
Bioactivity of Essential Oil from Citrus aurantium Peel against the Pulse Beetle Callosbruchus maculatus F. on Chickpea

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and EO Extraction
2.2. Analysis of EO
2.3. Strain of Callosobruchus Maculatus
2.4. Biological Tests
2.5. Data Analysis
3. Results
3.1. Chemical Composition of EO
3.2. Effects of Citrus aurantium Peel EOs on the Survival of Callosobruchus maculatus
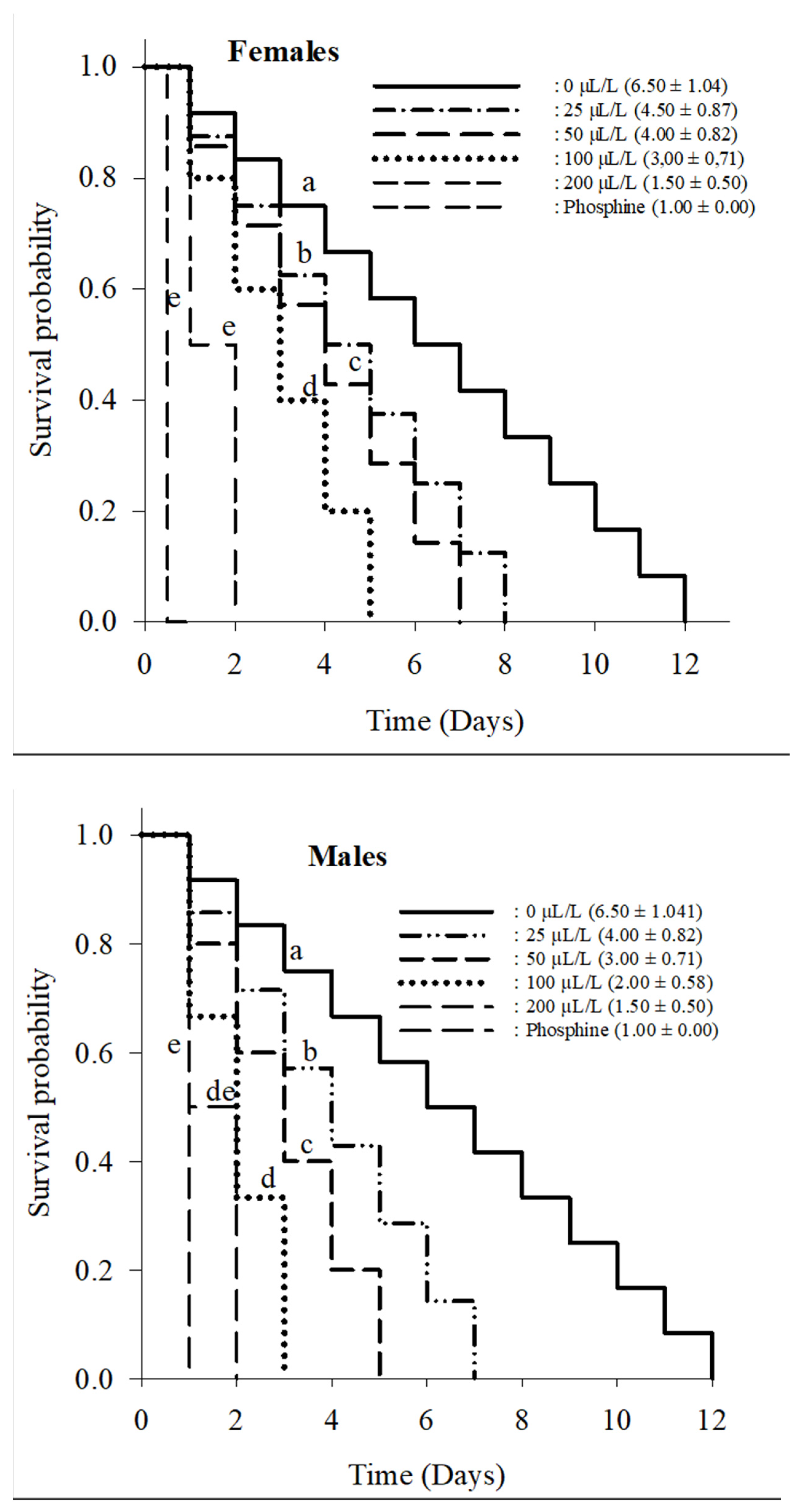
