
Effects of Inoculation with Plant Growth-Promoting Rhizobacteria on Chemical Composition of the Substrate and Nutrient Content in Strawberry Plants Growing in Different Water Conditions

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of the Experiment and Plant Material and Growth Conditions Employed
2.2. Experimental Factors
2.3. Measurement Methods
2.4. Statistical Methods
3. Results
3.1. Physicochemical Parameters of the Substrate
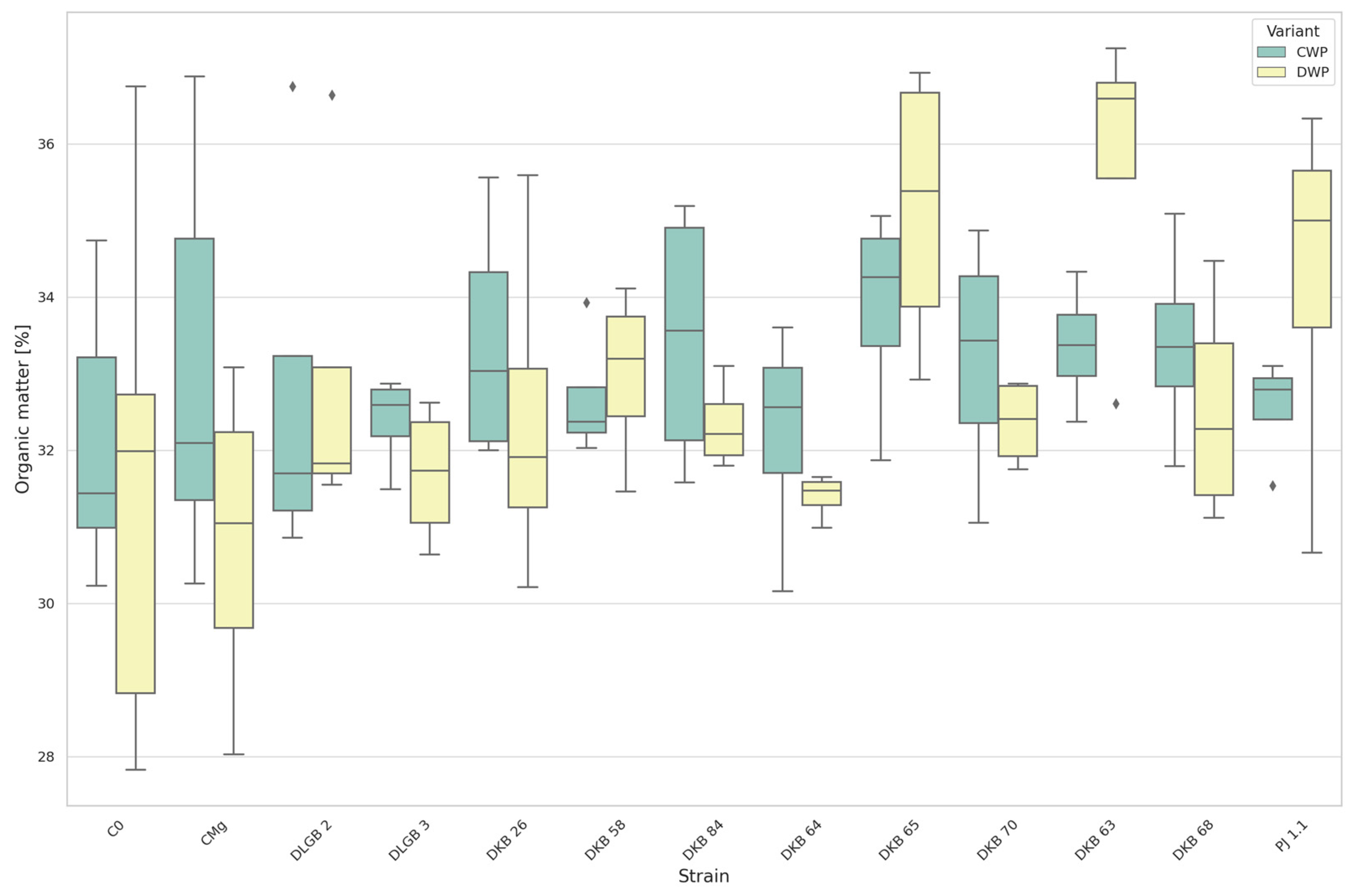
3.1.1. The Content of Organic Matter in Substrate
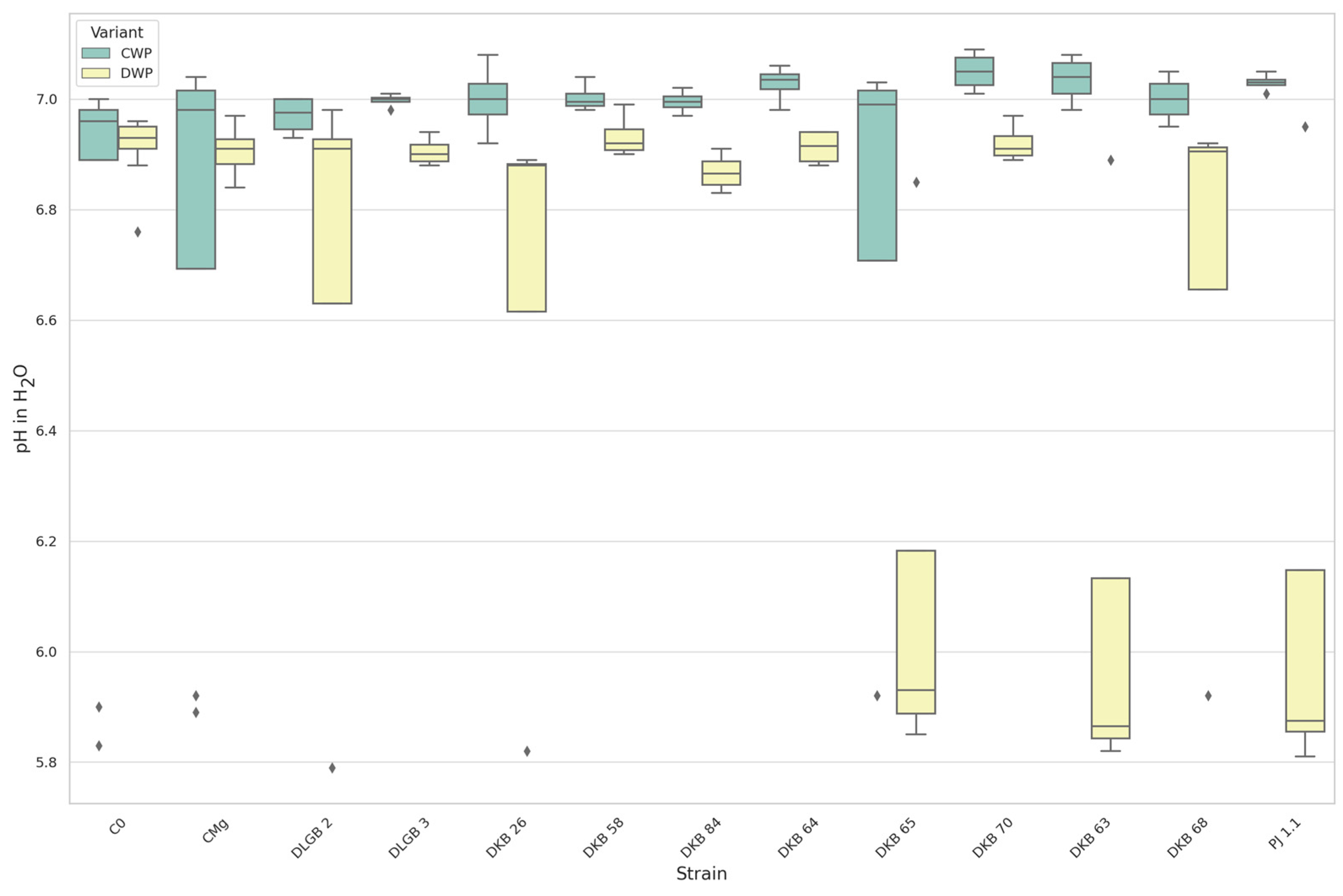
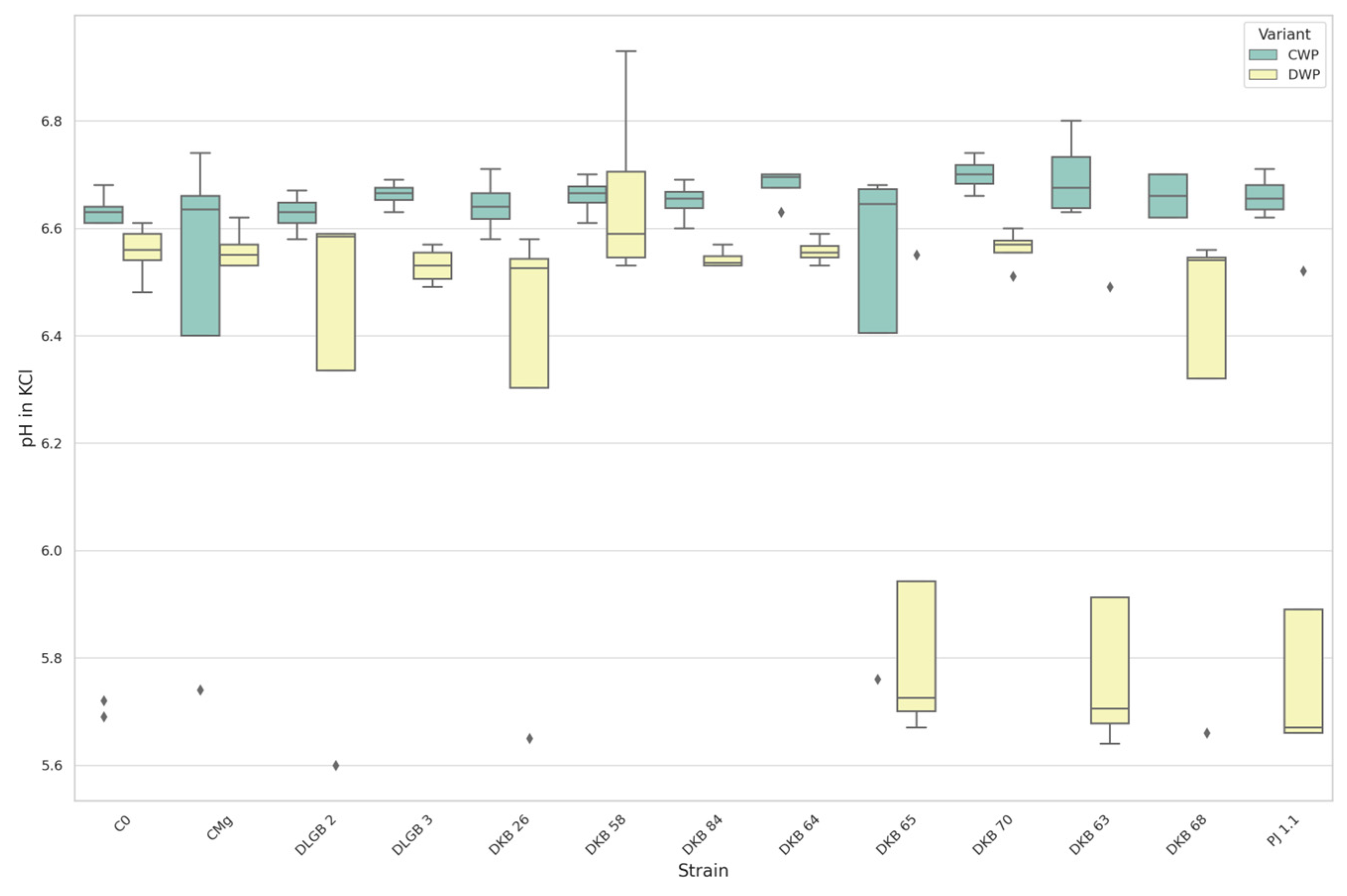
3.1.2. The pH of the Substrate
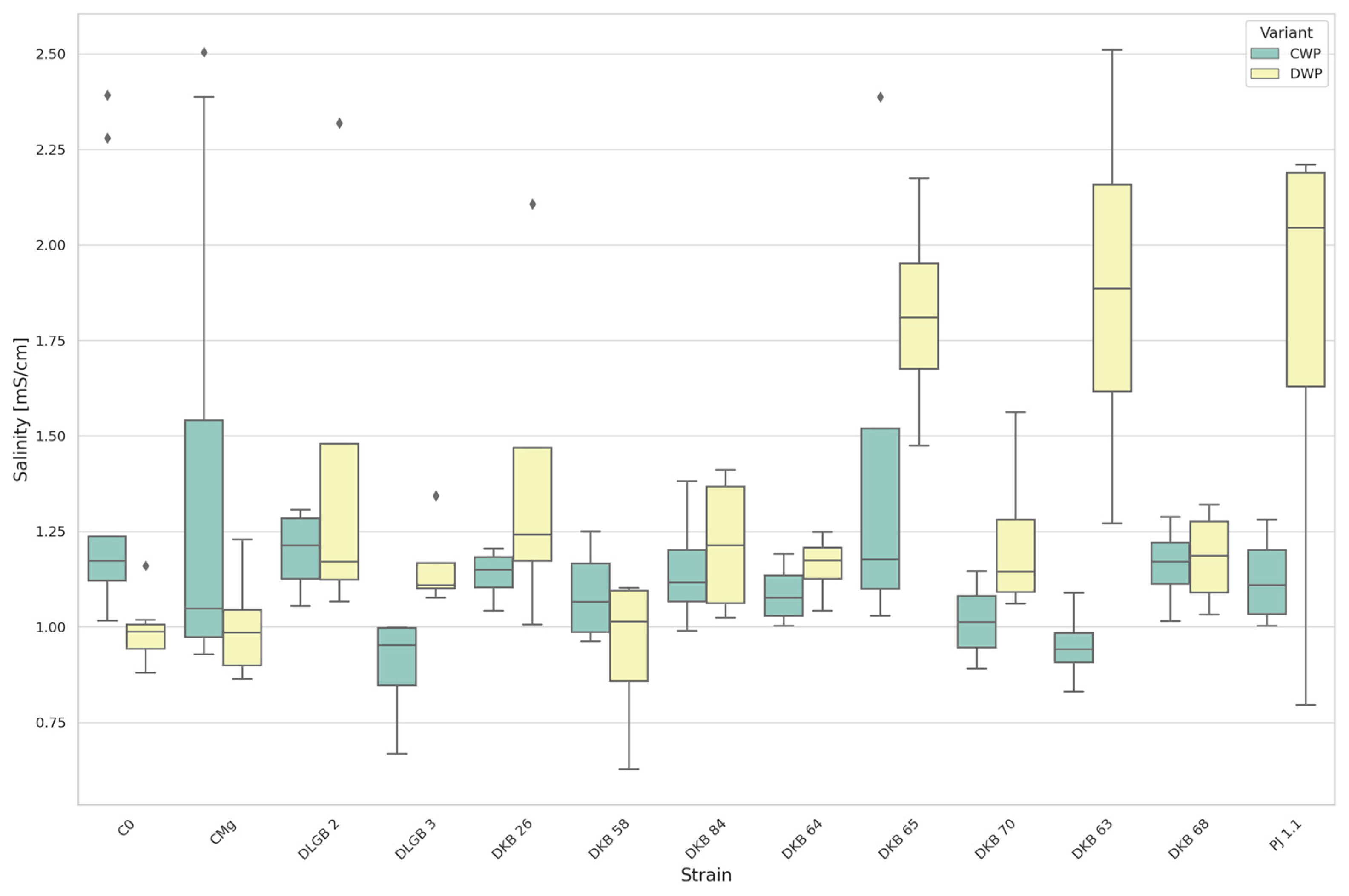
3.1.3. The Salinity of Substrate
