
Root Characteristics for Maize with the Highest Grain Yield Potential of 22.5 Mg ha−1 in China
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
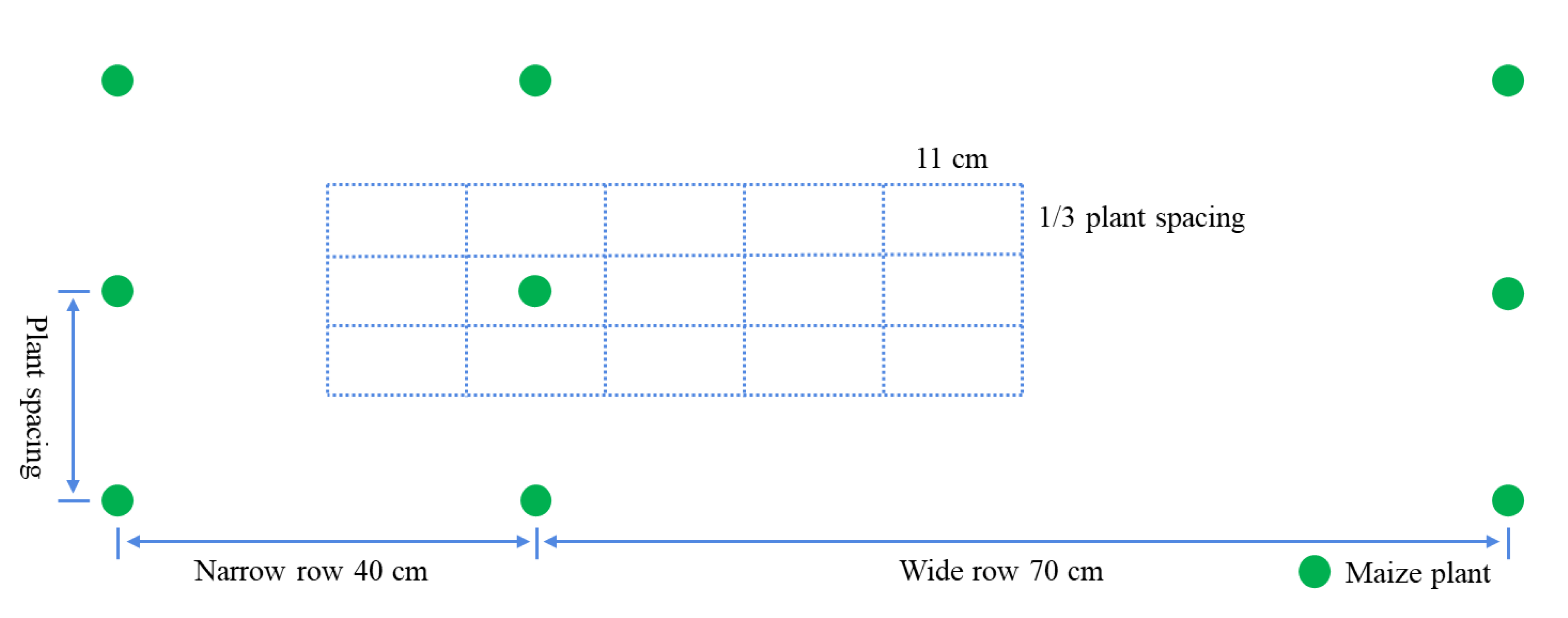
2.1. Experimental Design
2.2. Sampling and Measurements
2.2.1. Root Sampling Method
2.2.2. Root Index Analysis Methods
2.2.3. Grain Yield Determination Method
2.3. Statistical Analysis
3. Results
3.1. Grain Yield under Different Planting Densities
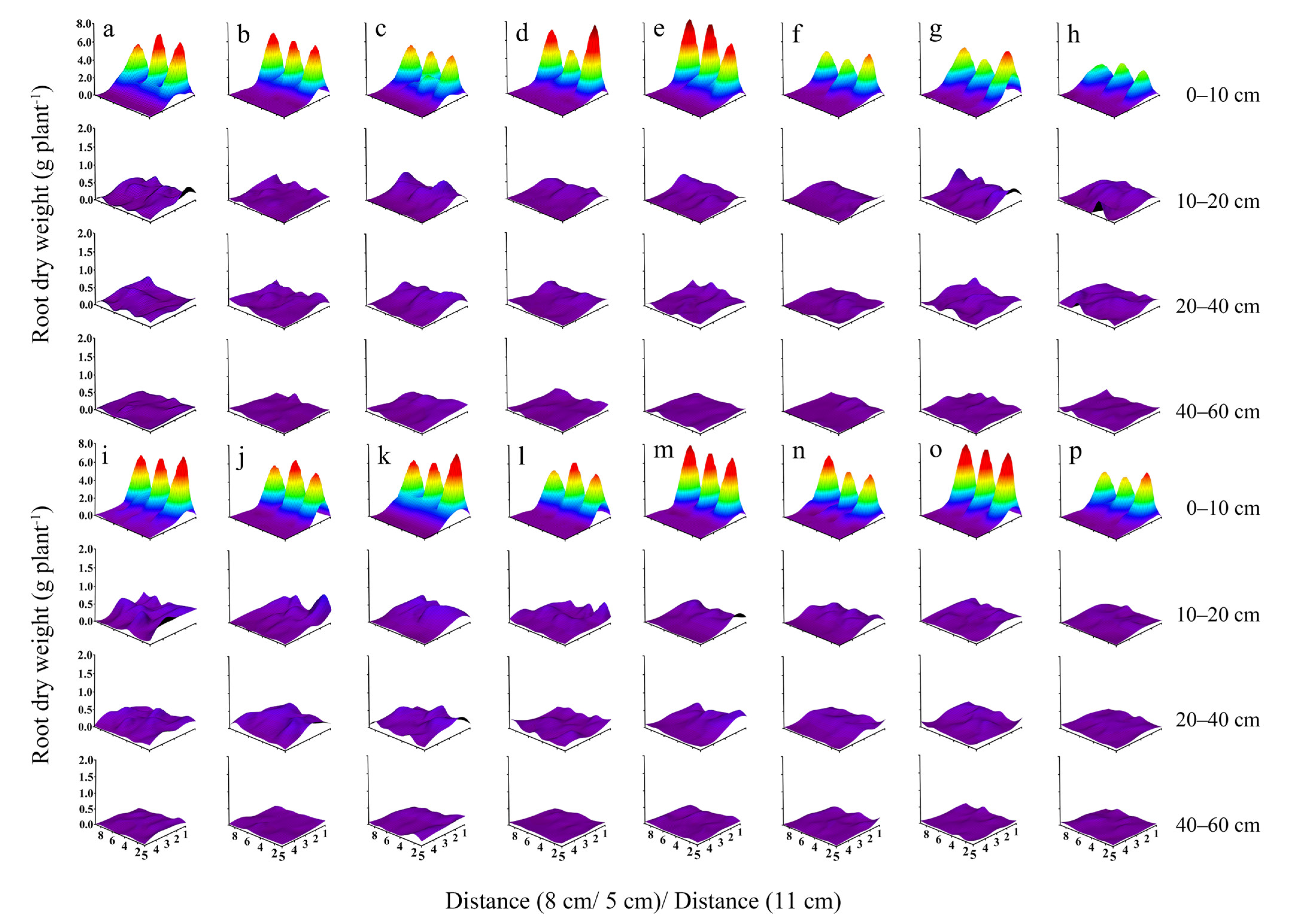
3.2. Root Dry Weight
3.3. Root Weight Density
3.4. Root Length
3.5. Root Length Density
3.6. Interaction Effects of Years, Planting Densities, and Cultivars on the Maize Root Characteristics
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, Y.; Lu, C. Future grain consumption trends and implications on grain security in China. Sustainability 2019, 11, 5165. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Chen, Y.; Wang, J. Global food security under COVID-19: Comparison and enlightenment of policy responses in different countries. Foods 2021, 10, 2850. [Google Scholar] [CrossRef] [PubMed]
- Smith, P. Delivering food security without increasing pressure on land. Glob. Food Secur. 2013, 2, 18–23. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, J.W.; LÜ, P.; Yang, J.S.; Liu, P.; Dong, S.T.; Li, D.H.; Sun, Q.Q. Effect of plant density on grain yield dry matter accumulation and partitioning in summer maize cultivar Denghai 661. Acta Agron. Sin. 2011, 37, 1301–1307. [Google Scholar] [CrossRef]
- Hou, P.; Liu, Y.; Liu, W.; Liu, G.; Xie, R.; Wang, K.; Ming, B.; Wang, Y.; Zhao, R.; Zhang, W.; et al. How to increase maize production without extra nitrogen input. Resour. Conserv. Recyclin. 2020, 160, 104913. [Google Scholar] [CrossRef]
- Wu, H.Y.; Qiao, M.Y.; Zhang, W.F.; Wang, K.R.; Li, S.K.; Jiang, C.D. Systemic regulation of photosynthetic function in maize plants at graining stage under a vertically heterogeneous light environment. J. Integr. Agric. 2022, 21, 666–676. [Google Scholar] [CrossRef]
- Ma, D.L.; Xie, R.Z.; Niu, X.K.; Li, S.K.; Long, H.L.; Liu, Y.E. Changes in the morphological traits of maize genotypes in China between the 1950s and 2000s. Eur. J. Agron. 2014, 58, 1–10. [Google Scholar] [CrossRef]
