

Dependency of Crops on Pollinators and Pollination Deficits: An Approach to Measurement Considering the Influence of Various Reproductive Traits


Abstract
:1. Introduction
2. Materials and Methods
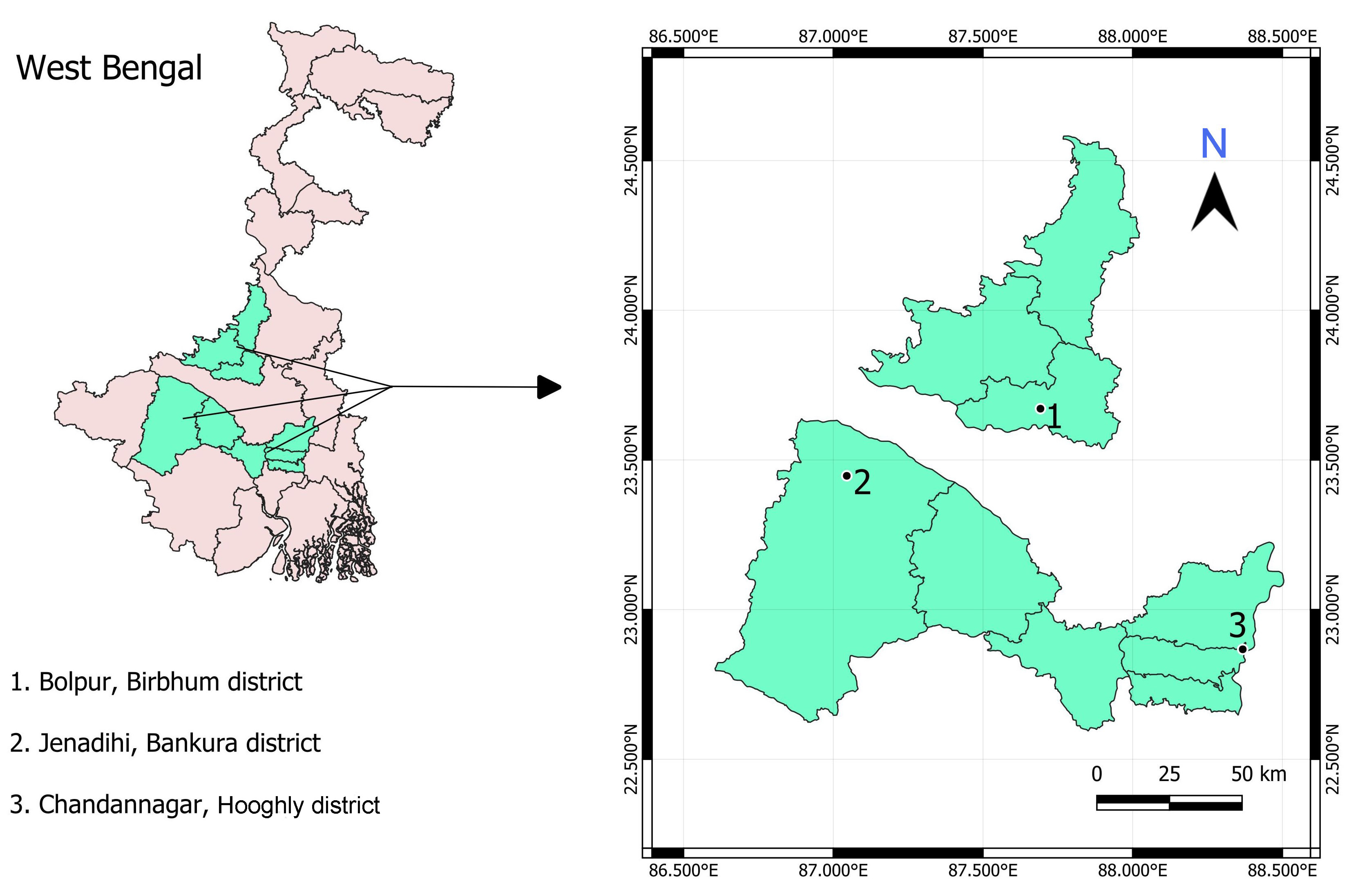
2.1. Experimental Site
2.2. Selected Crops
2.3. Dependency of Crops on Pollinators and Pollination Deficit
2.4. Data Collection about Reproductive Traits
2.5. Statistical Analyses
3. Results
3.1. Dependency of Crops on Pollinators and Pollination Deficit
3.2. Reproductive Traits of the Crops
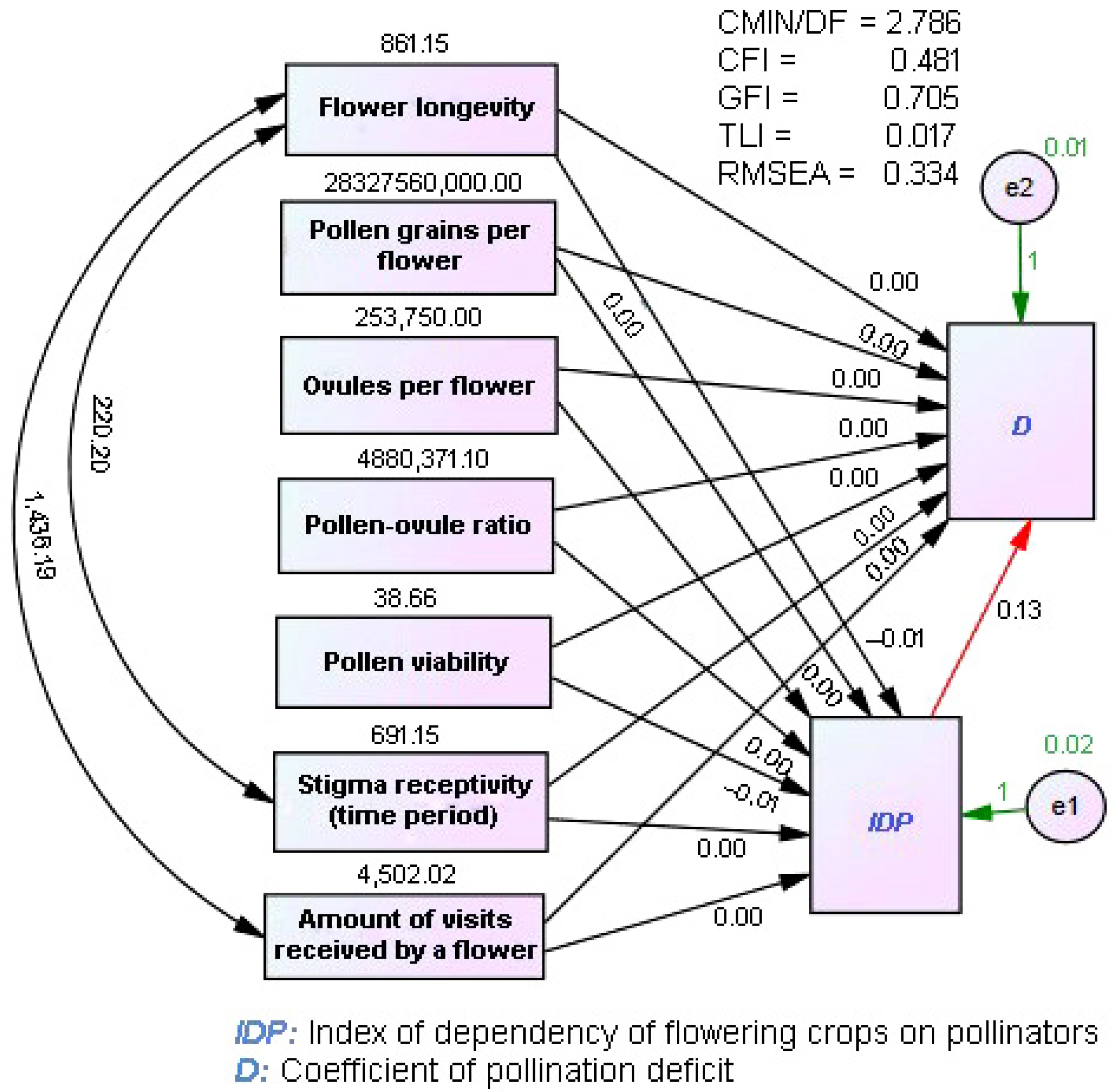
3.3. Influence of Reproductive Traits on a Crop’s Dependency on Pollinators and Pollination Deficit
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, A.M.; Vaissiere, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunninggham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.; Aizen, M.A.; Cunninggham, S.; Klein, A.M. Pollinator shortage and global crop yield. Commun. Integr. Biol. 2009, 2, 37–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodger, J.G.; Bennett, J.M.; Razanajatovo, M.; Knight, T.M.; van Kleunen, M.; Ashman, T.L.; Steets, J.A.; Hui, C.; Arceo-G’omez, G.; Burd, M.; et al. Widespread vulnerability of flowering plant seed production to pollinator declines. Sci. Adv. 2021, 7, eabd3524. [Google Scholar] [CrossRef] [PubMed]
- Baker, H.G. Self-compatibility and establishment after “long-distance” dispersal. Evolution 1955, 9, 347–349. [Google Scholar]
- Lloyd, D.G. Demographic factors and mating patterns in angiosperms. In Demography and Evolution in Plant Populations, Botanical Monographs; Solbrig, O.T., Ed.; Blackwell: Oxford, UK, 1980; pp. 67–88. [Google Scholar]
- Lloyd, D.G.; Schoen, D.J. Self- and cross-fertilization in plants. I. Functional dimensions. Int. J. Plant Sci. 1992, 153, 358–369. [Google Scholar] [CrossRef]
- Calviño, A.; Galetto, L. Cleistogamy in the rare high Andean perennial herb Cryptantha capituliflora (Boraginaceae). Plant Syst. Evol. 2003, 237, 41–50. [Google Scholar] [CrossRef]
- Knight, T.M.; Steets, J.A.; Vamosi, J.C.; Mazer, S.J.; Burd, M.; Campbell, D.R.; Dudash, M.R.; Johnston, M.O.; Mitchell, R.J.; Ashman, T.L. Pollen limitation of plant reproduction: Pattern and process. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 467–497. [Google Scholar] [CrossRef] [Green Version]