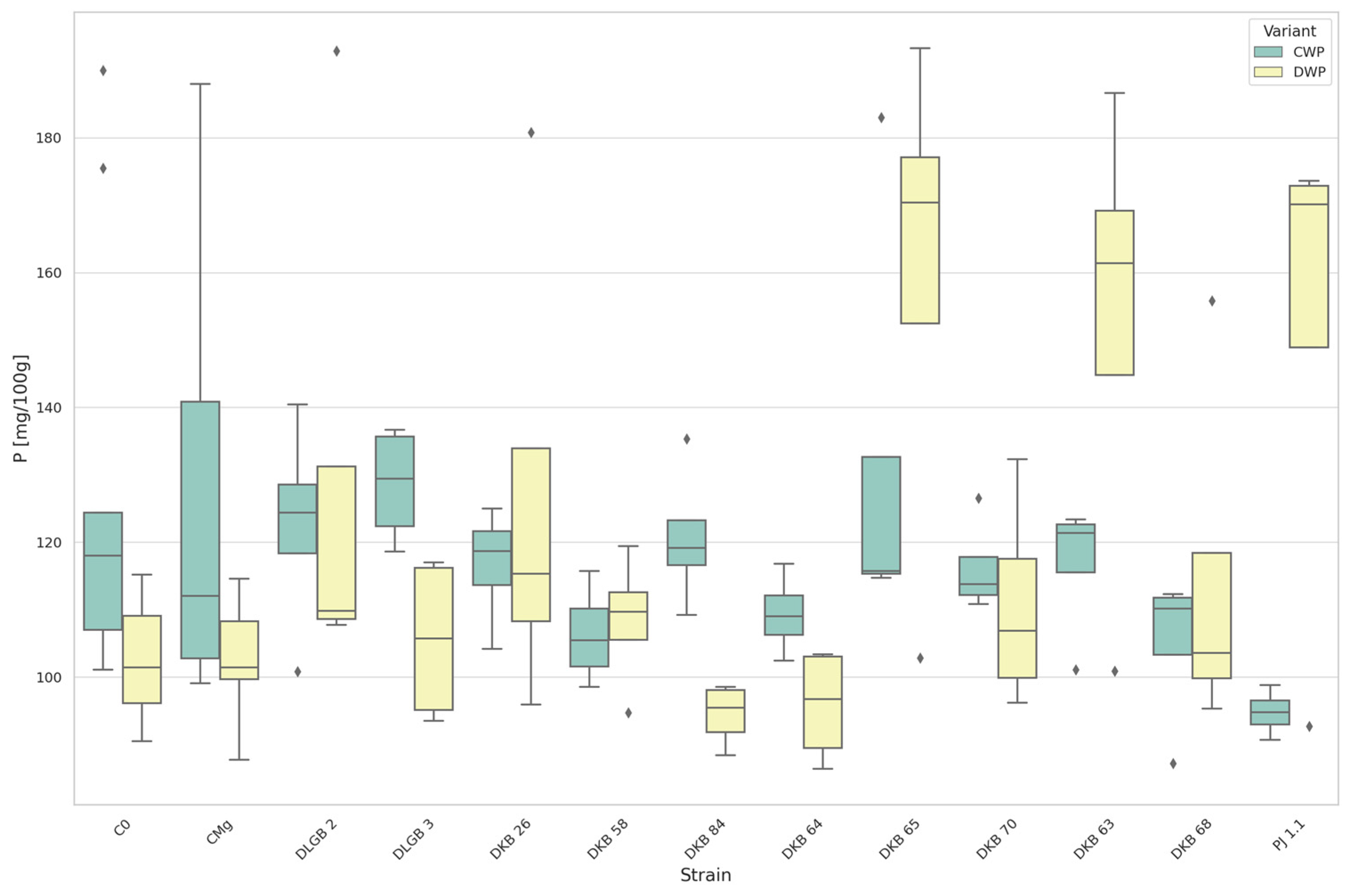
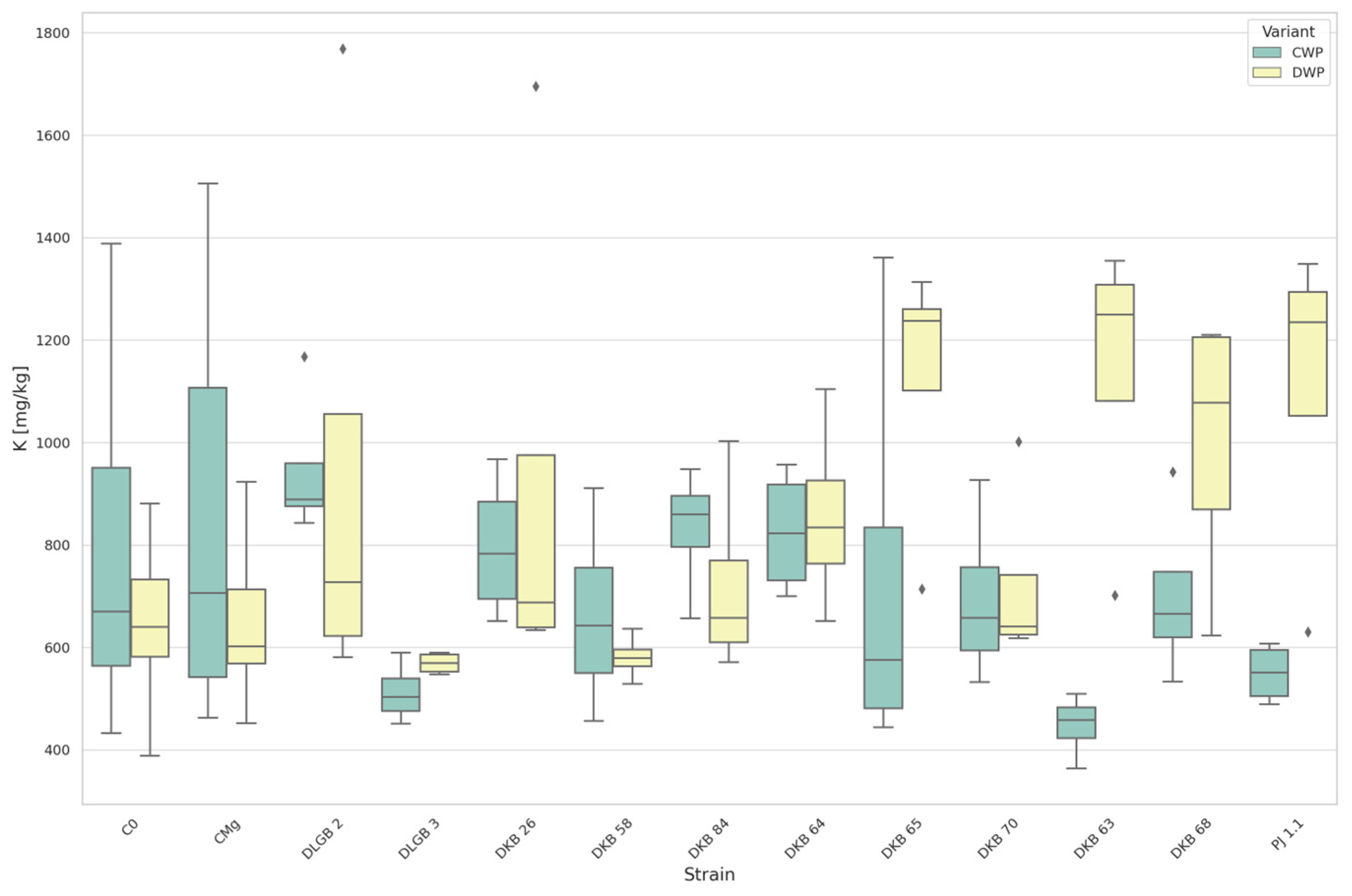
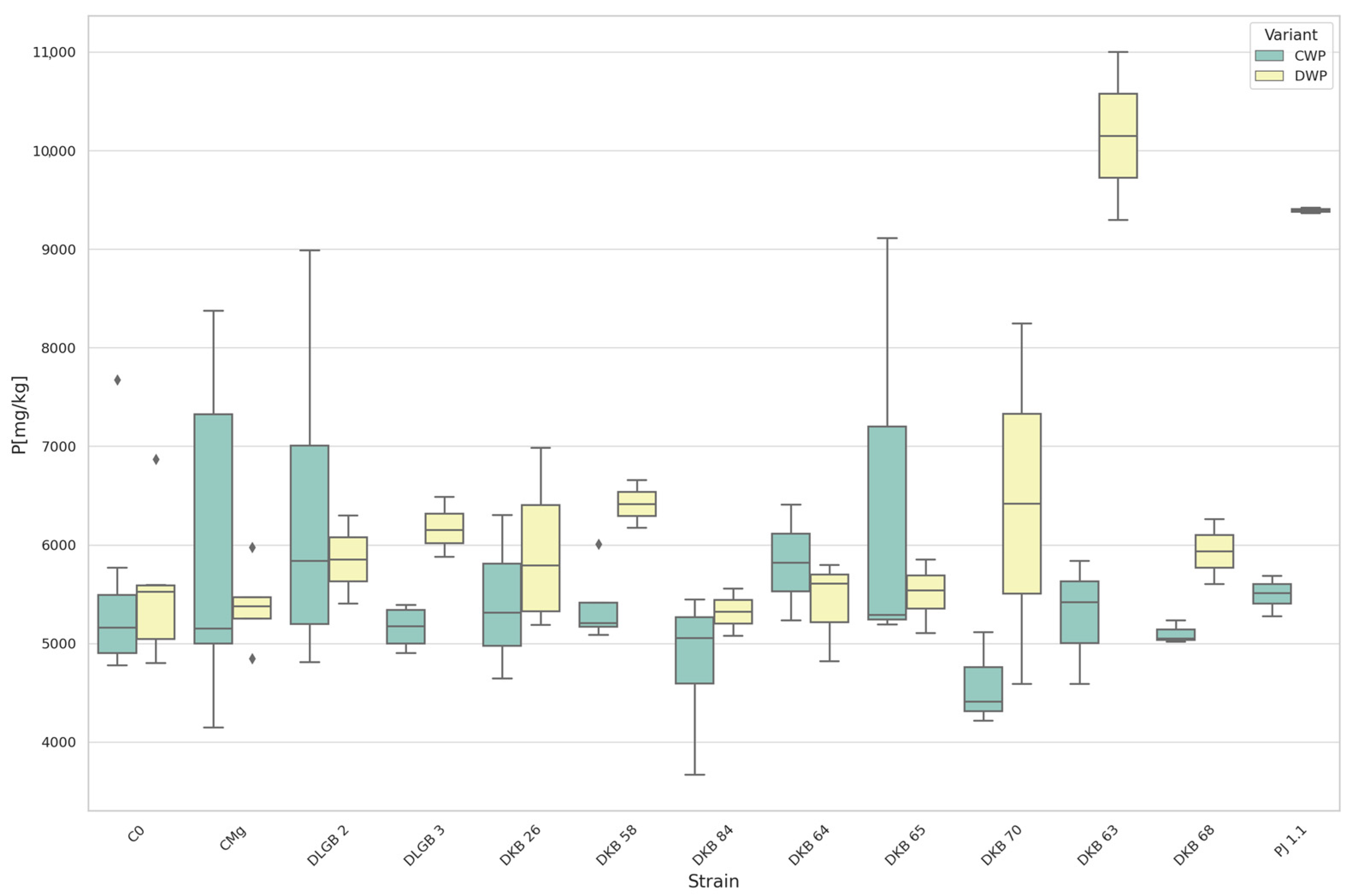
3.2. The Content of Macronutrients in the Substrate
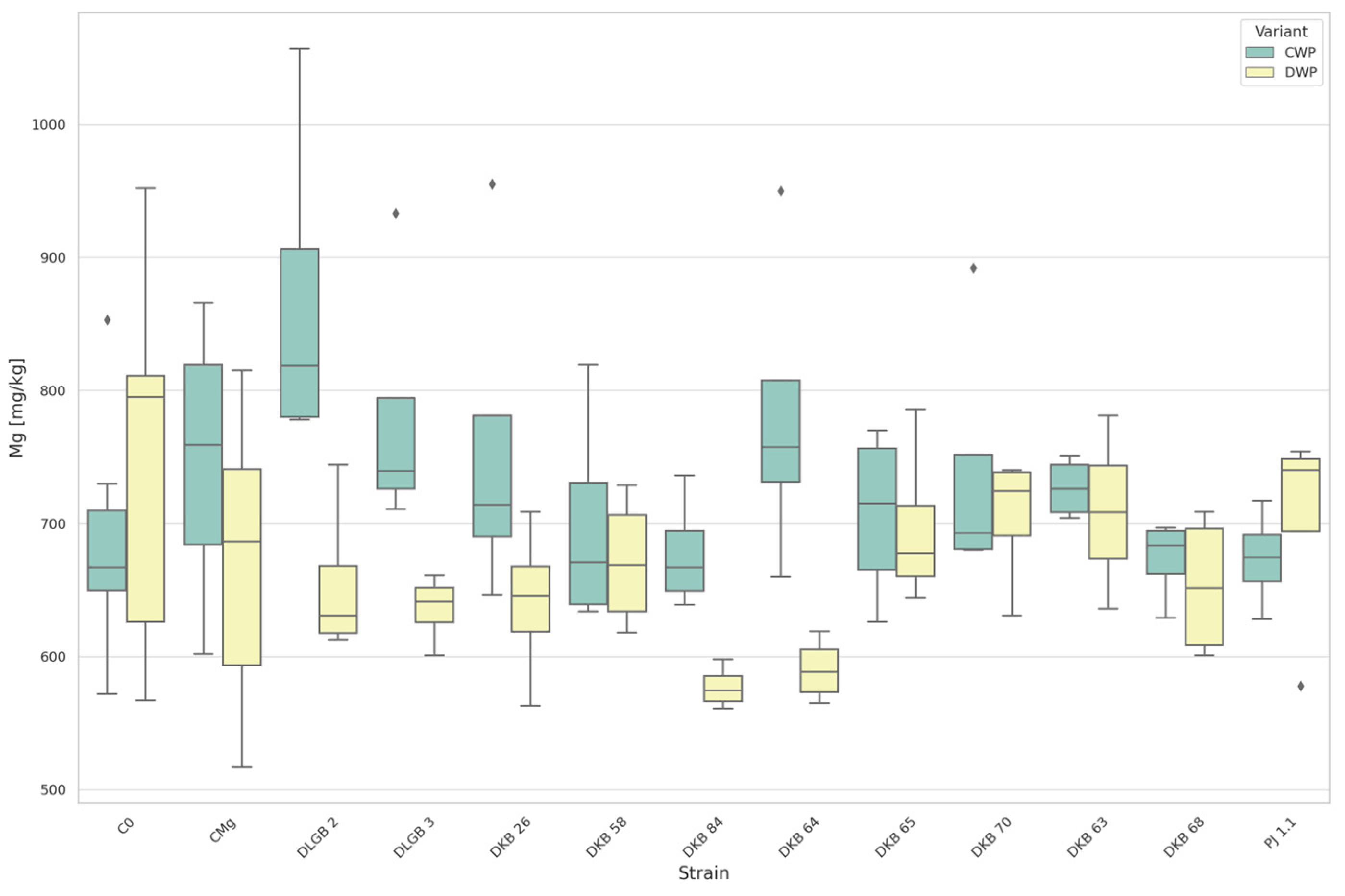
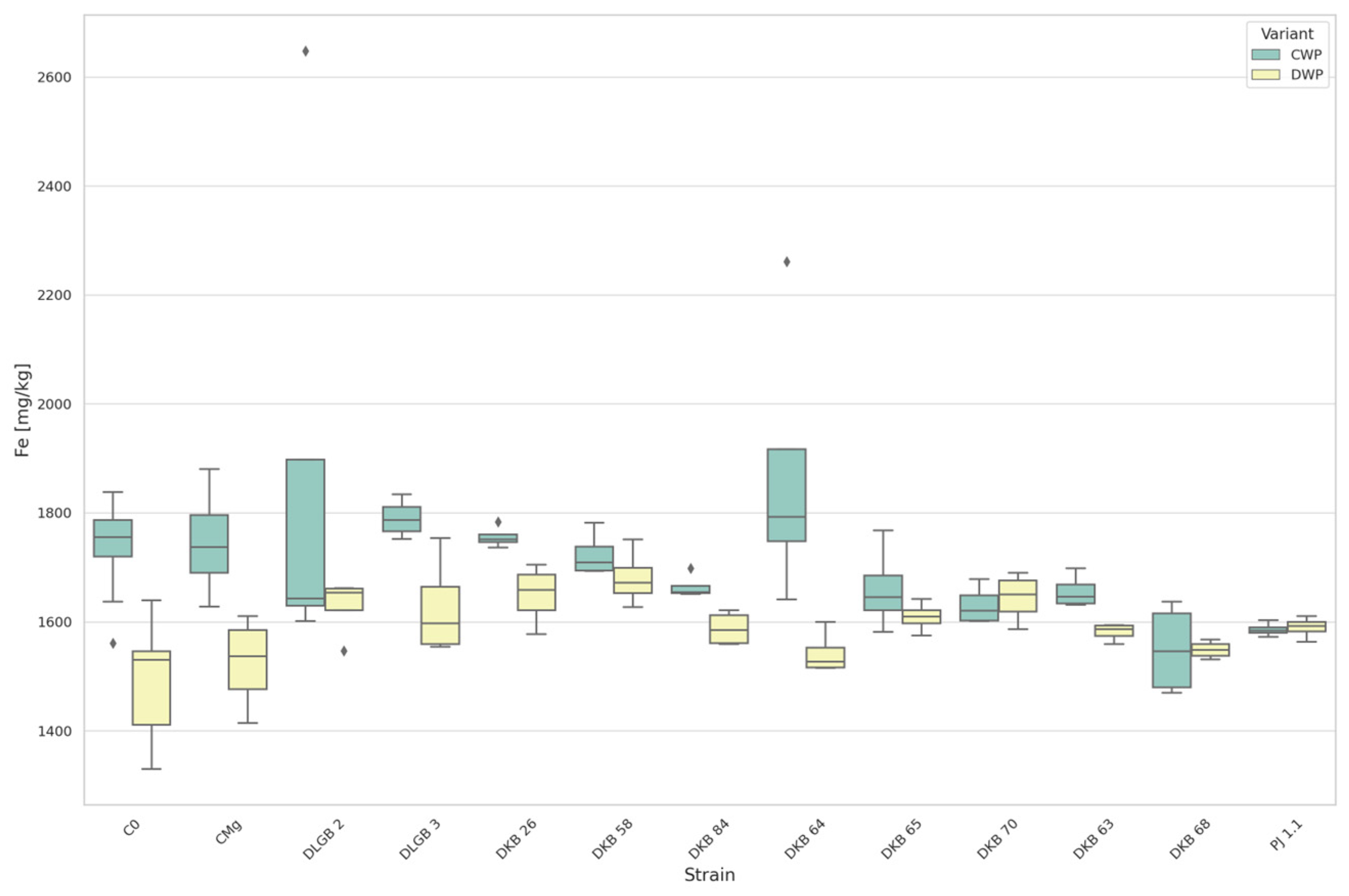
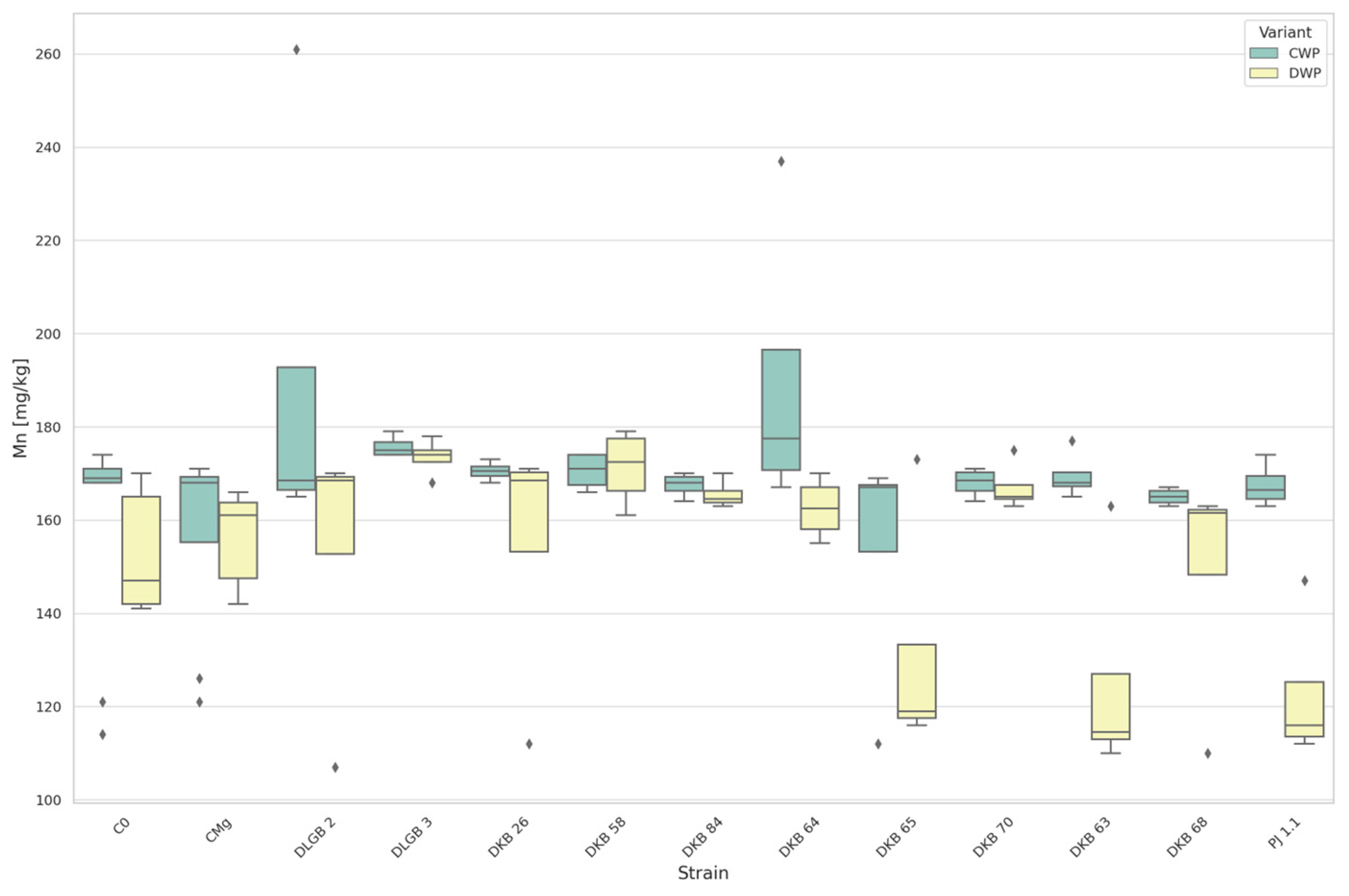
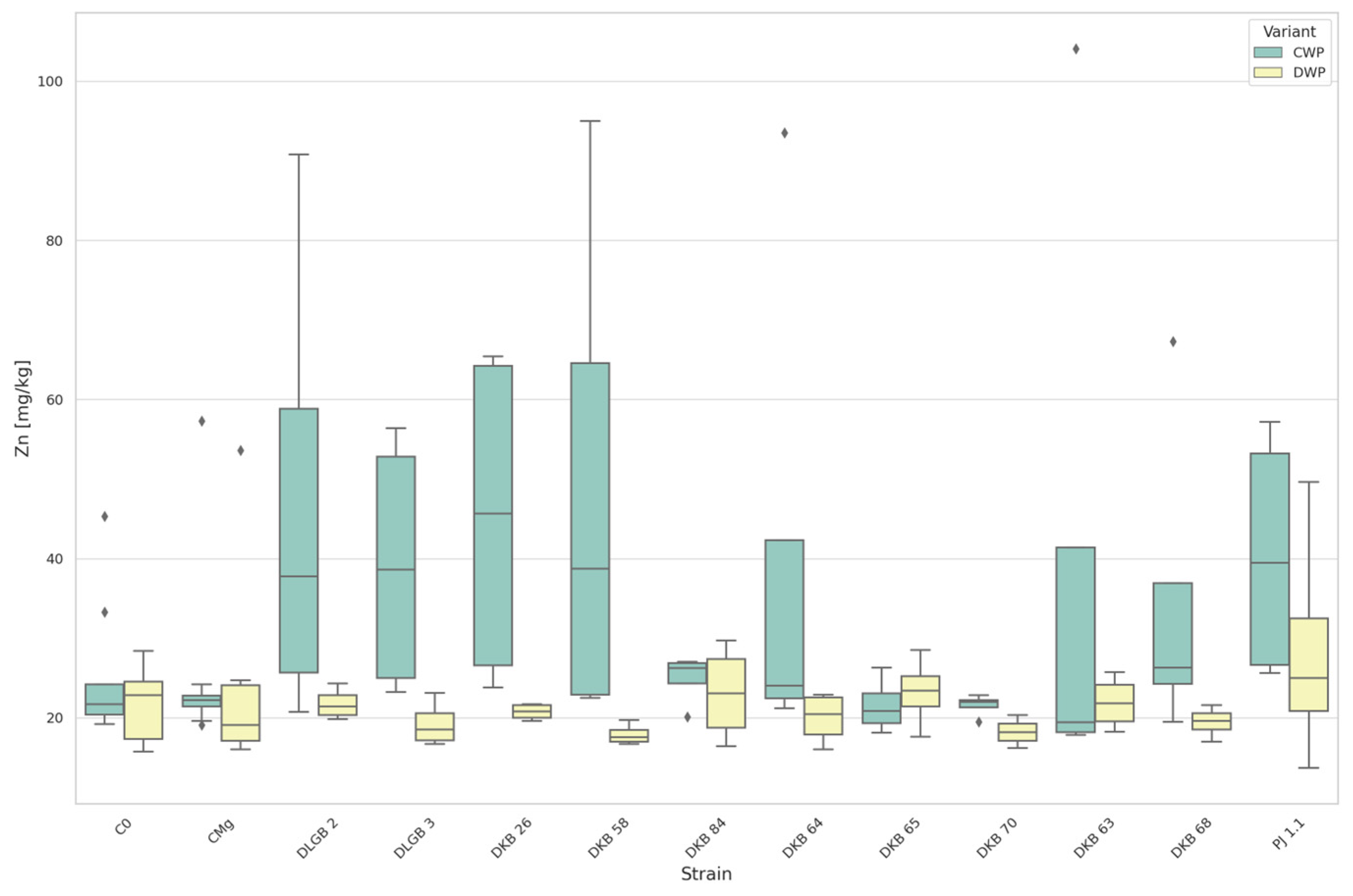
