
The Influence of Mixed Green Covers, a New Trend in Organic Olive Growing, on the Efficiency of Predatory Insects
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
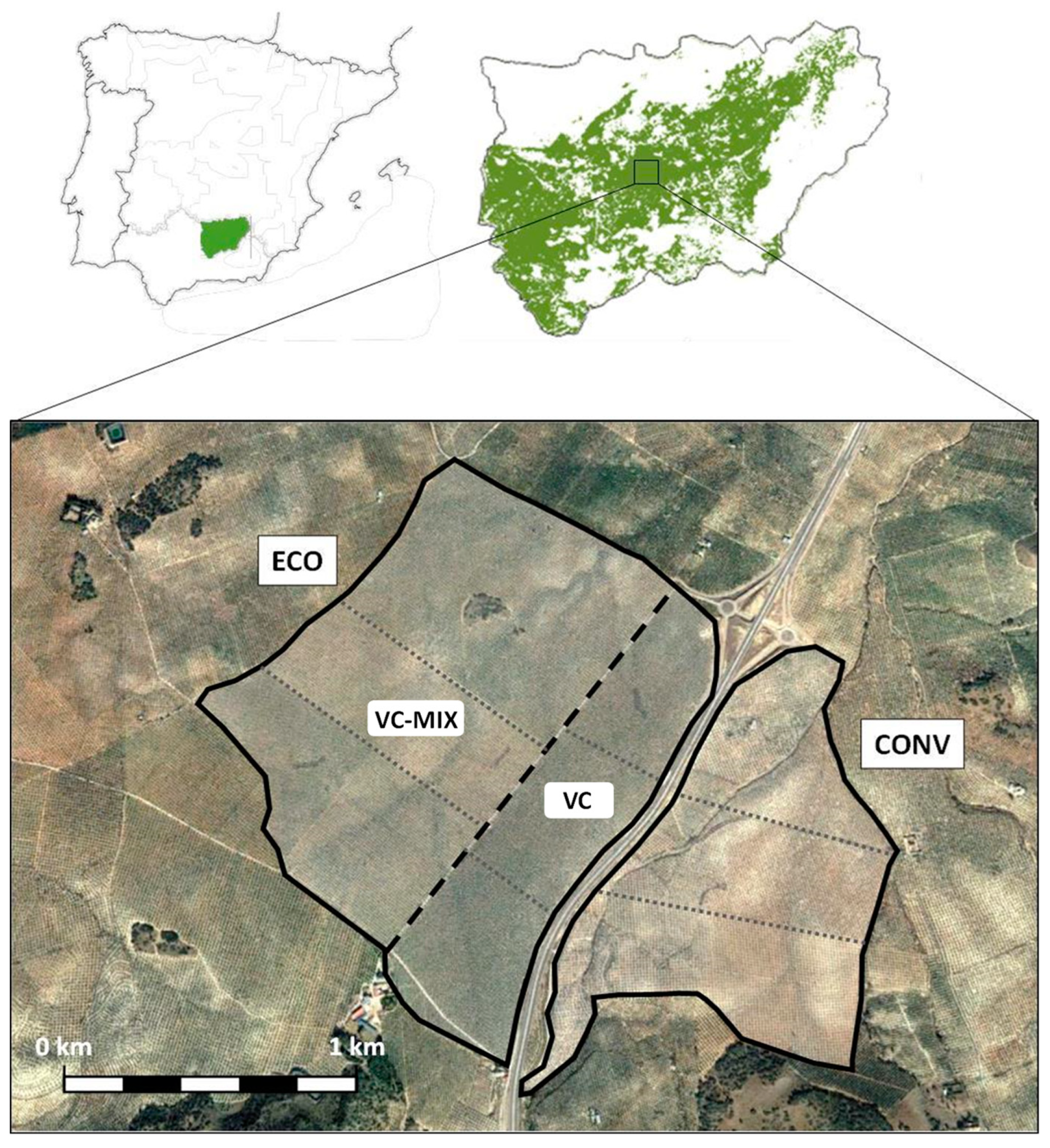
2.1. Description of the Study Area
2.1.1. Olive Groves with Ecological Farming (VC-MIX and VC)
2.1.2. Olive Grove with Conventional Farming
2.2. Experimental Design
2.2.1. Entomological Sampling
2.2.2. Sampling of the Vegetation (VC; VC-MIX Olive Groves)
2.2.3. Estimation of Oophagous Predation (VC, VC-MIX, and CONV Olive Groves)
- Population index: The total number of eggs/100 fruits. This density value reflects the relative density of the ovideponent females in the cultivated area.
- Potential attack: The number of fruits with any type of eggs (live, predated, or hatched) × 100/total number of fruits observed. This represents a value equivalent to the fruit drop that P. oleae would cause in the case of the total absence of oophagous predation activity.
- Hatching rate: The number of hatched eggs × 100/(sum of live and hatched eggs).
- Predation rate: The number of predated eggs/(sum of live eggs + predated + hatched). This allows for assessment of the predatory activity using an index of the activity of predatory eggs.
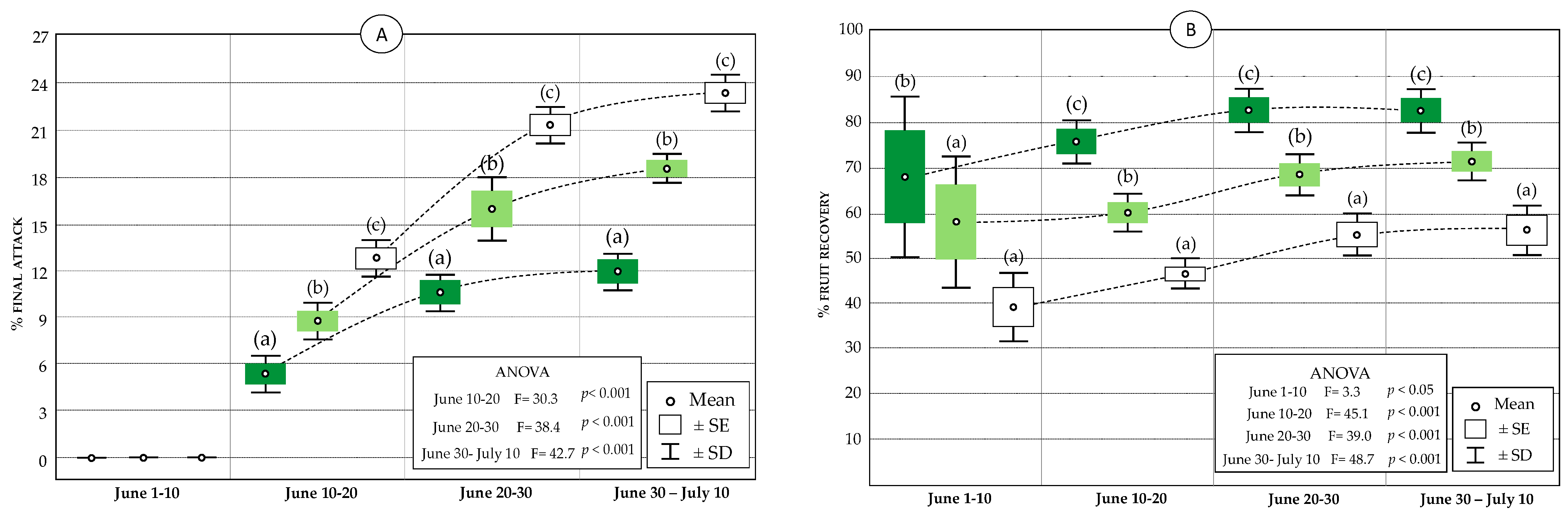
- Final attack: The number of fruits that contain at least one hatched egg × 100/(number of fruits observed). Given that the loss of fruit caused by P. oleae is exclusively due to the larvae emerging inside the attacked fruits once their larval development is complete, only hatched eggs were considered for its calculation. Therefore, the latter may have survived the predatory action of the natural enemies. For calculating this value, therefore, the live eggs were not accounted for, since these are likely to hatch or be predated. The entomophagous action allows for discarding a proportion of the population of P. oleae eggs; however, in the opposite case, their hatching results in establishing the larva in the endocarp of the fruit and its subsequent fall. The final attack describes the magnitude of this drop, providing a realistic estimation of the impact of P. oleae on the harvest.