- Harder, L.D.; Aizen, M.A. Floral adaptation and diversification under pollen limitation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Eckert, C.G.; Ozimec, B.; Herlihy, C.R.; Griffin, C.A.; Routley, M.B. Floral morphology mediates temporal variation in the mating system of a self-compatible plant. Ecology 2009, 90, 1540–1548. [Google Scholar] [CrossRef]
- Teixido, A.L.; Aizen, M.A. Reproductive assurance weakens pollinator-mediated selection on flower size in an annual mixed-mating species. Ann. Bot. 2019, 123, 1067–1077. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Millard, J.; Outhwaite, C.L.; Kinnersley, R.; Freeman, R.; Gregory, R.D.; Adedoja, O.; Gavini, S.; Kioko, E.; Kuhlmann, M.; Ollerton, J.; et al. Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 2021, 12, 2902. [Google Scholar] [CrossRef] [PubMed]
- Zattara, E.E.; Aizen, M.A. Worldwide occurrence records suggest a global decline in bee species richness. One Earth 2021, 4, 114–123. [Google Scholar] [CrossRef]
- Outhwaite, C.L.; McCann, P.; Newbold, T. Agriculture and climate change are reshaping insect biodiversity worldwide. Nature 2022, 605, 97–102. [Google Scholar] [CrossRef]
- Li, Q.; Sun, M.; Liu, Y.; Liu, B.; Bianchi, F.J.J.A.; van der Werf, W.; Lu, Y. High pollination deficit and strong dependence on honeybees in pollination of korla fragrant pear, Pyrus sinkiangensis. Plants 2022, 11, 1734. [Google Scholar] [CrossRef]
- Aizen, M.A.; Aguiar, S.; Biesmeijer, J.C.; Garibaldi, L.A.; Inouye, D.W.; Jung, C.; Martins, D.J.; Medel, R.; Morales, C.L.; Ngo, H.; et al. Global agricultural productivity is threatened by increasing pollinator dependence without a parallel increase in crop diversification. Glob. Chang. Biol. 2019, 25, 3516–3527. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.R.; Singh, G.M.; Mozaffarian, D.; Myers, S.S. Effects of decreases of animal pollinators on human nutrition and global health: A modelling analysis. Lancet 2015, 386, 1964–1972. [Google Scholar] [CrossRef]
- Garibaldi, L.; Aizen, M.; Klein, A.; Cunningham, S.; Harder, L. Global growth and stability of agricultural yield decrease with pollinator dependence. Proc. Natl. Acad. Sci. USA 2011, 108, 5909–5914. [Google Scholar] [CrossRef]