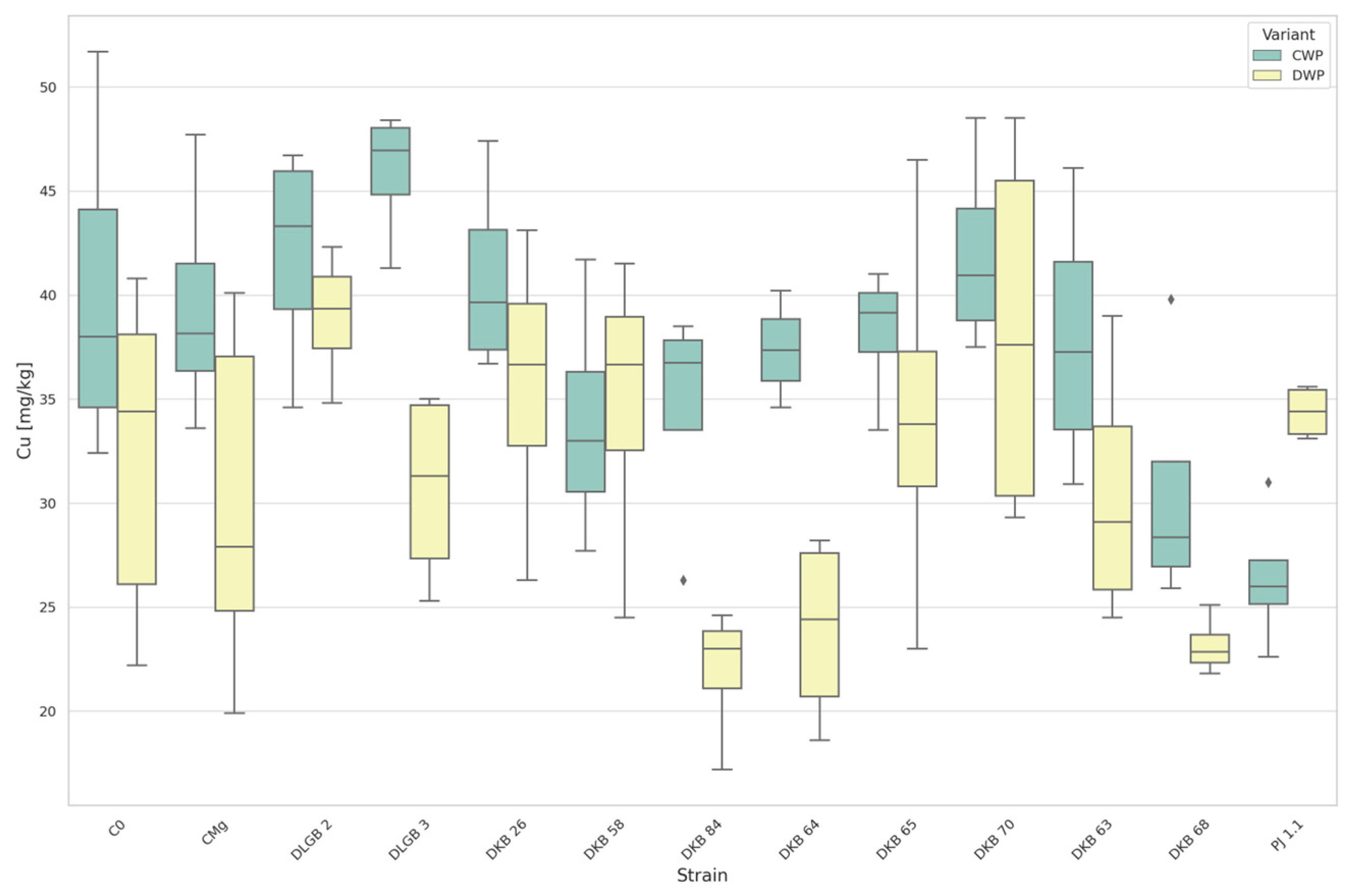
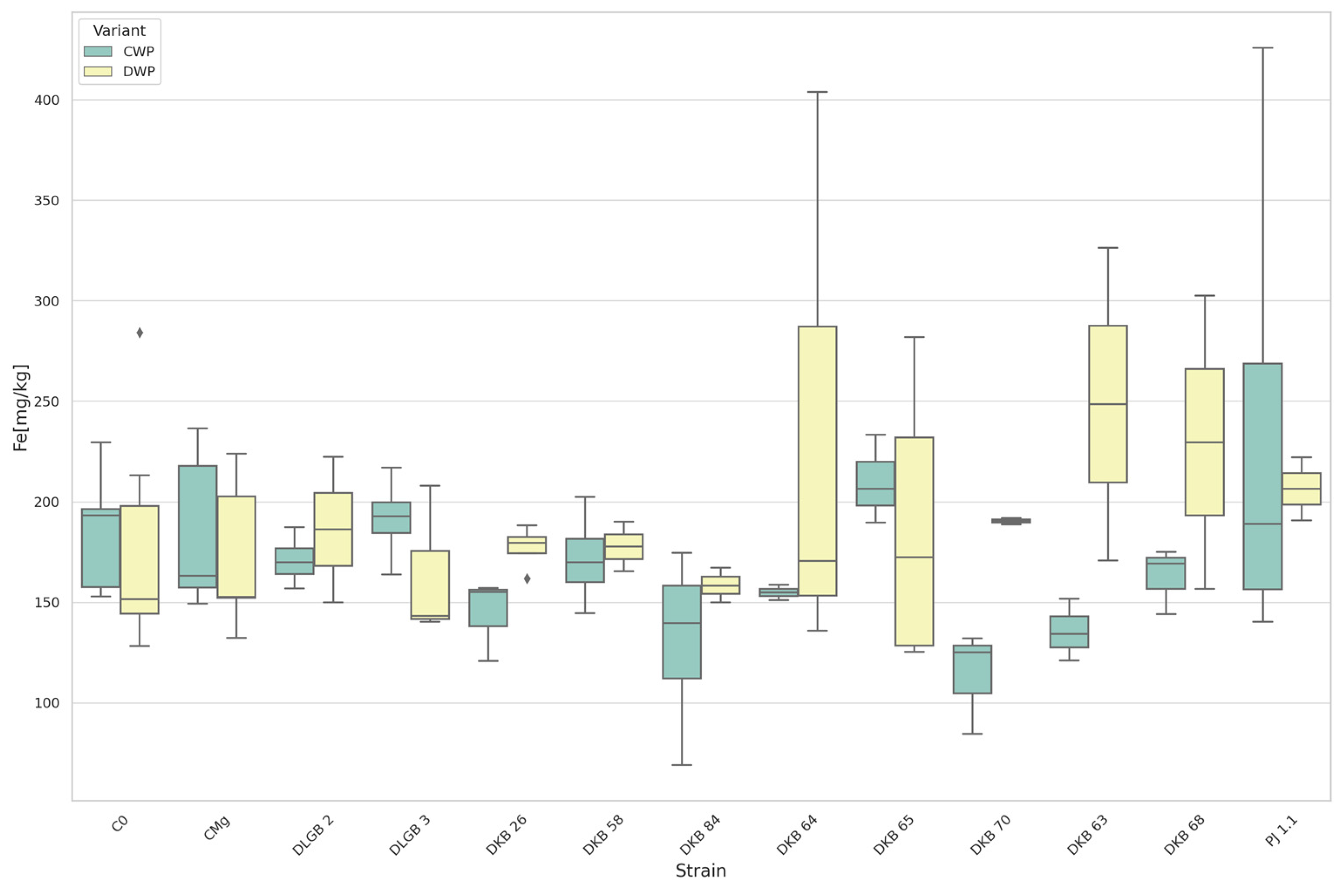
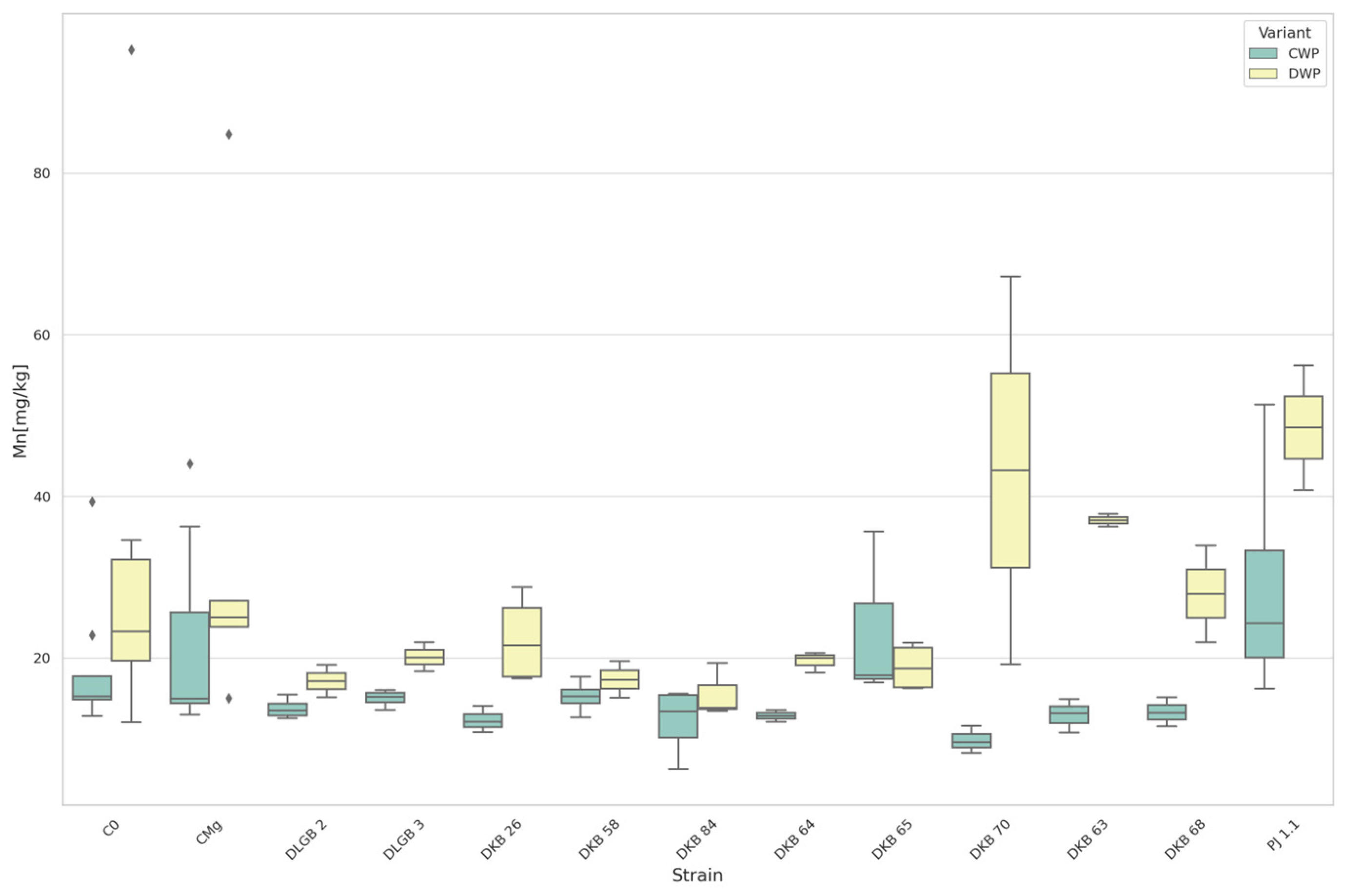
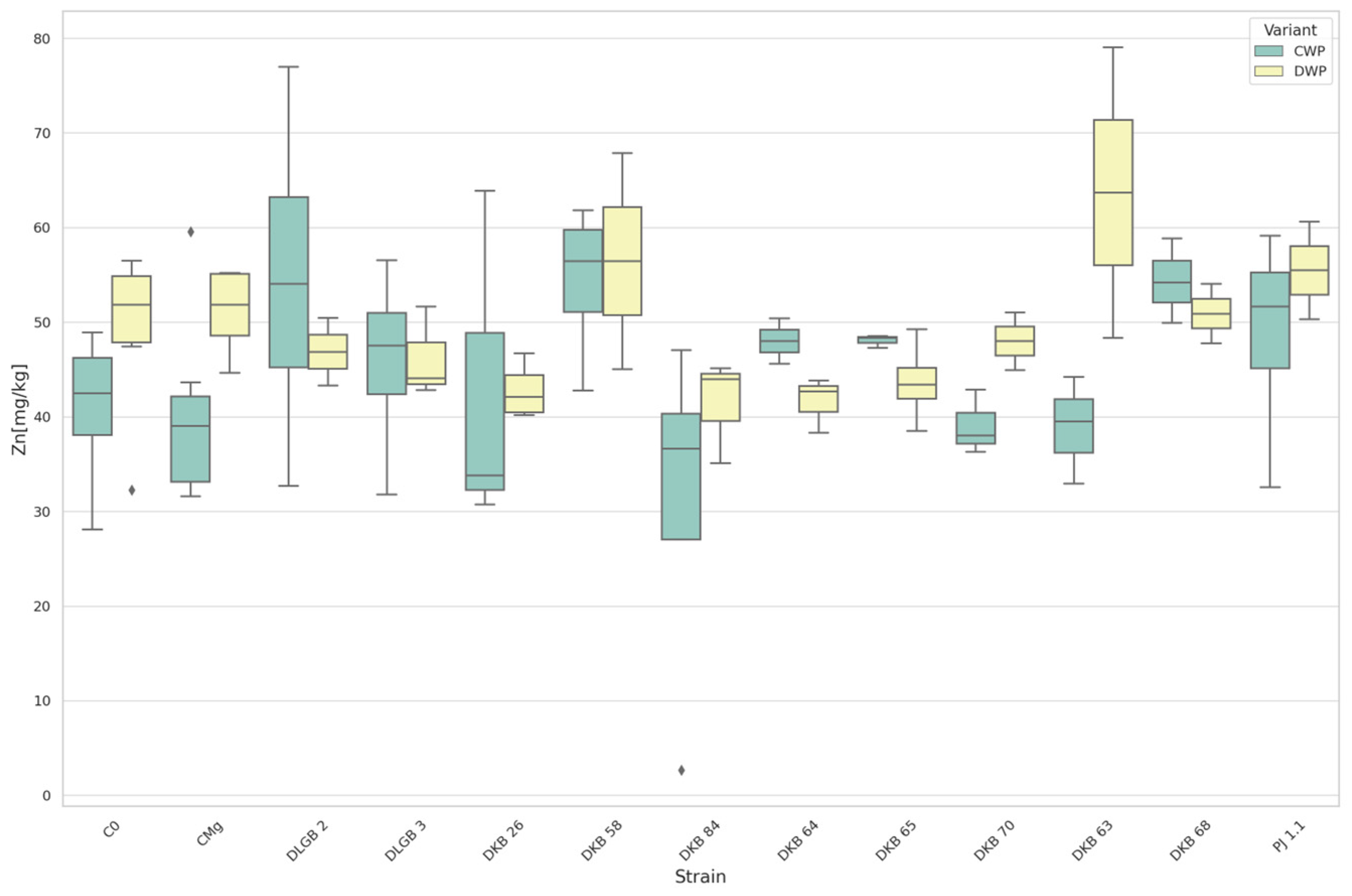
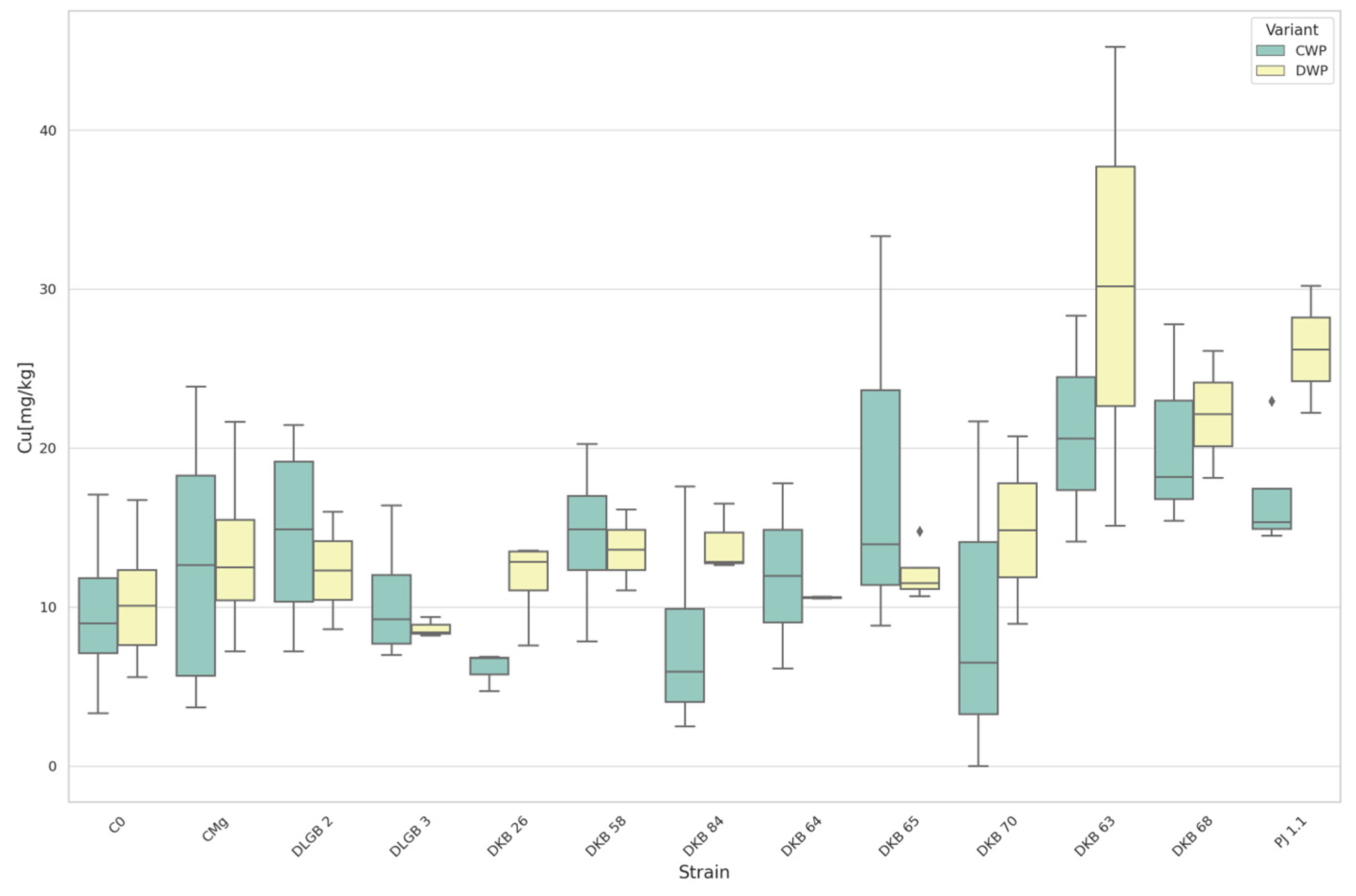
3.3. The Content of Micronutrients in the Substrate
3.4. Summary Analysis Effect Substrate
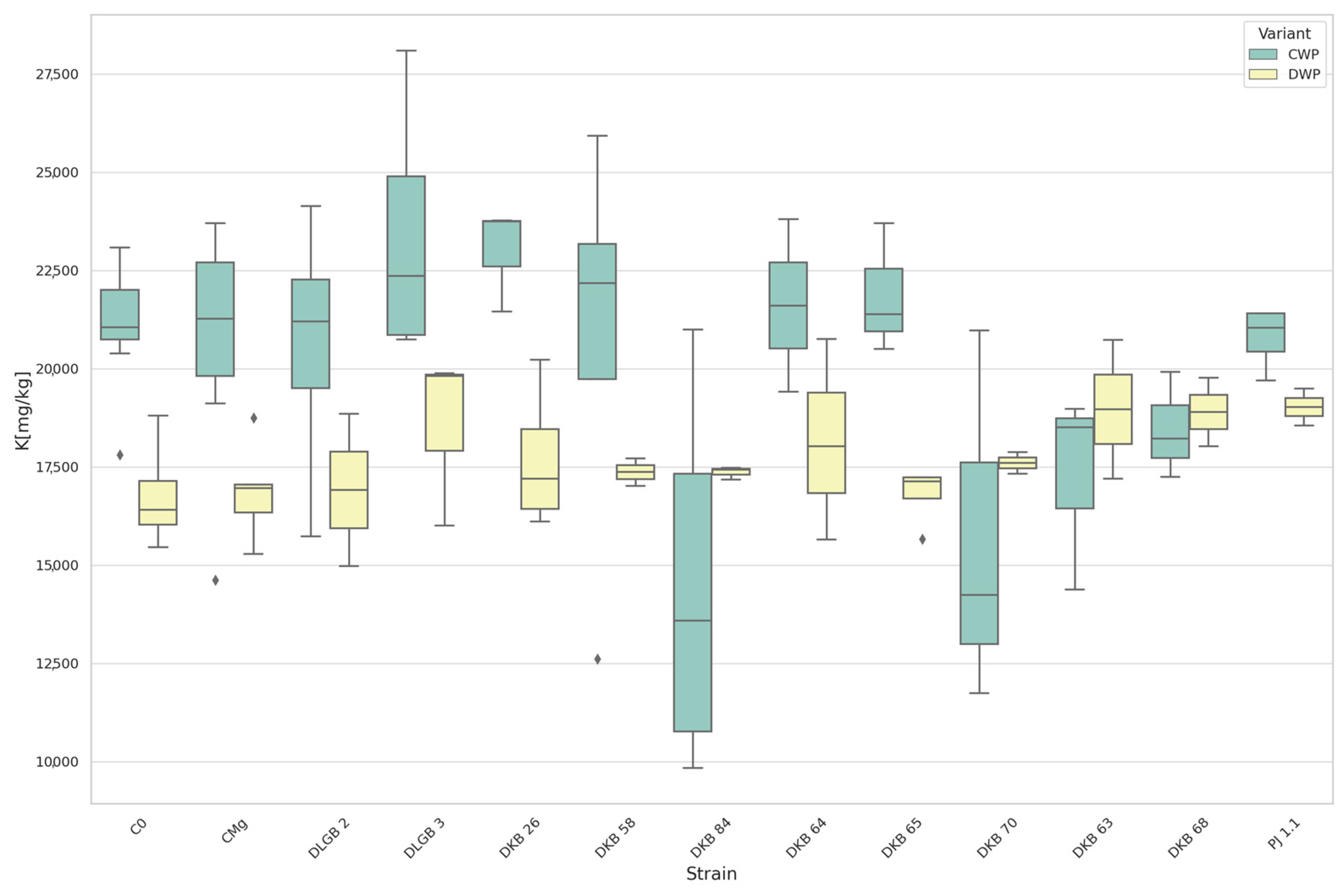
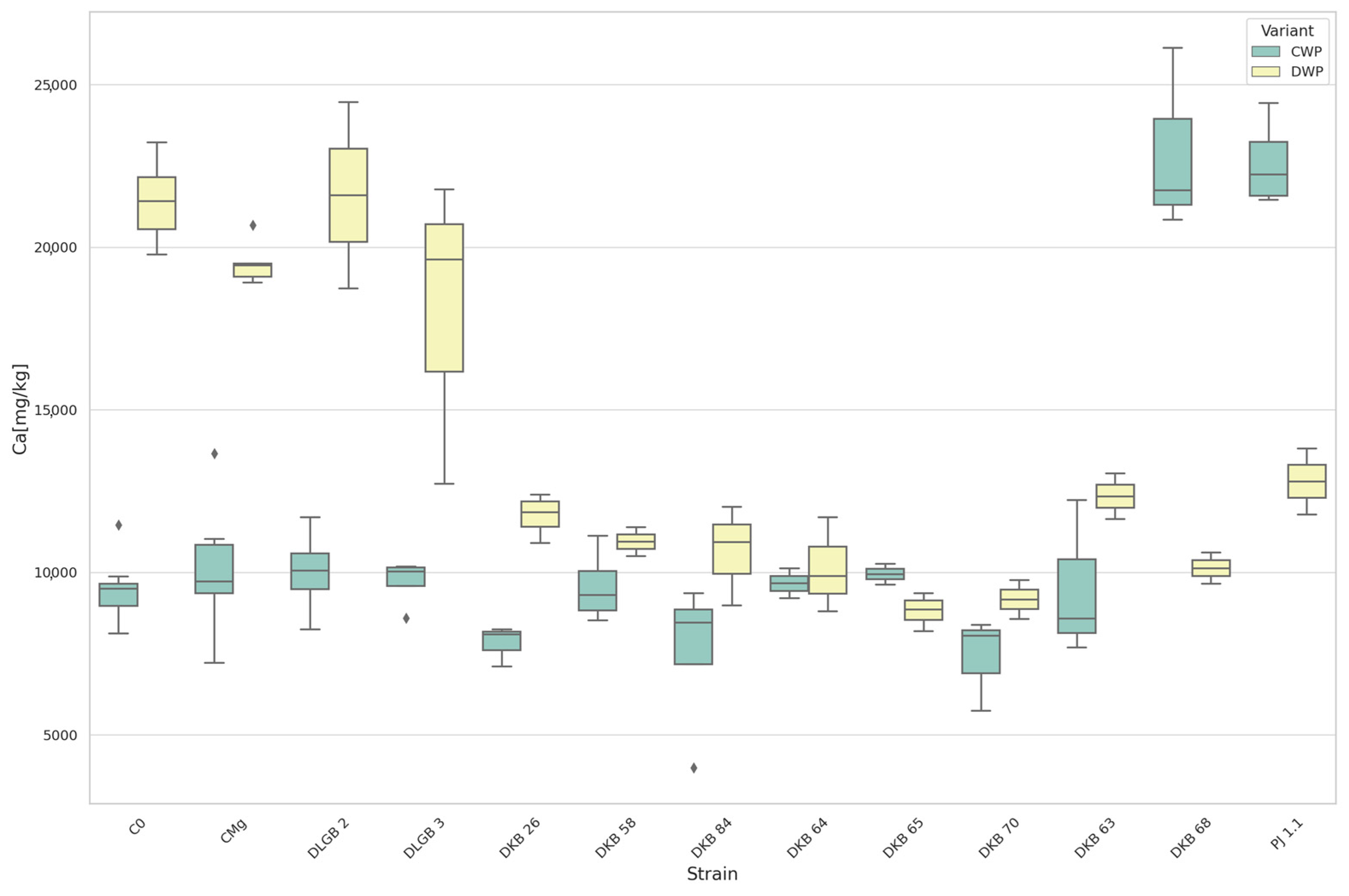
