
Effect of Dietary Starch-to-Fat Ratio on Lipid Metabolism, Inflammation, and Microbiota of Multiparous Sow and Newborn Piglets

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Design
2.2. Sow Performance
2.3. Sample Collection
2.4. Diets and Feces Nutrients Analysis
2.5. Metabolic Biomarkers Analysis
2.6. Fecal Microbiota Analysis
2.7. Statistical Analyses
3. Results
3.1. Sow Performance
3.2. Parameters Related to Glucolipid Metabolism
3.3. Antioxidant Enzymes and Inflammatory Cytokines
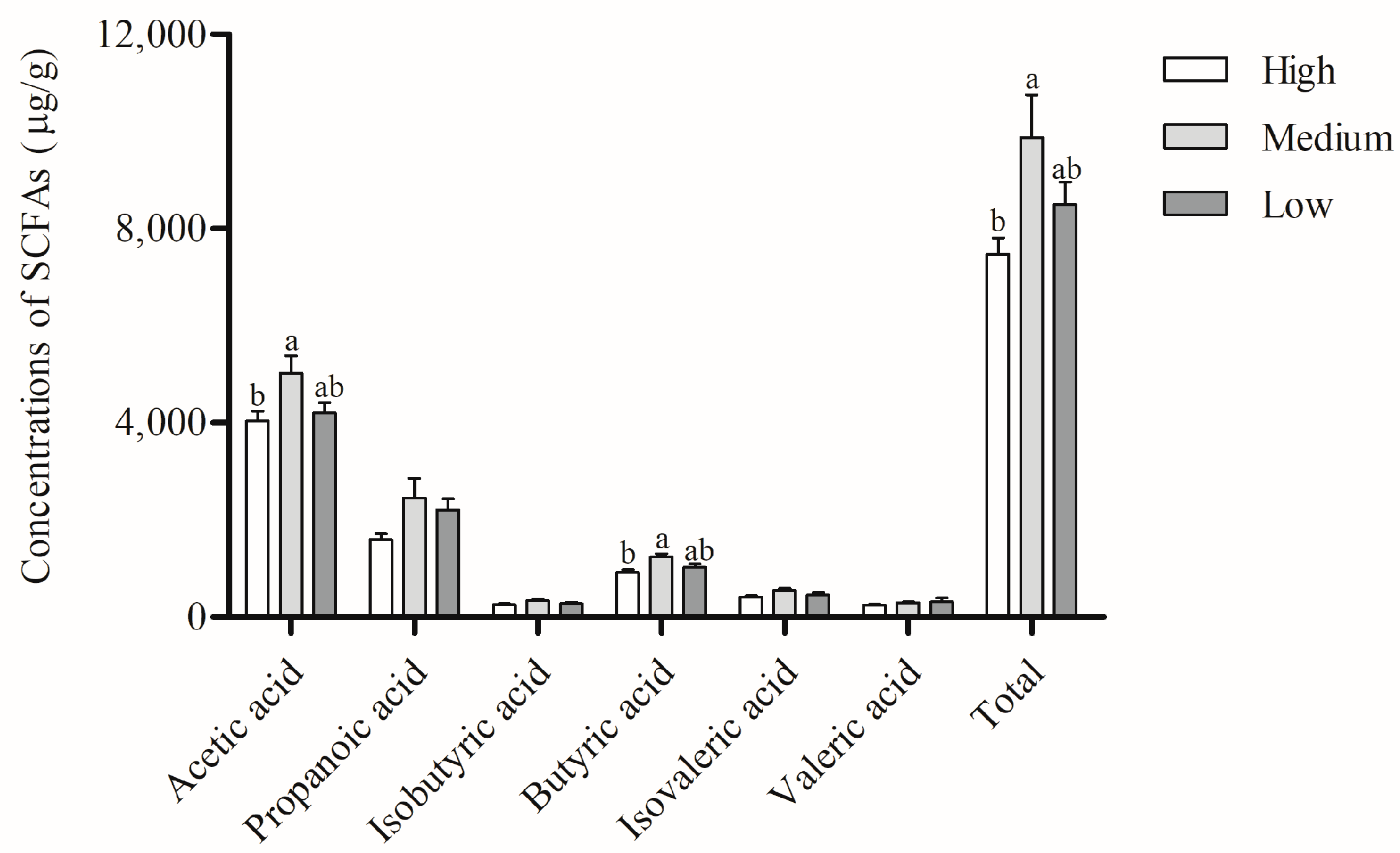
3.4. SCFAs in Feces of Sows
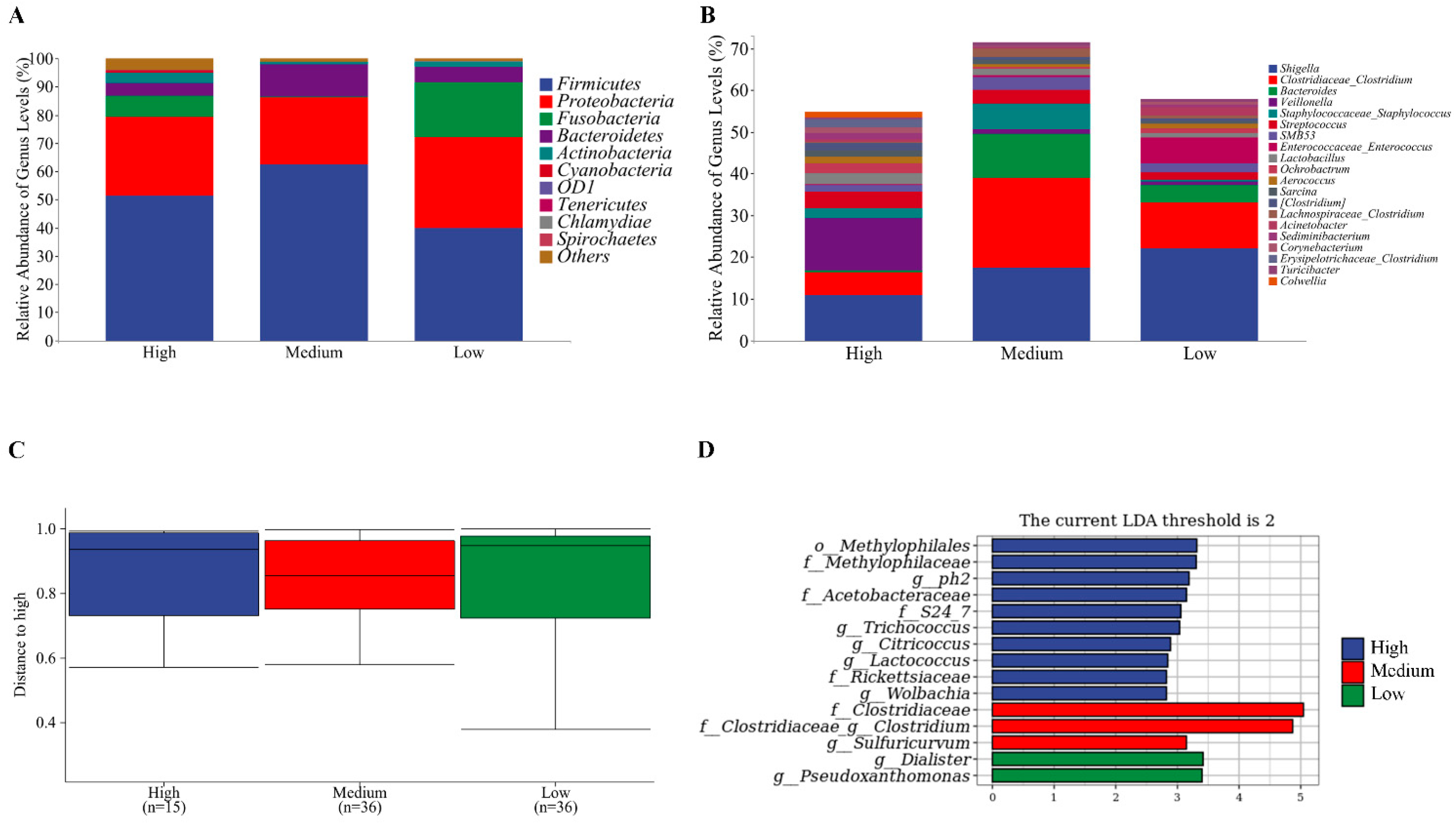
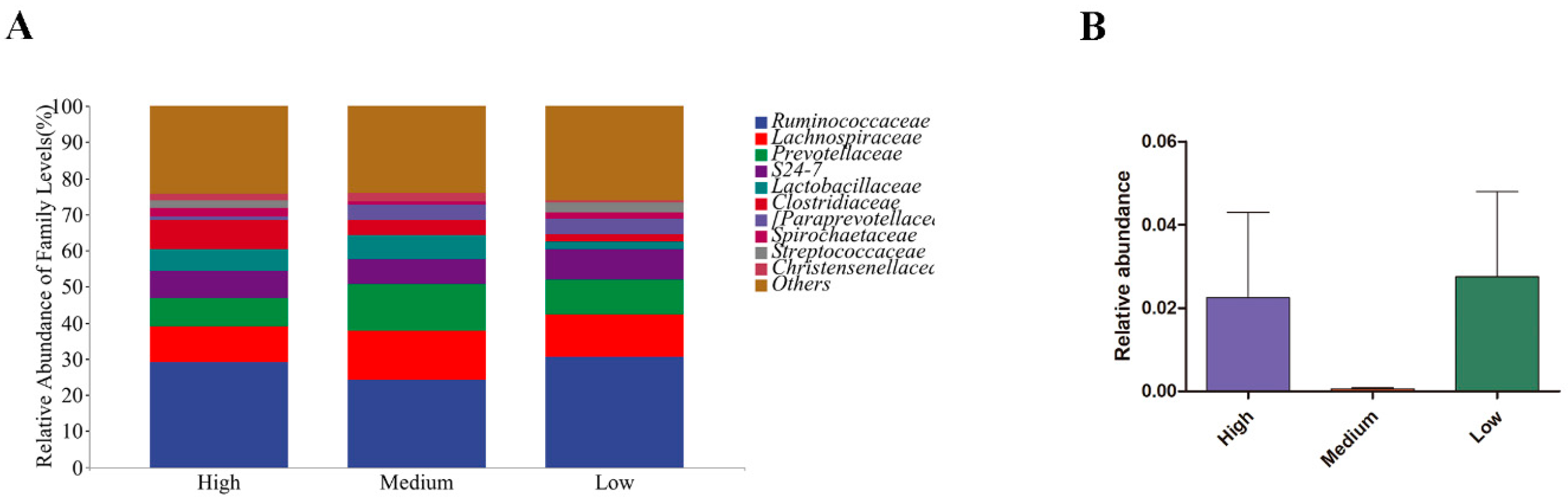
3.5. Microbial Flora of Sows and Newborn Piglets
4. Discussion
4.1. Sow Performance
4.2. Glucolipid Metabolism
4.3. Antioxidant Enzymes and Inflammatory Cytokines
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, D.J.P. The malnourished baby and infant. Br. Med. Bull. 2001, 60, 69–88. [Google Scholar] [CrossRef]
- Staessen, T.W.O.; Verdegem, M.C.J.; Nederlof, M.A.J.; Eding, E.H.; Schrama, J.W. Effect of type of dietary non-protein energy source (starch vs. fat) on the body bile acid pool size and composition, faecal bile acid loss and bile acid synthesis in rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2021, 27, 865–879. [Google Scholar] [CrossRef]
- Morris, D.L.; Brown-Brandl, T.M.; Hales, K.E.; Harvatine, K.J.; Kononoff, P.J. Effects of high-starch or high-fat diets formulated to be isoenergetic on energy and nitrogen partitioning and utilization in lactating Jersey cows. J. Dairy Sci. 2020, 103, 4378–4389. [Google Scholar] [CrossRef] [PubMed]
- Abbassi-Ghanavati, M.; Greer, L.G.; Cunningham, F.G. Pregnancy and Laboratory Studies A Reference Table for Clinicians. Obstet. Gynecol. 2009, 114, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Panarotto, D.; Remillard, P.; Bouffard, L.; Maheux, P. Insulin resistance affects the regulation of lipoprotein lipase in the postprandial period and in an adipose tissue-specific manner. Eur. J. Clin. Investig. 2002, 32, 84–92. [Google Scholar] [CrossRef]
- Gonzalez, C.G.; Alonso, A.; Balbin, M.; Diaz, F.; Fernandez, S.; Patterson, A.M. Effects of pregnancy on insulin receptor in liver, skeletal muscle and adipose tissue of rats. Gynecol. Endocrinol. 2002, 16, 193–205. [Google Scholar] [CrossRef]
- Ikeoka, D.; Krusinova, E. Insulin resistance and lipid metabolism. Rev. Assoc. Med. Bras. 2009, 55, 234. [Google Scholar] [CrossRef]
- Garduno-Alanis, A.; Vazquez-de Anda, G.; Valdes-Ramos, R.; Talavera, J.O.; Herrera-Villalobos, J.E.; Huitron-Bravo, G.G.; Hernandez-Garduno, E. Predictors of hyperlipidemia during the first half of pregnancy in Mexican women. Nutr. Hosp. 2015, 31, 508–513. [Google Scholar] [CrossRef]
- McMullin, T.S.; Lowe, E.R.; Bartels, M.J.; Marty, M.S. Dynamic changes in lipids and proteins of maternal, fetal, and pup blood and milk during perinatal development in CD and wistar rats. Toxicol. Sci. 2008, 105, 260–274. [Google Scholar] [CrossRef]