- Tamburini, G.; Bommarco, R.; Kleijn, D.; van der Putten, W.H.; Marini, L. Pollination contribution to crop yield is often context-dependent: A review of experimental evidence. Agric. Ecosyst. Environ. 2019, 280, 16–23. [Google Scholar] [CrossRef]
- Reilly, J.R.; Artz, D.R.; Biddinger, D.; Bobiwash, K.; Boyle, N.K.; Brittain, C.; Brokaw, J.; Campbell, J.W.; Daniels, J.; Elle, E.; et al. Crop production in the USA is frequently limited by a lack of pollinators. Proc. R. Soc. B. Biol. Sci. 2020, 287, 20200922. [Google Scholar] [CrossRef] [PubMed]
- Holland, J.M.; Sutter, L.; Albrecht, M.; Jeanneret, P.; Pfister, S.C.; Schirmel, J.; Entling, M.H.; Kaasik, R.; Kovacs, G.; Veromann, E.; et al. Moderate pollination limitation in some entomophilous crops of Europe. Agric. Ecosyst. Environ. 2020, 302, 107002. [Google Scholar] [CrossRef]
- Deprá, M.S.; Delaqua, G.G.; Freitas, L.; Gaglianone, M.C. Pollination deficit in open-field tomato crops (Solanum lycopersicum L., Solanaceae) in Rio de Janeiro state, southeast Brazil. J. Pollinat. Ecol. 2014, 12, 1–8. [Google Scholar] [CrossRef]
- Bisui, S.; Layek, U.; Karmakar, P. Utilization of Indian dammar bee (Tetragonula iridipennis Smith) as a pollinator of bitter gourd. Acta Agrobot. 2020, 73, 7316. [Google Scholar] [CrossRef]
- Layek, U.; Kundu, A.; Karmakar, P. Floral ecology, floral visitors and breeding system of Gandharaj lemon (Citrus × limon L. Osbeck). Bot. Pac. 2020, 9, 113–119. [Google Scholar] [CrossRef]
- Layek, U.; Kundu, A.; Das, N.; Mondal, R.; Karmakar, P. Intercropping with pigeonpea (Cajanus cajan L. Millsp.): An assessment of its influence on the assemblage of pollinators and yield of neighbouring non-leguminous crops. Life 2023, 13, 193. [Google Scholar] [CrossRef]
- Norton, J.D. Testing of plum pollen viability with tetrazolium salts. Am. Soc. Hort. Sci. 1966, 89, 132–134. [Google Scholar]
- Zeisler, M. Uber die abgrenzung des eigentlichen narbenflache mit hilfe von reaktionen. Beih. Bot. Cent. 1933, 85, 308–318. [Google Scholar]
