
Effects and Underlying Mechanisms of Zearalenone Mycotoxin at Concentrations Close to the EC Recommendation on the Colon of Piglets after Weaning

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Toxin Preparation
2.2. Experimental Design
2.3. Quantification of Gene Expression
2.4. Quantification of Protein Expression by Western Blot
2.5. Quantification of Protein Concentration (ELISA)
2.6. Assessment of Oxidative Stress
2.6.1. Lipid Peroxidation Measurement (Thiobarbituric Acid Reactive Substances-TBARS)
2.6.2. Protein Oxidation Measurement (Protein Carbonyl Analysis)
2.6.3. DNA Fragmentation Using Diphenylamine (DPA)
2.7. Assessment of Antioxidant Response
2.7.1. Determination of Total Antioxidant Status (TAC)
2.7.2. Determination of Antioxidant Enzyme Activity
2.8. Statistical Analyses
3. Results
3.1. Effect of ZEN on Innate Immunity
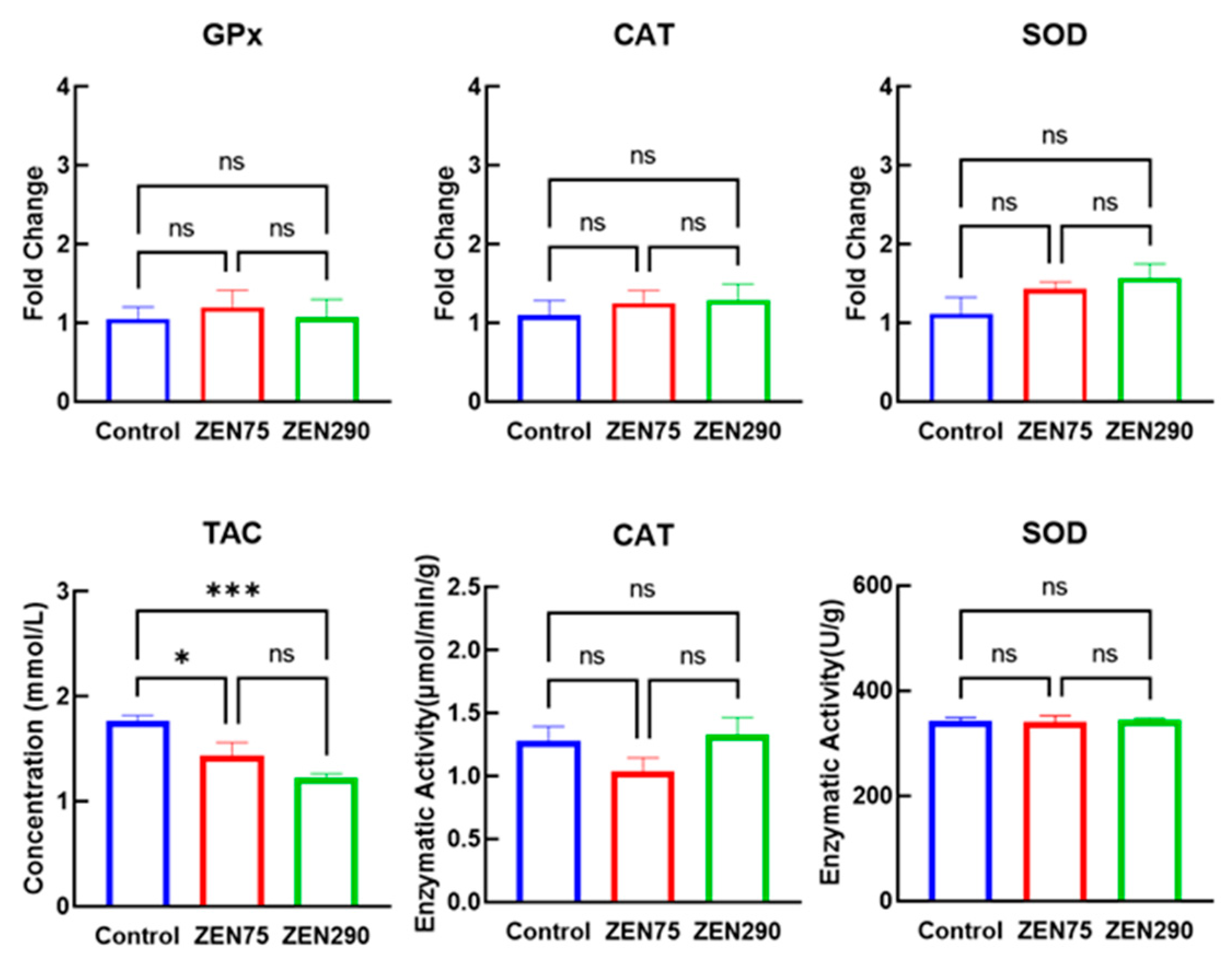
3.2. Effect of ZEN on Oxidative Response
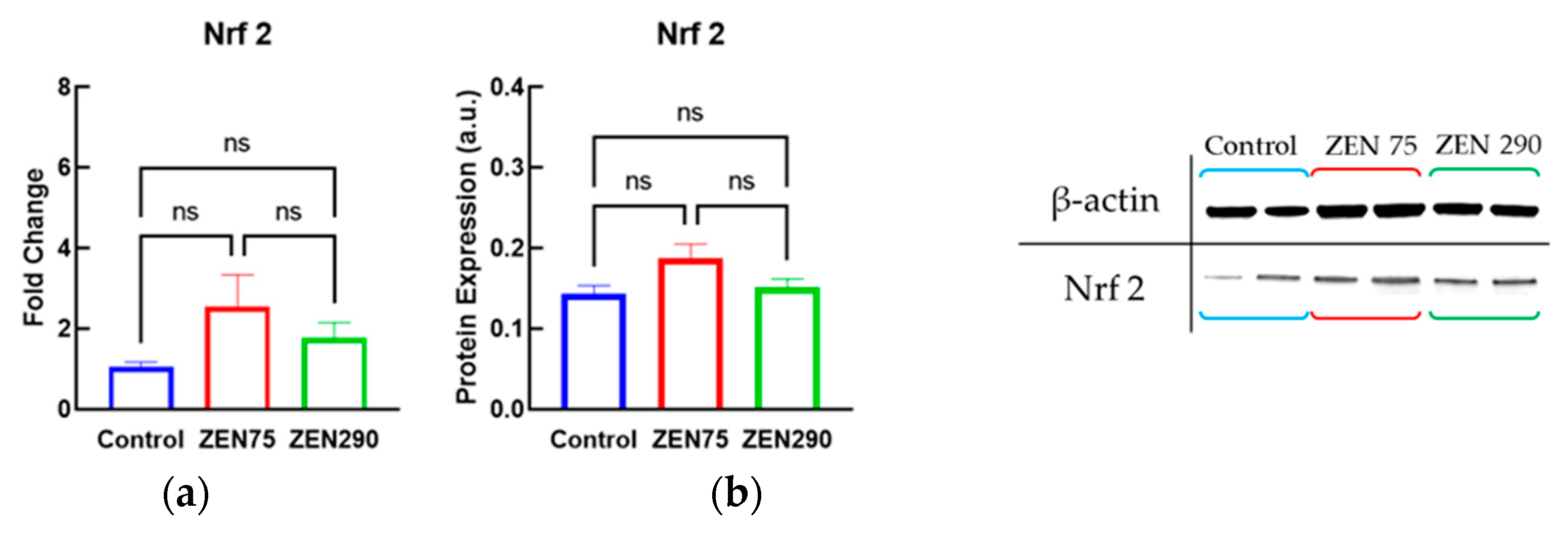
3.3. Effect of ZEN on Antioxidant Response
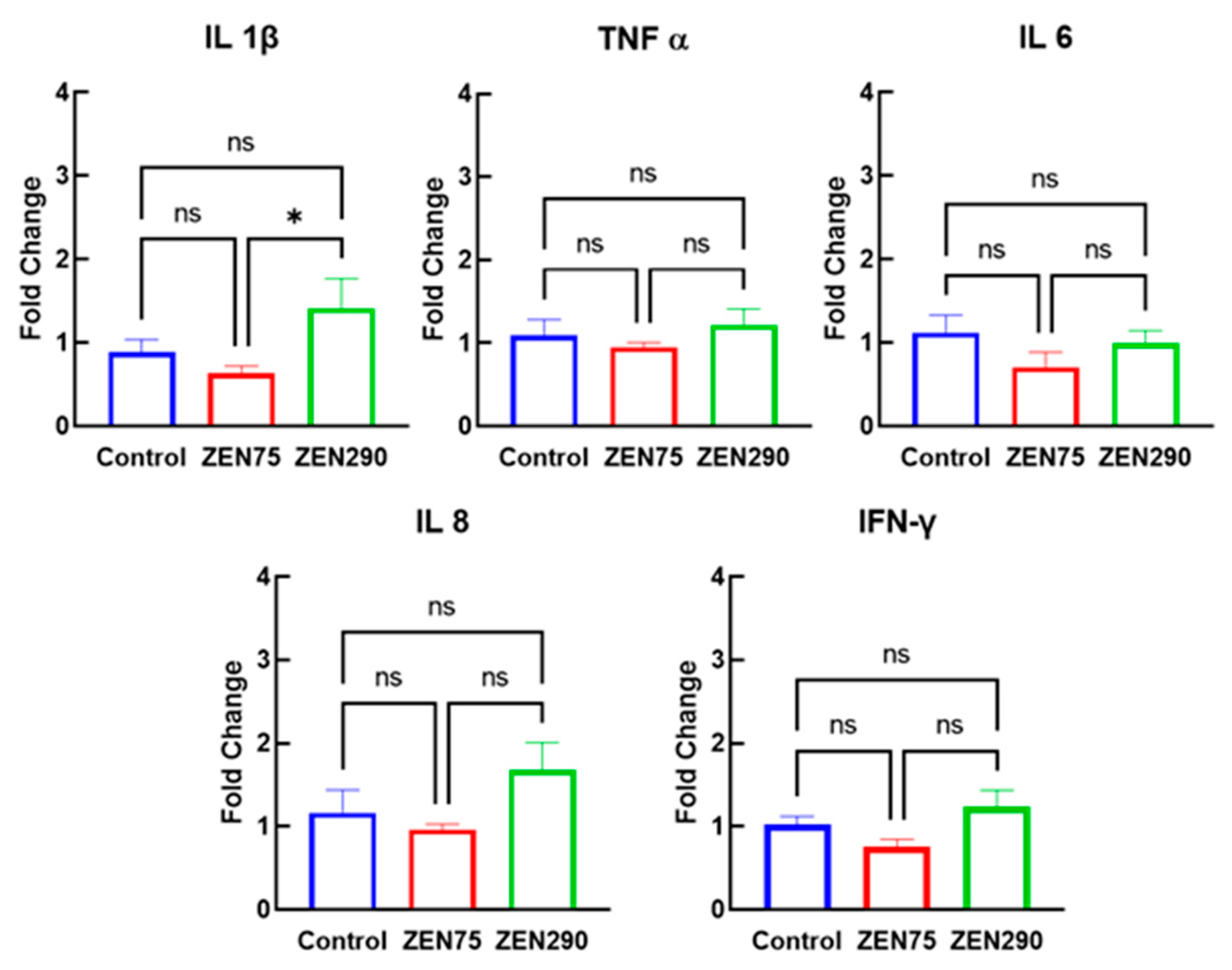
3.4. Effect of ZEN on Pro-Inflammatory Response
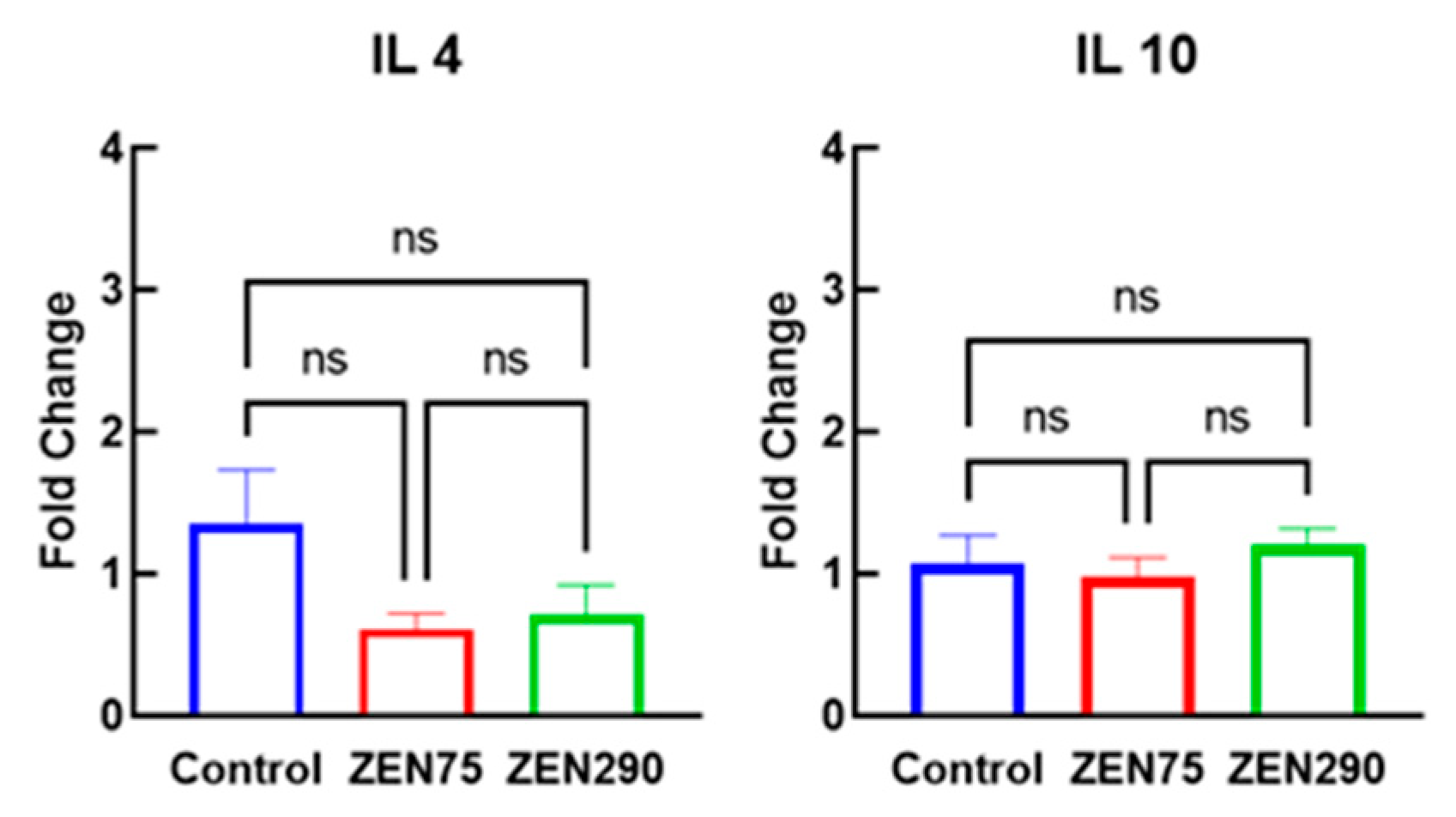
3.5. Effect of ZEN on Anti-Inflammatory Response
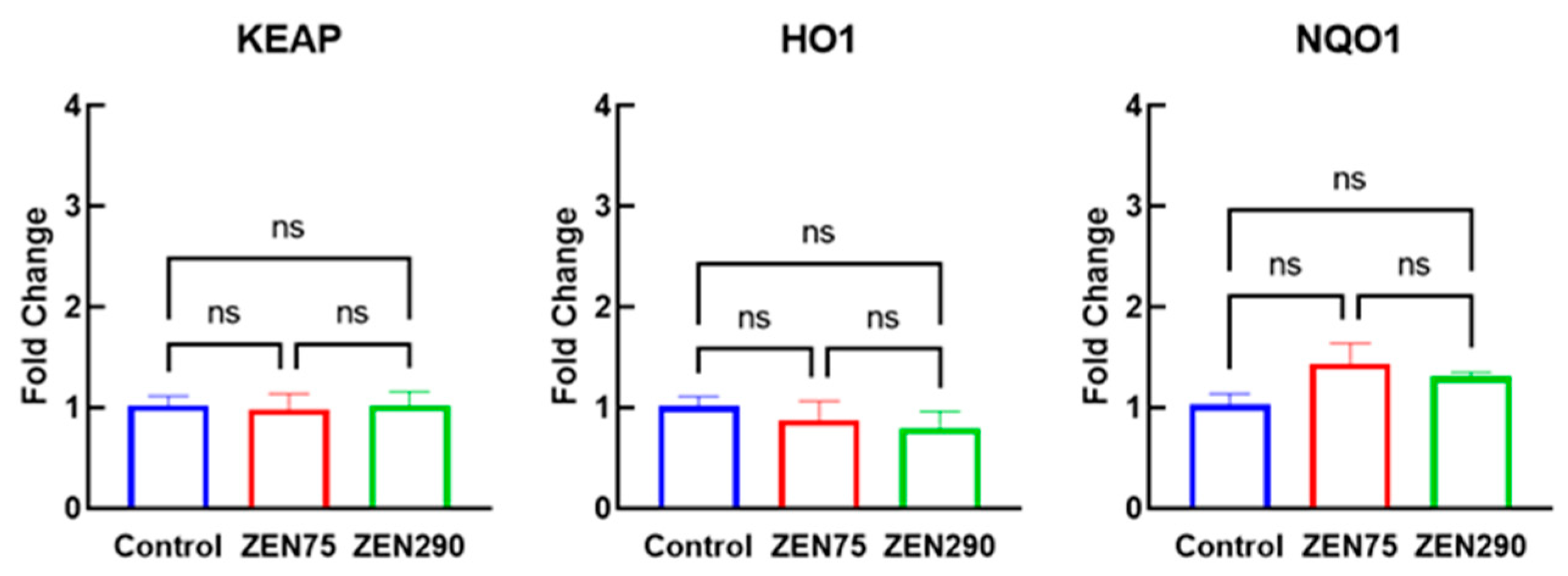
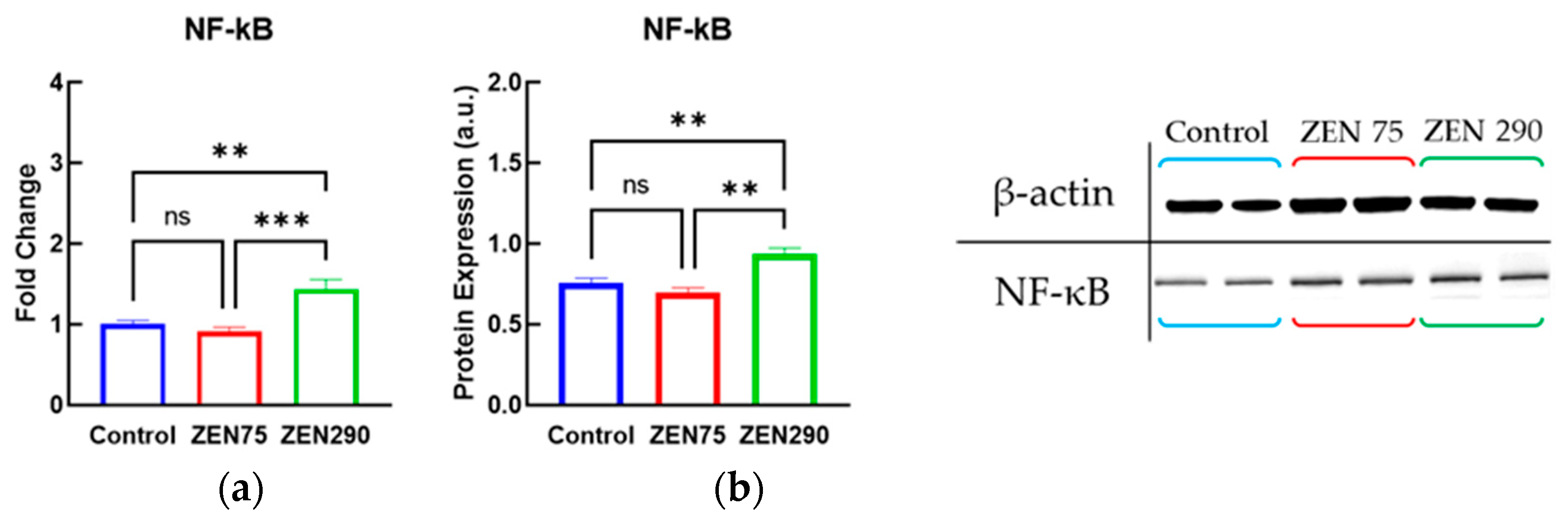
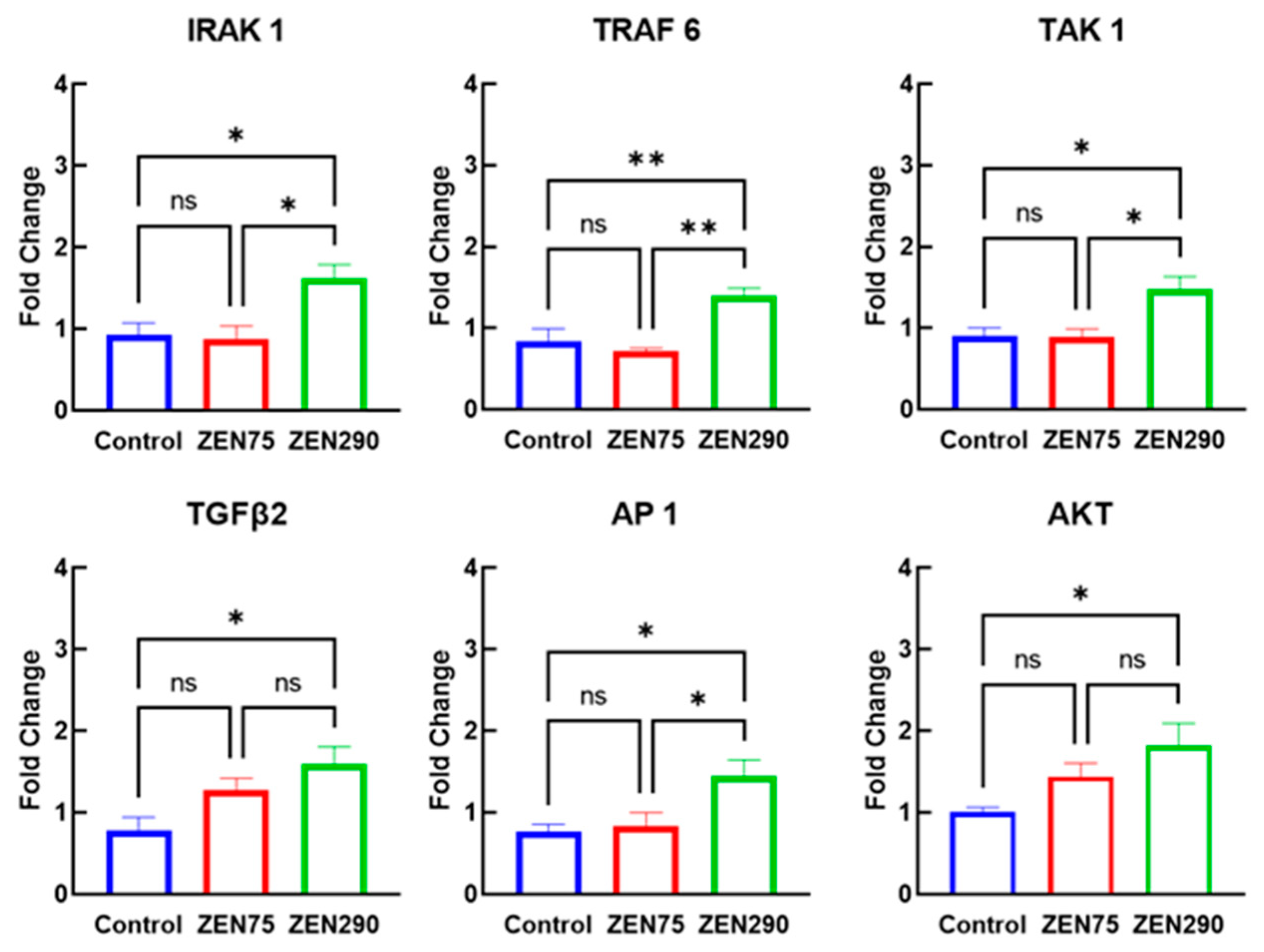
3.6. Effect of ZEN on Molecules Involved in Signaling Pathways
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Luo, S.; Terciolo, C.; Bracarense, A.P.F.L.; Payros, D.; Pinton, P.; Oswald, I.P. In Vitro and in Vivo Effects of a Mycotoxin, Deoxynivalenol, and a Trace Metal, Cadmium, Alone or in a Mixture on the Intestinal Barrier. Environ. Int. 2019, 132, 105082. [Google Scholar] [CrossRef] [PubMed]
- Bakker, M.G.; Brown, D.W.; Kelly, A.C.; Kim, H.-S.; Kurtzman, C.P.; Mccormick, S.P.; O’Donnell, K.L.; Proctor, R.H.; Vaughan, M.M.; Ward, T.J. Fusarium Mycotoxins: A Trans-Disciplinary Overview. Can. J. Plant Pathol. 2018, 40, 161–171. [Google Scholar] [CrossRef]
- Binder, S.B.; Schwartz-Zimmermann, H.E.; Varga, E.; Bichl, G.; Michlmayr, H.; Adam, G.; Berthiller, F. Metabolism of Zearalenone and Its Major Modified Forms in Pigs. Toxins 2017, 9, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frizzell, C.; Ndossi, D.; Verhaegen, S.; Dahl, E.; Eriksen, G.; Sørlie, M.; Ropstad, E.; Muller, M.; Elliott, C.T.; Connolly, L. Endocrine Disrupting Effects of Zearalenone, Alpha- and Beta-Zearalenol at the Level of Nuclear Receptor Binding and