- Liu, G.; Yang, Y.; Guo, X.; Liu, W.; Xie, R.; Ming, B.; Xue, J.; Wang, K.; Li, S.; Hou, P. Coordinating maize source and sink relationship to achieve yield potential of 22.5 Mg ha-1. Field Crops Res. 2022, 283, 108544. [Google Scholar] [CrossRef]
- Liu, G.; Hou, P.; Xie, R.; Ming, B.; Wang, K.; Liu, W.; Yang, Y.; Xu, W.; Chen, J.; Li, S. Nitrogen uptake and response to radiation distribution in the canopy of high-yield Maize. Crop Sci. 2019, 59, 1236–1247. [Google Scholar] [CrossRef]
- Shao, H.; Xia, T.; Wu, D.; Chen, F.; Mi, G. Root growth and root system architecture of field-grown maize in response to high planting density. Plant Soil. 2018, 430, 395–411. [Google Scholar] [CrossRef]
- Morris, E.C.; Griffiths, M.; Golebiowska, A.; Mairhofer, S.; Burr-Hersey, J.; Goh, T.; Wangenheim, D.V.; Atkinson, B.; Sturrock, C.J.; Lynch, J.P.; et al. Shaping 3D root system architecture. Curr. Biol. 2017, 27, 919–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, A.; Lynch, J.P. Reduced frequency of lateral root branching improves N capture from low-N soils in maize. J. Exp. Bot. 2015, 66, 2055–2065. [Google Scholar] [CrossRef] [Green Version]
- Ning, P.; Li, S.; White, P.J.; Li, C. Maize varieties released in different eras Have similar root length density distributions in the Soil, Which are negatively correlated with local concentrations of soil mineral nitrogen. PLoS ONE 2015, 10, e0121892. [Google Scholar] [CrossRef] [PubMed]
- Ning, P.; Li, S.; Li, X.; Li, C. New maize hybrids had larger and deeper post-silking root than old ones. Field Crops Res. 2014, 166, 66–71. [Google Scholar] [CrossRef]
- Reynolds, M.; Atkin, O.K.; Bennett, M.; Cooper, M.; Dodd, I.C.; Foulkes, M.J.; Frohberg, C.; Hammer, G.; Henderson, I.R.; Huang, B.; et al. Addressing research bottlenecks to crop productivity. Trends Plant Sci. 2021, 26, 607–630. [Google Scholar] [CrossRef] [PubMed]
- Grabarnik, P.; Pagèes, L.; Bengough, A.G. Geometrical properties of simulated maize root systems: Consequences for length density and intersection density. Plant Soil 1998, 200, 157–167. [Google Scholar] [CrossRef]
- Tang, L.; Sun, H.; Sun, R.; Niu, Y.; Song, J.; Li, S.; Shen, Y. Optimized nitrogen application increases soil water extraction by changing in-season maize root morphology and distribution in rainfed farmland. Agronomy 2020, 10, 1606. [Google Scholar] [CrossRef]