- Aizen, M.A.; Garibaldi, L.A.; Cunningham, S.A.; Klein, A.M. How much does agriculture depend on pollinators? Lessons from long-term trends in crop production. Ann. Bot. 2009, 103, 1579–1588. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Steffan-Dewenter, I.; Potts, S.G.; Packer, L. Pollinator diversity and crop pollination services are at risk. Trends Ecol. Evol. 2005, 20, 651–652. [Google Scholar] [CrossRef]
- Eilers, E.J.; Kremen, C.; Smith Greenleaf, S.; Garber, A.K.; Klein, A.M. Contribution of pollinator-mediated crops to nutrients in the human food supply. PLoS ONE 2011, 6, e21363. [Google Scholar]
- Toni, H.C.; Avohou, A.; Djossa, B.A. Contribution of animal pollination to food nutrient production in Benin-West Africa. J. Basic Appl. Zool. 2021, 82, 1–9. [Google Scholar] [CrossRef]
- Gallai, N.; Salles, J.M.; Settele, J.; Vaissière, B.E. Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol. Econ. 2009, 68, 810–821. [Google Scholar] [CrossRef]
- Denisow, B.; Wrzesien, M.; Cwener, A. Pollination and floral biology of Adonis vernalis L. (Ranunculaceae)-a case study of threatened species. Acta Soc. Bot. Pol. 2014, 83, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Cruden, R.W. Pollen-ovule ratios: A conservative indicator of breeding systems in flowering plants. Evolution 1977, 31, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Preston, R.E. Pollen-ovule ratios in the Cruciferae. Amer. J. Bot. 1986, 73, 1732–1740. [Google Scholar] [CrossRef]
- Gallardo, R.; Dominguez, E.; Muñoz, J.M. Pollen-ovule ratio, pollen size, and breeding system in Astragalus (Fabaceae) subgenus Epiglottis: A pollen and seed allocation approach. Amer. J. Bot. 1994, 81, 1611–1619. [Google Scholar] [CrossRef]
- Charnov, E.L. The theory of sex allocation; Princeton University Press: Princeton, NJ, USA, 1982. [Google Scholar]
- Queller, D.C. Pollen-ovule ratios and hermaphrodite sexual allocation strategies. Evolution 1984, 38, 1148–1151. [Google Scholar] [CrossRef] [Green Version]
- Pacini, E.; Franchi, G.G.; Lisci, M.; Nepi, M. Pollen viability related to type of pollination in six angiosperm species. Ann. Bot. 1997, 80, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Dafni, A.; Firmage, D.H. Pollen viability and longevity: Practical, ecological and evolutionary implications. Plant Syst. Evol. 2000, 222, 113–132. [Google Scholar] [CrossRef]