Steroidogenesis. Toxicol. Lett. 2011, 206, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Jia, R.; Liu, W.; Zhao, L.; Cao, L.; Shen, Z. Low Doses of Individual and Combined Deoxynivalenol and Zearalenone in Naturally Moldy Diets Impair Intestinal Functions via Inducing Inflammation and Disrupting Epithelial Barrier in the Intestine of Piglets. Toxicol. Lett. 2020, 333, 159–169. [Google Scholar] [CrossRef]
- Rogowska, A.; Pomastowski, P.; Sagandykova, G.; Buszewski, B. Zearalenone and Its Metabolites: Effect on Human Health, Metabolism and Neutralisation Methods. Toxicon 2019, 162, 46–56. [Google Scholar] [CrossRef]
- Reddy, K.E.; Jeong, J.Y.; Song, J.; Lee, Y.; Lee, H.-J.; Kim, D.-W.; Jung, H.-A.; Kim, K.H.; Kim, M.; Oh, Y.; et al. Colon Microbiome of Pigs Fed Diet Contaminated with Commercial Purified Deoxynivalenol and Zearalenone. Toxins 2018, 10, 347. [Google Scholar] [CrossRef] [Green Version]
- Commission Recommendation (EU) 576/2006. Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding (Text with EEA relevance). Off. J. Eur. Union 2006. [Google Scholar]
- Braicu, C.; Cojocneanu-Petric, R.; Jurj, A.; Gulei, D.; Taranu, I.; Gras, A.M.; Marin, D.E.; Berindan-Neagoe, I. Microarray Based Gene Expression Analysis of Sus Scrofa Duodenum Exposed to Zearalenone: Significance to Human Health. BMC Genom. 2016, 17, 646. [Google Scholar] [CrossRef] [Green Version]
- Taranu, I.; Gras, M.A.; Pistol, G.C.; Motiu, M.; Marin, D.; Stancu, M. Investigation of Zearalenone Tolerance Limit in the Feedstuffs for Weaned Pigs. Sci. Pap. Anim. Sci. Ser. 2016, 65, 14–18. [Google Scholar]
- Gajęcka, M.; Waśkiewicz, A.; Zielonka, Ł.; Goliński, P.; Rykaczewska, A.; Lisieska-Żołnierczyk, S.; Gajęcki, M.T. Mycotoxin Levels in the Digestive Tissues of Immature Gilts Exposed to Zearalenone and Deoxynivalenol. Toxicon 2018, 153, 1–11. [Google Scholar] [CrossRef]
- Jo, H.; Kong, C.; Song, M.; Kim, B.G. Effects of Dietary Deoxynivalenol and Zearalenone on Apparent Ileal Digestibility of Amino Acids in Growing Pigs. Anim. Feed Sci. Technol. 2016, 219, 77–82. [Google Scholar] [CrossRef]
- Xiong, X.; Tan, B.; Song, M.; Ji, P.; Kim, K.; Yin, Y.; Liu, Y. Nutritional Intervention for the Intestinal Development and Health of Weaned Pigs. Front. Vet. Sci. 2019, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Blachier, F.; Andriamihaja, M.; Kong, X.-F. Fate of Undigested Proteins in the Pig Large Intestine: What Impact on the Colon Epithelium? Anim. Nutr. 2022, 9, 110–118. [Google Scholar] [CrossRef]