- Hu, X.-Q.; Song, R.; Zhang, L. Effect of Oxidative Stress on the Estrogen-NOS-NO-K-Ca Channel Pathway in Uteroplacental Dysfunction: Its Implication in Pregnancy Complications. Oxid. Med. Cell Longev. 2019, 2019, 9194269. [Google Scholar] [CrossRef]
- Spiteller, P.; Kern, W.; Reiner, J.; Spiteller, G. Aldehydic lipid peroxidation products derived from linoleic acid. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2001, 1531, 188–208. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.D.; De Long, N.E.; Wang, R.C.; Yazdi, F.T.; Holloway, A.C.; Raha, S. Angiogenesis in the Placenta: The Role of Reactive Oxygen Species Signaling. BioMed Res. Int. 2015, 2015, 814543. [Google Scholar] [CrossRef] [PubMed]
- Mor, G. Inflammation and pregnancy-The role of toll-like receptors in trophoblast-immune interaction. In Assessment of Human Reproductive Function; Bulletti, C., Guller, S., DeZiegler, D., Lockwood, C.J., Eds.; Annals of the New York Academy of Sciences: Cambridge, MA, USA, 2008; Volume 1127, pp. 121–128. [Google Scholar]
- Heerwagen, M.J.R.; Miller, M.R.; Barbour, L.A.; Friedman, J.E. Maternal obesity and fetal metabolic programming: A fertile epigenetic soil. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 299, R711–R722. [Google Scholar] [CrossRef]
- Challis, J.R.; Lockwood, C.J.; Myatt, L.; Norman, J.E.; Strauss, J.F., III; Petraglia, F. Inflammation and Pregnancy. Reprod. Sci 2009, 16, 206–215. [Google Scholar] [CrossRef]
- Jordan, S.; Tung, N.; Casanova-Acebes, M.; Chang, C.; Cantoni, C.; Zhang, D.; Wirtz, T.; Naik, S.; Rose, S.; Brocker, C.; et al. Dietary intake regulates the circulating inflammatory monocyte pool. Eur. J. Immunol. 2019, 49, 178. [Google Scholar] [CrossRef] [PubMed]
- Laws, J.; Amusquivar, E.; Laws, A.; Herrera, E.; Lean, I.J.; Dodds, P.F.; Clarke, L. Supplementation of sow diets with oil during gestation: Sow body condition, milk yield and milk composition. Livest. Sci. 2009, 123, 88–96. [Google Scholar] [CrossRef]
- Rosero, D.S.; Odle, J.; Mendoza, S.M.; Boyd, R.D.; Fellner, V.; van Heugten, E. Impact of dietary lipids on sow milk composition and balance of essential fatty acids during lactation in prolific sows. J. Anim. Sci. 2015, 93, 2935–2947. [Google Scholar] [CrossRef] [PubMed]
- White, C.L.; Pistell, P.J.; Purpera, M.N.; Gupta, S.; Fernandez-Kim, S.-O.; Hise, T.L.; Keller, J.N.; Ingram, D.K.; Morrison, C.D.; Bruce-Keller, A.J. Effects of high fat diet on Morris maze performance, oxidative stress, and inflammation in rats: Contributions of maternal diet. Neurobiol. Dis. 2009, 35, 3–13. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef] [PubMed]