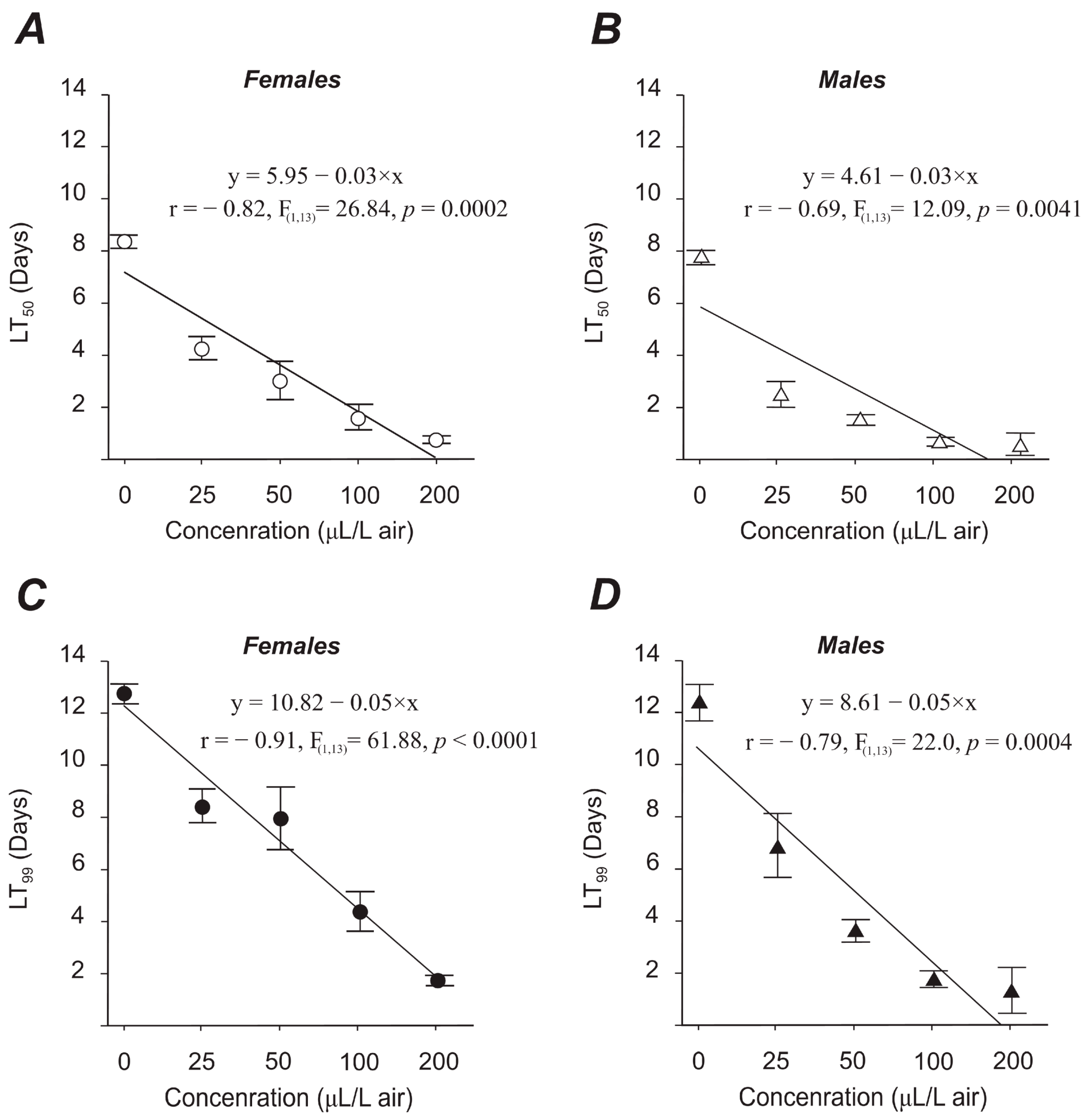
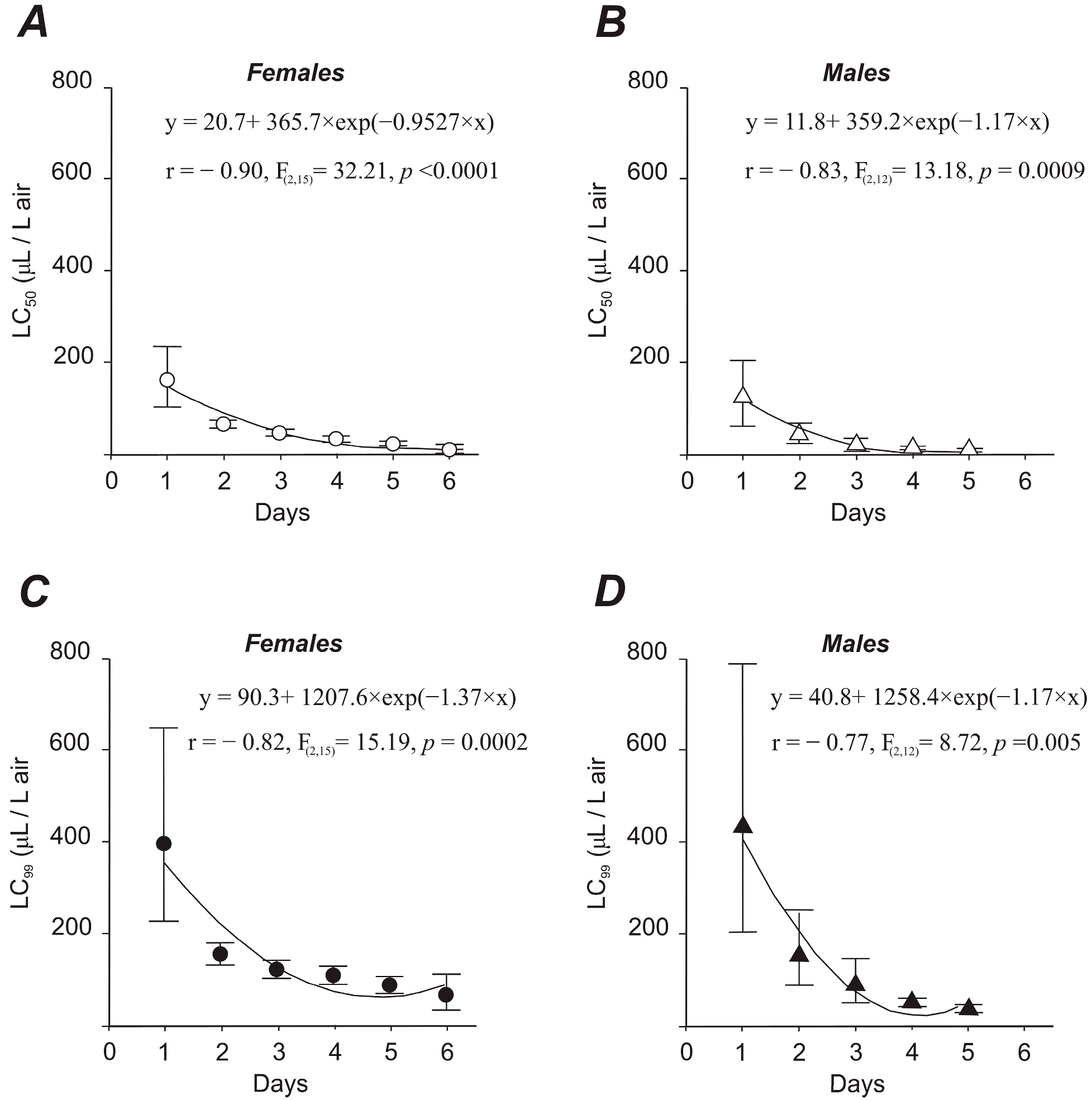
3.2.1. Adult Mortality
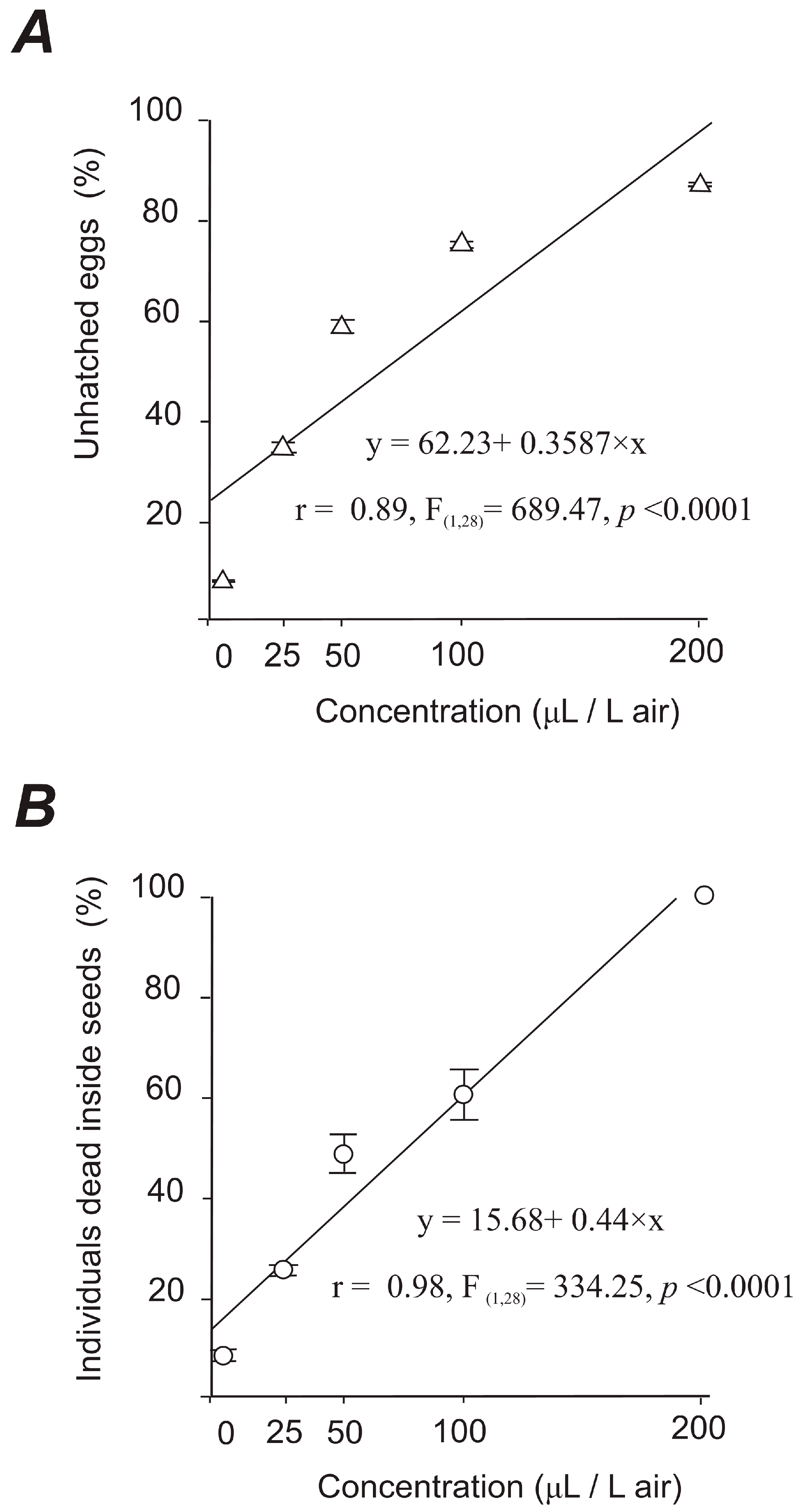
3.2.2. Non-Hatchability of Eggs and Mortality of Individuals Dead in Seeds
3.3. Effects of the Citrus aurantium Peel EO on Fecundity and Adult Emergence of Callosobruchus maculatus
3.3.1. Fecundity
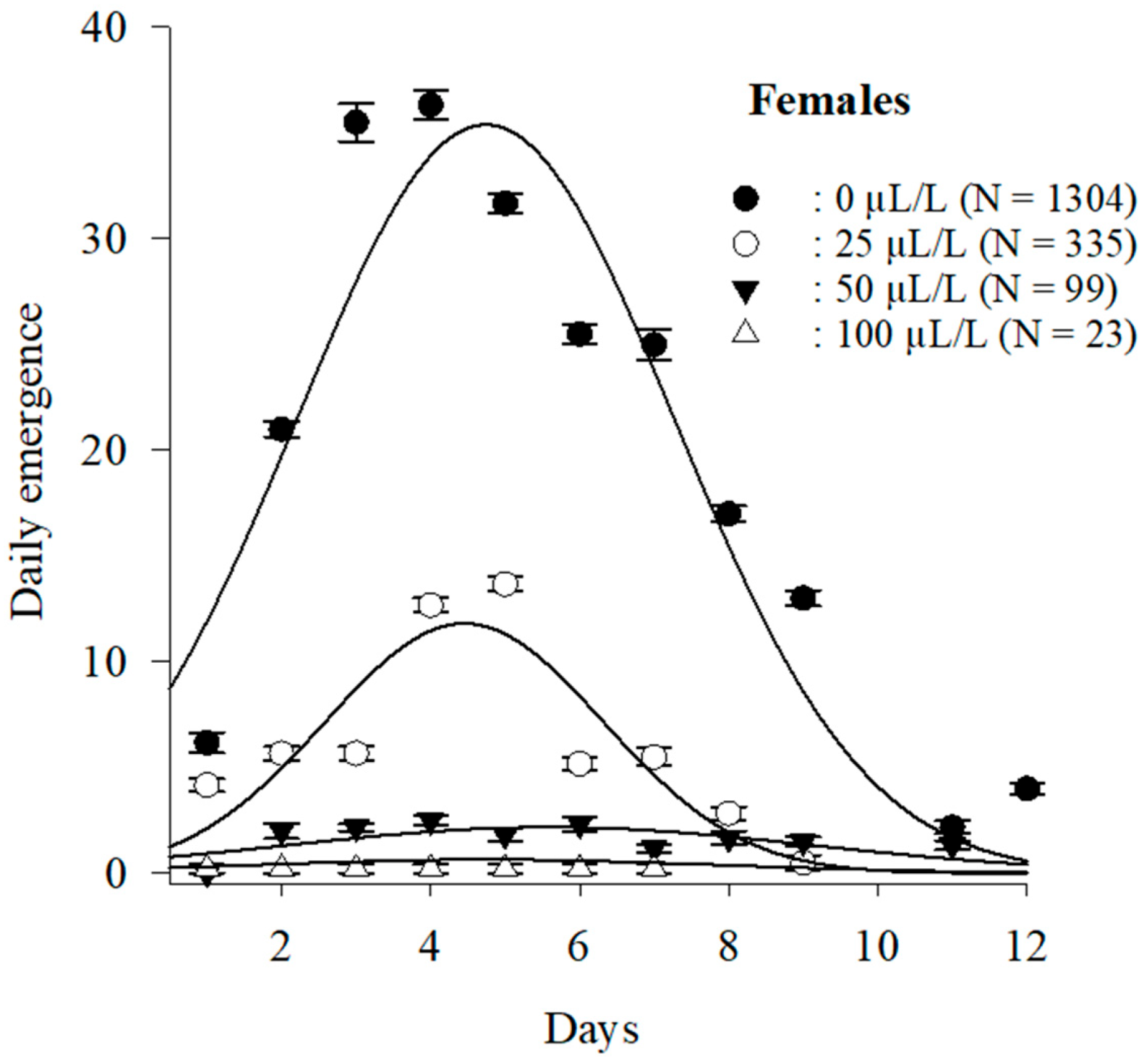
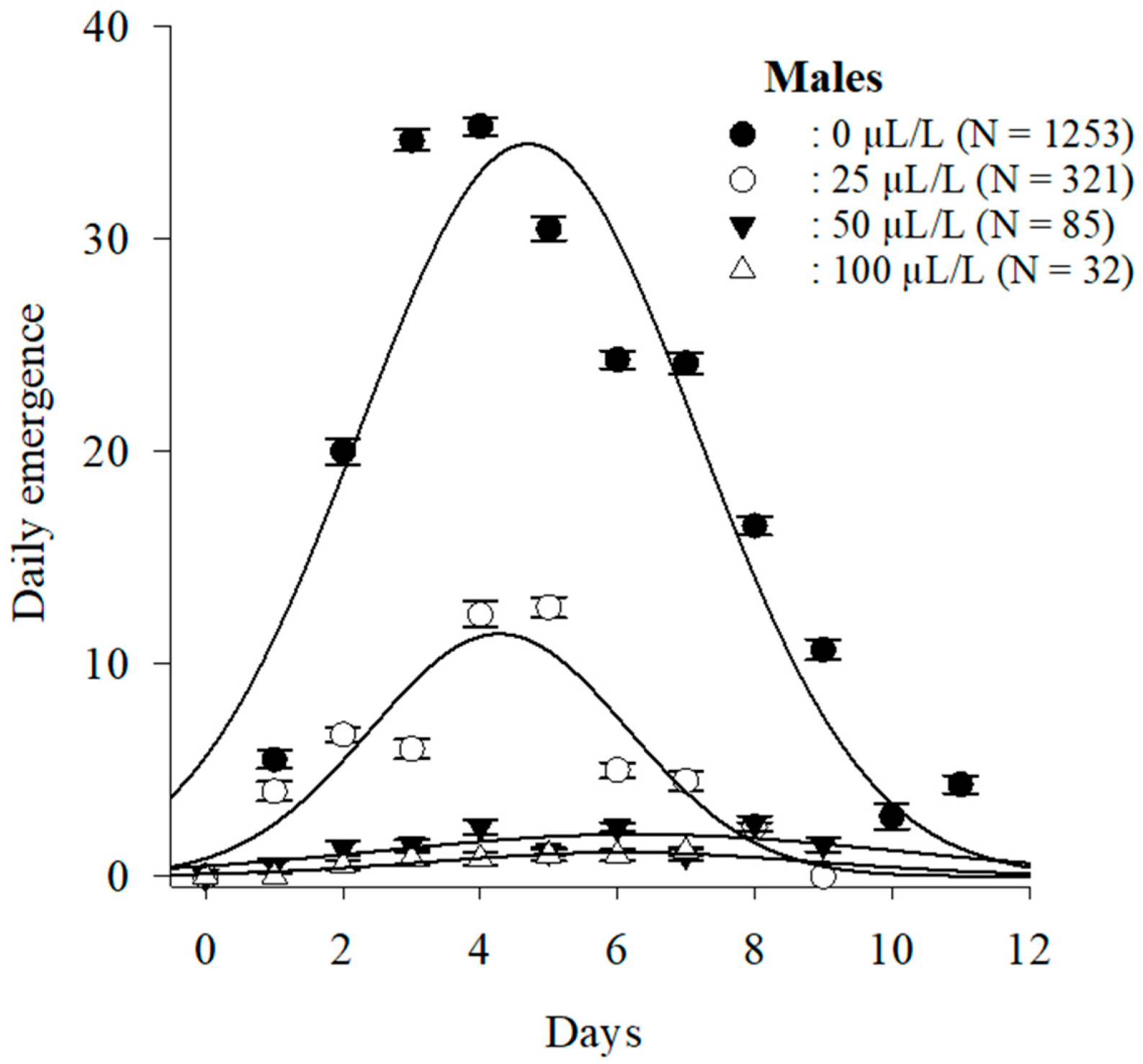
