
Positive Effect of High Zinc on Growth of Sedum alfredii

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Pre-Cultivation
2.2. Plant Growth and Biomass
2.3. Element Analysis
2.4. Metabolite Measurements
2.5. Data Processing
2.6. Statistical Analysis
3. Results
3.1. Effects of Zn on Plant Growth and Zn Concentration
3.2. Sample Quality Inspection and Overview of the Metabolites Profiles
3.3. Multidimensional Statistical Analysis
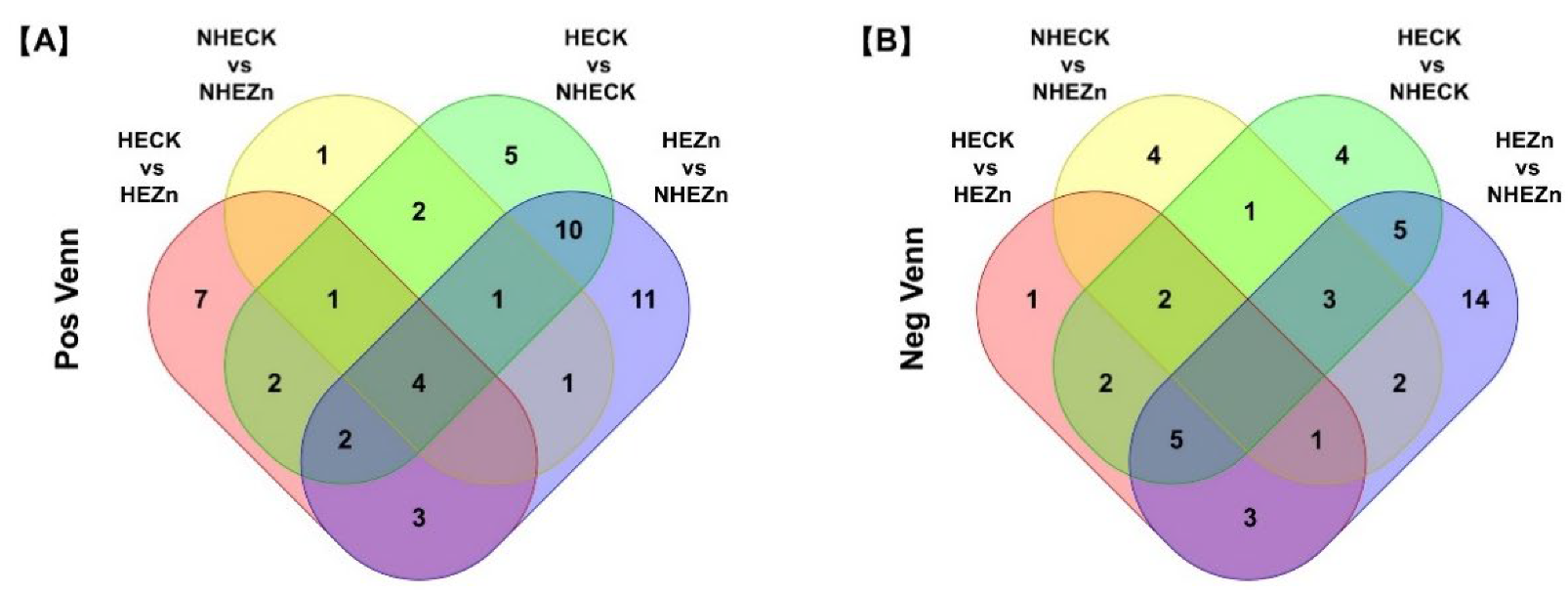
3.4. Fold Change Analysis
3.5. KEGG Enrichment Analysis
4. Discussion
4.1. Effect of High Zn on Growth of S. alfredii
4.2. Changes of Organic Acids and Their Derivatives Induced by Zn
4.3. Changes of Lipids and Lipid Molecules Induced by Zn
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef]
- Broadley, M.R.; White, P.J.; Hammond, J.P.; Zelko, I.; Lux, A. Zinc in plants. New Phytol. 2007, 173, 677–702. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.; Zahir, Z.A.; Ditta, A.; Tahir, M.U.; Ahmad, M.; Mumtaz, M.Z.; Hayat, K.; Hussain, S. Production and implication of bio-activated organic fertilizer enriched with zinc-solubilizing bacteria to boost up maize (Zea mays L.) production and biofortification under two cropping seasons. Agronomy 2020, 10, 39. [Google Scholar] [CrossRef]
- Noman, A.; Aqeel, M.; Khalid, N.; Islam, W.; Sanaullah, T.; Anwar, M.; Khan, S.; Ye, W.; Lou, Y. Zinc finger protein transcription factors: Integrated line of action for plant antimicrobial activity. Microb. Pathog. 2019, 132, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Hansch, R.; Mendel, R.R. Physiological functions of mineral micronutrients (Cu, Zn, Mn, Fe, Ni, Mo, B, Cl). Curr. Opin. Plant Biol. 2009, 12, 259–266. [Google Scholar] [CrossRef]
- Palmer, C.M.; Guerinot, M.L. Facing the challenges of Cu, Fe and Zn homeostasis in plants. Nat. Chem. Biol. 2009, 5, 333–340. [Google Scholar] [CrossRef]
- Cakmak, I. Possible roles of zinc in protecting plant cells from damage by reactive oxygen species. New Phytol. 2000, 146, 185–205. [Google Scholar] [CrossRef]
- Castillo-González, J.; Ojeda-Barrios, D.; Hernández-Rodríguez, A.; González-Franco, A.C.; Robles-Hernández, L.; López-Ochoa, G.R. Zinc Metalloenzymes in Plants. Interciencia 2018, 43, 242–248. [Google Scholar]
- Natasha, N.; Shahid, M.; Bibi, I.; Iqbal, J.; Khalid, S.; Murtaza, B.; Bakhat, H.F.; Farooq, A.B.U.; Amjad, M.; Hammad, H.M.; et al. Zinc in soil-plant-human system: A data-analysis review. Sci. Total Environ. 2021, 808, 152024. [Google Scholar] [CrossRef]