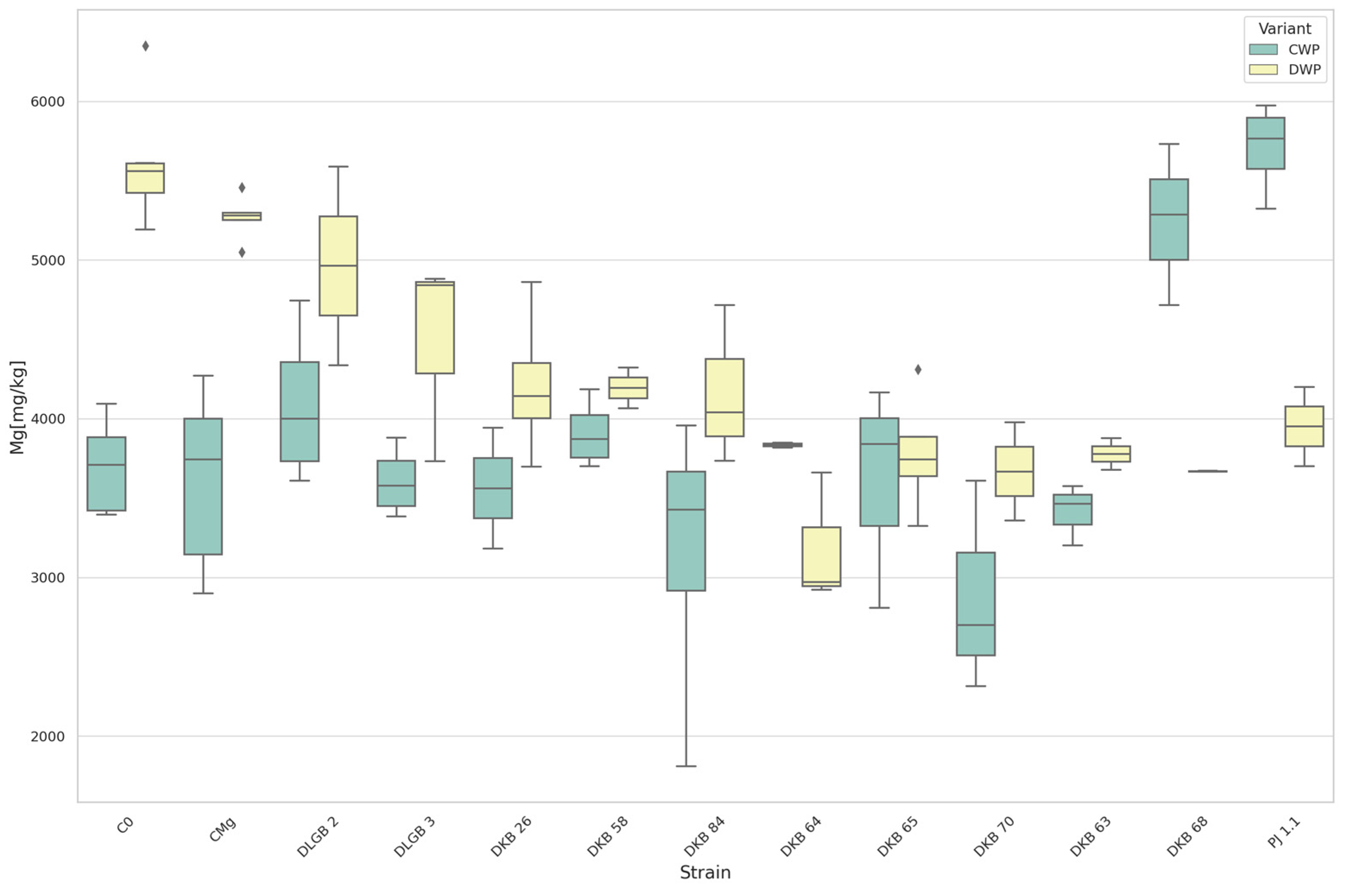
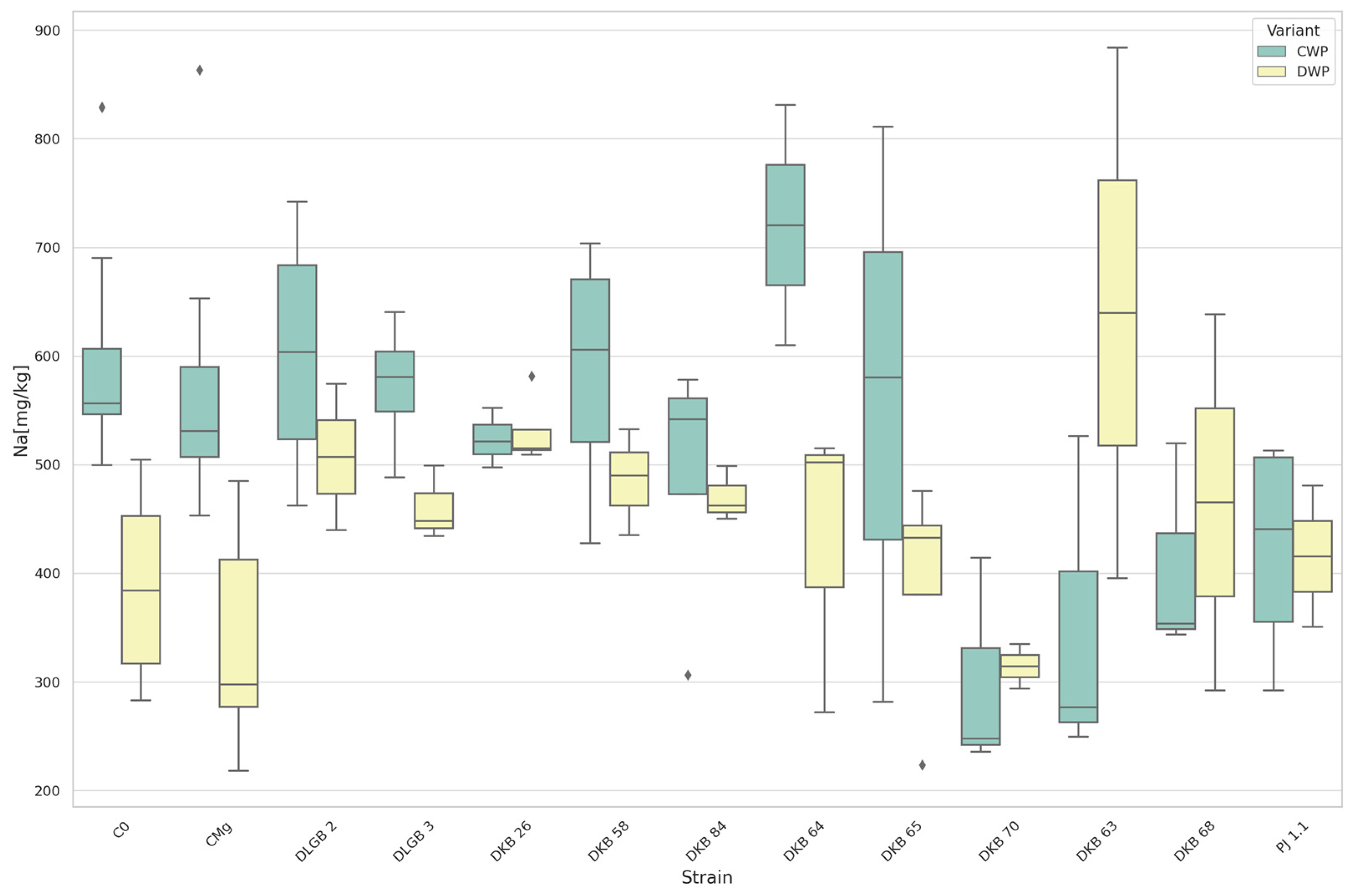
3.5. The Content of Macronutrients and Sodium in Strawberry Leaves
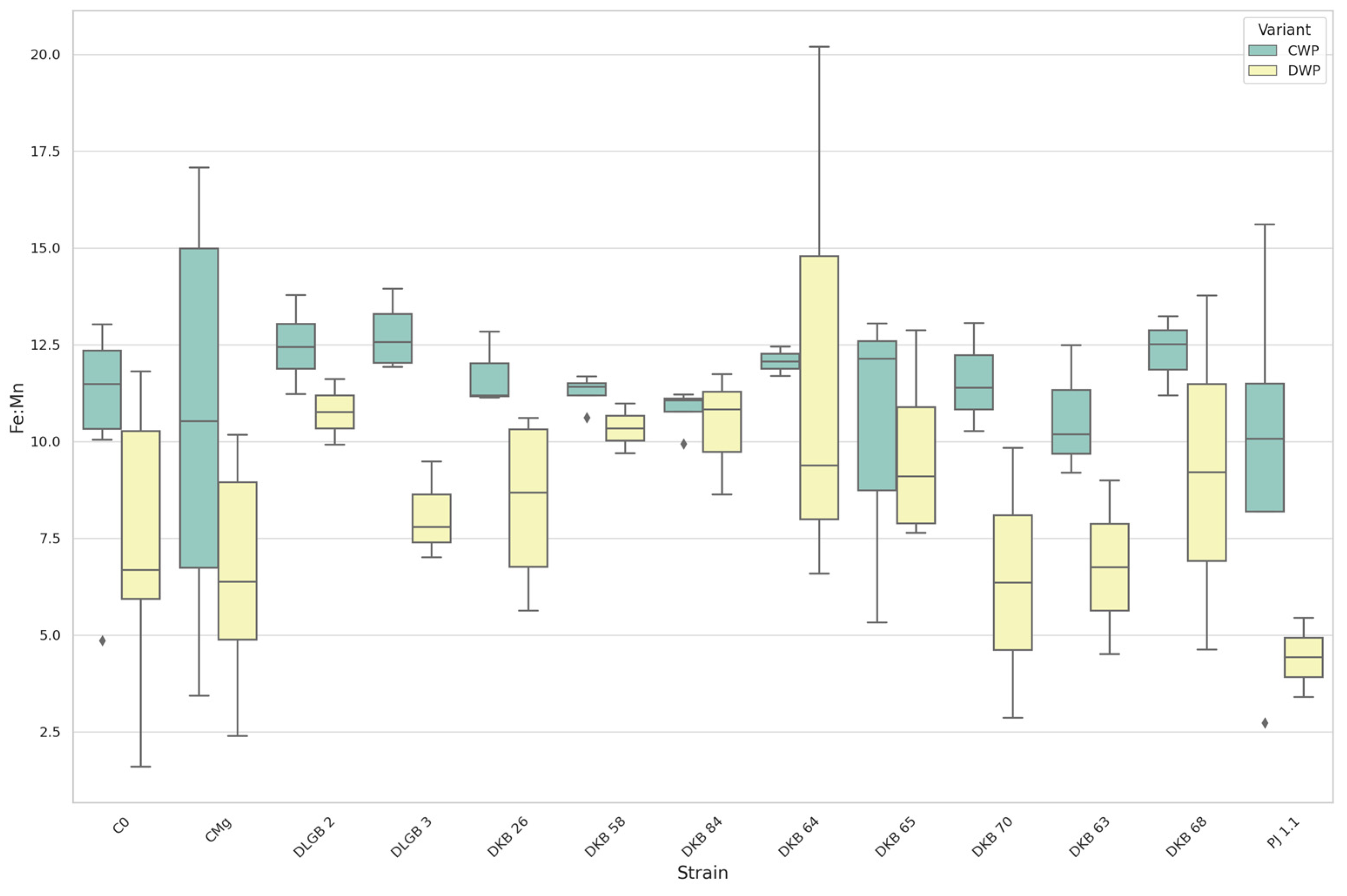
3.6. The Content of Micronutrients in Strawberry Leaves
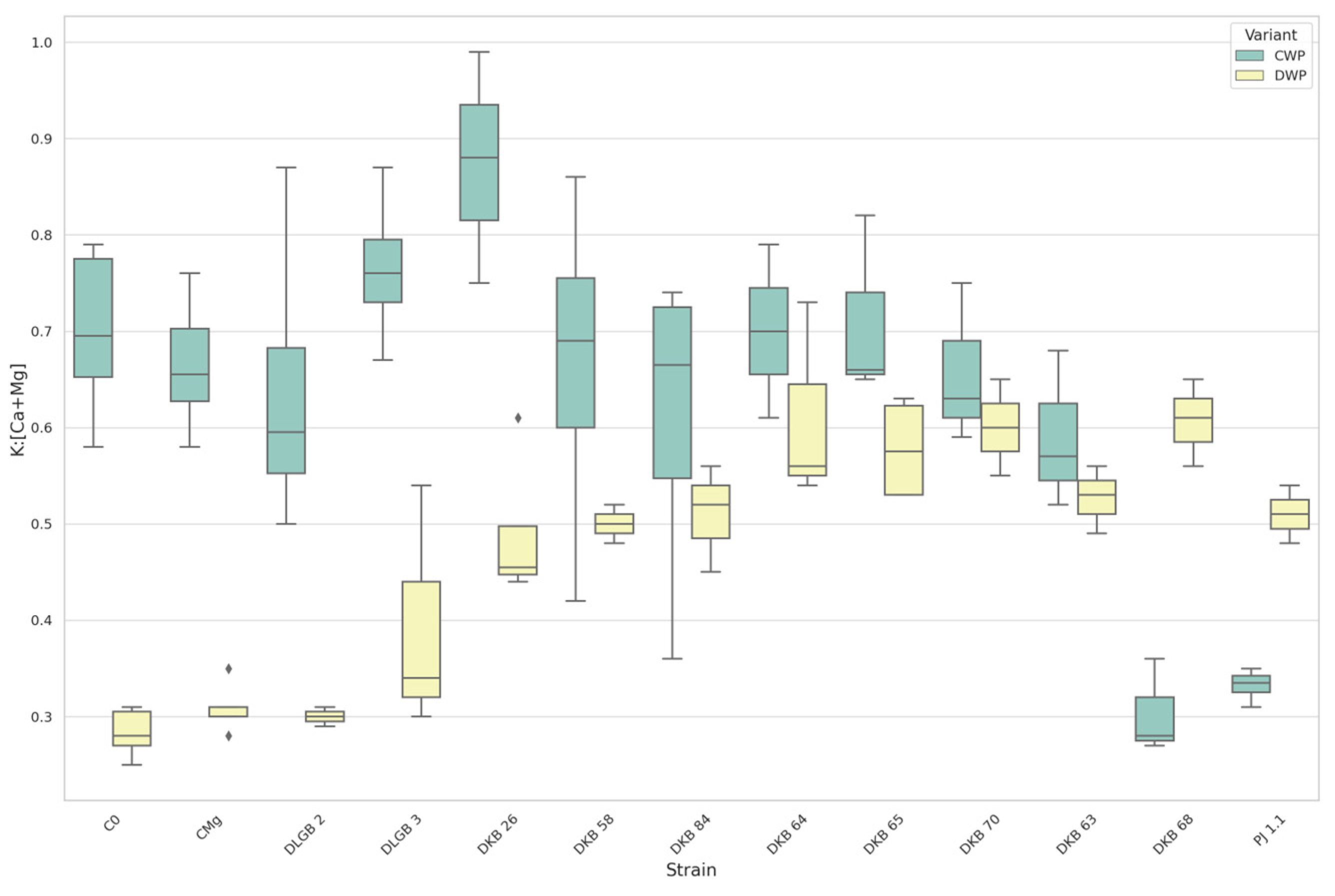
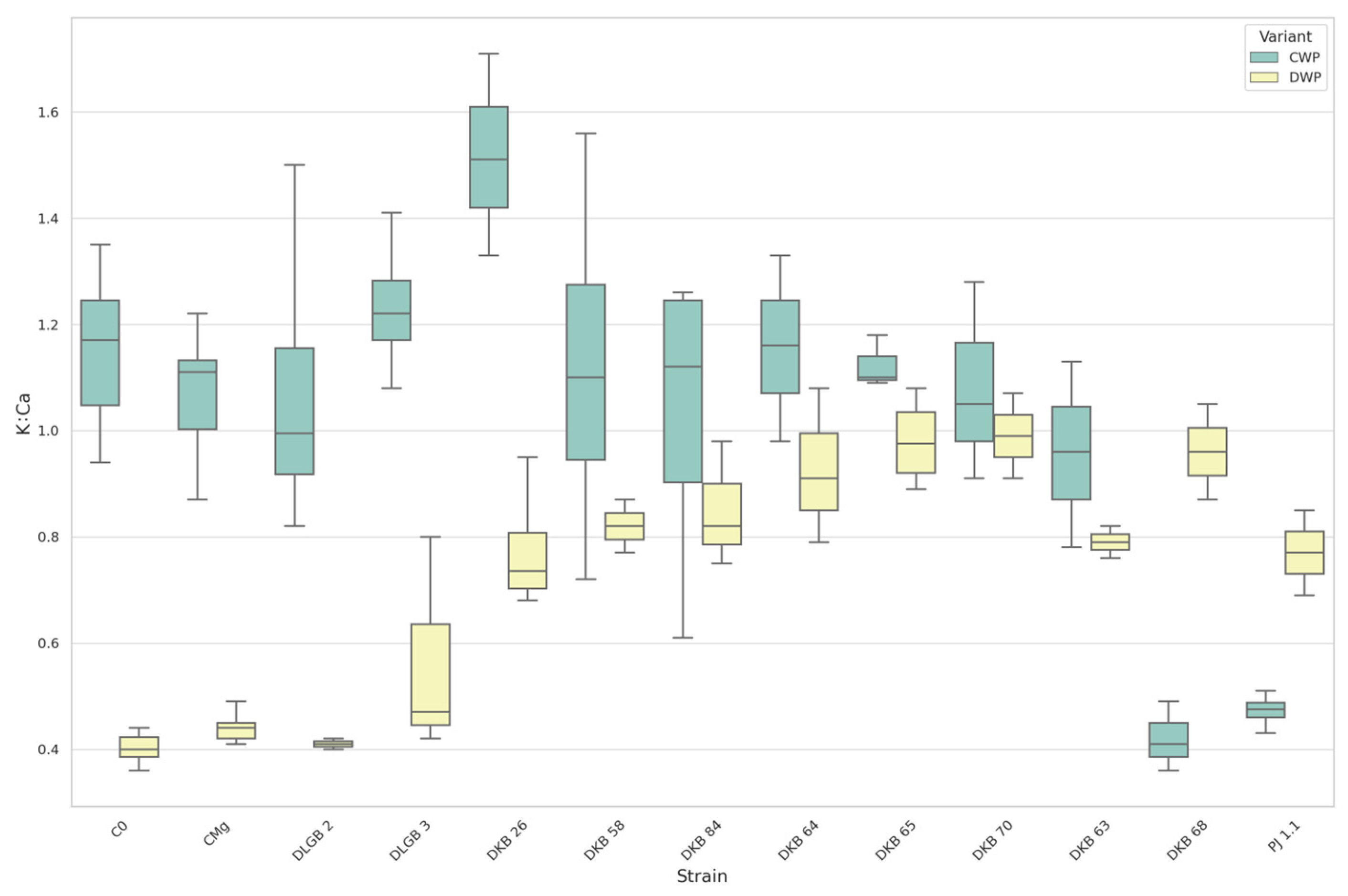