- Richards, J.D.; Gong, J.; de Lange, C.F.M. The Gastrointestinal Microbiota and Its Role in Monogastric Nutrition and Health with an Emphasis on Pigs: Current Understanding, Possible Modulations, and New Technologies for Ecological Studies. Can. J. Anim. Sci. 2005, 85, 421–435. [Google Scholar] [CrossRef]
- Boudry, G.; Péron, V.; Le Huërou-Luron, I.; Lallès, J.P.; Sève, B. Weaning Induces Both Transient and Long-Lasting Modifications of Absorptive, Secretory, and Barrier Properties of Piglet Intestine. J. Nutr. 2004, 134, 2256–2262. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.M.; Crenshaw, J.D.; Polo, J. The Biological Stress of Early Weaned Piglets. J. Anim. Sci. Biotechnol. 2013, 4, 19. [Google Scholar] [CrossRef] [Green Version]
- Panah, F.M.; Lauridsen, C.; Højberg, O.; Jensen, H.E.; Nielsen, T.S. Composition of Mucus- and Digesta-Associated Bacteria in Growing Pigs with and without Diarrhea Differed According to the Presence of Colonic Inflammation. BMC Microbiol. 2023, 23, 145. [Google Scholar] [CrossRef] [PubMed]
- Bauer, E.; Williams, B.A.; Smidt, H.; Verstegen, M.W.A.; Mosenthin, R. Influence of the Gastrointestinal Microbiota on Development of the Immune System in Young Animals. Curr. Issues Intest. Microbiol. 2006, 7, 35–52. [Google Scholar] [PubMed]
- Grosu, I.A.; Bulgaru, C.V.; Pistol, G.C.; Cismileanu, A.; Marin, D.E.; Taranu, I. Effects of Exposure to Low Zearalenone Concentrations Close to the EU Recommended Value on Weaned Piglets’ Colon. Toxins 2023, 15, 206. [Google Scholar] [CrossRef] [PubMed]
- Pistol, G.C.; Marin, D.E.; Rotar, M.C.; Ropota, M.; Taranu, I. Bioactive Compounds from Dietary Whole Grape Seed Meal Improved Colonic Inflammation via Inhibition of MAPKs and NF-KB Signaling in Pigs with DSS Induced Colitis. J. Funct. Foods 2020, 66, 103708. [Google Scholar] [CrossRef]
- Marin, D.E.; Pistol, G.C.; Gras, M.; Palade, M.; Taranu, I. A Comparison between the Effects of Ochratoxin A and Aristolochic Acid on the Inflammation and Oxidative Stress in the Liver and Kidney of Weanling Piglets. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Ben Salah-Abbès, J.; Belgacem, H.; Ezzdini, K.; Abdel-Wahhab, M.A.; Abbès, S. Zearalenone Nephrotoxicity: DNA Fragmentation, Apoptotic Gene Expression and Oxidative Stress Protected by Lactobacillus Plantarum MON03. Toxicon 2020, 175, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Taranu, I.; Pistol, G.C.; Palade, M.L.; Bulgaru, C.V.; Habeanu, M.; Anghel, A.C.; Marin, D. Dietary