- Wei, C.; Jiao, Q.; Agathokleous, E.; Liu, H.; Li, G.; Zhang, J.; Fahad, S.; Jiang, Y. Hormetic effects of zinc on growth and antioxidant defense system of wheat plants. Sci. Total Environ. 2022, 807 Pt 2, 150992. [Google Scholar] [CrossRef]
- Gu, H.-H.; Zhan, S.-S.; Wang, S.-Z.; Tang, Y.-T.; Chaney, R.L.; Fang, X.-H.; Cai, X.-D.; Qiu, R.-L. Silicon-mediated amelioration of zinc toxicity in rice (Oryza sativa L.) seedlings. Plant Soil 2012, 350, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Bankaji, I.; Pérez-Clemente, R.; Caçador, I.; Sleimi, N. Accumulation potential of Atriplex halimus to zinc and lead combined with NaCl: Effects on physiological parameters and antioxidant enzymes activities. S. Afr. J. Bot. 2019, 123, 51–61. [Google Scholar] [CrossRef]
- Chakraborty, S.; Mishra, A.K. Mitigation of zinc toxicity through differential strategies in two species of the cyanobacterium Anabaena isolated from zinc polluted paddy field. Environ. Pollut. 2020, 263 Pt B, 114375. [Google Scholar] [CrossRef]
- Mateos-Naranjo, E.; Perez-Romero, J.A.; Redondo-Gómez, S.; Mesa-Marín, J.; Castellanos, E.M.; Davy, A.J. Salinity alleviates zinc toxicity in the saltmarsh zinc-accumulator Juncus acutus. Ecotoxicol. Environ. Saf. 2018, 163, 478–485. [Google Scholar] [CrossRef] [PubMed]
- Bernardy, K.; Farias, J.G.; Pereira, A.S.; Dorneles, A.O.S.; Bernardy, D.; Tabaldi, L.A.; Neves, V.M.; Dressler, V.L.; Nicoloso, F.T. Plants’ genetic variation approach applied to zinc contamination: Secondary metabolites and enzymes of the antioxidant system in Pfaffia glomerata accessions. Chemosphere 2020, 253, 126692. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, G.P.S.; Bali, A.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Insights into the tolerance and phytoremediation potential of Coronopus didymus L. (Sm) grown under zinc stress. Chemosphere 2019, 244, 125350. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.E.; Long, X.X.; Ye, H.B.; He, Z.L.; Calvert, D.V.; Stoffella, P.J. Cadmium tolerance and hyperaccumulation in a new Zn-hyperaccumulating plant species (Sedum alfredii Hance). Plant Soil 2004, 259, 181–189. [Google Scholar] [CrossRef]
- Yang, X.E.; Li, T.Q.; Long, X.X.; Xiong, Y.H.; He, Z.L.; Stoffella, P.J. Dynamics of zinc uptake and accumulation in the hyperaccumulating and non-hyperaccumulating ecotypes of Sedum alfredii Hance. Plant Soil 2006, 284, 109–119. [Google Scholar] [CrossRef]
