lavermicocca, P.; Valerio, F.; Visconti, A. Antifungal activity of phenyllactic acid against molds isolated from bakery products. Appl. Environ. Microbiol. 2003, 69, 634–640. [Google Scholar] [CrossRef]
- Ström, K.; Sjögren, J.; Broberg, A.; Schnürer, J. Lactobacillus plantarum MiLAB 393 produces the antifungal cyclic dipeptides cyclo (L-Phe-L-Pro) and cyclo (L-Phe-trans-4-OH-L-Pro) and 3-phenyllactic acid. Appl. Environ. Microbiol. 2002, 68, 4322–4327. [Google Scholar] [CrossRef]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- De Vuyst, L.; Leroy, F. Bacteriocins from lactic acid bacteria: Production, purification, and food applications. Microb. Physiol. 2007, 13, 194–199. [Google Scholar] [CrossRef]
- Gerez, C.L.; Torino, M.I.; Rollán, G.; de Valdez, G.F. Prevention of bread mould spoilage by using lactic acid bacteria with antifungal properties. Food Control 2009, 20, 144–148. [Google Scholar] [CrossRef]
- Chen, J.; Guttieri, M.J.; Zhang, J.; Hole, D.; Souza, E.; Goates, B. A novel QTL associated with dwarf bunt resistance in Idaho 444 winter wheat. Theor. Appl. Genet. 2016, 129, 2313–2322. [Google Scholar] [CrossRef] [PubMed]
- Mourad, A.; Mahdy, E.; Bakheit, B.R.; Abo-Elwafaa, A.; Baenziger, P.S. Effect of common bunt infection on agronomic traits in wheat (Triticum aestivum L.). J. Plant Breed. Genet. 2018, 2, 1. [Google Scholar]
- Borgen, A.; Kristensen, L. Effect of seed treatment with milk powder and mustard flour in control of common bunt (Tilletia tritici) in wheat and stem smut (Urocystis occulta) in rye. In Proceedings from BCPC Symposium No. 76:“Seed Treatment: Challenges & Opportunities”; British Crop Protection Council: Farnham, UK, 2001. [Google Scholar]
- Arie, T. Fusarium diseases of cultivated plants, control, diagnosis, and molecular and genetic studies. J. Pestic. Sci. 2019, 44, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome perspective. Front. Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef]
- Johns, L.E.; Bebber, D.P.; Gurr, S.J.; Brown, N.A. Emerging health threat and cost of Fusarium mycotoxins in European wheat. Nat. Food 2022, 3, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Ajayi-Oyetunde, O.O.; Bradley, C.A. Rhizoctonia solani: Taxonomy, population biology and management of rhizoctonia seedling disease of soybean. Plant Pathol. 2018, 67, 3–17. [Google Scholar] [CrossRef]
- Kara, A.; Arici, Ş.E. Determination of Gamma Rays Efficiency Against Rhizoctonia solani in Potatoes. Open Chem. 2019, 17, 254–259. [Google Scholar] [CrossRef]
- Hou, Y.; Ma, X.; Wan, W.; Long, N.; Zhang, J.; Tan, Y.; Duan, S.; Zeng, Y.; Dong, Y. Comparative genomics of pathogens causing brown spot disease of tobacco: Alternaria longipes and Alternaria alternata. PLoS ONE 2016, 11, e0155258. [Google Scholar] [CrossRef]
- Nowicki, M.; Nowakowska, M.; Niezgoda, A.; Kozik, E. Alternaria black spot of crucifers: Symptoms, importance of disease, and perspectives of resistance breeding. Veg. Crops Res. Bull. 2012, 76, 5–19. [Google Scholar] [CrossRef]