- Fruit recovery: The number of fruits in which all the eggs have been predated × 100/(number of fruits that have contained eggs). This parameter indicates predation’s real effectiveness, since it corresponds to the percentage of fruits in which all the eggs have been predated. Once all the P. oleae eggs were eliminated, fruits were considered recovered, so, for practical purposes, the recovery percentage is a parameter that indicates the oophagous predation’s real effectiveness through the lacewings. Importantly, this value does not always correspond to the predation value, except in the cases in which the number of fruits attacked present infestation densities equal to 1 egg/fruit.
2.2.4. Statistical Analysis
3. Results
3.1. Diversity and Relative Abundance of Plant Species in Vegetation Cover (VC and VC-MIX)
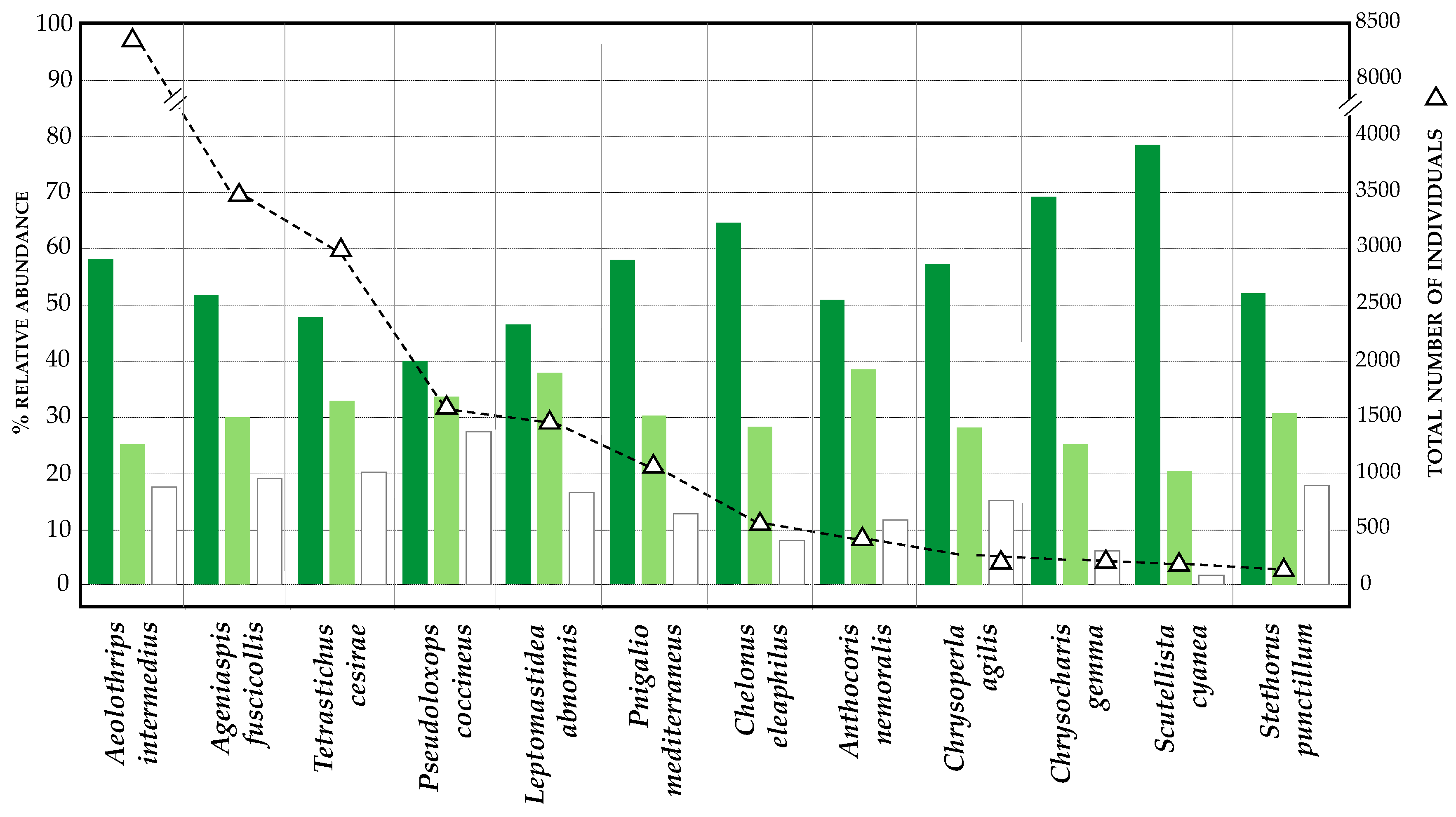
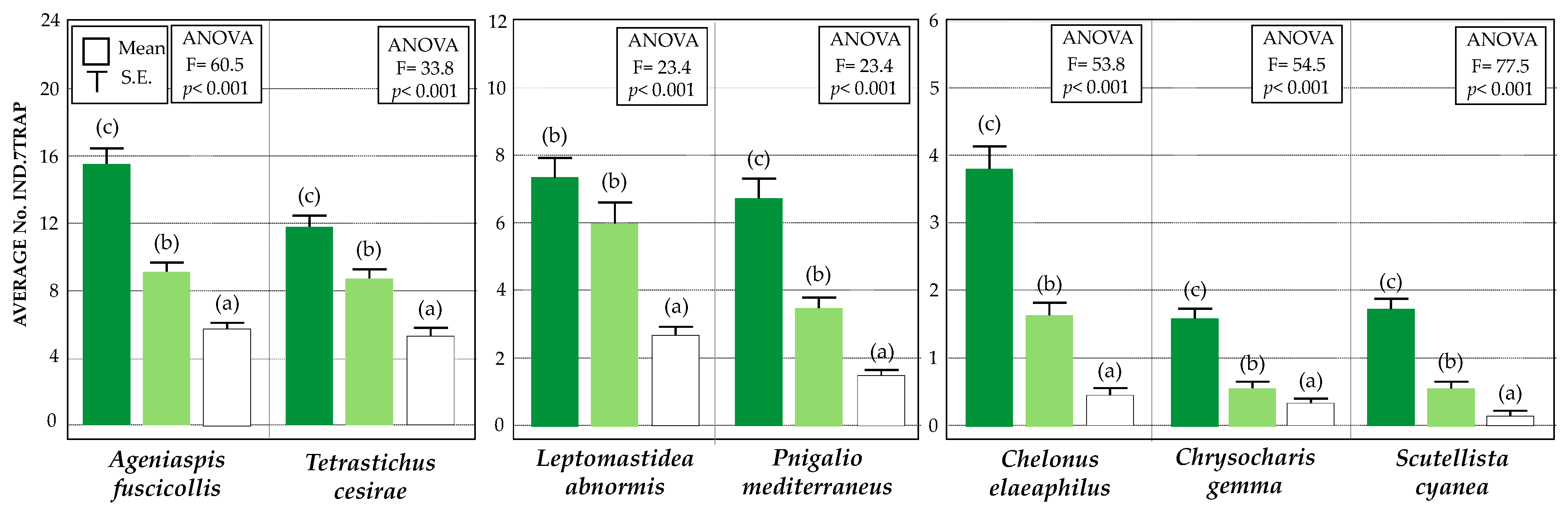
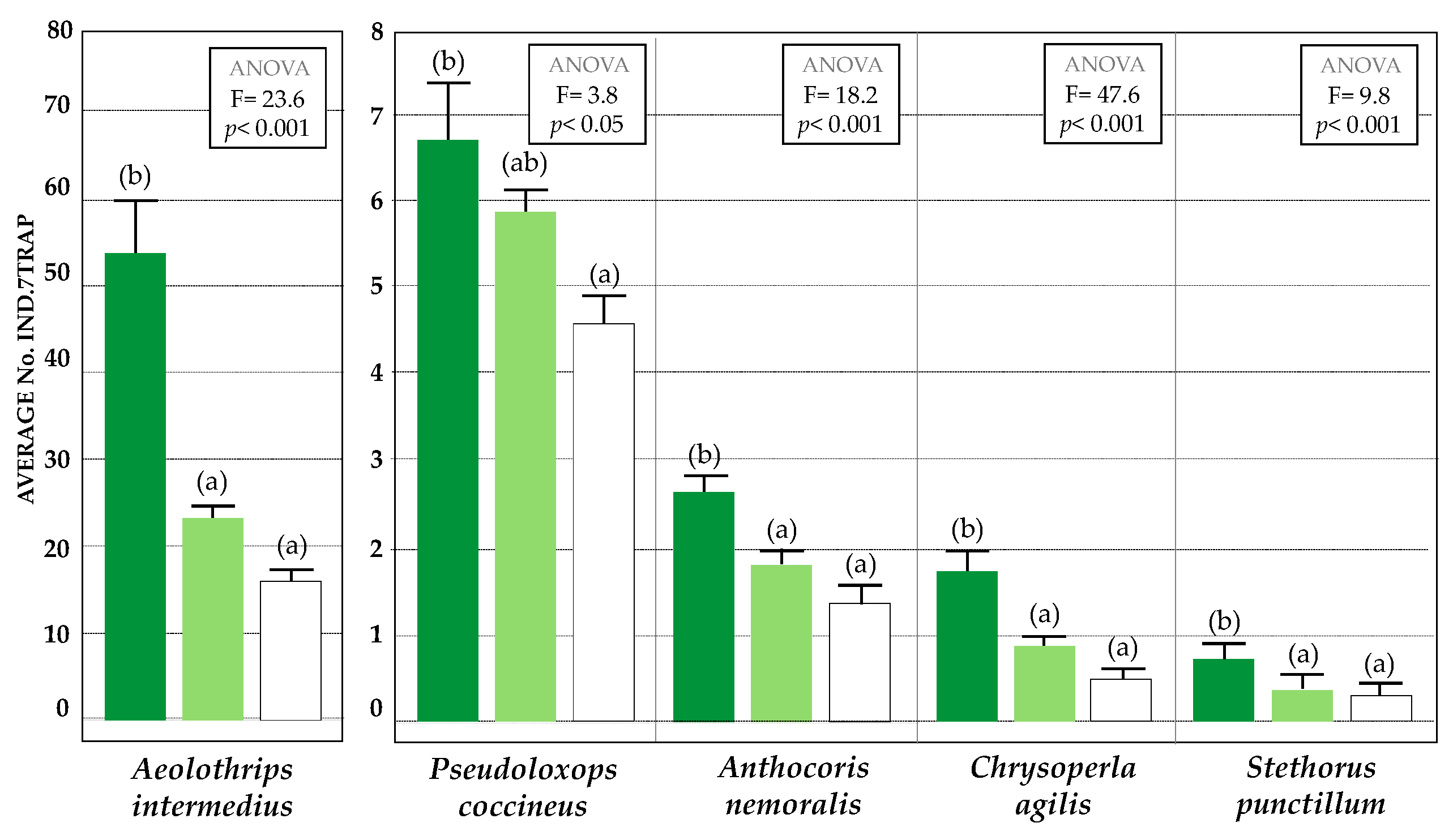
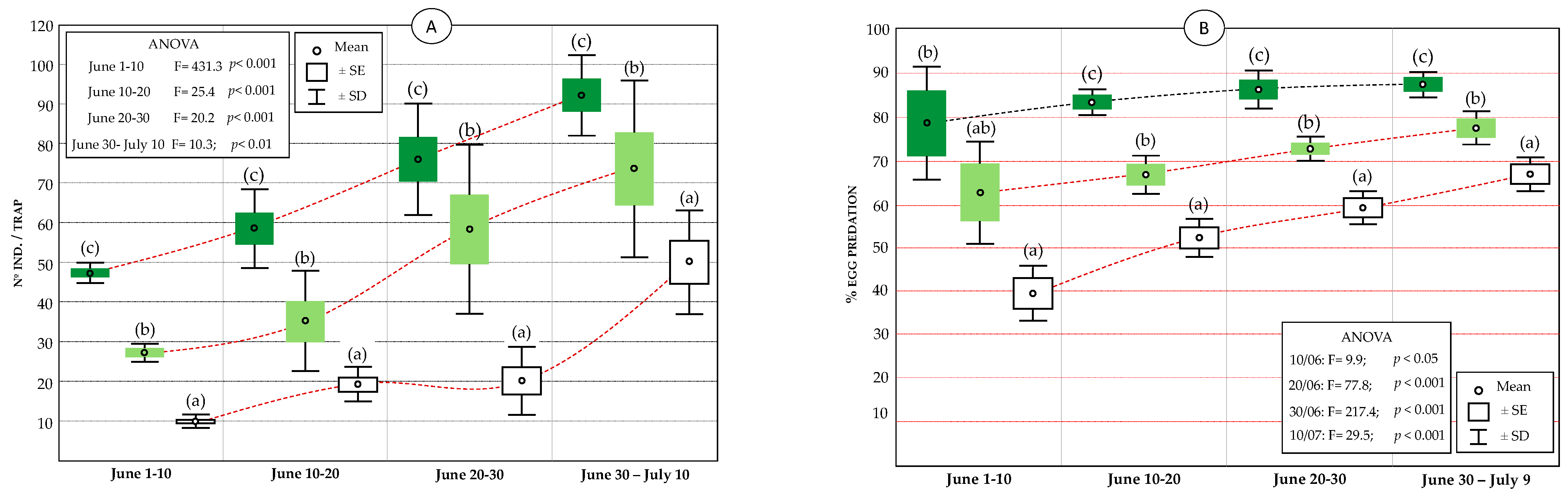
3.2. Diversity and Relative Abundance of Beneficial Insects (Chromatic Sticky Traps)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parasitoids spp. | Olive Pest Host | Host Association | |
|---|---|---|---|
| Cephalonomia cursor Westwood, 1881 (Hym.: Bethylidae) |  | Phloeotribus scarabaeoides | [36] Russo, 1938 |
| Apanteles xanthostigma (Haliday, 1834) (Hym.: Braconidae) |  | Prays oleae | [33] Campos, 1981 |
| Psyttalia concolor (Szepligeti, 1910) (Hym.: Braconidae) |  | Bactrocera oleae | [35] Kapatos, 1977 |
| Chelonus elaeaphilus Silvestri, 1908 (Hym.: Braconidae) |  | Prays oleae | [33] Campos, 1981 |
| Ageniaspis fuscicollis (Dalman, 1820) (Hym.: Encyrtidae) |  | Prays oleae | [33] Campos, 1981 |
| Diversinervus elegans Silvestri, 1915 (Hym.: Encyrtidae) |  | Saissetia oleae | [37] Panis, 1977 |
| Leptomastidea abnormis (Girault, 1915) (Hym.: Encyrtidae) |  | Saissetia oleae | [38] Copland, 1983 |
| Scutellista cyanea Motschulsky, 1859 (Hym.: Encyrtidae) |  | Saissetia oleae | [37] Panis, 1977 |