- Oikeh, S.O.; Kling, J.G.; Horst, W.J.; Chude, V.O.; Carsky, R.J. Growth and distribution of maize roots under nitrogen fertilization in plinthite soil. Field Crops Res. 1999, 62, 1–13. [Google Scholar] [CrossRef]
- Herder, G.D.; Isterdael, G.V.; Beeckman, T.; Smet, I.D. The roots of a new green revolution. Trends Plant Sci. 2010, 15, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Forde, B.; Lorenzo, H. The nutritional control of root development. Plant Soil 2001, 232, 51–68. [Google Scholar] [CrossRef]
- Hammer, G.L.; Dong, Z.; McLean, G.; Doherty, A.; Messina, C.; Schussler, J.; Zinselmeier, C.; Paszkiewicz, S.; Cooper, M. Can changes in canopy and/or root system architecture explain historical maize yield trends in the U.S. Corn Belt? Crop Sci. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Liu, G.; Hou, P.; Xie, R.; Ming, B.; Wang, K.; Xu, W.; Liu, W.; Yang, Y.; Li, S. Canopy characteristics of high-yield maize with yield potential of 22.5 Mg ha−1. Field Crops Res. 2017, 213, 221–230. [Google Scholar] [CrossRef]
- Guo, X.; Yang, Y.; Liu, H.; Liu, G.; Liu, W.; Wang, Y.; Zhao, R.; Ming, B.; Xie, R.; Wang, K.; et al. Effects of solar radiation on root and shoot growth of maize and the quantitative relationship between them. Crop Sci. 2020, 61, 1414–1425. [Google Scholar] [CrossRef]
- Liu, G.; Yang, Y.; Liu, W.; Guo, X.; Xie, R.; Ming, B.; Xue, J.; Zhang, G.; Li, R.; Wang, K.; et al. Optimized canopy structure improves maize grain yield and resource use efficiency. Food Energy Secur. 2022, 11, e375. [Google Scholar] [CrossRef]
- Liu, G.; Yang, Y.; Guo, X.; Liu, W.; Xie, R.; Ming, B.; Xue, J.; Wang, K.; Li, S.; Hou, P. A global analysis of dry matter accumulation and allocation for maize yield breakthrough from 1.0 to 25.0 Mg ha−1. Resour. Conserv. Recycl. 2023, 188, 106656. [Google Scholar] [CrossRef]
- Gao, J.; Shi, J.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Grain yield and root characteristics of summer maize (Zea mays L.) under shade stress conditions. J. Agron. Crop Sci. 2017, 203, 562–573. [Google Scholar] [CrossRef]
- Gao, Y.; Duan, A.; Qiu, X.; Liu, Z.; Sun, J.; Zhang, J.; Wang, H. Distribution of roots and root length density in a maize/soybean strip intercropping system. Agric. Water Manag. 2010, 98, 199–212. [Google Scholar] [CrossRef]