- Al-Lami, H.F.D.; You, M.P.; Barbetti, M.J. Incidence, pathogenicity and diversity of Alternaria spp. associated with Alternaria leaf spot of canola (Brassica napus) in Australia. Plant Pathol. 2019, 68, 492–503. [Google Scholar] [CrossRef]
- Wang, N.Y.; Forcelini, B.B.; Peres, N.A. Anthracnose fruit and root necrosis of strawberry are caused by a dominant species within the Colletotrichum acutatum species complex in the United States. Phytopathology 2019, 109, 1293–1301. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, G.; Tan, Q.; Schneider, V.; Ishii, H. Inherent tolerance of Colletotrichum gloeosporioides to fludioxonil. Pestic. Biochem. Physiol. 2021, 172, 104767. [Google Scholar] [CrossRef] [PubMed]
- Willbur, J.; McCaghey, M.; Kabbage, M.; Smith, D.L. An overview of the Sclerotinia sclerotiorum pathosystem in soybean: Impact, fungal biology, and current management strategies. Trop. Plant Pathol. 2019, 44, 3–11. [Google Scholar] [CrossRef]
- Parente, E.; Brienza, C.; Moles, M.; Ricciardi, A. A comparison of methods for the measurement of bacteriocin activity. J. Microbiol. Methods 1995, 22, 95–108. [Google Scholar] [CrossRef]
- Prema, P.; Smila, D.; Palavesam, A.; Immanuel, G. Production and characterization of an antifungal compound (3-phenyllactic acid) produced by Lactobacillus plantarum strain. Food Bioprocess Technol. 2010, 3, 379–386. [Google Scholar] [CrossRef]
- Dec, M.; Urban-Chmiel, R.; Gnat, S.; Puchalski, A.; Wernicki, A. Identification of Lactobacillus strains of goose origin using MALDI-TOF mass spectrometry and 16S–23S rDNA intergenic spacer PCR analysis. Res. Microbiol. 2014, 165, 190–201. [Google Scholar] [CrossRef]
- Dec, M.; Puchalski, A.; Urban-Chmiel, R.; Wernicki, A. 16S-ARDRA and MALDI-TOF mass spectrometry as tools for identification of Lactobacillus bacteria isolated from poultry. BMC Microbiol. 2016, 16, 105. [Google Scholar] [CrossRef]
- Leser, T.D.; Amenuvor, J.Z.; Jensen, T.K.; Lindecrona, R.H.; Boye, M.; Møller, K. Culture-independent analysis of gut bacteria: The pig gastrointestinal tract microbiota revisited. Appl. Environ. Microbiol. 2002, 68, 673–690. [Google Scholar] [CrossRef]
- Pang, H.; Qin, G.; Tan, Z.; Li, Z.; Wang, Y.; Cai, Y. Natural populations of lactic acid bacteria associated with silage fermentation as determined by phenotype, 16S ribosomal RNA and recA gene analysis. Syst. Appl. Microbiol. 2011, 34, 235–241. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- EPPO PP/1/181 (4). 2012. Available online: https://gepeu.files.wordpress.com/2014/11/pp1-181-4-e-conduct-and-reporting-of-efficacy-evaluation-trials-including-good-experimental-practice.pdf (accessed on 2 October 2023).
- EPPO PP 1/152 (4). 2012. Available online: https://pp1.eppo.int/standards/PP1-152-4 (accessed on 2 October 2023).
- EPPO PP/1/135 (4). 2014. Available online: https://pp1.eppo.int/standards/PP1-135-4 (accessed on 2 October 2023).
- EPPO PP/1/19 (4). 2003. Available online: https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1365-2338.2004.00691.x (accessed on 2 October 2023).
- BBCH. 2018. Available online: https://www.julius-kuehn.de/en/jki-publication-series/bbch-scale/ (accessed on 2 October 2023).