| Chrysocharis gemma (Walker, 1839) (Hym.: Eulophidae) |  | Prays oleae | [39] Noyes, 2019 |
| Pnigalio mediterraneus Ferrierre & Delucchi, 1957 (Hym.: Eulophidae) |  | Bactrocera oleae; Prays oleae | [40] Stavraki, 1970 |
| Tetrastichus cesirae Russo, 1938 (Hym.: Eulophidae) |  | Saissetia oleae; Bactrocera oleae; Liothrips oleae | [3] Arambourg, 1986 |
| Elasmus flabellatus (Fonscolombe, 1832) (Hym.: Eulophidae) |  | Prays oleae | [33] Campos, 1981 |
| Eupelmus urozonus (Dalman, 1820) (Hym.: Eupelmidae) |  | Bactrocera oleae | [33] Campos, 1981 |
| Diadegma semiclausum (Hellén, 1949) (Hym.: Ichneumonidae) |  | Prays oleae | [41] Torres, 2010 |
| Cheiropachus quadrum (Fabricius, 1787) (Hym.: Pteromalidae) |  | Phloeotribus scarabaeoides | [36] Russo, 1938 |
| Cyrtoptyx latipes (Rondani, 1874) (Hym.: Pteromalidae) |  | Bactrocera oleae | [42] Ranaldi and Santoni, 1987 |
| Predators spp. | Olive Pest Prey | Prey Association | |
|---|---|---|---|
| Coccinella septempunctata (L., 1758) (Col: Coccinellidae) |  | Saissetia oleae; Lepidosaphes ulmi; Parlatoria oleae; Aspidiotus nerii | [3] Arambourg, 1986 |
| Stethorus punctillum (Weise, 1891) (Col.: Coccinellidae) |  | Saissetia oleae; Lepidosaphes ulmi; Parlatoria oleae; Aspidiotus nerii | [3] Arambourg, 1986 |
| Prolasioptera berlesiana Paoli, 1907 (Dip.: Cecidomyiidae) |  | Bactrocera oleae | [43] Silvestri, 1945 |
| Anthocoris nemoralis (F., 1794) (Hem.: Anthocoridae) |  | Euphyllura olivina; Liothrips oleae; Prays oleae | [3] Arambourg, 1986 |
| Pseudoloxops coccineus (Meyer-Dür, 1843) (Hem.: Miridae) |  | Prays oleae | [44] Paredes et al., 2013 |
| Brachynotocoris ferreri Baena (Hem.: Miridae) |  | Prays oleae | [44] Paredes et al., 2013 |
| Chrysoperla agilis Henry et al., 2003 (Neu.: Chrysopidae) |  | Euphyllura olivina; Prays oleae | [30] Bozsik and González-Ruiz, 2006 |
| Harraphidia laufferi (Navás, 1915) (Rap.: Raphidiidae) |  | Phloeotribus scarabaeoides | [45] Rozas and González-Ruiz, 2017 |