- Galen, C.; Plowright, R.C. Testing the accuracy of using peroxidase activity to indicate stigma receptivity. Canad. J. Bot. 1987, 65, 107–111. [Google Scholar] [CrossRef]
- Dafni, A.; Maués, M.M. A rapid and simple procedure to determine stigma receptivity. Sex. Plant Reprod. 1998, 11, 177–180. [Google Scholar] [CrossRef] [Green Version]
- Heslop-Harrison, J.; Heslop-Harrison, Y.; Barber, J. The stigma surface in incompatibility responses. Proc. R. Soc. B Biol. Sci. 1975, 188, 287–297. [Google Scholar]
- Kulloli, S.K.; Ramasubbu, R.; Sreekala, A.K.; Pandurangan, A.G. Cytochemical localization of stigma-surface esterases in three species of Impatiens (Balsaminaceae) of Western Ghats. Asia J. Exp. Biol. Sci. 2010, 1, 106–111. [Google Scholar]
- Byers, K.J.; Bradshaw, H.D., Jr.; Riffell, J.A. Three floral volatiles contribute to differential pollinator attraction in monkeyflowers (Mimulus). J. Exp. Biol. 2014, 217, 614–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rering, C.C.; Franco, J.G.; Yeater, K.M.; Mallinger, R.E. Drought stress alters floral volatiles and reduces floral rewards, pollinator activity, and seed set in a global plant. Ecosphere 2020, 11, e03254. [Google Scholar] [CrossRef]
- Layek, U.; Bisui, S.; Mondal, R.; Das, N.; De, S.K.; Karmakar, P. Floral traits and chemical cues associated with rock bee (Apis dorsata Fabricius) for the host selection in West Bengal, India. Grana 2021, 60, 310–323. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Value of IDP | Category |
|---|---|
| >0.9–1 | Obligatory dependence on biotic pollinators |
| 0.6–0.9 | Highly dependent on biotic pollinators |
| >0.3–<0.6 | Moderately dependent on biotic pollinators |
| 0.1–0.3 | Low dependence on biotic pollinators |
| <0.1 | Negligible dependence on biotic pollinators |
| Value of D | Category |
| >0.5 | High pollination deficit |
| 0.3–0.5 | Medium pollination deficit |
| 0.3 > D ≥ 0.1 | Low pollination deficit |
| <0.1 | Negligible pollination deficit |
| Crop Species | IDP | D |
|---|---|---|
| Foeniculum vulgare Mill. | 0.47 | 0.19 |
| Brassica juncea (L.) Czern. | 0.67 | 0.09 |
| Raphanus sativus (L.) Domin | 0.70 | 0.11 |
| Cucumis sativus L. | 1 | 0.17 |