- Li, T.-Q.; Yang, X.-E.; Jin, X.-F.; He, Z.-L.; Stoffella, P.-J.; Hu, Q.-H. Root responses and metal accumulation in two contrasting ecotypes of Sedum alfredii Hance under lead and zinc toxic stress. J. Environ. Sci. Health Part A-Toxic/Hazard. Subst. Environ. Eng. 2005, 40, 1081–1096. [Google Scholar]
- Li, T.; Yang, X.; Lu, L.; Islam, E.; He, Z. Effects of zinc and cadmium interactions on root morphology and metal translocation in a hyperaccumulating species under hydroponic conditions. J. Hazard. Mater. 2009, 169, 734–741. [Google Scholar] [CrossRef]
- Lu, L.; Tian, S.; Zhang, J.; Yang, X.; Labavitch, J.M.; Webb, S.M.; Latimer, M.; Brown, P.H. Efficient xylem transport and phloem remobilization of Zn in the hyperaccumulator plant species Sedum alfredii. New Phytol. 2013, 198, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Patti, G.J.; Yanes, O.; Siuzdak, G. Innovation: Metabolomics: The apogee of the omics trilogy. Nat. Rev. Mol. Cell Biol. 2012, 13, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Xu, L.; Shen, H.; Wang, J.; Liu, W.; Zhu, X.; Wang, R.; Sun, X.; Liu, L. Metabolomic analysis with GC-MS to reveal potential metabolites and biological pathways involved in Pb &Cd stress response of radish roots. Sci. Rep. 2015, 5, 18296. [Google Scholar] [PubMed]
- Wang, J.; Chen, X.; Chu, S.; You, Y.; Chi, Y.; Wang, R.; Yang, X.; Hayat, K.; Zhang, D.; Zhou, P. Comparative cytology combined with transcriptomic and metabolomic analyses of Solanum nigrum L. in response to Cd toxicity. J. Hazard. Mater. 2022, 423, 127168. [Google Scholar] [CrossRef]
- Ghatak, A.; Chaturvedi, P.; Weckwerth, W. Metabolomics in Plant Stress Physiology. Adv. Biochem. Eng. Biotechnol. 2018, 164, 187–236. [Google Scholar]
- Tang, Q.; Zheng, X.; Chen, W.; Ye, X.; Tu, P. Metabolomics reveals key resistant responses in tomato fruit induced by Cryptococcus laurentii. Food Chem. Mol. Sci. 2022, 4, 100066. [Google Scholar] [CrossRef]
- Ogura, T.; Bamba, T.; Fukusaki, E. Development of a practical metabolite identification technique for non-targeted metabolomics. J. Chromatogr. A 2013, 1301, 73–79. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Ayala, J.; Liu, Y.; An, J.; Wang, D.; Cai, Z.; Hou, R.; Zhang, M. Possible Effects of Early Maternal Separation on the Gut Microbiota of Captive Adult Giant Pandas. Animals 2022, 12, 2587. [Google Scholar] [CrossRef]