3.3.2. Adult Emergence
3.4. Effects of the Citrus aurantium Peel EO on the Population Growth Parameters of Callosobruchus maculatus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Isman, M.B. Botanical Insecticides in the Twenty-First Century—Fulfilling Their Promise? Annu. Rev. Entomol. 2020, 65, 233–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haddi, K.; Turchen, L.M.; Viteri Jumbo, L.O.; Guedes, R.N.C.; Pereira, E.J.G.; Aguiar, R.W.S.; Oliveira, E.E. Rethinking Biorational Insecticides for Pest Management: Unintended Effects and Consequences. Pest Manag. Sci. 2020, 76, 2286–2293. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Benelli, G. Essential Oils as Ecofriendly Biopesticides? Challenges and Constraints. Trends Plant Sci. 2016, 21, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Campolo, O.; Giunti, G.; Russo, A.; Palmeri, V.; Zappalà, L. Essential Oils in Stored Product Insect Pest Control. J. Food Qual. 2018, 2018, 6906105. [Google Scholar] [CrossRef] [Green Version]
- De Andrade Dutra, K.; de Oliveira, J.V.; Navarro, D.M.D.A.F.; Santos, J.P.O. Control of Callosobruchus maculatus (FABR.) (Coleoptera: Chrysomelidae: Bruchinae) in Vigna unguiculata (L.) WALP. with Essential Oils from Four Citrus spp. Plants. J. Stored Prod. Res. 2016, 68, 25–32. [Google Scholar] [CrossRef]