| Aeolothrips intermedius Bagnall, 1934 (Thy.: Aeolothripidae) |  | Liothrips oleae; Aceria oleae; Oxycenus maxwelli | [46] De Liñán, 1998 |
| Beneficial Species | VC-MIX | VC | Conventional | |
|---|---|---|---|---|
| Predators | Coccinella septempunctata | X | X | - |
| Stethorus punctillum | X | X | X | |
| Prolasioptera berlesiana | X | - | - | |
| Anthocoris nemoralis | X | X | X | |
| Pseudoloxops coccineus | X | X | X | |
| Brachynotocoris ferreri | X | - | - | |
| Chrysoperla agilis | X | X | X | |
| Harraphidia laufferi | X | - | - | |
| Aeolothrips intermedius | X | X | X | |
| Parasitoids | Cephalonomia cursor | X | X | X |
| Apanteles xanthostigma | X | X | - | |
| Psyttalia concolor | X | X | - | |
| Chelonus elaeaphilus | X | X | X | |
| Ageniaspis fuscicollis | X | X | X | |
| Diversinervus elegans | X | X | - | |
| Leptomastidea abnormis | X | X | X | |
| Scutellista cyanea | X | X | X | |
| Chrysocharis gemma | X | X | X | |
| Pnigalio mediterraneus | X | X | X | |
| Tetrastichus cesirae | X | X | X | |
| Elasmus flabellatus | X | - | - | |
| Eupelmus urozonus | X | - | - | |
| Diadegma semiclausum | X | X | X | |
| Cheiropachus quadrum | X | X | X | |
| Cyrtoptyx latipes | X | - | - |
3.3. Attack Parameters of Olive Moth, P. oleae
3.3.1. Egg Population and Potential Attack
3.3.2. Evolution of Lacewing Catches (McPhail Traps) and Oophagous Larval Predation
4. Discussion
- -
- Plants provide a settlement for numerous phytophagous species which are potential prey/alternative hosts for numerous entomophagous species, which due to their relatively polyphagous nature, interact with the phytophagous species that are specifically associated with olive trees. Regarding conventional olive growing, in which there is no plant cover, implementing living plant covers creates a settlement for a wide community of species in the cultivation area, which causes the herbaceous plants of the herbaceous cover to be authentic insect reservoirs [12].