- Yeo, H.J.; Baek, S.-A.; Sathasivam, R.; Kim, J.K.; Park, S.U. Metabolomic analysis reveals the interaction of primary and secondary metabolism in white, pale green, and green pak choi (Brassica rapa subsp. chinensis). Appl. Biol. Chem. 2021, 64, 3. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant Secondary Metabolites as Defenses, Regulators, and Primary Metabolites: The Blurred Functional Trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Tian, S.; Lu, L.; Zhang, J.; Wang, K.; Brown, P.; He, Z.; Liang, J.; Yang, X. Calcium protects roots of Sedum alfredii H. against cadmium-induced oxidative stress. Chemosphere 2011, 84, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Li, L.; Tang, S.; Liu, C.; Fu, X.; Shi, Z.; Mao, H. Metabolomics Reveals that Crossbred Dairy Buffaloes Are More Thermotolerant than Holstein Cows under Chronic Heat Stress. J. Agric. Food Chem. 2018, 66, 12889–12897. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Gong, X.; Cai, Y.; Guo, Y.; Tu, J.; Li, H.; Zhang, T.; Wang, J.; Xue, F.; Zhu, Z.-J. Normalization and integration of large-scale metabolomics data using support vector regression. Metabolomics 2016, 12, 89. [Google Scholar] [CrossRef]
- Dos Santos, L.R.; da Silva, B.R.S.; Pedron, T.; Batista, B.L.; Lobato, A.K.D.S. 24-Epibrassinolide Improves Root Anatomy and Antioxidant Enzymes in Soybean Plants Subjected to Zinc Stress. J. Soil Sci. Plant Nutr. 2020, 20, 105–124. [Google Scholar] [CrossRef]
- Anjum, N.A.; Singh, H.P.; Khan, M.I.R.; Masood, A.; Per, T.S.; Negi, A.; Batish, D.R.; Khan, N.; Duarte, A.; Pereira, E.; et al. Too much is bad-an appraisal of phytotoxicity of elevated plant-beneficial heavy metal ions. Environ. Sci. Pollut. Res. 2015, 22, 3361–3382. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, Y.; Zhang, R.; Luo, J.; Song, Y.; Li, J.; Wu, K.; Peng, L.; Liu, Y.; Du, Y.; et al. Hemicellulose modification promotes cadmium hyperaccumulation by decreasing its retention on roots in Sedum alfredii. Plant Soil 2020, 447, 241–255. [Google Scholar] [CrossRef]
- Guo, X.; Luo, J.; Du, Y.; Li, J.; Liu, Y.; Liang, Y.; Li, T. Coordination between root cell wall thickening and pectin modification is involved in cadmium accumulation in Sedum alfredii. Environ. Pollut. 2021, 268, 115665. [Google Scholar] [CrossRef]
- Rauser, W.E. Structure and function ofmetal chelators produced by plants. The case for organic acids, amino acids, phytin and metallothioneins. Cell Biochem. Biophys. 1999, 31, 19–48. [Google Scholar] [CrossRef]
- Mathys, W. The Role of Malate, Oxalate, and Mustard Oil Glucosides in the Evolution of ZincResistance in Herbage Plants. Physiol. Plant. 1977, 40, 130–136. [Google Scholar] [CrossRef]