- Bowler, I. Developing sustainable agriculture. Geography 2002, 87, 205–212. [Google Scholar]
- Nandhini, S.; Sendhilvel, V.; Babu, S. Endophytic bacteria from tomato and their efficacy against Fusarium oxysporum f. sp. lycopersici, the wilt pathogen. J. Biopestic. 2012, 5, 178. [Google Scholar]
- Abdulkareem, M.; Aboud, H.M.; Saood, H.M.; Shibly, M.K. Antagonistic activity of some plant growth rhizobacteria to Fusarium graminearum. Int. J. Phytopathol. 2014, 3, 49–54. [Google Scholar] [CrossRef]
- Yezli, W.; Zebboudj-Yezli, N.; Hamini-Kadar, N.; Kihal, M.; Henni, J.E. An in vitro antagonistic activity evaluation of rhizobacteria against Fusarium oxysporum f. sp. radicis-lycopersici (Forl) isolated from the Algerian west. Int. J. Biosci. 2015, 7, 95–103. [Google Scholar]
- Moreno-Mateos, M.A.; Delgado-Jarana, J.; Codon, A.C.; Benítez, T. pH and Pac1 control development and antifungal activity in Trichoderma harzianum. Fungal Genet. Biol. 2007, 44, 1355–1367. [Google Scholar] [CrossRef]
- Shentu, X.; Zhan, X.; Ma, Z.; Yu, X.; Zhang, C. Antifungal activity of metabolites of the endophytic fungus Trichoderma brevicompactum from garlic. Braz. J. Microbiol. 2014, 45, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Raman, J.; Kim, J.S.; Choi, K.R.; Eun, H.; Yang, D.; Ko, Y.J.; Kim, S.J. Application of lactic acid bacteria (LAB) in sustainable agriculture: Advantages and limitations. Int. J. Mol. Sci. 2022, 23, 7784. [Google Scholar] [CrossRef]
- Magnusson, J.; Ström, K.; Roos, S.; Sjögren, J.; Schnürer, J. Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol. Lett. 2003, 219, 129–135. [Google Scholar] [CrossRef]
- Trias Mansilla, R.; Bañeras Vives, L.; Montesinos Seguí, E.; Badosa Romañó, E. Lactic acid bacteria from fresh fruit and vegetables as biocontrol agents of phytopathogenic bacteria and fungi. Int. Microbiol. 2008, 11, 231–236. [Google Scholar]
- Zaiton, H.; Kh, S.S.; Aqilah, Z.N.; Azfar, A.A. Effect of pH and heat treatment on antifungal activity of Lactobacillus fermentum Te007, Lactobacillus pentosus G004 and Pediococcus pentosaceus Te010. Innov. Rom. Food Biotechnol. 2011, 8, 41–53. [Google Scholar]
- Cortés-Zavaleta, O.; López-Malo, A.; Hernández-Mendoza, A.; García, H.S. Antifungal activity of lactobacilli and its relationship with 3-phenyllactic acid production. Int. J. Food Microbiol. 2014, 173, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Gwiazdowski, R.; Kluczyńska, K.; Gwiazdowska, D. Impact of the selected lactic acid bacteria on the growth of pathogens occurring in oilseed rape crops/Wpływ wybranych bakterii fermentacji mlekowej na wzrost patogenów występujących w uprawie rzepaku. Prog. Plant Prot. 2015, 55, 446–451. [Google Scholar]
- Dalie, D.K.D.; Deschamps, A.M.; Atanasova-Penichon, V.; Richard-Forget, F. Potential of Pediococcus pentosaceus (L006) isolated from maize leaf to suppress fumonisin-producing fungal growth. J. Food Prot. 2010, 73, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Steglińska, A.; Kołtuniak, A.; Motyl, I.; Berłowska, J.; Czyżowska, A.; Cieciura-Włoch, W.; Okrasa, M.; Kręgiel, D.; Gutarowska, B. Lactic acid bacteria as biocontrol agents against potato (Solanum tuberosum L.) pathogens. Appl. Sci. 2022, 12, 7763. [Google Scholar] [CrossRef]