Inclusion of Saccharomyces Cerevisiae Fermented Rapeseed Meal Modulated Immune, Oxidant and Antioxidant Indices in Piglets after Weaning. In Scientific Papers: Series D, Animal Science-The International Session of Scientific Communications of the Faculty of Animal Science; University of Agronomic Sciences and Veterinary Medicine of Bucharest: Bucharest, Romania, 2022; Volume 65. [Google Scholar]
- Chedea, V.S.; Palade, L.M.; Pelmus, R.S.; Dragomir, C.; Taranu, I. Red Grape Pomace Rich in Polyphenols Diet Increases the Antioxidant Status in Key Organs—Kidneys, Liver, and Spleen of Piglets. Animals 2019, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, J.; Duan, H.; Li, R.; Peng, W.; Wu, C. Activation of Nrf2/HO-1 Signaling: An Important Molecular Mechanism of Herbal Medicine in the Treatment of Atherosclerosis via the Protection of Vascular Endothelial Cells from Oxidative Stress. J. Adv. Res. 2021, 34, 43–63. [Google Scholar] [CrossRef]
- Bulgaru, C.V.; Marin, D.E.; Pistol, G.C.; Taranu, I. Zearalenone and the Immune Response. Toxins 2021, 13, 248. [Google Scholar] [CrossRef]
- Zinedine, A.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Review on the Toxicity, Occurrence, Metabolism, Detoxification, Regulations and Intake of Zearalenone: An Oestrogenic Mycotoxin. Food Chem. Toxicol. 2007, 45, 1–18. [Google Scholar] [CrossRef]
- Ropejko, K.; Twarużek, M. Zearalenone and Its Metabolites—General Overview, Occurrence, and Toxicity. Toxins 2021, 13, 35. [Google Scholar] [CrossRef]
- Koenderman, L.; Buurman, W.; Daha, M.R. The Innate Immune Response. Immunol. Lett. 2014, 162, 95–102. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Pathogen Recognition in the Innate Immune Response. Biochem. J. 2009, 420, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R. Toll-like Receptors and Innate Immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- Jia, Z.; Liu, M.; Qu, Z.; Zhang, Y.; Yin, S.; Shan, A. Toxic Effects of Zearalenone on Oxidative Stress, Inflammatory Cytokines, Biochemical and Pathological Changes Induced by This Toxin in the Kidney of Pregnant Rats. Environ. Toxicol. Pharmacol. 2014, 37, 580–591. [Google Scholar] [CrossRef]
- Taranu, I.; Marin, D.E.; Pistol, G.C.; Motiu, M.; Pelinescu, D. Induction of Pro-Inflammatory Gene Expression by Escherichia Coli and Mycotoxin Zearalenone Contamination and Protection by a Lactobacillus Mixture in Porcine IPEC-1 Cells. Toxicon 2015, 97, 53–63. [Google Scholar] [CrossRef]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like Receptors and Innate Immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef]