- Edwards, S.M.; Cunningham, S.A.; Dunlop, A.L.; Corwin, E.J. The maternal gut microbiome during pregnancy. MCN Am. J. Matern. Child Nurs. 2017, 42, 310–316. [Google Scholar] [CrossRef]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Distinct composition of gut microbiota during pregnancy in overweight and normal-weight women. Am. J. Clin. Nutr. 2008, 88, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Hou, C.; Li, N.; Zhang, X.; Zhang, G.; Yang, F.; Zeng, X.; Liu, Z.; Qiao, S. Microbial and metabolic alterations in gut microbiota of sows during pregnancy and lactation. FASEB J. 2019, 33, 4490–4501. [Google Scholar] [CrossRef] [PubMed]
- Gohir, W.; Whelan, F.J.; Surette, M.G.; Moore, C.; Schertzer, J.D.; Sloboda, D.M. Pregnancy-related changes in the maternal gut microbiota are dependent upon the mother’s periconceptional diet. Gut. Microbes. 2015, 6, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.D.; Bemis, T.; Brychta, R.; Chen, K.Y.; Courville, A.; Crayner, E.J.; Goodwin, S.; Guo, J.; Howard, L.; Knuth, N.D.; et al. Calorie for Calorie, Dietary Fat Restriction Results in More Body Fat Loss than Carbohydrate Restriction in People with Obesity. Cell Metab. 2015, 22, 427–436. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Z.; Ming, D.; Huang, C.; Xu, S.; Li, Z.; Wang, Z.; Liu, H.; Zeng, X.; Wang, F. Effect of maternal dietary starch-to-fat ratio and daily energy intake during late pregnancy on the performance and lipid metabolism of primiparous sows and newborn piglets. J. Anim. Sci. 2022, 100, skac033. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 18th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2007. [Google Scholar]
- Adeola, O.; Lewis, A.; Southern, L. Digestion and Balance Techniques in Pigs. In Swine Nutrition, 2nd ed.; CRC Press LLC, Inc.: Boca Raton, FL, USA, 2001; Chapter 40; p. 899. [Google Scholar]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef]
- Jin, S.S.; Jin, Y.H.; Jang, J.C.; Hong, J.S.; Jung, S.W.; Kim, Y.Y. Effects of dietary energy levels on physiological parameters and reproductive performance of gestating sows over three consecutive parities. Asian-Australas. J. Anim. Sci. 2018, 31, 410–420. [Google Scholar] [CrossRef]
- Noblet, J.; Etienne, M. Effect of energy-level in lactating sows on yield and composition of milk and nutrient balance of piglets. J. Anim. Sci. 1986, 63, 1888–1896. [Google Scholar] [CrossRef]
- Westman, E.C.; Feinman, R.D.; Mavropoulos, J.C.; Vernon, M.C.; Volek, J.S.; Wortman, J.A.; Yancy, W.S.; Phinney, S.D. Low-carbohydrate nutrition and metabolism. Am. J. Clin. Nutr. 2007, 86, 276–284. [Google Scholar] [CrossRef]
- Kim, J.S.; Hosseindoust, A.; Ju, I.K.; Yang, X.; Lee, S.H.; Noh, H.S.; Lee, J.H.; Chae, B.J. Effects of dietary energy levels and β-mannanase supplementation in a high mannan-based diet during lactation on reproductive performance, apparent total tract digestibility and milk composition in multiparous sows. Ital. J. Anim. Sci. 2017, 17, 128–134. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, C.J.; Zhao, X.; Xiao, K.; Deng, M.; Zhang, L.; Qiu, X.; Deng, J.; Yin, Y.; Tan, C. Dietary energy sources during late gestation and lactation of sows: Effects on performance, glucolipid metabolism, oxidative status of sows, and their offspring1. J. Anim. Sci. 2019, 97, 4608–4618. [Google Scholar] [CrossRef]