- Lačanin, I.; Mounier, J.; Pawtowski, A.; Dušková, M.; Kameník, J.; Karpíšková, R. Assessment of the antifungal activity of Lactobacillus and Pediococcus spp. for use as bioprotective cultures in dairy products. World J. Microbiol. Biotechnol. 2017, 33, 188. [Google Scholar] [CrossRef]
- De Simone, N.; Capozzi, V.; de Chiara, M.L.V.; Amodio, M.L.; Brahimi, S.; Colelli, G.; Drider, D.; Spano, G.; Russo, P. Screening of lactic acid bacteria for the bio-control of Botrytis cinerea and the potential of Lactiplantibacillus plantarum for eco-friendly preservation of fresh-cut kiwifruit. Microorganisms 2021, 9, 773. [Google Scholar] [CrossRef]
- Quattrini, M.; Bernardi, C.; Stuknytė, M.; Masotti, F.; Passera, A.; Ricci, G.; Vallone, L.; De Noni, I.; Brasca, M.; Fortina, M.G. Functional characterization of Lactobacillus plantarum ITEM 17215: A potential biocontrol agent of fungi with plant growth promoting traits, able to enhance the nutritional value of cereal products. Food Res. Int. 2018, 106, 936–944. [Google Scholar] [CrossRef]
- Kharazian, Z.A.; Jouzani, G.S.; Aghdasi, M.; Khorvash, M.; Zamani, M.; Mohammadzadeh, H. Biocontrol potential of Lactobacillus strains isolated from corn silages against some plant pathogenic fungi. Biol. Control 2017, 110, 33–43. [Google Scholar] [CrossRef]
- Barrios-Roblero, C.; Rosas-Quijano, R.; Salvador-Figueroa, M.; Gálvez-López, D.; Vázquez-Ovando, A. Antifungal lactic acid bacteria isolated from fermented beverages with activity against Colletotrichum gloeosporioides. Food Biosci. 2019, 29, 47–54. [Google Scholar] [CrossRef]
- Ramos-Pereira, J.; Mareze, J.; Fernández, D.; Rios, E.A.; Santos, J.A.; López-Díaz, T.M. Antifungal activity of lactic acid bacteria isolated from milk against Penicillium commune, P. nordicum, and P. verrucosum. Int. J. Food Microbiol. 2021, 355, 109331. [Google Scholar] [CrossRef] [PubMed]
- Byrne, M.B.; Thapa, G.; Doohan, F.M.; Burke, J.I. Lactic acid bacteria as potential biocontrol agents for Fusarium head blight disease of spring barley. Front. Microbiol. 2022, 13, 912632. [Google Scholar] [CrossRef] [PubMed]
- López-Seijas, J.; García-Fraga, B.; da Silva, A.F.; Sieiro, C. Wine lactic acid bacteria with antimicrobial activity as potential biocontrol agents against Fusarium oxysporum f. sp. lycopersici. Agronomy 2019, 10, 31. [Google Scholar] [CrossRef]
- Shrestha, A.; Kim, B.S.; Park, D.H. Biological control of bacterial spot disease and plant growth-promoting effects of lactic acid bacteria on pepper. Biocontrol Sci. Technol. 2014, 24, 763–779. [Google Scholar] [CrossRef]
- Mohd Taha, M.D.; Mohd Jaini, M.F.; Saidi, N.B.; Abdul Rahim, R.; Md Shah, U.K.; Mohd Hashim, A. Biological control of Erwinia mallotivora, the causal agent of papaya dieback disease by indigenous seed-borne endophytic lactic acid bacteria consortium. PLoS ONE 2019, 14, e0224431. [Google Scholar] [CrossRef]