- Liedgens, M.; Richner, W. Relation between maize (Zea mays L.) leaf area and root density observed with minirhizotrons. Eur. J. Agron. 2001, 15, 131–141. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, cheap and deep: An ideotype to optimize water and N acquisition by maize root systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Bates, T.R.; Lynch, J.P. Root hairs confer a competitive advantage under low phosphorus availability. Plant Soil 2001, 236, 243–250. [Google Scholar] [CrossRef]
- Guo, X.; Yang, Y.; Liu, H.; Liu, G.; Liu, W.; Wang, Y.; Zhao, R.; Ming, B.; Xie, R.; Wang, K.; et al. Effects of solar radiation on dry matter distribution and root morphology of high yielding maize cultivars. Agriculture 2022, 12, 299. [Google Scholar] [CrossRef]
- Borrell, A.; Hammer, G.; Oosterom, E.V. Stay-green: A consequence of the balance between supply and demand for nitrogen during grain filling? Ann. Appl. Biol. 2001, 138, 91–95. [Google Scholar] [CrossRef]
- Liu, G.; Yang, H.; Xie, R.; Yang, Y.; Liu, W.; Guo, X.; Xue, J.; Ming, B.; Wang, K.; Hou, P.; et al. Genetic gains in maize yield and related traits for high-yielding cultivars released during 1980s to 2010s in China. Field Crops Res. 2021, 270, 108223. [Google Scholar] [CrossRef]
- Huang, Z.H.; Wang, S.Y.; Bao, Y.; Liang, X.H.; Sun, G.; Shen, L.; Cao, Y.; Wu, C.S. Studies on dry matter accumulation and distributive characteristic in super high-yield maize. J. Maize Sci. 2007, 15, 95–98. [Google Scholar] [CrossRef]
- Zhang, L.; Liang, Z.Y.; He, X.M.; Meng, Q.F.; Hu, Y.; Schmidhalter, U.; Zhang, W.; Zou, C.Q.; Chen, X.P. Improving grain yield and protein concentration of maize (Zea mays L.) simultaneously by appropriate hybrid selection and nitrogen management. Field Crops Res. 2020, 249, 107754. [Google Scholar] [CrossRef]
- Borch, K.; Bouma, T.J.; Lynch, J.P.; Brown, K.M. Ethylene: A regulator of root architectural responses to soil phosphorus availability. Plant Cell Environ. 1999, 22, 425–431. [Google Scholar] [CrossRef]
- Qin, H.; He, L.; Huang, R. The coordination of ethylene and other hormones in primary root development. Front Plant Sci. 2019, 10, 874. [Google Scholar] [CrossRef]
- Zhang, F.L.; Niu, X.K.; Zhang, Y.M.; Xie, R.Z.; Liu, X.; Li, S.K.; Gao, S.J. Studies on the root characteristics of maize varieties of different eras. J. Integr. Agric. 2013, 12, 426–435. [Google Scholar] [CrossRef]