- -
- Secondly, herbaceous vegetation provides a food source for the adults of many predatory and parasitoid species [58]. These arthropods require an adequate intake to develop their fecundity, reach their potential longevity [59], and ensure post-diapause survival. The nectar of flowers is especially important for hymenopterous parasitoids [60,61] and predators [62] that are attracted by the release of semiochemical volatiles [63,64]. Several authors [65,66] have reported that Ch. carnea can be caught in traps baited with phenylacetaldehyde, which is a common constituent of a flower scent and is of great practical interest [64,65]. Thus, maintaining biodiversity is essential for achieving greater stability and can be used as an indicator of the system’s resistance to change [67,68].
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MAGRAMA. Ministerio de Agricultura, Alimentación y Medio Ambiente. Available online: https://www.mapa.gob.es/estadistica/pags/anuario/2012/AE_2012_13_12_01_01.pdf (accessed on 15 February 2023).
- García-Fuentes, A.; Lendínez, M.L.; Salazar, C. Pérdida de diversidad vegetal en los olivares del Alto Valle del Guadalquivir: Alternativas agroecológicas. In La Cultura del Olivo, Ecología, Economía, Sociedad; Anta, J.L., Palacios, J., Guerrero, F., Eds.; Universidad de Jaén: Jaén, Spain, 2005; pp. 300–430. ISBN 84-8439-250-3. [Google Scholar]
- Arambourg, Y. Traité D’entomologie Oléicole; Conseil oléicole International Publishing: Madrid, Spain, 1986; p. 360. ISBN 8439875835/9788439875833. [Google Scholar]
- Andrés-Cantero, F. Enfermedades y Plagas del Olivo, 1st ed.; Riquelme y Vargas Ediciones, SL: Jaén, Spain, 1997; p. 646. ISBN 8486216311/9788486216313. [Google Scholar]
- Gómez-Guzmán, J.A.; Sainz-Pérez, M.; González-Ruiz, R. Monitoring and inference of behavioral resistance in beneficial insects to insecticides in two pest control systems: IPM and organic. Agronomy 2022, 12, 538. [Google Scholar] [CrossRef]
- Vives, J.M. Control de plagas de insectos. Problemas y alternativas. In Insecticidas Biorracionales; Bellés, J., Ed.; Consejo Superior de Investigaciones Científicas; Colección Nuevas Tendencias: Madrid, Spain, 1988; pp. 3–13. [Google Scholar]
- Carrero, J.M. Lucha Integrada Contra Las Plagas Agrícolas y Forestales; Mundi-Prensa Ediciones: Madrid, Spain, 1996; p. 272. ISBN 8471146398/9788471146397. [Google Scholar]
- Gómez-Guzmán, J.A.; González-Ruiz, R. Side effects of insecticides on beneficial insects: A practical tool to identify organic agroecosystems. World J. Agric. Sci. 2019, 4, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Guzmán, J.A.; González-Ruiz, R. Determination of the sampling size for the reliable identification of organic crops by inducing sublethal effects in beneficial insects. In Proceedings of the IOP Conference Series: Earth and Enviromental Science, Prague, Czech Republic, 7–11 September 2020. [Google Scholar] [CrossRef]
- Gómez-Guzmán, J.A.; Sainz-Pérez, M.; González-Ruiz, R. Induction of sublethal effects for the characterization of olive groves under different pest management systems. Rev. Bras. Frutic. 2021, 43, 1–15. [Google Scholar] [CrossRef]
- Civantos-Ruiz, M.; Gómez-Guzmán, J.A.; Sainz-Pérez, M.; González-Ruiz, R. Application of accumulated heat units in the control of Prays oleae (Bern.) (Lep., Praydidae). Rev. Bras. Frutic. 2022, 44, e-804. [Google Scholar] [CrossRef]
- Hodkinson, I.D.; Hughes, M.K. Fitofagia en los Insectos, 2nd ed.; Oikos-Tau, S.A., Ed.; Ediciones: Barcelona, Spain, 1993; p. 104. ISBN 978-84-281-0803-4. [Google Scholar]
- Chauvin, R. The World of an Insect; Translated from the French by Harold Oldroyd; Weidenfeld and Nicolson: London, UK, 1967; p. 256. [Google Scholar]
- Cerdà, A.; Giménez-Morera, A.; Bodí, M.B.; Burguet, M.; García-López, J.; Jovani, C.; Segura, M. Pérdida de suelo y agua bajo cubierta de Quercus coccifera en la sierra de enguera. Valencia. C & G 2010, 24, 13–23. [Google Scholar]
- Rodríguez-Lizana, A.; Pereira, M.J.; Ribeiro, M.C.; Soares, A.; Márquez-García, F.; Ramos, A.; Gil-Ribes, J. Assessing Local Uncertainty of Soil Protection in an Olive Grove Area with Pruning Residues Cover: A Geostatistical Cosimulation Approach. Land Degrad. Dev. 2017, 28, 2086–2097. [Google Scholar] [CrossRef]
- Vanwalleghem, T.; Gómez, J.A.; Amate, J.I.; De Molina, M.G.; Vanderlinden, K.; Guzmán, G.; Laguna, A.; Giráldez, J.V. Impact of historical land use and soil management change on soil erosion and agricultural sustainability during the anthropocene. Anthropocene 2017, 17, 13–29. [Google Scholar] [CrossRef]
- Zema, D.A.; Labate, A.; Martino, D.; Zimbone, S.M. Comparing different infiltration methods of the HEC-HMS model: The case study of the mésima torrent (southern italy). Land Degrad. Dev. 2017, 28, 294–308. [Google Scholar] [CrossRef]
- Casado, G.G.; Pulido, L.F. Manejo de la Cubierta Vegetal en el Olivar Ecológico en Andalucía: Siembra de Leguminosas Entre Calles; Dirección General de Agricultura Ecológica; Consejería de Agricultura y Pesca: Junta de Andalucía, Spain, 2007; p. 78. [Google Scholar]