- Hamed, H.A.; Moustafa, Y.A.; Abdel-Aziz, S.M. In vivo efficacy of lactic acid bacteria in biological control against Fusarium oxysporum for protection of tomato plant. Life Sci. J. 2011, 8, 462–468. [Google Scholar]
- Zebboudj, N.; Yezli, W.; Hamini-Kadar, N.; Kihal, M. Antifungal activity of lactic acid bacteria against Fusarium species responsible for tomato crown and root rots. Environ. Exp. Biol. 2020, 18, 7–13. [Google Scholar]
- Kim, Y.C.; Anderson, A.J. Rhizosphere pseudomonads as probiotics improving plant health. Mol. Plant Pathol. 2018, 19, 2349–2359. [Google Scholar] [CrossRef]


{kind=link}
| LAB Strain | Antifungal Activity (AU/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| Fusarium graminearum | Fusarium culmorum | Fusarium oxysporum | Fusarium poae | Rhizoctonia solani | Alternaria alternata | Colletotrichum gleosporioides | Sclerotinia sclerotiorum | |
| P7.Z | 400 | 400 | 200 | 400 | 1600 | 800 | 800 | 800 |
| P7L | 400 | 400 | 200 | 400 | 1600 | 800 | 800 | 800 |
| P7 | 400 | 400 | 400 | 400 | 1600 | 800 | 800 | 800 |
| A2.P | 400 | 400 | 200 | 400 | 1600 | 400 | 400 | 400 |
| P40 | 400 | 400 | 200 | 400 | 400 | 800 | 800 | 800 |
| P41 | 400 | 400 | 400 | 400 | 1600 | 1600 | 800 | 800 |
| P42 | <200 | <200 | <200 | <200 | 400 | <200 | 200 | <200 |
| G13 | 800 | 400 | 200 | 400 | 800 | 400 | 800 | 800 |
| S12 | 800 | 400 | 400 | 400 | 800 | 400 | 400 | 400 |
| A7 | 400 | 400 | 400 | 400 | 800 | 400 | 400 | 800 |
| Treatment | Season 1 | Season 2 | ||
|---|---|---|---|---|
| Average % of Infected Stems | Average % of Efficacy | Average % of Infected Stems | Average % of Efficacy | |
| Control/Untreated | 11.3 a | - | 11.9 a | - |
| Lactiplantibacillus plantarum A7 | 0.6 b | 94.7 | 1.3 b | 89.1 |
| Lacticaseibacillus paracasei G13 | 0.6 b | 94.7 | 7.8 a | 34.5 |
| Lacticaseibacillus paracasei S12 | 1.1 b | 90.3 | 1.1 b | 90.8 |
| Lentilactobacillus buchneri P7 | 2.5 b | 77.9 | 1.5 b | 87.4 |
| Lentilactobacillus buchneri P41 | 5.2 ab | 54.0 | 2.1 b | 82.4 |
| Vibrance Gold 100 FS | 0.6 b | 94.7 | 0.4 b | 96.6 |
| LSD 0.05 | 7.06 | 4.46 | ||
| Treatment | Season 1 | Season 2 | ||
|---|---|---|---|---|
| Average % of Infected Ears | Average % of Efficacy | Average % of Infected Ears | Average % of Efficacy | |
| Control/Untreated | 57.5 a | - | 73.9 a | - |
| Lactiplantibacillus plantarum A7 | 28.7 bc | 50.1 | 0.3 c | 99.6 |
| Lacticaseibacillus paracasei G13 | 15.0 c | 73.9 | 55.6 b | 24.8 |
| Lacticaseibacillus paracasei S12 | 36.6 b | 36.3 | 1.9 c | 97.4 |
| Lentilactobacillus buchneri P7 | 27.3 bc | 52.5 | 1.0 c | 98.6 |
| Lentilactobacillus buchneri P41 | 32.7 b | 43.1 | 5.2 c | 93.0 |
| Vitavax 200 FS | 0.0 d | 100 | 1.4 c | 98.1 |
| LSD 0.05 | 14.49 | 8.35 | ||
| Treatment | Rye | Wheat | ||||||
|---|---|---|---|---|---|---|---|---|
| Season 1 | Season 2 | Season 1 | Season 2 | |||||
| Average % of Infected Plants | Average % of Efficacy | Average % of Infected Plants | Average % of Efficacy | Average % of Infected Plants | Average % of Efficacy | Average % of Infected Plants | Average % of Efficacy | |
| Control/Untreated | 25.8 a | - | 28.6 a | - | 43.5 a | - | 23.9 a | - |