- Papanastasiou, S.A.; Bali, E.-M.D.; Ioannou, C.S.; Papachristos, D.P.; Zarpas, K.D.; Papadopoulos, N.T. Toxic and Hormetic-like Effects of Three Components of Citrus Essential Oils on Adult Mediterranean Fruit Flies (Ceratitis capitata). PLoS ONE 2017, 12, e0177837. [Google Scholar] [CrossRef] [Green Version]
- Chaieb, I.; Zarrad, K.; Sellam, R.; Tayeb, W.; Hammouda, A.B.; Laarif, A.; Bouhachem, S. Chemical Composition and Aphicidal Potential of Citrus aurantium Peel Essential Oils. Entomol. Gen. 2018, 37, 63–75. [Google Scholar] [CrossRef]
- Zarrad, K.; Hamouda, A.B.; Chaieb, I.; Laarif, A.; Jemâa, J.M.-B. Chemical Composition, Fumigant and Anti-Acetylcholinesterase Activity of the Tunisian Citrus aurantium L. Essential Oils. Ind. Crops Prod. 2015, 76, 121–127. [Google Scholar] [CrossRef]
- FAOSTAT. Cultures Et Produits Animaux. Available online: http://www.fao.org/faostat/fr/?#data/QC (accessed on 20 November 2022).