- Alcántara, C.; Pujadas, A.; Saavedra, M. Management of cruciferous cover crops by mowing for soil and water conservation in southern Spain. Agric. Water Manag. 2011, 98, 1071–1080. [Google Scholar] [CrossRef]
- García-Martín, J.F.; Cuevas, M.; Feng, C.H.; Álvarez-Mateos, P.; Torres, M.; Sánchez, S. Energetic valorisation of olive biomass: Olive-tree pruning, olive stones and pomaces. Processes 2020, 8, 511. [Google Scholar] [CrossRef]
- Iranzo, M.; Canizares, J.V.; Roca-Perez, L.; Sainz-Pardo, I.; Mormeneo, S.; Boluda, R. Characteristics of rice straw and sewage sludge as composting materials in Valencia (Spain). Bioresour. Technol. 2004, 95, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Ribo, M.; Albiach, R.; Pomares, F.; Canet, R. Alternativas de gestión de la paja de arroz en la Albufera de Valencia. Vida Rural 2017, 430, 56–60. [Google Scholar]
- Lull, C.; Bautista, I.; Segarra, R.; Monzó, J.M.; Seguí, J.V.; Lidón, A. Efecto de una cubierta de paja de arroz sobre la respiración del suelo. In Proceedings of the IX Simposio Nacional Sobre Control de la Degradación y Recuperación de Suelos, Elche, Spain, 27–29 May 2021. [Google Scholar]
- Campos, M.; Ramos, P. Some relationships between the number of Prays oleae eggs laid on olive fruits and their predation by Chrysoperla carnea. In Integrated Pesticide Control Olive-groves. In Proceedings of the CEC/FAO/IOBC International Joint Meeting, Pisa, Italy, 3–6 April 1985. [Google Scholar]
- Bozsik, A.; González-Ruiz, R.; Lara, B.H. Distribution of the Chysoperla carnea complex in southern Spain (Neuroptera: Chrysopidae). An. Univ. Oradea Fasc. Protectia Mediu. 2009, 14, 60–65. [Google Scholar]
- Trdan, S.; Andjus, L.; Raspudić, E.; Kač, M. Distribution of Aeolothrips intermedius Bagnall (Thysanoptera: Aeolothripidae) and its potential prey Thysanoptera species on different cultivated host plants. J. Pest. Sci. 2005, 78, 217–226. [Google Scholar] [CrossRef]
- González-Ruiz, R.; Gómez-Guzmán, J.A. Agricultural Management Greatly Affects the Beneficial Entomofauna of the Olive Groves. Am. J. Biomed. Sci. Res. 2019, 1, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Ebeling, W.; Wagner, R.; Reierson, D.A. Influence of repellency on the efficacy of blatticides. I. Learned modification of behavior of the German cockroach. J. Econ. Entomol. 1966, 59, 1374–1388. [Google Scholar] [CrossRef]
- Ail-Catzim, C.E.; Cerna-Chávez, E.; Landeros-Flores, J.; Ochoa-Fuentes, Y.; García-López, A.M.; González, R.E. Efecto de Insecticidas Sobre la Mortalidad y Depredación de Chrysoperla carnea (Neuroptera: Chrysopidae). Southwest. Entomol. 2015, 40, 565–574. [Google Scholar] [CrossRef]
- Bozsik, A.; González-Ruiz, R. First data on the sibling species of the common green lacewings in Spain (Neuroptera: Chrysopidae): (The taxonomic status of the most important cryptic species of Chrysoperla carnea complex in Spain). In Proceedings of the 4th International Plant Protection Symposium at Debrecen University and 11th Trans-Tisza Plant Protection Forum, Debrecen, Hungary, 18–19 October 2006. [Google Scholar]
- González Ruiz, R.; Al-Asaad, S.; Bozsik, A. Influencia de las masas forestales en la diversidad y abundancia de los crisópidos (Neur. Chrysopidae) del olivar. Cuad. Soc. Esp. Cienc. For. 2008, 26, 33–38. [Google Scholar]
- Ramos, P.; Ramos, J.M. Veinte años de observaciones sobre la depredación oófaga en Prays oleae Bern. Granada (España), 1970–1989. Boletín Sanid. Veg. Plagas 1990, 16, 119–127. [Google Scholar]
- Campos, M. Contribución al estudio de la entomocenosis de Prays oleae Bern. (Lep. Hyponomeutidae) en Granada (España). Acta Oecol. 1981, 2, 27–35. [Google Scholar]
- Varela, J.L.; González, R. Estudio sobre la entomofauna de un olivar en la provincia de Granada, durante el periodo de vuelo de la generación antófaga de Prays oleae (Lep. Yponomeutidae). Phytoma 1999, 111, 42–55. [Google Scholar]
- Kapatos, E.; Fletcher, B.S.; Pappas, S.; Laudeho, Y. Release of Opius concolor and O. concolor var. siculus [Hym.: Braconidae] against the spring generation of Dacus oleae [Dipt.: Trypetidae] on Corfu. Entomophaga 1977, 22, 265–270. [Google Scholar] [CrossRef]
- Russo, G. VI Contributo alla conoscenza dei coleotteri Scolitidi: Phloeotribus scarabaeoides (Bern.) Fauv. II. Biografia, simbionti, danni e lotta. Boll. R. Lab. Entomol. Agric. Portici 1938, 2, 3–420. [Google Scholar]
- Panis, A. Contribución al conocimiento de la biología de la «cochinilla negra de los agrios» (Saissetia oleae Olivier). Bol. Serv. Plagas 1977, 3, 199–205. [Google Scholar]
- Copland, J.S. Restricciones de temperatura en el control de cochinillas y cochinillas. Bol. SROP 1983, 6, 142–145. [Google Scholar]
- Noyes, J.S. Universal Chalcidoidea Database; World Wide Web Electronic Publication, 2019. Available online: https://www.nhm.ac.uk/our-science/data/chalcidoids/ (accessed on 15 February 2023).