- Camhi, S.L.; Lee, P.; Choi, A.M. The Oxidative Stress Response. New Horiz. 1995, 3, 170–182. [Google Scholar]
- Liu, X.; Xu, C.; Yang, Z.; Yang, W.; Huang, L.; Wang, S.; Liu, F.; Liu, M.; Wang, Y.; Jiang, S. Effects of Dietary Zearalenone Exposure on the Growth Performance, Small Intestine Disaccharidase, and Antioxidant Activities of Weaned Gilts. Animals 2020, 10, 2157. [Google Scholar] [CrossRef]
- Qin, X.; Cao, M.; Lai, F.; Yang, F.; Ge, W.; Zhang, X.; Cheng, S.; Sun, X.; Qin, G.; Shen, W.; et al. Oxidative Stress Induced by Zearalenone in Porcine Granulosa Cells and Its Rescue by Curcumin In Vitro. PLoS ONE 2015, 10, e0127551. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Gao, J.; Liu, F.X.; Broomhead, J.; Chi, F. Effects of Purified Zearalenone on Growth Performance, Organ Size, Serum Metabolites, and Oxidative Stress in Postweaning Gilts1. J. Anim. Sci. 2011, 89, 3008–3015. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.E.; Pistol, G.C.; Neagoe, I.V.; Calin, L.; Taranu, I. Effects of Zearalenone on Oxidative Stress and Inflammation in Weanling Piglets. Food Chem. Toxicol. 2013, 58, 408–415. [Google Scholar] [CrossRef]
- Nathan, C. Points of Control in Inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Targets-Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Nordgreen, J.; Edwards, S.A.; Boyle, L.A.; Bolhuis, J.E.; Veit, C.; Sayyari, A.; Marin, D.E.; Dimitrov, I.; Janczak, A.M.; Valros, A. A Proposed Role for Pro-Inflammatory Cytokines in Damaging Behavior in Pigs. Front. Vet. Sci. 2020, 7, 646. [Google Scholar] [CrossRef] [PubMed]
- Oswald, I.P. Role of Intestinal Epithelial Cells in the Innate Immune Defence of the Pig Intestine. Vet. Res. 2006, 37, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marin, D.E.; Motiu, M.; Taranu, I. Food Contaminant Zearalenone and Its Metabolites Affect Cytokine Synthesis and Intestinal Epithelial Integrity of Porcine Cells. Toxins 2015, 7, 1979–1988. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.E.; Taranu, I.; Burlacu, R.; Manda, G.; Motiu, M.; Neagoe, I.; Dragomir, C.; Stancu, M.; Calin, L. Effects of Zearalenone and Its Derivatives on Porcine Immune Response. Toxicol. Vitr. 2011, 25, 1981–1988. [Google Scholar] [CrossRef]