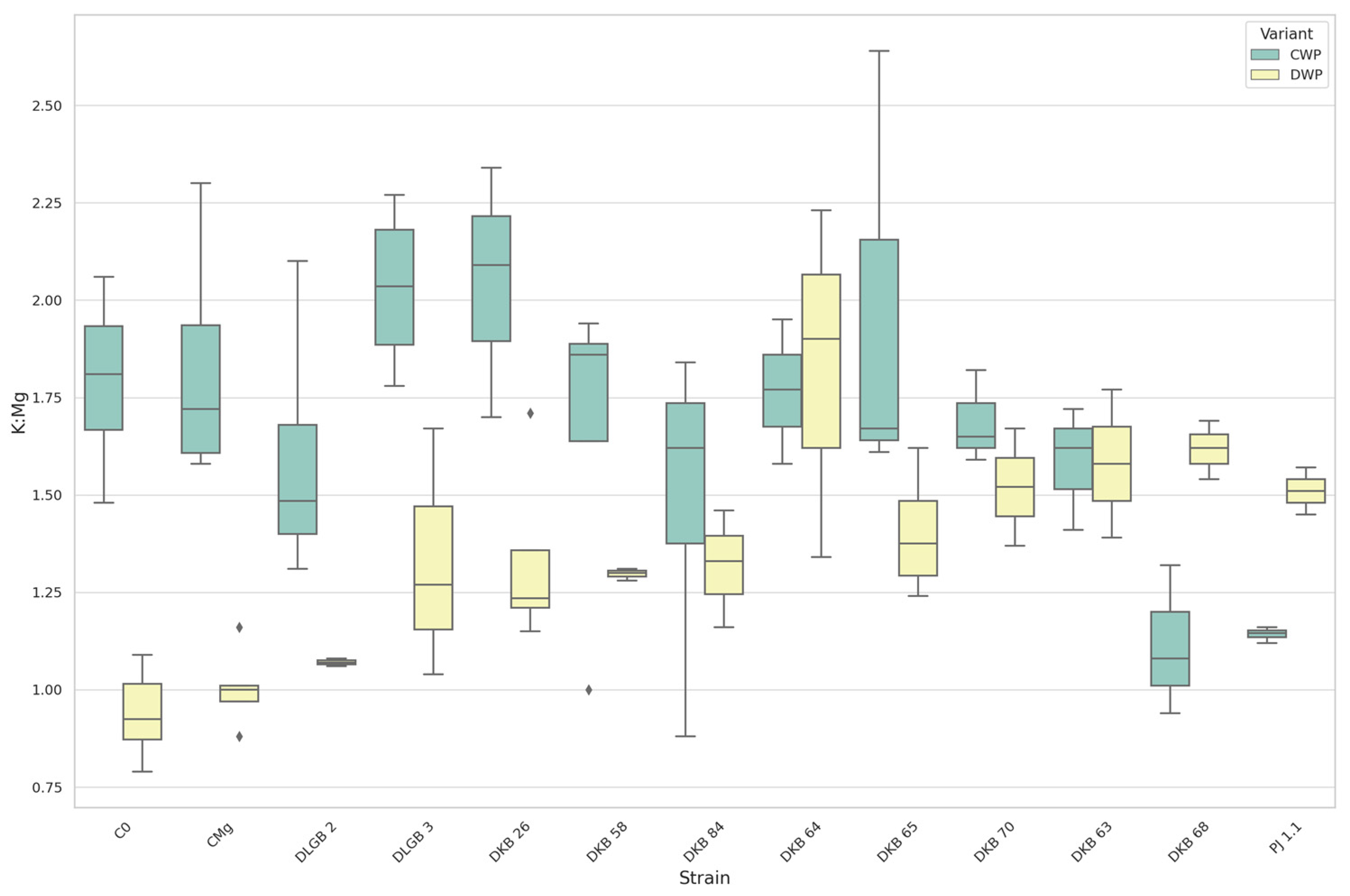
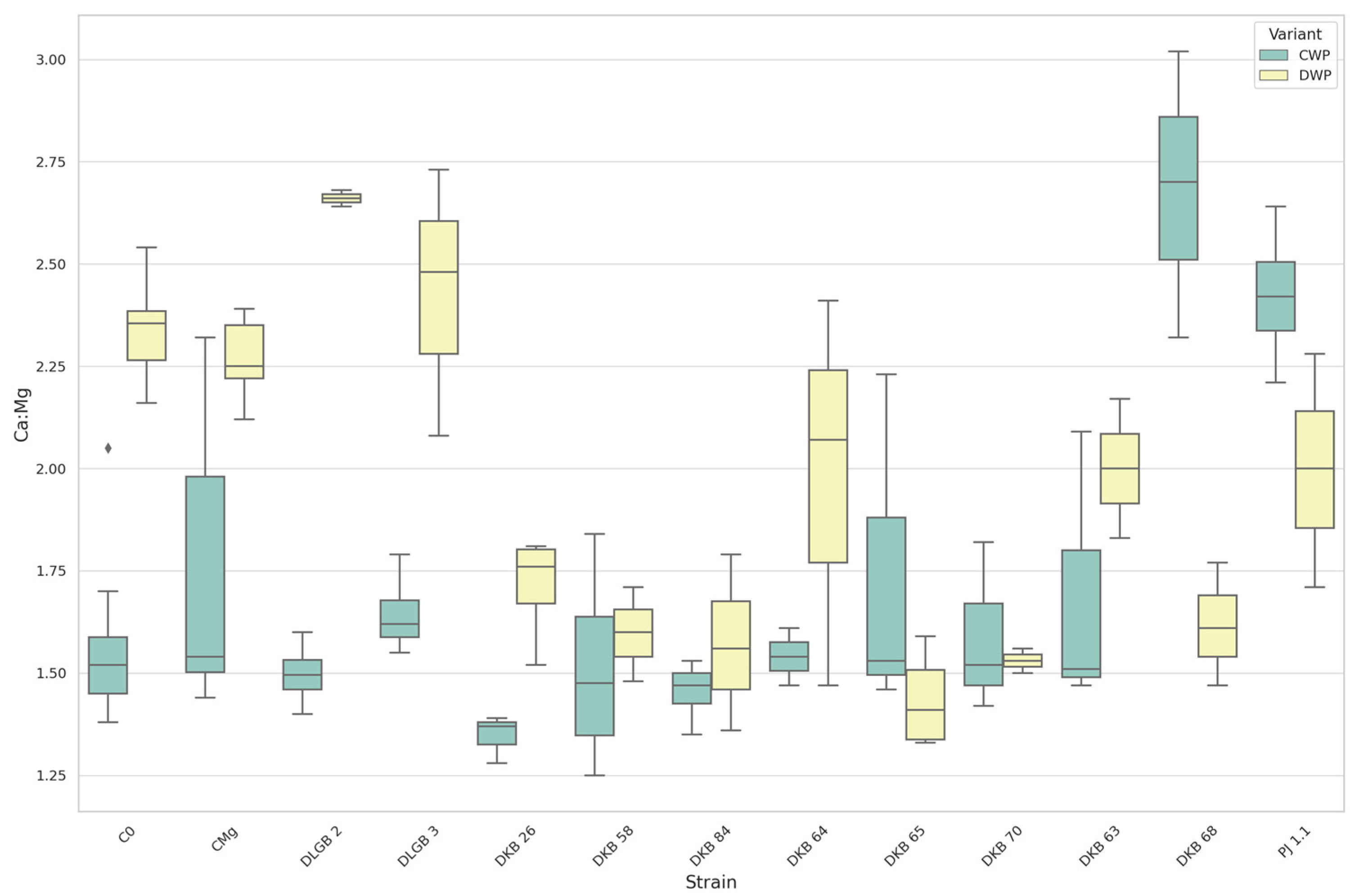
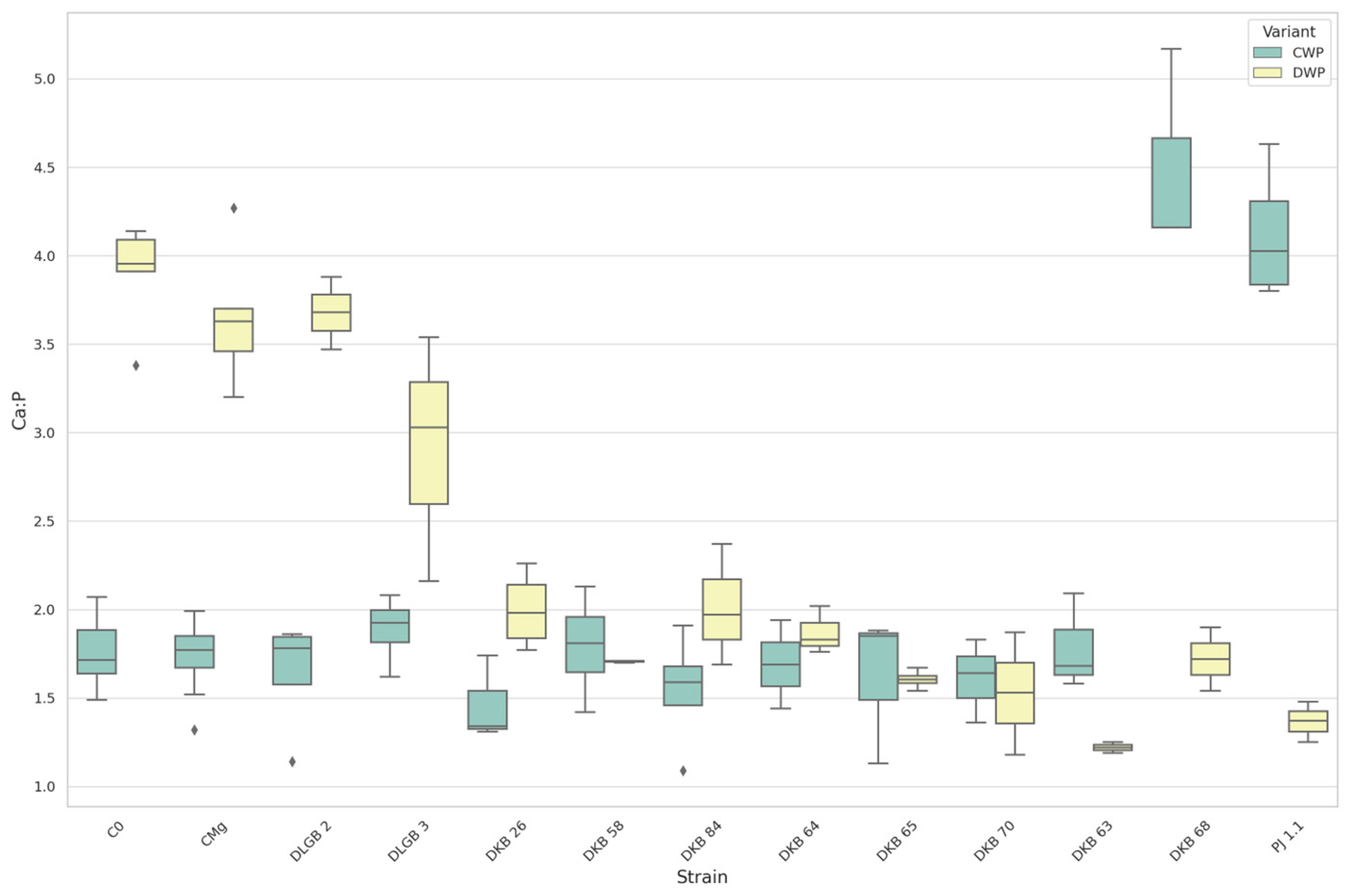
3.7. The Leaf Nutrients Ratios
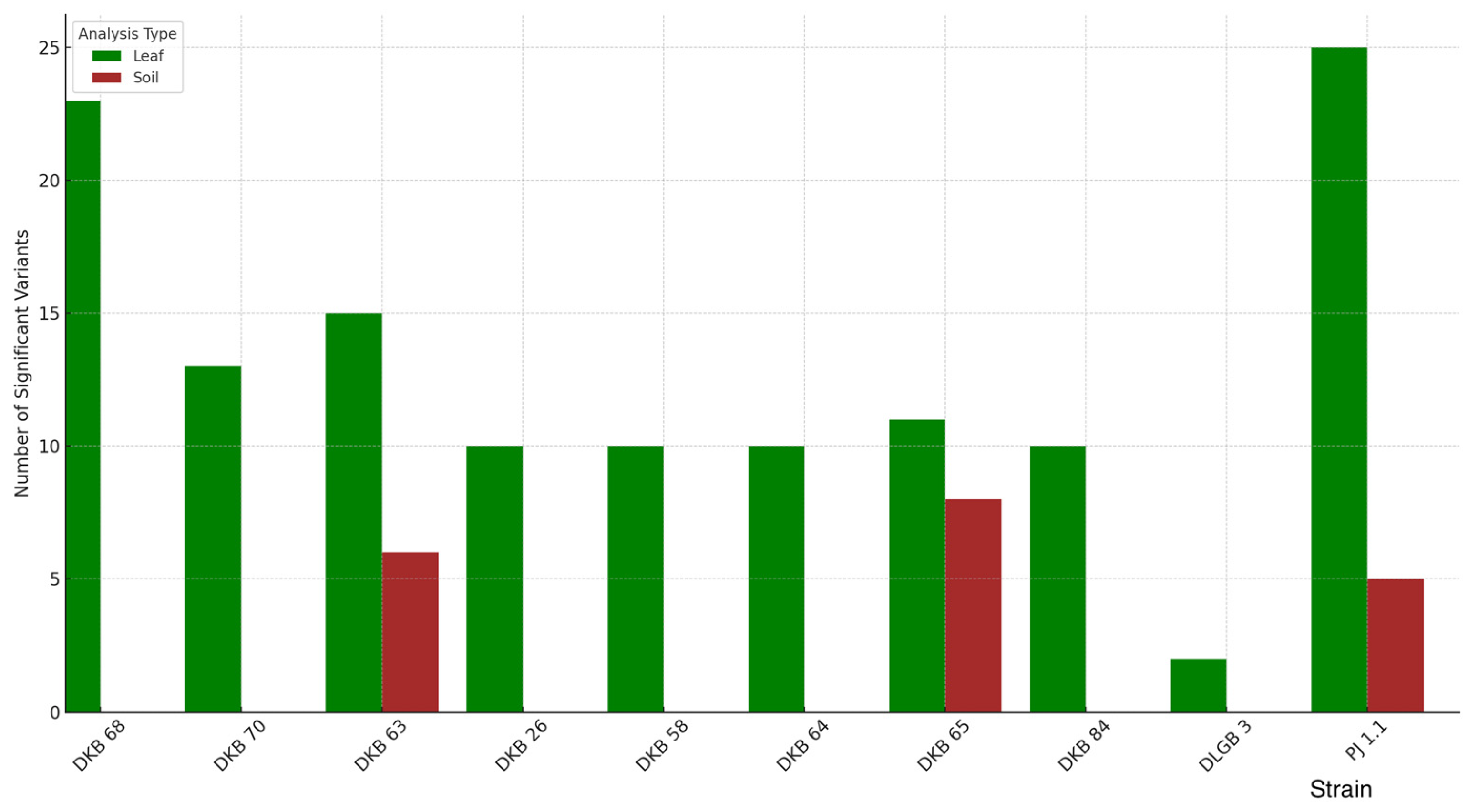
3.8. Leaf Analysis Effect Summary
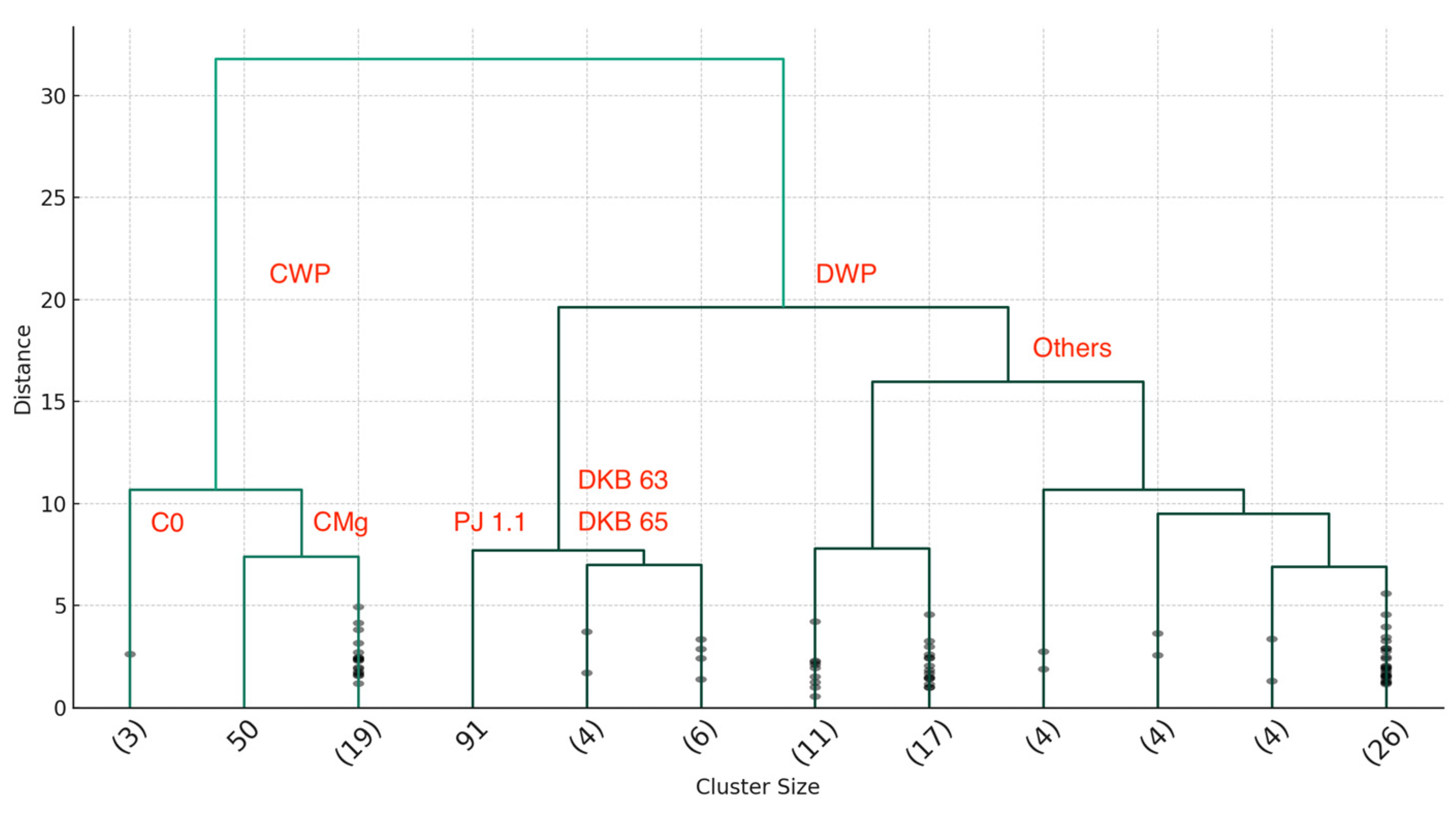
3.9. Cluster Analysis
3.9.1. Substrate Composition Clusters
- Optimal Condition Cluster
- 2.