- Ci, L.; Sun, H.; Huang, Y.; Guo, J.; Albrecht, E.; Zhao, R.; Yang, X. Maternal dietary fat affects the LT muscle fatty acid composition of progeny at weaning and finishing stages in pigs. Meat Sci. 2014, 96, 1141–1146. [Google Scholar] [CrossRef]
- Wang, Y.; Miura, Y.; Kaneko, T.; Li, J.; Qin, L.Q.; Wang, P.Y.; Matsui, H.; Sato, A. Glucose intolerance induced by a high-fat/low-carbohydrate diet in rats-Effects of nonesterified fatty acids. Endocrine 2002, 17, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Deng, M.; Chen, J.; Zhao, X.; Xiao, K.; He, W.; Qiu, X.; Xu, Y.; Yin, Y.; Tan, C. Starch supplementation improves the reproductive performance of sows in different glucose tolerance status. Anim. Nutr. 2021, 7, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- O’Malley, E.G.; Reynolds, C.M.E.; Killalea, A.; O’Kelly, R.; Sheehan, S.R.; Turner, M.J. Maternal obesity and dyslipidemia associated with gestational diabetes mellitus (GDM). Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 246, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Fang, Z.; Lin, Y.; Che, L.; Wu, C.; Xu, S.; Feng, B.; Li, J.; Wu, D. Influence of dietary fat source on sow and litter performance, colostrum and milk fatty acid profile in late gestation and lactation. Anim. Sci. J. 2017, 88, 1768–1778. [Google Scholar] [CrossRef] [PubMed]
- Saleri, R.; Sabbioni, A.; Cavalli, V.; Superchi, P. Monitoring blood plasma leptin and lactogenic hormones in pregnant sows. Animal 2015, 9, 629–634. [Google Scholar] [CrossRef]
- Forhead, A.J.; Fowden, A.L. The hungry fetus? Role of leptin as a nutritional signal before birth. J. Physiol. 2009, 587, 1145–1152. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Pérez, A.; Vilariño-García, T.; Guadix, P.; Dueñas, J.L.; Sánchez-Margalet, V. Leptin and Nutrition in Gestational Diabetes. Nutrients 2020, 12, 1970. [Google Scholar] [CrossRef]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef] [PubMed]
- Arita, Y.; Kihara, S.; Ouchi, N.; Takahashi, M.; Maeda, K.; Miyagawa, J.; Hotta, K.; Shimomura, I.; Nakamura, T.; Miyaoka, K.; et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem. Biophys. Res. Commun. 1999, 257, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, T.; Cai, A.; Wu, Y.; Wei, H.; Jiang, S.; Peng, J. Excessive backfat of sows at 109 d of gestation induces lipotoxic placental environment and is associated with declining reproductive performance. J. Anim. Sci. 2018, 96, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Feillet-Coudray, C.; Fouret, G.; Vigor, C.; Bonafos, B.; Jover, B.; Blachnio-Zabielska, A.; Rieusset, J.; Casas, F.; Gaillet, S.; Landrier, J.F.; et al. Long-Term Measures of Dyslipidemia, Inflammation, and Oxidative Stress in Rats Fed a High-Fat/High-Fructose Diet. Lipids 2019, 54, 81–97. [Google Scholar] [CrossRef]