- Stavraki, H. Contribution a l’inventaire du complexe parasitaire de quelques insectes noisibles a l’olivier en Grèce. Entomophaga 1970, 15, 225–231. [Google Scholar] [CrossRef]
- Torres, M.R. Parasitoides de plagas identificados en la provincia de Jaén (España). Bol. Soc. Entomol. Aragonesa 2010, 46, 597–601. [Google Scholar]
- Ranaldi, F.Y.; Santoni, M. Los parasitoides de la mosca del olivo Dacus oleae (Gmel). Inf. Fitopatol. 1987, 37, 15–18. [Google Scholar]
- Silvestri, F. Contribution à la biologie de la petite Cécidomyie des olives (Prolasioptera Berlesiana Paoli) en Italie. Monit. Int. De La Prot. Des Plantes 1945, 19, 73–76. [Google Scholar]
- Paredes, D.; Cayuela, L.; Campos, M. Synergistic effects of ground cover and adjacent vegetation on natural enemies of olive insect pests. Agric. Ecosyst. Environ. 2013, 173, 72–80. [Google Scholar] [CrossRef]
- Rozas, L.; González-Ruiz, R. Primeros datos sobre la influencia de las cubiertas vegetales en la presencia de Raphidioptera (Insecta: Neuropteroidea) en olivares de Jaén. In Proceedings of the XVI Scientific-Technical Symposium on Olive Oil, Fundación del Olivar Jaén, Spain, 8–10 May 2013. [Google Scholar]
- Liñan de, C. Entomología Agroforestal: Insectos y Ácaros que Dañan Montes, Cultivos y Jardines; Ediciones Agrotécnicas, S.L.: Madrid, Spain, 1998; p. 1309. ISBN 9788487480546. [Google Scholar]
- Álvarez, H.A.; Jiménez-Muñoz, R.; Morente, M.; Campos, M.; Ruano, F. La presencia de cobertura vegetal en olivares ecológicos afecta a la interacción de los enemigos naturales contra Prays oleae, favoreciendo una depredación eficaz de los huevos. Agric. Ecosistemas Y Medio Ambiente 2021, 315, 107441. [Google Scholar]
- Azam, F. Comparative effects of organic and inorganic nitrogen sources applied to a flooded soil on rice yield and availability of N. Plant Soil 1990, 125, 255–262. [Google Scholar] [CrossRef]
- Singh, H. Nitrogen mineralization, microbial biomass and crop yield affected by the placement of wheat residues and fertilizers in a semi-arid tropical soil with minimum tillage. J. Appl. Ecol. 1995, 32, 588–595. [Google Scholar] [CrossRef]
- Rodríguez-Lizana, A.; Espejo-Pérez, A.J.; González-Fernández, P.; Ordóñez-Fernández, R. Los residuos de poda como alternativa al laboreo tradicional para reducir la erosión y la dispersión de contaminantes en los olivares. Contam. Del Agua El Aire Y El Suelo 2008, 193, 165–173. [Google Scholar]
- Burkert, A.; Bationo, A.; Dossa, K. Mechanisms of cereal growth induced by residue mulch increases in West Africa. Soil Sci. Soc. Am. J. 2000, 64, 346–358. [Google Scholar] [CrossRef] [Green Version]
- Shah, Z.; Shah, S.H.; Peoples, M.B.; Schwenke, G.D.; Herridge, D.F. Crop residue and fertiliser N effects on nitrogen fixation and yields of legume–cereal rotations and soil organic fertility. Field Crops Res. 2003, 83, 1–11. [Google Scholar] [CrossRef]
- Shafi, M.; Bakht, J.; Jan, M.T.; Shah, Z. Soil C and N dynamics and maize (Zea may L.) yield as affected by cropping systems and residue management in North-western Pakistan. Soil Tillage Res. 2007, 94, 520–529. [Google Scholar] [CrossRef]
- Calatrava, J.; Franco, J.A. Using pruning residues as mulch: Analysis of its adoption and process of diffusion in southern spain olive orchards. J. Environ. Manag. 2011, 92, 620–629. [Google Scholar] [CrossRef]
- Repullo, M.A.; Carbonell, R.; Hidalgo, J.; Rodríguez-Lizana, A.; Ordóñez, R. Using olive pruning residues to cover soil and improve fertility. Soil Tillage Res. 2012, 124, 36–46. [Google Scholar] [CrossRef]
- Taguas, E.V.; Gómez, J.A. Vulnerabilidad de los olivares bajo la actual normativa de la PAC (Política Agraria Común) sobre la erosión del suelo: Un caso de estudio en el Sur de España. Política De Uso De La Tierra 2015, 42, 683–694. [Google Scholar]
- Medina, J.; Monreal, C.; Barea, J.M.; Arriagada, C.; Borie, F.; Cornejo, P. Estabilización de residuos de cultivos y aplicación a suelos agrícolas y degradados: Una revisión. Gestión De Residuos 2015, 42, 41–54. [Google Scholar]
- Chang, Y.F.; Tauber, M.J.; Tauber, C.A. Reproduction and quality of F1 offspring in Chrysoperla carnea: Differential influence of quiescence, artificially-induced diapause, and natural diapause. J. Insect Physiol. 1996, 42, 521–528. [Google Scholar] [CrossRef]
- González-Ruiz, R.; Campos, M. Rearing of Cheiropachus quadrum (Hym.: Pteromalidae) from the olive beetle, Phloeotribus scarabaeoides (Col.: Scolytidae). Potential biological control agent. Redia 1990, 73, 495–505. [Google Scholar]
- Manojlovic, B.; Zabel, A.; Kostic, M.; Stankovic, S. Effect of nutrition of parasites with nectar of melliferous plants on parasitism of the elm bark beetles (col., scolytidae). J. Appl. Entomol. 2000, 124, 155–161. [Google Scholar] [CrossRef]
- Manojlović, B.; Zabel, A.; Stanković, S.; Kostić, M. Ecphylus silesiacus (ratz.) (hymenoptera, braconidae), an important elm bark beetle parasitoid. Agric. For. Entomol. 2000, 2, 63–67. [Google Scholar] [CrossRef]
- Udayagiri, S. Behavioral manipulation of natural enemies: Potential use in insect pest management. Agric. Zool. Rev. 1996, 7, 181–216. [Google Scholar]
- Tóth, M.; Bozsik, A.; Szentkirályi, F.; Letardi, A. Phenylacetaldehyde: A chemical attractant for common green lacewings (Chrysoperla carnea s.l., Neuroptera: Chrysopidae). Eur. J. Entomol. 2006, 103, 267. [Google Scholar] [CrossRef] [Green Version]
- Tóth, M.; Szentkirályi, F.; Vuts, J.; Letardi, A.; Tabilio, M.R.; Jaastad, G.; Knudsen, G.K. Optimization of a phenylacetaldehyde-based attractant for common green lacewings (Chrysoperla carnea sl). J. Chem. Ecol. 2009, 35, 449–458. [Google Scholar] [CrossRef]
- Bruce, T.J.; Cork, A.; Hall, D.R.; Dunkelblum, E. Laboratory and field evaluation of floral odours from African marigold, Tagetes erecta, and sweet pea, Lathyrus odoratus, as kairomones for the cotton bollworm Helicoverpa armigera. IOBC WPRS Bull. 2001, 25, 315–322. [Google Scholar]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents—A checklist of volatile compounds isolated by headspace techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Gliessman, S. La biodiversidad y la estabilidad de los agroecosistemas. In La Práctica de la Agricultura y la Ganadería Ecológicas; Cornejo, J., Ed.; CAAE: Sevilla, Spain, 2001; pp. 69–89. [Google Scholar]
- Foraster-Pulido, L. Las Cubiertas Vegetales en el Rediseño del Olivar para una Transición Agroecológica; Universidad Internacional de Andalucía: Seville, Spain, 2010; p. 104. ISBN 978-84-7993-173-5. Available online: https://core.ac.uk/download/pdf/72018622.pdf (accessed on 17 January 2023).