- Broadley, M.R.; Rose, T.; Frei, M.; Pariasca-Tanaka, J.; Yoshihashi, T.; Thomson, M.; Hammond, J.P.; Aprile, A.; Close, T.J.; Ismail, A.M.; et al. Response to zinc deficiency of two rice lines with contrasting tolerance is determined by root growth maintenance and organic acid exudation rates, and not by zinc-transporter activity. New Phytol. 2010, 186, 400–414. [Google Scholar]
- Lavell, A.A.; Benning, C. Cellular Organization and Regulation of Plant Glycerolipid Metabolism. Plant Cell Physiol. 2019, 60, 1176–1183. [Google Scholar] [CrossRef] [PubMed]
- Ruelland, E.; Kravets, V.; Derevyanchuk, M.; Martinec, J.; Zachowski, A.; Pokotylo, I. Role of phospholipid signalling in plant environmental responses. Environ. Exp. Bot. 2015, 114, 129–143. [Google Scholar] [CrossRef]
- Rontani, J.; Rabourdin, A.; Pinot, F.; Kandel, S.; Aubert, C. Visible light-induced oxidation of unsaturated components of cutins: A significant process during the senescence of higher plants. Phytochemistry 2005, 66, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Moon, H.; Chowrira, G.; Kunst, L. Heterologous expression of a fatty acid hydroxylase gene in developing seeds of Arabidopsis thaliana. Planta 2003, 217, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.A.; Khalil, I.; Varanini, Z.; Pinton, R. Increase in NAD(P)H-dependent generation of active oxygen species and changes in lipid composition of microsomes isolated from roots of zinc-deficient bean plants. J. Plant Nutr. 2000, 23, 285–295. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adduct | Name | VIP | FC | p | m/z | rt (s) |
|---|---|---|---|---|---|---|
| [M+H]+ | Tetradecanoylcarnitine | 3.76 | 0.21 | 0.00 | 372 | 10.99 |
| [M+H−H2O]+ | 12,19,20-Trihydroxy-14-methylenegeranylnerol | 3.84 | 0.28 | 0.00 | 337 | 10.88 |
| [M+H]+ | 1-Hexadecanoyl-sn-glycero-3-phosphoethanolamine | 1.44 | 0.57 | 0.00 | 454 | 10.89 |
| [M+H]+ | Glyceryl monooleate | 1.34 | 0.34 | 0.00 | 357 | 11.25 |
| [M+H]+ | 3alpha,17beta-Dihydroxy-5alpha-androstane | 2.06 | 0.48 | 0.00 | 293 | 11.51 |
| [M+H]+ | Anileridine | 1.65 | 0.44 | 0.00 | 353 | 7.35 |
| [M+H]+ | 1-Stearoyl-sn-glycero-3-phosphocholine | 1.05 | 0.56 | 0.00 | 524 | 11.42 |
| [M+H]+ | 2-Aminooctadecane-1,3,4-triol | 2.21 | 0.57 | 0.00 | 318 | 8.50 |
| [M]+ | Lauramidopropylbetaine | 1.18 | 0.21 | 0.00 | 343 | 8.88 |
| [M+H]+ | [4,6-diethyl-6-(4-ethyl-2-methyloctyl)-3H-1,2-dioxin-3-yl]acetic acid | 1.02 | 0.53 | 0.00 | 355 | 10.30 |
| [M+NH4]+ | 1-O-linoleoyl-3-O-beta-D-galactopyranosyl-syn-glycerol | 1.06 | 0.63 | 0.00 | 534 | 10.55 |