| L. plantarum A7 | 15.7 ab | 39.1 | 16.0 bc | 44.1 | 21.6 b | 50.3 | 14.1 ab | 41.0 |
| L. paracasei G13 | 25.7 a | 0.4 | 18.3 b | 36.0 | 22.0 b | 49.4 | 12.6 b | 47.3 |
| L. paracasei S12 | 22.5 a | 12.8 | 14.2 bc | 50.3 | 22.4 b | 48.5 | 15.4 a | 35.6 |
| L. buchneri P7 | 21.6 a | 16.3 | 22.9 a | 19.9 | 27.3 b | 37.2 | 7.7 b | 67.8 |
| L. buchneri P41 | 22.5 a | 12.8 | 20.4 ab | 28.7 | 23.2 b | 46.7 | 12.6 a | 47.3 |
| Vibrance Gold 100 FS | 11.3 b | 56.2 | 7.2 c | 74.8 | nt. | nt. | nt. | nt. |
| Vitavax 100 FS | nt. | nt. | nt. | nt. | 18.7 b | 57.0 | 8.0 a | 66.5 |
| LSD 0.05 | 10.68 | 9.52 | 10.18 | 9.82 | ||||
| Treatment | Season 1 | Season 2 | ||
|---|---|---|---|---|
| Average Number of Infected Stems per Plot | Average % of Efficacy | Average Number of Infected Stems per Plot | Average % of Efficacy | |
| Control/Untreated | 503.3 a | - | 91.8 a | - |
| Lactiplantibacillus plantarum A7 | 0.0 b | 100 | 0.5 b | 99.5 |
| Lacticaseibacillus paracasei G13 | 16.5 b | 96.7 | 1.8 b | 98.0 |
| Lacticaseibacillus paracasei S12 | 24.8 b | 95.1 | 4.0 b | 95.6 |
| Lentilactobacillus buchneri P7 | 57.8 b | 88.5 | 0.0 b | 100 |
| Lentilactobacillus buchneri P41 | 66.0 b | 86.9 | 1.3 b | 98.6 |
| Vibrance Gold 100 FS | 0.0 b | 100 | 0.0 b | 100 |
| LSD 0.05 | 125.68 | 19.50 | ||
| Treatment | Season 1 | Season 2 | ||
|---|---|---|---|---|
| Average % of Infected Ears | Average % of Efficacy | Average % of Infected Ears | Average % of Efficacy | |
| Control/Untreated | 17.5 a | - | 56.5 a | - |
| Lactiplantibacillus plantarum A7 | 0.5 d | 97.1 | 5.5 b | 90.3 |
| Lacticaseibacillus paracasei G13 | 1.5 c | 91.4 | 31.5 ab | 44.2 |
| Lacticaseibacillus paracasei S12 | 2.0 c | 88.6 | 19.5 b | 65.5 |
| Lentilactobacillus buchneri P7 | 4.0 c | 77.1 | 0.5 b | 99.1 |
| Lentilactobacillus buchneri P41 | 6.5 bc | 62.9 | 7.0 b | 87.6 |
| Vitavax 200 FS | 0.0 d | 100 | 0.0 b | 100 |
| LSD 0.05 | 5.11 | 25.29 | ||
| Treatment | Season 1 | Season 2 | ||
|---|---|---|---|---|
| Plant Emergence * | Overwintering * | Plant Emergence * | Overwintering * | |
| 12 November 2015 BBCH 12 | 17 March 2016 BBCH 23 | 12 December 2016 BBCH 12 | 31 March 2017 BBCH 23 | |
| Control/Untreated | 94.00 a | 66.75 a | 55.00 a | 52.88 a |
| Lactiplantibacillus plantarum A7 | 85.00 ab | 74.00 a | 44.38 a | 43.00 a |
| Lacticaseibacillus paracasei G13 | 76.75 ab | 65.25 a | 46.63 a | 45.00 a |
| Lacticaseibacillus paracasei S12 | 87.50 ab | 70.25 a | 47.00 a | 44.75 a |
| Lentilactobacillus buchneri P7 | 75.25 b | 57.75 a | 49.50 a | 46.25 a |
| Lentilactobacillus buchneri P41 | 82.00 ab | 65.75 a | 46.63 a | 44.63 a |
| Vitavax 200 FS | 95.75 a | 77.50 a | 49.50 a | 47.00 a |
| LSD 0.05 | 18.382 | 16.557 | 11.039 | 11.256 |
| Treatment | Season 1 | Season 2 | ||
|---|---|---|---|---|
| Plant Emergence * | Overwintering * | Plant Emergence * | Overwintering * | |
| 12 November 2015 BBCH 12 | 17 March 2016 BBCH 23 | 12 December 2016 BBCH 12 | 31 March 2017 BBCH 23 | |
| Control/Untreated | 80.50 a | 66.50 a | 44.13 a | 42.38 a |
| Lactiplantibacillus plantarum A7 | 59.25 b | 52.00 ab | 30.25 b | 28.38 c |