- Vetterlein, D.; Phalempin, M.; Lippold, E.; Schlüter, S.; Schreiter, S.; Ahmed, M.A.; Carminati, A.; Duddek, P.; Jorda, H.; Bienert, G.P.; et al. Root hairs matter at field scale for maize shoot growth and nutrient uptake, but root trait plasticity is primarily triggered by texture and drought. Plant Soil 2022, 478, 119–141. [Google Scholar] [CrossRef]
- Stirbet, A.; Lazár, D.; Guo, Y.; Govindjee, G. Photosynthesis: Basics, history and modelling. Ann. Bot. 2020, 126, 511–537. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Organic Matter (g kg−1) | Alkaline N (mg kg−1) | Olsen P (mg kg−1) | Available K (mg kg−1) | Bulk Density (g cm−3) | pH |
|---|---|---|---|---|---|---|
| 2019 | 14.1 | 87.6 | 53.8 | 108.6 | 1.37 | 7.97 |
| 2021 | 17.8 | 78.0 | 66.3 | 330.1 | 1.34 | 7.75 |
| Year | Tmax (°C) | Tmin (°C) | Td (°C) | Sr (M Jm−2 day−1) | Pre (mm) |
|---|---|---|---|---|---|
| 2019 | 27.1 | 12.3 | 14.8 | 9.8 | 189.1 |
| 2021 | 28.5 | 10.1 | 18.4 | 9.4 | 159.8 |
| Sources of Variation | Root Dry Weight | Root Weight Density | Root Length | Root Length Density |
|---|---|---|---|---|
| Year | ** | * | ns | ns |
| Planting density | ** | ** | ** | ** |
| Cultivar | ns | ns | ** | ** |
| Year × Planting density | ns | ns | ns | ns |
| Year × Cultivar | ns | ns | * | * |
| Planting density × Cultivar | ns | ns | ns | ns |
| Year × Planting density × Cultivar | ns | ns | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Liu, G.; Yang, Y.; Guo, X.; Jin, S.; Xie, R.; Ming, B.; Xue, J.; Wang, K.; Li, S.; et al. Root Characteristics for Maize with the Highest Grain Yield Potential of 22.5 Mg ha−1 in China. Agriculture 2023, 13, 765. https://doi.org/10.3390/agriculture13040765
Zhang L, Liu G, Yang Y, Guo X, Jin S, Xie R, Ming B, Xue J, Wang K, Li S, et al. Root Characteristics for Maize with the Highest Grain Yield Potential of 22.5 Mg ha−1 in China. Agriculture. 2023; 13(4):765. https://doi.org/10.3390/agriculture13040765
Chicago/Turabian StyleZhang, Long, Guangzhou Liu, Yunshan Yang, Xiaoxia Guo, Shuai Jin, Ruizhi Xie, Bo Ming, Jun Xue, Keru Wang, Shaokun Li, and et al. 2023. "Root Characteristics for Maize with the Highest Grain Yield Potential of 22.5 Mg ha−1 in China" Agriculture 13, no. 4: 765. https://doi.org/10.3390/agriculture13040765