| Cucurbita maxima Duchesne | 1 | 0.11 |
| Luffa aegyptiaca Mill. | 1 | 0.11 |
| Momordica charantia L. | 1 | 0.12 |
| Cajanas cajan L. Millsp. | 0.55 | 0.21 |
| Cicer arietinum L. | 0.45 | 0.12 |
| Lablab purpureus (L.) Sweet | 0.11 | 0.06 |
| Pisum sativum L. | 0.53 | 0.06 |
| Nigella sativa L. | 0.18 | 0.06 |
| Citrus × aurantiifolia (Christm.) Swingle | 0.74 | 0.15 |
| Citrus × limon L. Osbeck | 0.81 | 0.59 |
| Citrus maxima (Burm.) Merr. | 0.57 | 0.28 |
| Solanum lycopersicum L. | 0.47 | 0.13 |
| Solanum melongena L. | 0.48 | 0.14 |
| Crops | Flower Longevity (h) | Pollen/Flower | Ovule/Flower | Pollen–Ovule Ratio | Pollen Viability | Stigma Receptivity Time (h) | Visits/Flower |
|---|---|---|---|---|---|---|---|
| Foeniculum vulgare | 68.40 d ± 11.38 | 9783.92 e ± 1307.57 | 2.00 g ± 0 | 4891.96 | 85.63 ab ± 2.80 | 64.80 b ± 8.39 | 223.26 a ± 41.05 |
| Brassica juncea | 49.20 f ± 14.37 | 36,612.88 de ± 5952.24 | 20.20 g ± 0.95 | 1812.52 | 89.28 a ± 4.03 | 43.20 c ± 10.12 | 183.32 b ± 41.50 |
| Raphanus sativus | 52.80 ef ± 11.59 | 32,433.25 de ± 3554.95 | 7.50 g ± 0.89 | 4324.43 | 86.69 ab ± 4.46 | 44.40 c ± 8.10 | 172.97 bc ± 36.28 |
| Cucumis sativus | 7.80 h ± 1.48 | 7511.91 e ± 1026.68 | 79.85 def ± 11.59 | 94.08 | 75.53 efg ± 6.88 | 7.20 g ± 1.03 | 34.73 gh ± 17.10 |
| Cucurbita maxima | 7.20 h ± 1.40 | 112,829.10 c ± 17,942.11 | 248.70 b ± 35.83 | 453.68 | 88.49 a ± 5.98 | 6.60 g ± 0.97 | 38.88 gh ± 16.22 |
| Luffa aegyptiaca | 9.60 h ± 1.58 | 28,530.46 de ± 6572.95 | 176 c ± 28.43 | 162.10 | 77.72 def ± 5.39 | 9.20 fg ± 1.03 | 44.87 g ± 18.35 |
| Momordica charantia | 11.20 h ± 1.93 | 17,485.52 e ± 2817.90 | 22.05 g ± 3.02 | 792.99 | 69.93 h ± 3.95 | 10.80 fg ± 1.40 | 22.38 gh ± 6.75 |
| Cajanas cajan | 7.80 h ± 1.48 | 30,371.99 de ± 4500.18 | 4.95 g ± 0.83 | 6135.76 | 82.54 bcd ± 5.41 | 21.60 e ± 7.59 | 24.20 gh ± 8.82 |
| Cicer arietinum | 15.60 h ± 5.80 | 5835.61 e ± 842.80 | 2.35 g ± 0.49 | 2483.24 | 71.21 gh ± 3.38 | 14.40 f ± 5.06 | 38.56 gh ± 11.36 |
| Lablab purpureus | 33.60 g ± 8.58 | 28,419.86 de ± 5278.89 | 8.30 g ± 1.03 | 3424.08 | 86.55 ab ± 6.31 | 27.60 e ± 5.80 | 43.34 gh ± 11.88 |
| Pisum sativum | 34.80 g ± 8.39 | 34,224.06 de ± 5424.89 | 7.15 g ± 0.81 | 4786.58 | 84.74 abc ± 6.19 | 34.80 d ± 6.81 | 47.19 g ± 11.32 |