- Liu, M.; Zhu, D.; Guo, T.; Zhang, Y.; Shi, B.; Shan, A.; Chen, Z. Toxicity of Zearalenone on the Intestines of Pregnant Sows and Their Offspring and Alleviation with Modified Halloysite Nanotubes. J. Sci. Food Agric. 2018, 98, 698–706. [Google Scholar] [CrossRef]
- Benthem de Grave, X.; Saltzmann, J.; Laurain, J.; Rodriguez, M.A.; Molist, F.; Dänicke, S.; Santos, R.R. Transmission of Zearalenone, Deoxynivalenol, and Their Derivatives from Sows to Piglets during Lactation. Toxins 2021, 13, 37. [Google Scholar] [CrossRef]
- Lingappan, K. NF-ΚB in Oxidative Stress. Curr. Opin. Toxicol. 2018, 7, 81–86. [Google Scholar] [CrossRef]
- Arsura, M.; Panta, G.R.; Bilyeu, J.D.; Cavin, L.G.; Sovak, M.A.; Oliver, A.A.; Factor, V.; Heuchel, R.; Mercurio, F.; Thorgeirsson, S.S.; et al. Transient Activation of NF-ΚB through a TAK1/IKK Kinase Pathway by TGF-Β1 Inhibits AP-1/SMAD Signaling and Apoptosis: Implications in Liver Tumor Formation. Oncogene 2003, 22, 412–425. [Google Scholar] [CrossRef] [Green Version]
- Shen, T.; Miao, Y.; Ding, C.; Fan, W.; Liu, S.; Lv, Y.; Gao, X.; De Boevre, M.; Yan, L.; Okoth, S.; et al. Activation of the P38/MAPK Pathway Regulates Autophagy in Response to the CYPOR-Dependent Oxidative Stress Induced by Zearalenone in Porcine Intestinal Epithelial Cells. Food Chem. Toxicol. 2019, 131, 110527. [Google Scholar] [CrossRef]
- Pistol, G.C.; Braicu, C.; Motiu, M.; Gras, M.A.; Marin, D.E.; Stancu, M.; Calin, L.; Israel-Roming, F.; Berindan-Neagoe, I.; Taranu, I. Zearalenone Mycotoxin Affects Immune Mediators, MAPK Signalling Molecules, Nuclear Receptors and Genome-Wide Gene Expression in Pig Spleen. PLoS ONE 2015, 10, e0127503. [Google Scholar] [CrossRef] [Green Version]
- Pistol, G.C.; Gras, M.A.; Marin, D.E.; Israel-Roming, F.; Stancu, M.; Taranu, I. Natural Feed Contaminant Zearalenone Decreases the Expressions of Important Pro- and Anti-Inflammatory Mediators and Mitogen-Activated Protein Kinase/NF-ΚB Signalling Molecules in Pigs. Br. J. Nutr. 2014, 111, 452–464. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Lv, Y.; Ren, S.; Shao, M.; Shen, T.; Huang, K.; Zhou, J.; Yan, L.; Song, S. Zearalenone (ZEA)-Induced Intestinal Inflammation Is Mediated by the NLRP3 Inflammasome. Chemosphere 2018, 190, 272–279. [Google Scholar] [CrossRef]
- Zheng, D.; Kern, L.; Elinav, E. The NLRP6 Inflammasome. Immunology 2021, 162, 281–289. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions, and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions, or products referred to in the content. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Orientation | Tm (°C) | Amplicon Length (bp) |