- Water Deficit Clusters
- 3.
- Diverse Response Clusters (‘Others’):
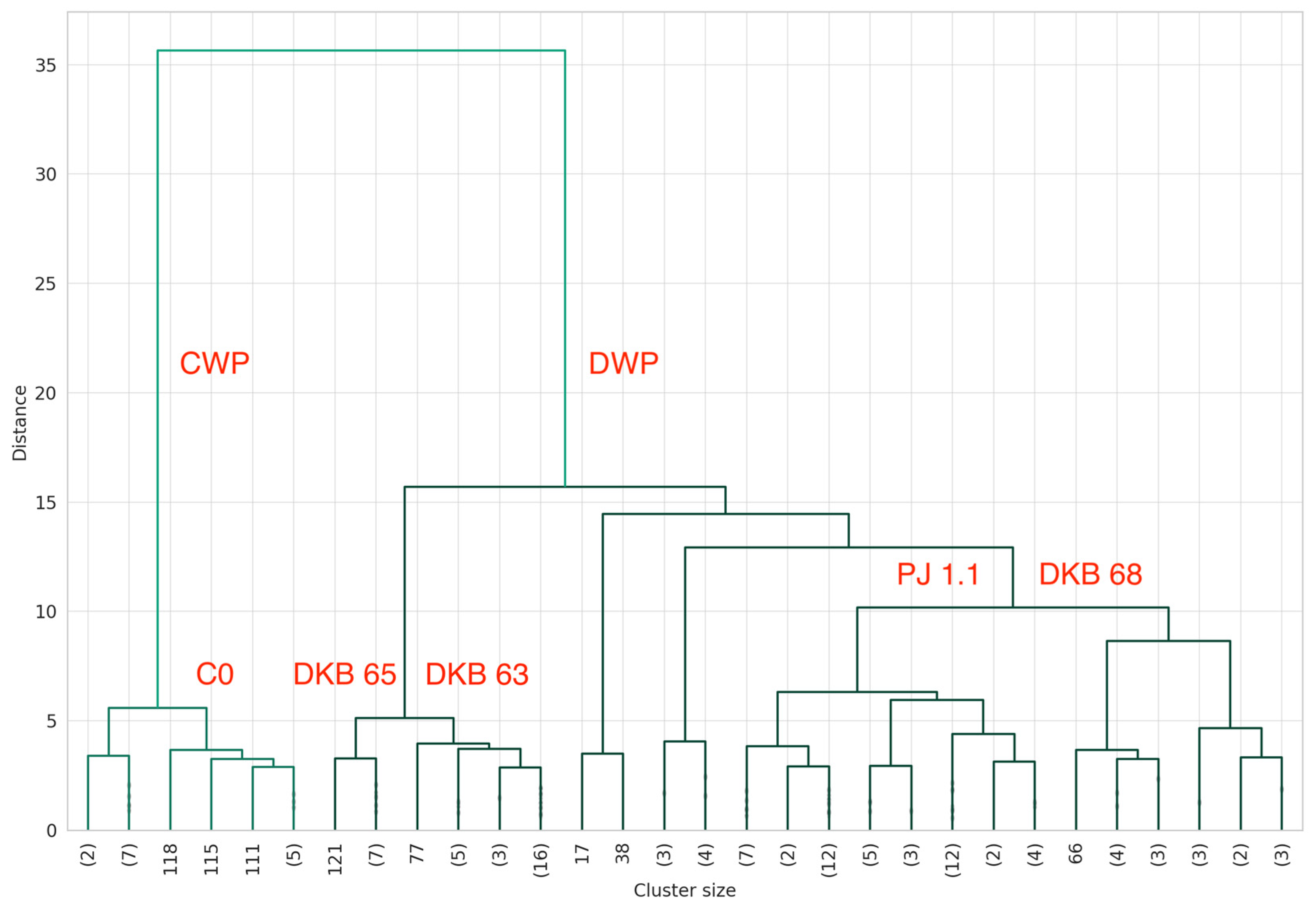
3.9.2. Ward Dendrogram Interpretation for Plant Tissue Analysis
- Optimal Branch:
- 2.
- Water Deficit Branch:
3.9.3. Scientific Importance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ozturk, M.; Turkyilmaz Unal, B.; García-Caparrós, P.; Khursheed, A.; Gul, A.; Hasanuzzaman, M. Osmoregulation and its actions during the drought stress in plants. Physiol. Plantarum. 2021, 172, 1321–1335. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Faaek, M.F.; Pirlak, L. Morphological and physiological effects of drought stress on some strawberry cultivars. Selcuk J. Agric. Food Sci. 2021, 35, 194–201. [Google Scholar] [CrossRef]
- World Strawberry Production by Country. Available online: https://www.atlasbig.com/en-us/countries-strawberry-production (accessed on 13 November 2023).
- Rutkowski, K.P.; Kruczyńska, D.E.; Żurawicz, E. Quality and shelf life of strawberry cultivars in Poland. Acta Hortic. 2006, 708, 329–332. [Google Scholar] [CrossRef]
- Chaves, V.C.; Calvete, E.; Reginatto, F.H. Quality properties and antioxidant activity of seven strawberry (Fragaria × ananassa Duch.) cultivars. Sci. Horticult. 2017, 225, 293–298. [Google Scholar] [CrossRef]
- Unal, N.; Okatan, V.; Bilgin, J.; Kahramanoğlu, I.; Hajizadeh, H.S. Impacts of different planting times on fruit quality and some bioactive contents of different strawberry cultivars. Folia Hortic. 2023, 35, 221–231. [Google Scholar] [CrossRef]
- Gondek, K.; Mierzwa-Hersztek, M.; Kopeć, M.; Zaleski, T.; Bogdał, S.; Bieniasz, M.; Błaszczyk, J.; Kaczmarczyk, E.; Kowalczyk, B.; Knaga, J.; et al. Mineral composition of fruits and leaves of San Andreas® everbearing strawberry in soilless cultivation. J. Elem. 2020, 25, 1333–1347. [Google Scholar] [CrossRef]
- Wysocki, K.; Kopytowski, J.; Bieniek, A.; Bojarska, J. The effect of substrates on yield and quality of strawberry fruits cultivated in a heated foil tunnel. Zemdirbyste 2017, 104, 283–286. [Google Scholar] [CrossRef]
- Mikiciuk, G.; Mikiciuk, M.; Hawrot-Paw, M. Influence of superabsorbent polymers on the chemical composition of strawberry (Fragaria × ananassa Duch.) and biological activity in the soil. Folia Hortic. 2015, 27, 63–69. [Google Scholar] [CrossRef]
- Kour, D.; Yadav, A.N. Bacterial mitigation of drought stress in plants: Current perspectives and future challenges. Curr. Microbiol. 2022, 79, 248. [Google Scholar] [CrossRef] [PubMed]
- Azeem, M.; Haider, M.Z.; Javed, S.; Saleem, M.H.; Alatawi, A. Drought stress amelioration in maize (Zea mays L.) by inoculation of Bacillus spp. strains under sterile soil conditions. Agriculture 2022, 12, 50. [Google Scholar] [CrossRef]
- Goswami, M.; Suresh, D. Plant growth-promoting rhizobacteria—Alleviators of abiotic stresses in soil: A review. Pedosphere 2020, 30, 40–61. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef] [PubMed]
- García-Caparrós, P.; Romero, M.J.; Llanderal, A.; Cermeño, P.; Lao, M.T.; Segura, M.L. Effects of drought stress on biomass, essential oil content, nutritional parameters, and costs of production in six Lamiaceae species. Water 2019, 11, 573. [Google Scholar] [CrossRef]
- Vandana, U.K.; Singha, B.; Gulzar, A.B.M.; Mazumder, P.B. Molecular mechanisms in plant growth promoting bacteria (PGPR) to resist environmental stress in plants. In Molecular Aspects of Plant Beneficial Microbes in Agriculture; Sharma, V., Salwan, R., Tawfeeq, L., Eds.; Academic Press: Cambridge, MA, USA; Elsevier Inc.: Hoboken, NJ, USA, 2020; pp. 221–233. [Google Scholar] [CrossRef]
- Kaur, H.; Kaur, J.; Gera, R. Plant growth promoting rhizobacteria: A boon to agriculture. Int. J. Cell Sci. Biotechnol. 2016, 5, 17–22. [Google Scholar]
- Mustafa, S.; Kabir, S.; Shabbir, U.; Batool, R. Plant growth promoting rhizobacteria in sustainable agriculture: From theoretical to pragmatic approach. Symbiosis 2019, 78, 115–123. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Ali, S.; Babar, M.A. Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regul. 2020, 90, 189–203. [Google Scholar] [CrossRef]
- Sindhu, S.S.; Dua, S.; Verma, M.K.; Khandelwal, A. Growth promotion of legumes by inoculation of rhizosphere bacteria. In Microbes for Legume Improvement; Khan, M.S., Musarrat, J., Zaidi, A., Eds.; Springer: Vienna, Austria, 2010; pp. 195–235. [Google Scholar] [CrossRef]
- Miller, S.H.; Browne, P.; Prigent-Combaret, C.; Combes-Meynet, E.; Morrissey, J.P.; O’Gara, F. Biochemical and genomic comparison of inorganic phosphate solubilisation in Pseudomonas species. Environ. Microbiol. Rep. 2009, 2, 403–411. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress: Significance of mineral nutrients. In Water Stress and Crop Plants: A Sustainable Approach; Ahmad, P., Ed.; Wiley Blackwell: Weinheim, Germany, 2016; pp. 649–668. [Google Scholar] [CrossRef]
- Etesami, H.; Adl, S.M. Plant growth-promoting rhizobacteria (PGPR) and their action mechanisms in availability of nutrients to plants. In Phyto-Microbiome in Stress Regulation; Kumar, M., Kumar, V., Prasad, R., Eds.; Springer: Singapore, 2020; pp. 147–203. [Google Scholar] [CrossRef]
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Hossain, M., Wani, S., Bhattacharjee, S., Burritt, D., Tran, L.S., Eds.; Springer: Cham, Switzerland, 2016; pp. 1–16. [Google Scholar] [CrossRef]
- Tadayyon, A.; Nikneshan, P.; Pessarakli, M. Effects of drought stress on concentration of macro-and micro-nutrients in castor (Ricinus communis L.) plant. J. Plant Nutr. 2018, 41, 304–310. [Google Scholar] [CrossRef]
- Sun, F.; Ou, Q.; Wang, N.; Guo, Z.X.; Ou, Y.; Li, N.; Peng, C. Isolation and identification of potassium-solubilizing bacteria from Mikania micrantha rhizospheric soil and their effect on M. micrantha plants. Glob. Ecol. Conser. 2020, 23, e01141. [Google Scholar] [CrossRef]
- Kaur, T.; Devi, R.; Kour, D.; Yadav, A.; Yadav, A.N. Plant growth promotion of barley (Hordeum vulgare L.) by potassium solubilizing bacteria with multifarious plant growth promoting attributes. Plant Sci. Today 2021, 8, 17–24. [Google Scholar] [CrossRef]
- Verma, P.; Yadav, A.N.; Khannam, K.S.; Saxena, A.K.; Suman, A. Potassium-solubilizing microbes: Diversity, distribution, and role in plant growth promotion. In Microorganisms for Green Revolution. Volume 1: Microbes for Sustainable Crop Production; Panpatte, D.G., Jhala, Y.K., Vyas, R.V., Shelat, H.N., Eds.; Springer: Singapore, 2017; pp. 125–149. [Google Scholar] [CrossRef]
- Singh, G.; Biswas, D.R.; Marwaha, T.S. Mobilization of potassium from waste mica by plant growth promoting rhizobacteria and its assimilation by maize (Zea mays) and wheat (Triticum aestivum): A hydroponics study under phytotron growth chamber. J. Plant Nutr. 2010, 33, 236–1251. [Google Scholar] [CrossRef]
- Hussain, H.A.; Hussain, S.; Khaliq, A.; Ashraf, U.; Anjum, S.A.; Men, S.; Wang, L. Chilling and drought stresses in crop plants: Implications, cross talk, and potential management opportunities. Front. Plant Sci. 2018, 9, 393. [Google Scholar] [CrossRef] [PubMed]
- D’Oria, A.; Courbet, G.; Billiot, B.; Jing, L.; Pluchon, S.; Arkoun, M.; Maillard, A.; Roux, C.P.; Trouverie, J.; Etienne, P.; et al. Drought specifically downregulates mineral nutrition: Plant ionomic content and associated gene expression. Plant Direct 2022, 6, e402. [Google Scholar] [CrossRef] [PubMed]
- Ghavami, N.; Alikhani, H.A.; Pourbabaee, A.A.; Besharati, H. Study the effects of siderophore-producing bacteria on zinc and phosphorous nutrition of canola and maize plants. Commun. Soil Sci. Plant Anal. 2016, 47, 1517–1527. [Google Scholar] [CrossRef]
- Radzki, W.; Manero, F.G.; Algar, E.; García, J.L.; García-Villaraco, A.; Solano, B.R. Bacterial siderophores efficiently provide iron to iron-starved tomato plants in hydroponics culture. Antonie Van Leeuwenhoek 2013, 104, 321–330. [Google Scholar] [CrossRef]
- Paliwoda, D.; Mikiciuk, G.; Mikiciuk, M.; Kisiel, A.; Sas-Paszt, L.; Miller, T. Effects of rhizosphere bacteria on strawberry plants (Fragaria × ananassa Duch.) under water deficit. Int. J. Mol. Sci. 2022, 23, 10449. [Google Scholar] [CrossRef] [PubMed]
- PN-EN 15935:2022-01; Soil, Waste, Treated Bio-Waste and Sewage Sludge—Determination of Loss on Ignition (LOI). Polish Committee for Standardization: Warszawa, Poland, 2022.
- PN-EN ISO 10390:2022-09; Soil, Treated Bio-Waste and Sewage Sludge—Determination of pH. Polish Committee for Standardization: Warszawa, Poland, 2022.
- PN-ISO 11265:1997; Soil Quality—Determination of the Specific Electrical Conductivity. Polish Committee for Standardization: Warszawa, Poland, 1997.