| Nigella sativa | 115.20 a ± 18.07 | 55,420.15 d ± 6804.97 | 96.25 de ± 13.57 | 575.79 | 71.31 gh ± 6.17 | 9.60 fg ± 1.58 | 172.80 bc ± 48.44 |
| Citrus × aurantiifolia | 97.20 b ± 14.37 | 7536.38 e ± 673.64 | 27.40 g ± 5.70 | 275.05 | 74.06 fgh ± 6.01 | 86.40 a ± 12.39 | 153.96 cd ± 25.53 |
| Citrus × limon | 79.20 c ± 11.59 | 9090.86 e ± 918.03 | 70.25 ef ± 9.50 | 129.41 | 85.18 ab ± 5.19 | 82.80 a ± 8.85 | 124.22 e ± 23.07 |
| Citrus maxima | 61.20 de ± 8.85 | 10,805.85 e ± 2996.74 | 104.25 d ± 13.18 | 103.65 | 84.37 abc ± 5.36 | 66 b ± 6.32 | 84.37 f ± 5.36 |
| Solanum lycopersicum | 13.80 h ± 1.75 | 345,440.30 b ± 81,326.76 | 56.75 f ± 9.04 | 6087.05 | 81.93 bcd ± 7.23 | 11 fg ± 1.41 | 17.39 h ± 9.29 |
| Solanum melongena | 58.80 e ± 14.37 | 681,055.60 a ± 95,092.40 | 2180.60 a ± 219.56 | 312.32 | 79.45 cde ± 6.92 | 45.60 c ± 12.39 | 141.47 de ± 38.01 |
| Reproductive Traits | Value of Correlation Coefficient (r) | |
|---|---|---|
| IDP | D | |
| Flower longevity | −0.398 | 0.227 |
| Pollen/flower | −0.185 | −0.119 |
| Ovule/flower | −0.082 | −0.035 |
| Pollen–ovule ratio | −0.430 | −0.253 |
| Pollen viability | −0.070 | 0.139 |
| Stigma receptivity time | −0.071 | 0.562 |
| Numberof visits received by a flower | −0.234 | 0.101 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Layek, U.; Baghira, N.K.; Das, A.; Kundu, A.; Karmakar, P. Dependency of Crops on Pollinators and Pollination Deficits: An Approach to Measurement Considering the Influence of Various Reproductive Traits. Agriculture 2023, 13, 1563. https://doi.org/10.3390/agriculture13081563
Layek U, Baghira NK, Das A, Kundu A, Karmakar P. Dependency of Crops on Pollinators and Pollination Deficits: An Approach to Measurement Considering the Influence of Various Reproductive Traits. Agriculture. 2023; 13(8):1563. https://doi.org/10.3390/agriculture13081563
Chicago/Turabian StyleLayek, Ujjwal, Nitol Krishna Baghira, Alokesh Das, Arijit Kundu, and Prakash Karmakar. 2023. "Dependency of Crops on Pollinators and Pollination Deficits: An Approach to Measurement Considering the Influence of Various Reproductive Traits" Agriculture 13, no. 8: 1563. https://doi.org/10.3390/agriculture13081563