- Yang, X.; Ma, X.; Wang, L.; Gao, K.; Jiang, Z. A high-fat diet expands body fat mass and up-regulates expression of genes involved in adipogenesis and inflammation in a genetically lean pig. J. Anim. Sci. 2017, 95, 223. [Google Scholar] [CrossRef]
- Chung, Y.L.; Lai, H.M. Molecular and granular characteristics of corn starch modified by HCl-methanol at different temperatures. Carbohydr. Polym. 2006, 63, 527–534. [Google Scholar] [CrossRef]
- Sandhu, K.S.; Singh, N. Some properties of corn starches II: Physicochemical, gelatinization, retrogradation, pasting and gel textural properties. Food Chem. 2007, 101, 1499–1507. [Google Scholar] [CrossRef]
- Yan, H.; Lu, H.; Almeida, V.V.; Ward, M.G.; Adeola, O.; Nakatsu, C.H.; Ajuwon, K.M. Effects of dietary resistant starch content on metabolic status, milk composition, and microbial profiling in lactating sows and on offspring performance. J. Anim. Physiol. Anim. Nutr. 2017, 101, 190–200. [Google Scholar] [CrossRef]
- Klingbeil, E.A.; Cawthon, C.; Kirkland, R.; de La Serre, C.B. Potato-Resistant Starch Supplementation Improves Microbiota Dysbiosis, Inflammation, and Gut-Brain Signaling in High Fat-Fed Rats. Nutrients 2019, 11, 2710. [Google Scholar] [CrossRef]
- Martínez, I.; Perdicaro, D.J.; Brown, A.W.; Hammons, S.; Carden, T.J.; Carr, T.P.; Eskridge, K.M.; Walter, J. Diet-induced alterations of host cholesterol metabolism are likely to affect the gut microbiota composition in hamsters. Appl. Environ. Microbiol. 2013, 79, 516–524. [Google Scholar] [CrossRef] [PubMed]
- Haenen, D.; Zhang, J.; Souza da Silva, C.; Bosch, G.; van der Meer, I.M.; van Arkel, J.; van den Borne, J.J.; Pérez Gutiérrez, O.; Smidt, H.; Kemp, B.; et al. A diet high in resistant starch modulates microbiota composition, SCFA concentrations, and gene expression in pig intestine. J. Nutr. 2013, 143, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Gohir, W.; Kennedy, K.M.; Wallace, J.G.; Saoi, M.; Bellissimo, C.J.; Britz-McKibbin, P.; Petrik, J.J.; Surette, M.G.; Sloboda, D.M. High-fat diet intake modulates maternal intestinal adaptations to pregnancy and results in placental hypoxia, as well as altered fetal gut barrier proteins and immune markers. J. Physiol. 2019, 597, 3029–3051. [Google Scholar] [CrossRef] [PubMed]
- Layden, B.T.; Angueira, A.R.; Brodsky, M.; Durai, V.; Lowe, W.L., Jr. Short chain fatty acids and their receptors: New metabolic targets. Transl. Res. 2013, 161, 131–140. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Starch-to-Fat Ratio | ||
|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |
| Ingredients | |||
| Corn | 43.20 | 53.00 | 53.20 |
| Soybean meal | 13.10 | 17.00 | 14.10 |