- PN-R-04024:1997; Analysis of the Chemical-Agricultural Soil—Determination of the Content of Available Phosphorus, Potassium, Magnesium and Manganese in Organic Soils. Polish Committee for Standardization: Warszawa, Poland, 1997. (In Polish)
- Hunter, J.D. Matplotlib: A 2D graphics environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Waskom, M.L. Seaborn: Statistical data visualization. J. Open Source Softw. 2021, 6, 3021. [Google Scholar] [CrossRef]
- Virtanen, P.; Gommers, R.; Oliphant, T.E.; Haberland, M.; Reddy, T.; Cournapeau, D.; Burovski, E.; Peterson, P.; Weckesser, W.; Bright, J.; et al. SciPy 1.0: Fundamental algorithms for scientific computing in Python. Nat. Methods 2020, 17, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Seabold, S.; Perktold, J. Statsmodels: Econometric and statistical modeling with Python. In Proceedings of the 9th Python in Science Conference, SciPy, Austin, TX, USA, 28 June–3 July 2010; Volume 57, p. 61. [Google Scholar] [CrossRef]
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, B.; Muhammad, B.L.; Jakada, B.H.; Vyas, N.L. Rhizosphere microbiome and plant nutrition. IJETST 2015, 2, 3208–3216. [Google Scholar] [CrossRef]
- Gupta, A.; Mishra, R.; Rai, S.; Bano, A.; Pathak, N.; Fujita, M.; Kumar, M.; Hasanuzzaman, M. Mechanistic insights of Plant Growth Promoting Bacteria mediated drought and salt stress tolerance in plants for sustainable agriculture. Int. J. Mol. Sci. 2022, 23, 3741. [Google Scholar] [CrossRef] [PubMed]
- Batool, M.; El-Badri, A.M.; Hassan, M.U.; Haiyun, Y.; Chunyun, W.; Zhenkun, Y.; Jie, K.; Wang, B.; Zhou, G. Drought stress in Brassica napus: Effects, tolerance mechanisms, and management strategies. J. Plant Growth Regul. 2022, 42, 1–25. [Google Scholar] [CrossRef]
- Al-Kaisi, M.M.; Elmore, R.W.; Guzman, J.G.; Hanna, H.M.; Hart, C.E.; Helmers, M.J.; Hodgson, E.W.; Lenssen, A.W.; Mallarino, A.P.; Robertson, A.E.; et al. Drought impact on crop production and the soil environment: 2012 experiences from Iowa. J. Soil Water Conserv. 2013, 68, 19A–24A. [Google Scholar] [CrossRef]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry, and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resour. Environ. Sustain. 2021, 5, 100032. [Google Scholar] [CrossRef]
- Babu, A.N.; Jogaiah, S.; Ito, S.-I.; Nagaraj, A.K.; Tran, L.-S. Improvement of growth, fruit weight and early blight disease protection of tomato plants by rhizosphere bacteria is correlated with their beneficial traits and induced biosynthesis of antioxidant peroxidase and polyphenol oxidase. Plant Sci. 2015, 231, 62–73. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El Enshasy, H.A.; Dailin, D.J.; Suriani, N. Recent understanding of soil Acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Tewari, S.; Arora, N.K. Multifunctional exopolysaccharides from Pseudomonas aeruginosa PF23 involved in plant growth stimulation, biocontrol and stress amelioration in sunflower under saline conditions. Curr. Microbiol. 2014, 69, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Raheem, A.; Shaposhnikov, A.; Belimov, A.A.; Dodd, I.C.; Ali, B. Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arch. Agron. Soil Sci. 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Gontia-Mishra, I.; Sapre, S.; Deshmukh, R.; Sikdar, S.; Tiwari, S. Microbe-mediated drought tolerance in plants: Current developments and future challenges. In Plant Microbiomes for Sustainable Agriculture; Yadav, A.N., Singh, J., Rastegari, A.A., Yadav, N., Eds.; Springer: Cham, Switzerland, 2020; pp. 351–379. [Google Scholar] [CrossRef]
- Zhao, K.; Penttinen, P.; Zhang, X.; Ao, X.; Liu, M.; Yu, X.; Chen, Q. Maize rhizosphere in Sichuan, China, hosts plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microbiol. Res. 2014, 169, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [PubMed]
- Billah, M.; Khan, M.; Bano, A.; Ul Hassan, T.; Munir, A.; Gurmani, A. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol. J. 2019, 36, 904–916. [Google Scholar] [CrossRef]
- Singha, K.M.; Singh, B.; Pandey, P. Host specific endophytic microbiome diversity and associated functions in three varieties of scented black rice are dependent on growth stage. Sci. Rep. 2021, 11, 12259. [Google Scholar] [CrossRef] [PubMed]
- Karlidag, H.; Esitken, A.; Turan, M.; Sahin, F. Effects of root inoculation of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient element contents of leaves of apple. Sci. Hortic. 2007, 114, 16–20. [Google Scholar] [CrossRef]
- Terzano, R.; Cuccovillo, G.; Gattullo, C.E.; Medici, L.; Tomasi, N.; Pinton, R.; Mimmo, T.; Cesco, S. Combined effect of soil organic acids and flavonoids on the mobilization of major and trace elements from soil. Biol. Fertil. Soils 2015, 51, 685–695. [Google Scholar] [CrossRef]
- Çakmakçı, R.; Haliloglu, K.; Türkoğlu, A.; Özkan, G.; Kutlu, M.; Varmazyari, A.; Molnar, Z.; Jamshidi, B.; Pour-Aboughadareh, A.; Bocianowski, J. Effect of different Plant Growth-Promoting Rhizobacteria on biological soil properties, growth, yield and quality of oregano (Origanum onites L.). Agronomy 2023, 13, 2511. [Google Scholar] [CrossRef]
- Adedeji, A.A.; Häggblom, M.M.; Babalola, O.O. Sustainable agriculture in Africa: Plant growth-promoting rhizobacteria (PGPR) to the rescue. Sci. Afr. 2020, 9, e00492. [Google Scholar] [CrossRef]
- Creus, C.M.; Pereyra, M.A.; Casanovas, E.M.; Sueldo, R.J.; Barassi, C.A. Plant growth-promoting effects of rhizobacteria on abiotic stressed plants. Azospirillum-grasses model. Am. J. Plant Sci. Biotech. 2010, 4, 49–59. [Google Scholar]
- Naveed, M.; Hussain, M.B.; Zahir, Z.A.; Mitter, B.; Sessitsch, A. Drought stress amelioration in wheat through inoculation with Burkholderia phytofirmans strain PsJN. Plant Growth Regul. 2014, 73, 121–131. [Google Scholar] [CrossRef]
- Fonseca, M.d.C.d.; Bossolani, J.W.; de Oliveira, S.L.; Moretti, L.G.; Portugal, J.R.; Scudeletti, D.; de Oliveira, E.F.; Crusciol, C.A.C. Bacillus subtilis inoculation improves nutrient uptake and physiological activity in sugarcane under drought stress. Microorganisms 2022, 10, 809. [Google Scholar] [CrossRef] [PubMed]
- Perin, E.C.; Messias, R.D.S.; Galli, V.; Borowski, J.M.; Souza, E.R.D.; Avila, L.O.D.; Bamberg, A.L.; Rombaldi, C.V. Mineral content and antioxidant compounds in strawberry fruit submitted to drough stress. Food Sci. Technol. 2019, 39, 245–254. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed]
- Dos Lopes, M.J.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful Plant Growth-Promoting Microbes: Inoculation methods and abiotic factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Omar, A.F.; Rehan, M.; AL-Turki, A. Alleviating the adverse effects of plant pathogens, drought and salinity stress factors using plant growth promoting bacteria. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12911. [Google Scholar] [CrossRef]
- Dotaniya, M.L.; Meena, V.D. Rhizosphere effect on nutrient availability in soil and its uptake by plants: A review. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 85, 1–12. [Google Scholar] [CrossRef]
- Aketi, R.; Sharma, S.K.; Sharma, M.P.; Namrata, Y.; Joshi, O.P. Inoculation of zinc solubilizing Bacillus aryabhattai strains for improved growth, mobilization and biofortification of zinc in soybean and wheat cultivated in vertisols of central India. Appl. Soil Ecol. 2014, 73, 87–96. [Google Scholar] [CrossRef]
- Armada, E.; Roldán, A.; Azcon, R. Differential activity of autochthonous bacteria in controlling drought stress in native Lavandula and Salvia plants species under drought conditions in natural arid soil. Microb. Ecol. 2014, 67, 410–420. [Google Scholar] [CrossRef]
- Pitura, K.; Michałojć, Z. Influence of nitrogen doses on the chemical composition and proportions of nutrients in selected vegetable species. J. Elem. 2015, 20, 667–676. [Google Scholar] [CrossRef]






























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Variant | Strain 1 | Strain 2 | Mean 1 | Mean 2 | Effect |
|---|---|---|---|---|---|---|
| pH in KCl | DWP * | DKB63 | C0 | 5.885 | 6.561 | DOWN |
| DKB63 | CMg | 5.885 | 6.559 | |||
| DKB65 | C0 | 5.918 | 6.561 | |||
| DKB65 | CMg | 5.918 | 6.559 | |||
| C0 | PJ1.1 | 6.561 | 5.880 | |||
| CMg | PJ1.1 | 6.559 | 5.880 | |||
| pH in H2O | DWP | DKB63 | C0 | 6.110 | 6.911 | DOWN |
| DKB63 | CMg | 6.110 | 6.906 | |||
| DKB65 | C0 | 6.140 | 6.911 | |||
| DKB65 | CMg | 6.140 | 6.906 | |||
| C0 | PJ1.1 | 6.911 | 6.127 | |||
| CMg | PJ1.1 | 6.906 | 6.127 | |||
| Salinity [mS∙cm−1] | DWP | DKB63 | C0 | 1888.250 | 986.222 | UP |
| DKB63 | CMg | 1888.250 | 996.125 | |||
| DKB65 | C0 | 1817.500 | 986.222 | |||
| DKB65 | CMg | 1817.500 | 996.125 | |||
| Fe [mg·kg−1] | CWP * | CMg | C0 | 1743.625 | 1732.444 | DOWN |
| Cu [mg·kg−1] | CWP | C0 | PJ1.1 | 40.178 | 26.400 | DOWN |
| P [mg·100 g−1] | DWP | DKB65 | C0 | 159.200 | 102.811 | UP |
| DKB65 | CMg | 159.200 | 102.538 |
| Variable | Variant | Strain 1 | Strain 2 | Mean Value 1 | Mean Value 2 | Effect |
|---|---|---|---|---|---|---|
| Mg | CWP * | DKB68 | C0 | 5244.40 | 3693.88 | UP |
| DKB68 | CMg | 5244.40 | 3617.79 | |||
| C0 | PJ1.1 | 3693.88 | 5706.55 | |||
| CMg | PJ1.1 | 3617.79 | 5706.55 | |||
| Ca | CWP | DKB68 | C0 | 22,911.27 | 9477.90 | UP |
| DKB68 | CMg | 22,911.27 | 10,103.08 | |||
| C0 | PJ1.1 | 9477.90 | 22,587.73 | |||
| CMg | PJ1.1 | 10,103.08 | 22,587.73 | |||
| Na | CWP | DKB70 | C0 | 299.19 | 600.02 | DOWN |
| Ca:P | CWP | DKB68 | C0 | 4.50 | 1.76 | UP |
| DKB68 | CMg | 4.50 | 1.73 | |||
| C0 | PJ1.1 | 1.76 | 4.12 | |||
| CMg | PJ1.1 | 1.73 | 4.12 | |||
| K:Ca | CWP | DKB68 | C0 | 0.42 | 1.15 | UP |
| DKB68 | CMg | 0.42 | 1.06 | DOWN | ||
| C0 | PJ1.1 | 1.15 | 0.47 | DOWN | ||
| K:Mg | CWP | CMg | PJ1.1 | 1.06 | 0.47 | DOWN |
| DKB68 | CMg | 1.11 | 1.81 | |||
| C0 | PJ1.1 | 1.80 | 1.14 | |||
| CMg | PJ1.1 | 1.81 | 1.14 | |||
| Ca:Mg | CWP | DKB68 | C0 | 2.68 | 1.58 | UP |
| DKB68 | CMg | 2.68 | 1.73 | |||
| C0 | PJ1.1 | 1.58 | 2.42 | |||
| CMg | PJ1.1 | 1.73 | 2.42 | |||
| Cu | DWP * | DKB63 | C0 | 30.18 | 10.39 | UP |
| DKB63 | CMg | 30.18 | 13.44 | |||
| C0 | PJ1.1 | 10.39 | 26.20 | |||
| Mg | DWP | DKB26 | C0 | 4210.68 | 5610.40 | DOWN |
| DKB26 | CMg | 4210.68 | 5267.34 | |||
| DKB58 | C0 | 4193.80 | 5610.40 | |||
| DKB58 | CMg | 4193.80 | 5267.34 | |||
| DKB63 | C0 | 3777.40 | 5610.40 | |||
| DKB63 | CMg | 3777.40 | 5267.34 | |||
| DKB64 | C0 | 3183.37 | 5610.40 | |||
| DKB64 | CMg | 3183.37 | 5267.34 | |||
| DKB65 | C0 | 3780.70 | 5610.40 | |||
| DKB65 | CMg | 3780.70 | 5267.34 | |||
| DKB68 | C0 | 3668.17 | 5610.40 | |||
| DKB68 | CMg | 3668.17 | 5267.34 | |||
| DKB70 | C0 | 3667.00 | 5610.40 | |||
| DKB70 | CMg | 3667.00 | 5267.34 | |||
| DKB84 | C0 | 4163.43 | 5610.40 | |||
| DKB84 | CMg | 4163.43 | 5267.34 | |||
| DLGB3 | C0 | 4484.60 | 5610.40 | |||
| C0 | PJ1.1 | 5610.40 | 3950.80 | |||
| CMg | PJ1.1 | 5267.34 | 3950.80 | |||
| Ca | DWP | DKB26 | C0 | 11,745.15 | 21,415.13 | DOWN |
| DKB26 | CMg | 11,745.15 | 19,522.86 | |||
| DKB58 | C0 | 10,942.10 | 21,415.13 | |||
| DKB58 | CMg | 10,942.10 | 19,522.86 | |||
| DKB63 | C0 | 12,337.80 | 21,415.13 | |||
| DKB63 | CMg | 12,337.80 | 19,522.86 | |||
| DKB64 | C0 | 10,129.50 | 21,415.13 | |||
| DKB64 | CMg | 10,129.50 | 19,522.86 | |||
| DKB65 | C0 | 8819.90 | 21,415.13 | |||
| DKB65 | CMg | 8819.90 | 19,522.86 | |||
| Ca | DWP | DKB68 | C0 | 10,129.40 | 21,415.13 | DOWN |
| DKB68 | CMg | 10,129.40 | 19,522.86 | |||
| DKB70 | C0 | 9166.80 | 21,415.13 | |||
| DKB70 | CMg | 9166.80 | 19,522.86 | |||
| DKB84 | C0 | 10,643.60 | 21,415.13 | |||
| DKB84 | CMg | 10,643.60 | 19,522.86 | |||
| C0 | PJ1.1 | 21,415.13 | 12,796.30 | |||
| CMg | PJ1.1 | 19,522.86 | 12,796.30 | |||
| Na | DWP | DKB63 | CMg | 639.75 | 338.09 | UP |
| P | DWP | DKB63 | C0 | 10,149.00 | 5534.48 | UP |
| DKB63 | CMg | 10,149.00 | 5383.28 | |||
| C0 | PJ1.1 | 5534.48 | 9392.40 | |||
| CMg | PJ1.1 | 5383.28 | 9392.40 | |||
| Ca:P | DWP | DKB26 | C0 | 2.00 | 3.91 | DOWN |
| DKB26 | CMg | 2.00 | 3.65 | |||
| DKB58 | C0 | 1.71 | 3.91 | |||
| DKB58 | CMg | 1.71 | 3.65 | |||
| DKB63 | C0 | 1.22 | 3.91 | |||
| DKB63 | CMg | 1.22 | 3.65 | |||
| DKB64 | C0 | 1.87 | 3.91 | |||
| DKB64 | CMg | 1.87 | 3.65 | |||
| DKB65 | C0 | 1.61 | 3.91 | |||
| DKB65 | CMg | 1.61 | 3.65 | |||
| DKB68 | C0 | 1.72 | 3.91 | |||
| DKB68 | CMg | 1.72 | 3.65 | |||
| DKB70 | C0 | 1.53 | 3.91 | |||
| DKB70 | CMg | 1.53 | 3.65 | |||
| DKB84 | C0 | 2.01 | 3.91 | |||
| DKB84 | CMg | 2.01 | 3.65 | |||
| DLGB3 | C0 | 2.91 | 3.91 | |||
| C0 | PJ1.1 | 3.91 | 1.37 | |||
| CMg | PJ1.1 | 3.65 | 1.37 | |||
| K:Ca | DWP | DKB26 | C0 | 0.78 | 0.40 | UP |
| DKB26 | CMg | 0.78 | 0.44 | |||
| DKB58 | C0 | 0.82 | 0.40 | |||
| DKB58 | CMg | 0.82 | 0.44 | |||
| DKB63 | C0 | 0.79 | 0.40 | |||
| DKB63 | CMg | 0.79 | 0.44 | |||
| DKB64 | C0 | 0.93 | 0.40 | |||
| DKB64 | CMg | 0.93 | 0.44 | |||
| K:Ca | DWP | DKB65 | C0 | 0.98 | 0.40 | UP |
| DKB65 | CMg | 0.98 | 0.44 | |||
| DKB68 | C0 | 0.96 | 0.40 | |||
| DKB68 | CMg | 0.96 | 0.44 | |||
| DKB70 | C0 | 0.99 | 0.40 | |||
| DKB70 | CMg | 0.99 | 0.44 | |||
| DKB84 | C0 | 0.85 | 0.40 | |||
| DKB84 | CMg | 0.85 | 0.44 | |||
| C0 | PJ1.1 | 0.40 | 0.77 | |||
| CMg | PJ1.1 | 0.44 | 0.77 | |||
| K:Mg | DWP | DKB63 | C0 | 1.58 | 0.94 | UP |
| DKB63 | CMg | 1.58 | 1.00 | |||
| DKB64 | C0 | 1.82 | 0.94 | |||
| DKB64 | CMg | 1.82 | 1.00 | |||
| DKB65 | C0 | 1.40 | 0.94 | |||
| DKB68 | C0 | 1.62 | 0.94 | |||
| DKB68 | CMg | 1.62 | 1.00 | |||
| DKB70 | C0 | 1.52 | 0.94 | |||
| DKB70 | CMg | 1.52 | 1.00 | |||
| C0 | PJ1.1 | 0.94 | 1.51 | |||
| CMg | PJ1.1 | 1.00 | 1.51 | |||
| Ca:Mg | DWP | DKB26 | C0 | 1.71 | 2.34 | DOWN |
| DKB26 | CMg | 1.71 | 2.27 | |||
| DKB58 | C0 | 1.60 | 2.34 | |||
| DKB58 | CMg | 1.60 | 2.27 | |||
| DKB65 | C0 | 1.44 | 2.34 | |||
| DKB65 | CMg | 1.44 | 2.27 | |||
| DKB68 | C0 | 1.62 | 2.34 | |||
| DKB68 | CMg | 1.62 | 2.27 | |||
| DKB70 | C0 | 1.53 | 2.34 | |||
| DKB70 | CMg | 1.53 | 2.27 | |||
| DKB84 | C0 | 1.57 | 2.34 | |||
| DKB84 | CMg | 1.57 | 2.27 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paliwoda, D.; Mikiciuk, G.; Chudecka, J.; Tomaszewicz, T.; Miller, T.; Mikiciuk, M.; Kisiel, A.; Sas-Paszt, L. Effects of Inoculation with Plant Growth-Promoting Rhizobacteria on Chemical Composition of the Substrate and Nutrient Content in Strawberry Plants Growing in Different Water Conditions. Agriculture 2024, 14, 46. https://doi.org/10.3390/agriculture14010046
Paliwoda D, Mikiciuk G, Chudecka J, Tomaszewicz T, Miller T, Mikiciuk M, Kisiel A, Sas-Paszt L. Effects of Inoculation with Plant Growth-Promoting Rhizobacteria on Chemical Composition of the Substrate and Nutrient Content in Strawberry Plants Growing in Different Water Conditions. Agriculture. 2024; 14(1):46. https://doi.org/10.3390/agriculture14010046
Chicago/Turabian StylePaliwoda, Dominika, Grzegorz Mikiciuk, Justyna Chudecka, Tomasz Tomaszewicz, Tymoteusz Miller, Małgorzata Mikiciuk, Anna Kisiel, and Lidia Sas-Paszt. 2024. "Effects of Inoculation with Plant Growth-Promoting Rhizobacteria on Chemical Composition of the Substrate and Nutrient Content in Strawberry Plants Growing in Different Water Conditions" Agriculture 14, no. 1: 46. https://doi.org/10.3390/agriculture14010046