|---|---|---|---|---|
| β-2 microglobulin | TTCTACCTTCTGGTCCACACTGA | 5′-3′ | 50 | 162 |
| TCATCCAACCCAGATGCA | 3′-5′ | 54 | ||
| GAPDH | ACTCACTCTTCTACCTTTGATGCT | 5′-3′ | 49 | 100 |
| TGTTGCTGTAGCCAAATTCA | 3′-5′ | 56 | ||
| Cyclophilin A | CCCACCGTCTTCTTCGACAT | 5′-3′ | 54 | 92 |
| TCTGCTGTCTTTGGAACTTTGTCT | 3′-5′ | 55 | ||
| β-actin | GGACTTCGAGCAGGAGATGG | 5′-3′ | 60 | 230 |
| GCACCGTGTTTGCGTAGAGG | 3′-5′ | 62 | ||
| HGPRT-1 | TGGAAAGAATGTCTTGATTGTTGAAG | 5′-3′ | 58.57 | 93 |
| ATCTTTGGATTATGCTGCTTGACC | 3′-5′ | 59.66 | ||
| RPL 32 | TGCTCTCAGACCCCTTGTGAAG | 5′-3′ | 61.93 | 106 |
| TTTCCGCCAGTTCCGCTTA | 3′-5′ | 59.63 | ||
| TNF-α | ACTGCACTTCGAGGTTATCGG | 5′-3′ | 60 | 118 |
| GGCGACGGGCTTATCTGA | 3′- 5′ | 60 | ||
| IL-8 | GCTCTCTGTGAGGCTGCAGTTC | 5′-3′ | 58 | 79 |
| AAGGTGTGGAATGCGTATTTATGC | 3′-5′ | 54 | ||
| IL-6 | GGCAAAAGGGAAAGAATCCAG | 5′-3′ | 57 | 87 |
| CGTTCTGTGACTGCAGCTTATCC | 3′-5′ | 61 | ||
| IL-1β | ATGCTGAAGGCTCTCCACCTC | 5′-3′ | 62 | 89 |
| TTGTTGCTATCATCTCCTTGCAC | 3′-5′ | 59 | ||
| IFNγ | TGGTAGCTCTGGGAAACTGAATG | 5′-3′ | 54 | 79 |
| GGCTTTGCGCTGGATCTG | 3′-5′ | 55 | ||
| IL-4 | CAACCCTGGTCTGCTTACTG | 5′-3′ | 52 | 173 |
| CTTCTCCGTCGTGTTCTCTG | 3′-5′ | 52 | ||
| IL-10 | GGCCCAGTGAAGAGTTTCTTTC | 5′-3′ | 54 | 51 |
| CAACAAGTCGCCCATCTGGT | 3′-5′ | 55 | ||
| NF-kB | CGAGAGGAGCACGGATACCA | 5′-3′ | 55 | 62 |
| GCCCCGTGTAGCCATTGA | 3′-5′ | 54 | ||
| CAT | CTTGGAACATTGTACCCGCT | 5′-3′ | 62 | 241 |
| GTCCAGAAGAGCCTGAATGC | 3′-5′ | 62 | ||
| GPx | GGAGATCCTGAATTGCCTCAAG | 5′-3′ | 50 | 62 |
| GCATGAAGTTGGGCTCGAA | 3′-5′ | 58 | ||
| SOD | GAGACCTGGGCAATGTGACT | 5′-3′ | 62 | 139 |
| CTGCCCAAGTCATCTGGTTT | 3′-5′ | 60 | ||
| Nrf2 | CCCATTCACAAAAGACAAACATTC | 5′-3′ | 57 | 72 |
| GCTTTTGCCCTTAGCTCATCTC | 3′-5′ | 59 | ||
| AP1 | CCCAAGATCCTGAAGCAGAG | 5′-3′ | 62 | 136 |
| GATGTGCCCGTTACTGGACT | 3′-5′ | 62 | ||
| p38α | TGCAAGGTCTCTGGAGGAAT | 5′-3′ | 52 | 109 |
| CTGAACGTGGTCATCCGTAA | 3′-5′ | 52 | ||
| TGFβ2 | CGATGATGATGTTGATGATGG | 5′-3′ | 55 | 69 |
| GCAAGGCTTTCTTGTATTTTCTTG | 3′-5′ | 58 | ||
| TAK1 | TGCCCAAACTCCAAAGAATC | 5′-3′ | 56 | 151 |
| TTTGCTGGTCCTTTTCATCC | 3′-5′ | 56 | ||
| ERK1 | CTACCTGGACCAGCTCAACC | 5′-3′ | 60 | 85 |
| CACTGTGATCCGTTTGTTGG | 3′-5′ | 60 | ||
| ERK2 | TGACATTCAACCCTCACAAGA | 5′-3′ | 42.86 | 198 |
| ATCTGTATCCTGGCTGGAATC | 3′-5′ | 47.62 | ||
| JNK1 | TGCTTTGTGGAATCAAGCAC | 5′-3′ | 51 | 60 |