| Wheat bran | 10.00 | 13.00 | 21.30 |
| Fish meal | 4.60 | 0.00 | 0.00 |
| Soybean oil | 0.00 | 2.00 | 5.60 |
| Corn starch | 25.00 | 9.00 | 0.00 |
| Oil powder | 1.00 | 2.30 | 1.30 |
| Limestone | 1.00 | 1.30 | 1.50 |
| Dicalcium phosphate | 1.10 | 1.40 | 1.00 |
| Salt | 0.30 | 0.30 | 0.30 |
| Vitamin and mineral premix 1 | 0.50 | 0.50 | 0.50 |
| Choline chloride | 0.20 | 0.20 | 0.20 |
| L-Lysine HCL | 0.00 | 0.00 | 1.00 |
| Total | 100.00 | 100.00 | 100.00 |
| Analyzed levels | |||
| ME, MJ/kg2 | 13.90 | 13.92 | 13.91 |
| Gross energy, MJ/kg | 15.54 | 15.88 | 16.80 |
| Crude protein | 14.68 | 14.65 | 14.97 |
| Ether extract | 2.86 | 6.41 | 9.16 |
| Starch | 57.45 | 50.21 | 42.53 |
| Neutral detergent fiber | 16.14 | 16.61 | 16.98 |
| Acid detergent fiber | 3.72 | 5.42 | 5.37 |
| Calcium | 0.89 | 0.87 | 0.85 |
| Total phosphorus | 0.64 | 0.63 | 0.73 |
| Amino acids 2 | |||
| Lys | 0.79 | 0.73 | 0.71 |
| Met | 0.26 | 0.23 | 0.23 |
| Thr | 0.53 | 0.53 | 0.52 |
| Trp | 0.15 | 0.16 | 0.16 |
| Item | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| Number of sows | 25 | 25 | 25 | ||
| Average of parity | 4.5 | 4.3 | 4.6 | ||
| Body weight, kg | |||||
| D 85 of gestation | 249.1 | 247.2 | 251.4 | 2.68 | 0.819 |
| D 107 of gestation | 277.8 | 276.4 | 282.7 | 2.74 | 0.623 |
| Gain during gestation | 28.7 | 29.2 | 31.3 | 1.11 | 0.605 |
| Postpartum | 261.2 | 258.8 | 254.4 | 2.58 | 0.986 |
| Backfat thickness, mm | |||||
| D85 of gestation | 19.11 | 18.05 | 18.73 | 0.66 | 0.807 |
| D107 of gestation | 19.17 | 19.30 | 19.22 | 0.63 | 0.996 |
| Gain during gestation | 0.06 | 1.26 | 0.49 | 0.38 | 0.439 |
| Postpartum | 20.36 | 18.26 | 17.15 | 0.59 | 0.072 |
| No. of pigs per litter | |||||
| Total piglets born | 15.8 | 15.8 | 16.7 | 0.35 | 0.508 |
| Piglets born alive | 14.6 | 15.1 | 14.4 | 0.30 | 0.569 |
| Stillbirth | 0.9 | 0.6 | 1.1 | 0.11 | 0.221 |
| Mummy | 0.3 | 0.1 | 0.2 | 0.06 | 0.399 |
| Piglet birth weight | 1.45 a | 1.29 b | 1.35 ab | 0.02 | 0.010 |
| Litter birth weight, kg | 21.22 a | 19.11 b | 20.81 a | 0.38 | 0.045 |
| Item, % | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| Fat | 5.27 b | 5.74 ab | 6.46 a | 0.20 | 0.046 |
| Protein | 16.61 | 17.55 | 16.74 | 0.37 | 0.558 |
| Lactose | 2.28 a | 1.78 b | 1.99 ab | 0.08 | 0.027 |
| Item, % | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| Dry matter | 85.88 a | 82.37 b | 78.27 c | 0.008 | <0.001 |
| Crude protein | 85.51 | 83.87 | 83.98 | 0.003 | 0.093 |
| Ether extracts | 64.93 | 68.22 | 61.93 | 0.017 | 0.365 |
| Gross energy | 86.88 a | 83.61 b | 79.58 c | 0.008 | <0.001 |
| Neutral detergent fiber | 51.32 a | 38.59 b | 41.41 b | 0.016 | <0.001 |
| Acid detergent fiber | 44.62 a | 39.59 a | 20.39 b | 0.027 | <0.001 |
| Organic matter | 88.29 a | 85.64 b | 81.15 c | 0.008 | <0.001 |
| Item, mmol/L | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| D 107 of gestation | |||||