| [M+H-H2O]+ | 5-dodecyl-4-hydroxy-4-methylcyclopent-2-en-1-one | 4.37 | 0.72 | 0.00 | 263 | 10.88 |
| [M+Na]+ | Deoxykhivorin | 1.02 | 0.16 | 0.00 | 593 | 11.71 |
| [M+H]+ | 6-{9a,11a-dimethyl-1H,2H,3H,3aH,3bH,4H,8H,9H,9bH,10H,11H-cyclopenta[a]phenanthren-1-yl}-3-ethyl-2-methylheptane | 1.57 | 1.18 | 0.01 | 397 | 12.65 |
| [M+H]+ | Lupenone | 1.26 | 0.28 | 0.01 | 425 | 12.98 |
| [M+Na]+ | 3-Deoxo-3-beta-Acetoxydeoxydihydrogedunin | 2.06 | 0.05 | 0.02 | 535 | 12.17 |
| [M+H-H2O]+ | DG(18:2(9Z,12Z)/18:2(9Z,12Z)) | 3.11 | 0.73 | 0.02 | 600 | 12.27 |
| [M+H]+ | Fenpropimorph | 2.00 | 0.85 | 0.03 | 304 | 11.19 |
| [M+H]+ | PC 32:4|PC 14:1_18:3 | 2.84 | 0.62 | 0.04 | 727 | 12.16 |
| [M-H]− | Linoleic acid | 2.07 | 0.53 | 0.00 | 279 | 11.30 |
| [M−H]− | 2-Azaniumylethyl (3-hexadecanoyloxy-2-hydroxypropyl) phosphate | 1.58 | 0.46 | 0.00 | 452 | 10.88 |
| [M−H]− | (Z)-5,8,11-Trihydroxyoctadec-9-enoic acid | 2.41 | 0.40 | 0.00 | 329 | 7.35 |
| [M−H]− | (15Z)-9,12,13-Trihydroxy-15-octadecenoic acid | 6.88 | 0.38 | 0.00 | 329 | 7.85 |
| [M−H]− | PA 36:5|PA 18:2_18:3 | 1.84 | 0.37 | 0.00 | 693 | 12.14 |
| [M−H]− | PA 36:4|PA 18:2_18:2 | 1.35 | 0.53 | 0.00 | 695 | 12.25 |
| [M−H]− | 9,12,13-Trihydroxyoctadec-10-enoic acid | 1.82 | 0.37 | 0.00 | 329 | 7.53 |
| [M+HCOO]− | [2-Hydroxy-3-[3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxypropyl] octadeca-9,12-dienoate | 1.05 | 0.58 | 0.00 | 561 | 10.54 |
| [M−H]− | 9-Hexadecenoic acid | 1.59 | 0.26 | 0.00 | 253 | 11.15 |
| [M+HCOO]− | [2-Hydroxy-3-[3,4,5-trihydroxy-6-[[3,4,5-trihydroxy-6-(hydroxymethyl)oxan-2-yl]oxymethyl]oxan-2-yl]oxypropyl] octadeca-9,12,15-trienoate | 1.10 | 0.54 | 0.00 | 721 | 9.88 |
| [M−H]− | L-Malic acid | 2.60 | 0.49 | 0.00 | 133 | 0.74 |
| [M+H]+ | PE 34:2|PE 16:0_18:2 | 1.06 | 0.39 | 0.05 | 717 | 12.34 |
| [M+H]+ | Palmitic amide | 4.91 | 0.83 | 0.08 | 256 | 11.07 |
| [M+ACN+H]+ | 5-[(1S,4aS,8aS)-Decahydro-5,5,8a-trimethyl-2-methylene-1-naphthalenyl]-3-methyl-, (2E)-2-penten-1-ol | 1.25 | 0.84 | 0.09 | 332 | 11.67 |
| [M−H]− | PMeOH 34:3|PMeOH 16:0_18:3 | 1.75 | 0.55 | 0.06 | 683 | 12.27 |
| [M−H]− | PMeOH 34:2|PMeOH 16:0_18:2 | 3.68 | 0.65 | 0.09 | 685 | 12.39 |
| Adduct | Name | VIP | FC | p | m/z | rt (s) |
|---|---|---|---|---|---|---|
| [M+H]+ | MMPE 34:2|MMPE 16:0_18:2 | 1.27 | 1.98 | 0.00 | 731 | 12.40 |
| [M+H]+ | Tetradecanoylcarnitine | 2.09 | 0.63 | 0.00 | 372 | 10.99 |
| [M+H]+ | PC 36:4|PC 18:2_18:2 | 3.79 | 15.91 | 0.00 | 783 | 12.89 |