| Lacticaseibacillus paracasei G13 | 50.75 b | 45.50 b | 44.00 a | 41.00 ab |
| Lacticaseibacillus paracasei S12 | 45.75 b | 37.50 b | 33.50 b | 29.88 c |
| Lentilactobacillus buchneri P7 | 58.00 b | 54.00 a | 29.00 b | 27.50 c |
| Lentilactobacillus buchneri P41 | 65.25 ab | 54.75 a | 34.75 b | 32.00 bc |
| Vibrance Gold FS | 83.75 a | 74.75 a | 50.63 a | 48.25 a |
| LSD 0.05 | 16.583 | 15.041 | 9.350 | 10.077 |
| Treatment | Grain Yield | Weight of 1000 Grains | ||||||
|---|---|---|---|---|---|---|---|---|
| Season 1 | Season 2 | Season 1 | Season 2 | |||||
| t/ha | % Increase * | t/ha | % Increase * | g | % Increase * | g | % Increase * | |
| Control/Untreated | 6.43 b | - | 5.63 c | - | 33.31 b | - | 36.35 b | - |
| L. plantarum A7 | 6.30 c | −1.95 | 5.88 c | 4.44 | 36.21 a | 8.71 | 35.43 b | −2.53 |
| L. paracasei G13 | 6.75 a | 5.06 | 5.78 c | 2.67 | 35.93 a | 7.87 | 36.45 ab | 0.27 |
| L. paracasei S12 | 6.43 b | 0.00 | 5.60 c | −0.44 | 36.69 a | 10.16 | 37.82 a | 4.04 |
| L. buchneri P7 | 6.28 c | −2.33 | 5.78 c | 2.67 | 37.07 a | 11.28 | 38.81 a | 6.75 |
| L. buchneri P41 | 6.23 c | −3.11 | 6.75 a | 20.00 | 33.30 b | −0.03 | 38.96 a | 7.16 |
| Vibrance Gold 100 FS | 6.18 c | −3.89 | 6.38 b | 13.33 | 33.12 b | −0.56 | 37.02 a | 1.83 |
| LSD 0.05 | 0.122 | 0.282 | 2.043 | 2.529 | ||||
| Treatment | Grain Yield | Weight of 1000 Grains | ||||||
|---|---|---|---|---|---|---|---|---|
| Season 1 | Season 2 | Season 1 | Season 2 | |||||
| t/ha | % Increase * | t/ha | % Increase * | g | % Increase * | g | % Increase * | |
| Control/Untreated | 1.15 e | - | 2.28 d | - | 34.61 b | - | 35.23 a | - |
| L. plantarum A7 | 3.00 b | 160.87 | 5.40 a | 137.36 | 37.12 ab | 7.26 | 36.61 a | 3.94 |
| L. paracasei G13 | 3.40 a | 195.65 | 3.25 c | 42.86 | 38.31 ab | 10.68 | 35.93 a | 2.01 |
| L. paracasei S12 | 2.50 c | 117.39 | 5.13 a | 125.27 | 43.43 a | 25.48 | 38.23 a | 8.53 |
| L. buchneri P7 | 2.53 c | 119.57 | 5.25 a | 130.77 | 35.82 b | 3.51 | 35.50 a | 0.78 |
| L. buchneri P41 | 2.20 d | 91.30 | 4.78 b | 109.89 | 35.10 b | 1.41 | 35.60 a | 1.07 |
| Vitavax 200 FS | 3.50 a | 204.35 | 5.20 a | 128.57 | 38.86 ab | 12.27 | 36.26 a | 2.94 |
| LSD 0.05 | 0.216 | 0.309 | 7.911 | 3.516 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gwiazdowski, R.; Kubiak, K.; Juś, K.; Marchwińska, K.; Gwiazdowska, D. The Biocontrol of Plant Pathogenic Fungi by Selected Lactic Acid Bacteria: From Laboratory to Field Study. Agriculture 2024, 14, 61. https://doi.org/10.3390/agriculture14010061
Gwiazdowski R, Kubiak K, Juś K, Marchwińska K, Gwiazdowska D. The Biocontrol of Plant Pathogenic Fungi by Selected Lactic Acid Bacteria: From Laboratory to Field Study. Agriculture. 2024; 14(1):61. https://doi.org/10.3390/agriculture14010061
Chicago/Turabian StyleGwiazdowski, Romuald, Krzysztof Kubiak, Krzysztof Juś, Katarzyna Marchwińska, and Daniela Gwiazdowska. 2024. "The Biocontrol of Plant Pathogenic Fungi by Selected Lactic Acid Bacteria: From Laboratory to Field Study" Agriculture 14, no. 1: 61. https://doi.org/10.3390/agriculture14010061