| Total cholesterol | 2.53 b | 2.57 b | 3.08 a | 0.09 | 0.019 |
| Triglyceride | 0.62 b | 0.87 ab | 1.36 a | 0.10 | 0.017 |
| HDL-C | 0.64 | 0.71 | 0.68 | 0.02 | 0.299 |
| LDL-C | 1.62 | 1.58 | 1.70 | 0.04 | 0.371 |
| Glucose | 4.05 b | 4.18 ab | 4.62 a | 0.09 | 0.014 |
| NEFA, μmol/L | 155.19 | 167.73 | 165.18 | 2.34 | 0.066 |
| Insulin, mIU/mL | 24.31 | 33.41 | 33.53 | 1.90 | 0.072 |
| HOMA-IR | 4.38 b | 6.26 ab | 7.02 a | 0.53 | 0.044 |
| Leptin | 1.88 | 2.04 | 2.04 | 0.03 | 0.101 |
| Adiponectin | 1.63 | 1.68 | 1.70 | 0.03 | 0.544 |
| Parturition | |||||
| Total cholesterol | 1.97 | 1.95 | 2.23 | 0.06 | 0.079 |
| Triglyceride | 0.32 | 0.36 | 0.37 | 0.02 | 0.708 |
| HDL-C | 0.49 | 0.57 | 0.6 | 0.02 | 0.077 |
| LDL-C | 1.17 b | 1.22 b | 1.41 a | 0.03 | 0.004 |
| Glucose | 5.11 | 5.37 | 5.35 | 0.16 | 0.782 |
| NEFA, μmol/L | 236.22 a | 232.72 a | 165.92 b | 6.48 | 0.001 |
| Insulin, mIU/mL | 25.09 | 26.83 | 28.37 | 2.13 | 0.828 |
| HOMA-IR | 5.63 | 6.06 | 6.71 | 0.46 | 0.638 |
| Leptin | 1.78 b | 1.84 b | 1.99 a | 0.03 | 0.008 |
| Adiponectin | 2.05 a | 2.08 a | 1.78 b | 0.04 | 0.012 |
| Item | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| lipoprotein lipase (U/g) | 42.70 | 48.80 | 47.03 | 2.49 | 0.619 |
| Leptin (ng/mg) | 0.86 | 1.03 | 0.95 | 0.04 | 0.175 |
| Adiponectin (μg/mg) | 0.77 | 0.94 | 0.88 | 0.04 | 0.133 |
| Item, mmol/L | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| Total cholesterol | 2.39 | 2.75 | 2.85 | 0.19 | 0.607 |
| Triglyceride | 1.21 | 1.58 | 1.61 | 0.12 | 0.350 |
| HDL-C | 0.44 | 0.54 | 0.62 | 0.05 | 0.349 |
| LDL-C | 1.46 | 1.65 | 1.73 | 0.08 | 0.281 |
| Glucose | 6.14 | 6.46 | 7.13 | 0.20 | 0.110 |
| NEFA, μmol/L | 180.28 | 184.23 | 190.00 | 2.74 | 0.361 |
| Insulin, mIU/mL | 47.91 ab | 44.55 b | 66.38 a | 4.13 | 0.039 |
| HOMA-IR | 13.14 b | 13.36 b | 21.23 a | 1.61 | 0.042 |
| Leptin | 1.43 b | 1.50 ab | 1.56 a | 0.02 | 0.033 |
| Adiponectin | 1.76 a | 1.67 ab | 1.61 b | 0.02 | 0.014 |
| Item | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| D 107 of gestation | |||||
| T-AOC, U/mL | 10.34 | 10.31 | 10.29 | 0.18 | 0.995 |
| SOD, U/mL | 244.70 | 232.25 | 249.84 | 5.67 | 0.444 |
| GSH-Px, umol/L | 21.51 b | 25.61 a | 27.40 a | 0.61 | 0.001 |
| MDA, nmol/mL | 1.49 | 1.53 | 1.65 | 0.05 | 0.392 |
| Parturition | |||||
| T-AOC, U/mL | 9.68 | 10.05 | 10.58 | 0.24 | 0.301 |
| SOD, U/mL | 243.20 | 269.97 | 245.50 | 5.45 | 0.080 |
| GSH-Px, umol/L | 26.94 | 25.06 | 26.81 | 0.53 | 0.282 |
| MDA, nmol/mL | 1.43 | 1.63 | 1.52 | 0.05 | 0.208 |
| Newborn piglets | |||||
| T-AOC, U/mL | 10.42 | 10.71 | 10.31 | 0.20 | 0.740 |
| SOD, U/mL | 258.34 | 250.69 | 264.59 | 4.01 | 0.424 |
| GSH-Px, umol/L | 13.02 | 13.48 | 14.86 | 0.68 | 0.441 |
| MDA, nmol/mL | 1.51 | 1.48 | 1.37 | 0.04 | 0.355 |
| Item, ng/L | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| D 107 of gestation | |||||