| [M+H]+ | 3-{7-[5-(1-hydroxyhenicosa-4,8-dien-1-yl)oxolan-2-yl]heptyl}-5-methyl-5H-furan-2-one | 1.40 | 1.58 | 0.00 | 573 | 12.15 |
| [M+NH4]+ | 4-[5-[[4-[5-[Acetyl(hydroxy)amino]pentylamino]-4-oxobutanoyl]-hydroxyamino]pentylamino]-4-oxobutanoic acid | 3.77 | 1.86 | 0.00 | 478 | 10.46 |
| [M+H-H2O]+ | 12,19,20-Trihydroxy-14-methylenegeranylnerol | 1.83 | 0.61 | 0.00 | 337 | 10.88 |
| [M+H]+ | 2-Aminooctadecane-1,3,4-triol | 1.35 | 1.37 | 0.00 | 318 | 8.50 |
| [M+H]+ | Cyclic AMP | 2.35 | 0.84 | 0.01 | 330 | 10.46 |
| [M+H-H2O]+ | DG(18:2(9Z,12Z)/18:2(9Z,12Z)) | 3.47 | 1.31 | 0.01 | 600 | 12.27 |
| [M+H]+ | PC 32:4|PC 14:1_18:3 | 1.09 | 1.25 | 0.04 | 727 | 12.16 |
| [M−H]− | PG 34:2|PG 16:0_18:2 | 1.47 | 2.71 | 0.00 | 746 | 12.29 |
| [M−CH3] − | Phosphatidylcholine lyso 18:2 | 1.25 | 1.98 | 0.00 | 504 | 10.43 |
| [M−H]− | PMeOH 34:2|PMeOH 16:0_18:2 | 5.73 | 1.45 | 0.00 | 685 | 12.39 |
| [M−H]− | 9-Hexadecenoic acid | 1.24 | 0.75 | 0.01 | 253 | 11.15 |
| [M−H]− | PMeOH 36:4|PMeOH 18:2_18:2 | 4.10 | 1.28 | 0.02 | 709 | 12.28 |
| [M−H]− | Linoleic acid | 3.95 | 0.82 | 0.02 | 279 | 11.30 |
| [M−H]− | Linolenic acid | 1.57 | 0.73 | 0.02 | 277 | 10.97 |
| [M−H]− | PE 36:4|PE 18:2_18:2 | 2.00 | 1.26 | 0.02 | 739 | 12.01 |
| [M+HCOO]− | (2-Hydroxy-3-octadeca-9,12-dienoyloxypropyl) 2-(trimethylazaniumyl)ethyl phosphate | 1.60 | 1.55 | 0.03 | 564 | 10.59 |
| [M−H]− | PI 34:2 | 3.49 | 1.48 | 0.04 | 834 | 12.27 |
| [M−H]− | 9-Hydroxy-10E,12Z-octadecadienoic acid | 1.84 | 0.79 | 0.04 | 295 | 9.76 |
| [M−H]- | PA 36:5|PA 18:2_18:3 | 1.35 | 1.26 | 0.04 | 693 | 12.14 |
| [M−H]− | PMeOH 34:3|PMeOH 16:0_18:3 | 1.38 | 1.27 | 0.04 | 683 | 12.27 |
| [M+H]+ | Erucamide | 7.83 | 1.14 | 0.05 | 338 | 12.07 |
| [M+H]+ | 3alpha,17beta-Dihydroxy-5alpha-androstane | 1.04 | 0.78 | 0.06 | 293 | 11.51 |
| [M+H]+ | N-(2-Phenylethyl)hexadecanamide | 2.66 | 0.93 | 0.10 | 360 | 12.07 |
| [M−H]− | [3-[2-Aminoethoxy(hydroxy)phosphoryl]oxy-2-hydroxypropyl] octadeca-9,12-dienoate | 2.18 | 1.50 | 0.08 | 476 | 10.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, C.; Yu, H.; Lu, L. Positive Effect of High Zinc on Growth of Sedum alfredii. Agriculture 2023, 13, 400. https://doi.org/10.3390/agriculture13020400
Xiao C, Yu H, Lu L. Positive Effect of High Zinc on Growth of Sedum alfredii. Agriculture. 2023; 13(2):400. https://doi.org/10.3390/agriculture13020400
Chicago/Turabian StyleXiao, Chun, Haiyue Yu, and Lingli Lu. 2023. "Positive Effect of High Zinc on Growth of Sedum alfredii" Agriculture 13, no. 2: 400. https://doi.org/10.3390/agriculture13020400