| TGGGCTTTAAGTCCCGATG | 3′-5′ | 51 | ||
| JNK2 | TATTATCGGGCACCAGAAGTC | 5′-3′ | 51 | 97 |
| AACCTTTCACCAGCTCTCTCA | 3′- 5′ | 53 | ||
| JNK 3 | TGCCTATGACGCTGTTCTTG | 5′-3′ | 58.27 | 180 |
| TGAAACTCCTCCAGCGTCTT | 3′-5′ | 58.95 | ||
| HO-1 | ATGTGAATGCAACCCTGTGA | 5′-3′ | 57.71 | 89 |
| GGAAGCCAGTCAAGAGACCA | 3′-5′ | 59.31 | ||
| NQO1 | GTATCCTGCCGAGACTGCTC | 5′-3′ | 59.97 | 134 |
| TAGCAGGGACTCCAAACCAC | 3′-5′ | 59.31 | ||
| KEAP | ACGACGTGGAGACAGAAACGT | 5′-3′ | 61.94 | 56 |
| GCTTCGCCGATGCTTCA | 3′-5′ | 58.07 | ||
| Akt | AAGGCCACGGGCCGCTACTA | 5′-3′ | 65.94 | 100 |
| GGAGGACGCGGTTCTCCGT | 3′-5′ | 64.2 | ||
| MyD88 | GCAGCTGGAACAGACCAACT | 5′-3′ | 60 | 66 |
| GTGCCAGGCAGGACATCT | 3′-5′ | 59 | ||
| MD-2 | CCTTGTTTTCTTCCATATTTACTG | 5′-3′ | 54 | 63 |
| CATCAGAGGAATTGCAGATCCA | 3′- 5′ | 58 | ||
| IRAK1 | CAAGGCAGGTCAGGTTTCGT | 5′-3′ | 55 | 115 |
| TTCGTGGGGCGTGTAGTGT | 3′-5′ | 58 | ||
| TRAF-6 | CAAGAGAATACCCAGTCGCACA | 5′-3′ | 50 | 122 |
| ATCCGAGACAAAGGGGAAGAA | 3′-5′ | 48 | ||
| TLR2 | TCACTTGTCTAACTTATCATCCTCTTG | 5′-3′ | 59 | 162 |
| TCAGCGAAGGTGTCATTATTGC | 3′-5′ | 59 | ||
| TLR4 | GCCATCGCTGCTAACATCATC | 5′-3′ | 60 | 108 |
| CTCATACTCAAAGATACACCATCGG | 3′-5′ | 59 | ||
| TLR5 | CCTTCCTGCTTCTTTGATGG | 5′-3′ | 56 | 124 |
| CTGTGACCGTCCTGATGTAG | 3′-5′ | 57 | ||
| TLR9 | CACGACAGCCGAATAGCAC | 5′-3′ | 59 | 121 |
| GGGAACAGGGAGCAGAGC | 3′-5′ | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bulgaru, V.C.; Pertea, A.M.; Grosu, I.A.; Anghel, A.C.; Pistol, G.C.; Marin, D.E.; Dinischiotu, A.; Taranu, I. Effects and Underlying Mechanisms of Zearalenone Mycotoxin at Concentrations Close to the EC Recommendation on the Colon of Piglets after Weaning. Agriculture 2023, 13, 1372. https://doi.org/10.3390/agriculture13071372
Bulgaru VC, Pertea AM, Grosu IA, Anghel AC, Pistol GC, Marin DE, Dinischiotu A, Taranu I. Effects and Underlying Mechanisms of Zearalenone Mycotoxin at Concentrations Close to the EC Recommendation on the Colon of Piglets after Weaning. Agriculture. 2023; 13(7):1372. https://doi.org/10.3390/agriculture13071372
Chicago/Turabian StyleBulgaru, Valeria Cristina, Ana Maria Pertea, Iulian Alexandru Grosu, Andrei Cristian Anghel, Gina Cecilia Pistol, Daniela Eliza Marin, Anca Dinischiotu, and Ionelia Taranu. 2023. "Effects and Underlying Mechanisms of Zearalenone Mycotoxin at Concentrations Close to the EC Recommendation on the Colon of Piglets after Weaning" Agriculture 13, no. 7: 1372. https://doi.org/10.3390/agriculture13071372