| Interleukin-1β | 87.12 | 93.00 | 88.65 | 1.64 | 0.326 |
| Interleukin-6 | 44.70 | 44.67 | 44.20 | 0.76 | 0.957 |
| Interleukin-10 | 21.38 | 21.32 | 21.94 | 0.33 | 0.718 |
| Tumor necrosis factor-α | 47.39 | 49.72 | 50.79 | 0.79 | 0.201 |
| Parturition | |||||
| Interleukin-1β | 85.21 b | 91.24 a | 90.94 a | 1.11 | 0.039 |
| Interleukin-6 | 41.41 b | 42.29 b | 45.10 a | 0.58 | 0.022 |
| Interleukin-10 | 22.43 | 21.61 | 21.71 | 0.36 | 0.608 |
| Tumor necrosis factor-α | 49.59 b | 54.83 a | 56.21 a | 0.83 | 0.001 |
| Newborn piglets | |||||
| Interleukin-1β | 71.22 b | 75.60 ab | 78.04 a | 1.05 | 0.022 |
| Interleukin-6 | 33.51 b | 35.22 ab | 36.18 a | 0.34 | 0.003 |
| Interleukin-10 | 16.36 | 17.26 | 17.86 | 0.30 | 0.264 |
| Tumor necrosis factor-α | 50.09 | 49.27 | 49.04 | 0.66 | 0.803 |
| Item, ng/L | Starch-to-Fat Ratio | SEM | p Value | ||
|---|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | |||
| D 107 of gestation | |||||
| Interleukin-1β | 35.86 b | 41.08 ab | 45.26 a | 1.62 | 0.049 |
| Interleukin-6 | 16.10 | 19.24 | 18.40 | 0.69 | 0.162 |
| Interleukin-10 | 8.88 | 10.85 | 9.09 | 0.42 | 0.108 |
| Tumor necrosis factor-α | 27.21 | 32.35 | 30.13 | 1.29 | 0.275 |
| Item | Starch-to-Fat Ratio | p Value | ||
|---|---|---|---|---|
| 20:1 | 10:1 | 5:1 | ||
| Sow | ||||
| Chao1 | 4261.78 | 5347.26 | 4597.68 | 0.281 |
| Faith_pd | 142.44 | 161.13 | 158.73 | 0.368 |
| Goods_coverage | 0.96 | 0.95 | 0.96 | 0.343 |
| Observed_species | 3733.48 | 4459.72 | 3913.87 | 0.182 |
| Pielou_e | 0.83 | 0.84 | 0.83 | 0.423 |
| Shannon | 9.81 | 10.20 | 9.86 | 0.082 |
| Simpson | 0.99 | 0.99 | 0.99 | 0.653 |
| Newborn piglets | ||||
| Chao1 | 1231.97 | 1513.94 | 1042.04 | 0.402 |
| Faith_pd | 682.00 | 194.83 | 162.29 | 0.291 |
| Goods_coverage | 0.99 | 0.99 | 0.99 | 0.135 |
| Observed_species | 1114.27 | 1293.00 | 893.60 | 0.423 |
| Pielou_e | 0.63 | 0.63 | 0.48 | 0.476 |
| Shannon | 6.38 | 6.50 | 4.78 | 0.532 |
| Simpson | 0.87 | 0.93 | 0.73 | 0.470 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Yu, Z.; Yin, X.; Wang, Z.; Xu, S.; Shi, C.; Zang, J.; Liu, H.; Wang, F. Effect of Dietary Starch-to-Fat Ratio on Lipid Metabolism, Inflammation, and Microbiota of Multiparous Sow and Newborn Piglets. Agriculture 2023, 13, 1069. https://doi.org/10.3390/agriculture13051069
Wang W, Yu Z, Yin X, Wang Z, Xu S, Shi C, Zang J, Liu H, Wang F. Effect of Dietary Starch-to-Fat Ratio on Lipid Metabolism, Inflammation, and Microbiota of Multiparous Sow and Newborn Piglets. Agriculture. 2023; 13(5):1069. https://doi.org/10.3390/agriculture13051069
Chicago/Turabian StyleWang, Wenhui, Zirou Yu, Xindi Yin, Zijie Wang, Song Xu, Chenyu Shi, Jianjun Zang, Hu Liu, and Fenglai Wang. 2023. "Effect of Dietary Starch-to-Fat Ratio on Lipid Metabolism, Inflammation, and Microbiota of Multiparous Sow and Newborn Piglets" Agriculture 13, no. 5: 1069. https://doi.org/10.3390/agriculture13051069