- Henry, C.S.; Brooks, S.J.; Duelli, P.; Johnson, J.B. A lacewing with the wanderlust: The European song species ‘Maltese’, Chrysoperla agilis, sp. n., of the carnea group of Chrysoperla (Neuroptera: Chrysopidae). Syst. Entomol. 2003, 28, 131–148. [Google Scholar] [CrossRef]
- Parrilla-González, J.A.; Ortega-Alonso, D. Social Innovation in Olive Oil Cooperatives: A Case Study in Southern Spain. Sustainability 2021, 13, 3934. [Google Scholar] [CrossRef]
- Alzoheiry, A.; Ghonimy, M.; El Rahman, E.A.; Abdelwahab, O.; Hassan, A. Improving olive mechanical harvesting using appropriate natural frequency. J. Agric. Eng. 2020, 51, 148–154. [Google Scholar] [CrossRef]
- Weihrauch, F. Overwintering of common green lacewings in hibernation shelters in the Hallertau hop growing area. Bull. Insectol. 2008, 61, 67–71. [Google Scholar]
- McEwen, P.K.; Åkerberg, C.; Bozsik, A.; James, C.J.; Eccleston, L.; Lenartsson, M.; Rossiter, P.; Tuovinen, T. Artificial overwintering chambers for green lacewings: Results of international trials and implications for pest control. J. Appl. Entomol. 2001, 123, 525–527. [Google Scholar] [CrossRef]
- Şengonca, Ç.; Frings, B. Enhancement of the green lacewing Chrysoperla carnea (Stephens) by providing artificial facilities for hibernation. Turk. Entomol. Derg. 1989, 13, 245–250. [Google Scholar]
- Şengonca, Ç.; Henze, M. Conservation and enhancement of Chrysoperla carnea (Stephens) (Neuroptera: Chrysopidae) in the field by providing hibernation shelters. J. Appl. Entomol. 1992, 114, 497–501. [Google Scholar] [CrossRef]
- Varela-Martínez, J.L.; González-Ruiz, R. Bases metodológicas para la evaluación del impacto ocasionado por las aplicaciones insecticidas sobre los enemigos naturales de las plagas del olivo (II). Phytoma 1999, 112, 32–42. [Google Scholar]
- Varela-Martínez, J.L.; González-Ruiz, R. La lucha química contra Prays oleae (lep., Yponomeutidae) y su influencia en los enemigos naturales de las plagas del olivar (y III). Phytoma 2000, 115, 35–47. [Google Scholar]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]








| Asteraceae | S1 | S2 | S3 | S4 | S5 | Total | t-Test | |
|---|---|---|---|---|---|---|---|---|
| Filago fuscescens Pomel | VC-MIX | 13 | 14 | 16 | 22 | 27 | 92 | p < 0.001 |
| VC | 5 | 0 | 2 | 0 | 4 | 11 | ||
| Filago pyramidata L. | VC-MIX | 105 | 73 | 137 | 64 | 111 | 490 | p < 0.001 |
| VC | 8 | 27 | 29 | 27 | 25 | 116 | ||
| Leontodon longirostris | VC-MIX | 0 | 0 | 0 | 0 | 0 | 0 | |
| (Finch & P. D. Sell) Talavera | VC | 0 | 9 | 1 | 0 | 0 | 10 | |
| Brassicaceae | ||||||||
| Diplotaxis virgata (Cav.) DC. | VC-MIX | 0 | 0 | 0 | 0 | 0 | 0 | p < 0.01 |
| VC | 5 | 15 | 5 | 6 | 4 | 35 | ||
| Caryophyllaceae | ||||||||
| Herniaria cinerea D. C. | VC-MIX | 12 | 5 | 39 | 5 | 12 | 73 | p > 0.05 |
| VC | 1 | 9 | 3 | 5 | 1 | 19 | ||
| Stellaria pallida (Dumort.) Piré | VC-MIX | 0 | 1 | 0 | 0 | 0 | 1 | |
| VC | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Fabaceae | ||||||||
| Astragalus hamosus L. | VC-MIX | 25 | 18 | 40 | 21 | 38 | 142 | p < 0.01 |
| VC | 0 | 1 | 1 | 3 | 2 | 7 | ||
| Melilotus elegans | VC-MIX | 0 | 0 | 2 | 0 | 0 | 2 | p > 0.05 |
| Salzm. ex Ser. | VC | 5 | 1 | 0 | 2 | 1 | 9 | |
| Ononis biflora Desf. | VC-MIX | 0 | 0 | 0 | 0 | 5 | 5 | |
| VC | 5 | 2 | 1 | 1 | 5 | 14 | ||
| Trigonella monspeliaca L. | VC-MIX | 1 | 0 | 1 | 2 | 2 | 6 | p < 0.01 |
| Geraniaceae | ||||||||
| Erodium malacoides (L.) L’Hér. | VC-MIX | 0 | 0 | 0 | 0 | 0 | 0 | |
| VC | 2 | 4 | 2 | 3 | 1 | 12 | ||
| Malvaceae | ||||||||
| Malva parviflora L. | VC-MIX | 80 | 43 | 30 | 62 | 49 | 264 | p < 0.05 |
| VC | 51 | 30 | 16 | 35 | 20 | 152 | ||
| Plantaginaceae | ||||||||
| Plantago lagopus L. | VC-MIX | 0 | 0 | 8 | 9 | 5 | 22 | |
| VC | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Poaceae | ||||||||
| Bromus rubens L. | VC-MIX | 30 | 25 | 15 | 106 | 50 | 226 | p > 0.05 |
| VC | 20 | 22 | 18 | 9 | 10 | 79 | ||
| Crucianella patula L. | VC-MIX | 0 | 0 | 0 | 0 | 0 | 0 | |
| VC | 6 | 10 | 24 | 2 | 5 | 47 | ||
| Hordeum murinum L. leporinum (Link) Arcang. | VC-MIX | 5 | 0 | 11 | 11 | 2 | 29 | p > 0.05 |
| VC | 0 | 2 | 1 | 0 | 2 | 5 | ||
| Phalaris minor Retz. | VC-MIX | 1 | 0 | 5 | 3 | 2 | 11 | p > 0.05 |
| VC | 1 | 3 | 1 | 2 | 5 | 12 | ||
| Trisetaria panicea (Lam.) Paunero | VC-MIX | 1 | 0 | 5 | 3 | 2 | 11 | p > 0.05 |
| VC | 1 | 3 | 1 | 2 | 5 | 12 | ||
| Primulaceae | ||||||||
| Anagallis foemina Mill. | VC-MIX | 0 | 0 | 1 | 0 | 0 | 1 | |
| VC | 0 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Ruiz, R.; Gómez-Guzmán, J.A.; Martínez-Rojas, M.; García-Fuentes, A.; Cordovilla, M.d.P.; Sainz-Pérez, M.; Sánchez-Solana, A.M.; Carlos-Hervás, J.; Rodríguez-Lizana, A. The Influence of Mixed Green Covers, a New Trend in Organic Olive Growing, on the Efficiency of Predatory Insects. Agriculture 2023, 13, 785. https://doi.org/10.3390/agriculture13040785
González-Ruiz R, Gómez-Guzmán JA, Martínez-Rojas M, García-Fuentes A, Cordovilla MdP, Sainz-Pérez M, Sánchez-Solana AM, Carlos-Hervás J, Rodríguez-Lizana A. The Influence of Mixed Green Covers, a New Trend in Organic Olive Growing, on the Efficiency of Predatory Insects. Agriculture. 2023; 13(4):785. https://doi.org/10.3390/agriculture13040785
Chicago/Turabian StyleGonzález-Ruiz, Ramón, José Alfonso Gómez-Guzmán, Minerva Martínez-Rojas, Antonio García-Fuentes, María del Pilar Cordovilla, María Sainz-Pérez, Antonio M. Sánchez-Solana, Juan Carlos-Hervás, and Antonio Rodríguez-Lizana. 2023. "The Influence of Mixed Green Covers, a New Trend in Organic Olive Growing, on the Efficiency of Predatory Insects" Agriculture 13, no. 4: 785. https://doi.org/10.3390/agriculture13040785