- Boughdad, A. Ravageurs Des Légumineuses Alimentaires Au Maroc. In Le Secteur des Légumineuses Alimentaires au Maroc; Actes (Eds) IAV Hasan II: Rabat, Morocco, 1992; pp. 315–338. [Google Scholar]
- De Melo, B.A.; Molina-Rugama, A.J.; Haddi, K.; Leite, D.T.; de Oliveira, E.E. Repellency and Bioactivity of Caatinga Biome Plant Powders against Callosobruchus maculatus (Coleoptera: Chrysomelidae: Bruchinae). Fla. Entomol. 2015, 98, 417–423. [Google Scholar] [CrossRef]
- Massango, H.G.L.L.; Faroni, L.R.A.; Haddi, K.; Heleno, F.F.; Viteri Jumbo, L.O.; Oliveira, E.E. Toxicity and Metabolic Mechanisms Underlying the Insecticidal Activity of Parsley Essential Oil on Bean Weevil, Callosobruchus maculatus. J. Pest Sci. 2017, 90, 723–733. [Google Scholar] [CrossRef]
- Haddi, K.; Viteri Jumbo, L.O.; Costa, M.S.; Santos, M.F.; Faroni, L.R.A.; Serrão, J.E.; Oliveira, E.E. Changes in the Insecticide Susceptibility and Physiological Trade-Offs Associated with a Host Change in the Bean Weevil Acanthoscelides obtectus. J. Pest Sci. 2018, 91, 459–468. [Google Scholar] [CrossRef]
- Nayak, M.K.; Daglish, G.J.; Phillips, T.W.; Ebert, P.R. Resistance to the Fumigant Phosphine and Its Management in Insect Pests of Stored Products: A Global Perspective. Annu. Rev. Entomol. 2020, 65, 333–350. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Palni, U.T.; Tripathi, N.N. Repellent Activity of Some Essential Oils against Two Stored Product Beetles Callosobruchus chinensis L. and C. maculatus F. (Coleoptera: Bruchidae) with Reference to Chenopodium ambrosioides L. Oil for the Safety of Pigeon Pea Seeds. J. Food Sci. Technol. 2014, 51, 4066–4071. [Google Scholar] [CrossRef] [Green Version]
- Nenaah, G.E.; Ibrahim, S.I.A.; Al-Assiuty, B.A. Chemical Composition, Insecticidal Activity and Persistence of Three Asteraceae Essential Oils and Their Nanoemulsions against Callosobruchus maculatus (F.). J. Stored Prod. Res. 2015, 61, 9–16. [Google Scholar] [CrossRef]
- Pourya, M.; Sadeghi, A.; Ghobari, H.; Taning, C.N.T.; Smagghe, G. Bioactivity of Pistacia atlantica Desf. Subsp. Kurdica (Zohary) Rech. F. and Pistacia Khinjuk Stocks Essential Oils against Callosobruchus maculatus (F, 1775) (Coloeptera: Bruchidae) under Laboratory Conditions. J. Stored Prod. Res. 2018, 77, 96–105. [Google Scholar] [CrossRef]
- de Souza Alves, M.; Campos, I.M.; de Brito, D.d.M.C.; Cardoso, C.M.; Pontes, E.G.; de Souza, M.A.A. Efficacy of Lemongrass Essential Oil and Citral in Controlling Callosobruchus maculatus (Coleoptera: Chrysomelidae), a Post-Harvest Cowpea Insect Pest. Crop Prot. 2019, 119, 191–196. [Google Scholar] [CrossRef]
- Chauhan, N.; Kashyap, U.; Dolma, S.K.; Reddy, S.G.E. Chemical Composition, Insecticidal, Persistence and Detoxification Enzyme Inhibition Activities of Essential Oil of Artemisia maritima against the Pulse Beetle. Molecules 2022, 27, 1547. [Google Scholar] [CrossRef]
- Van den Dool, H.; Kratz, P. A Generalization of the Retention Index System Including Linear Temperature Programmed Gas—Liquid Partition Chromatography. J. Chromatogr. A 1963, 11, 463–471. [Google Scholar] [CrossRef]
- Linstorm, P. NIST Chemistry Webbook, NIST Standard Reference Database Number 69. J. Phys. Chem. Ref. Data Monograph 1998, 9, 1–1951. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 9781932633214. [Google Scholar]
- ONSSA. Available online: http://eservice.onssa.gov.ma/IndPesticide.aspx (accessed on 20 November 2022).
- Robertson, J.L.; Jones, M.M.; Olguin, E.; Alberts, B. Bioassays with Arthropods; CRC Press: Boca Raton, FL, USA, 2017; ISBN 1315373777. [Google Scholar]
- González-Mas, M.C.; Rambla, J.L.; López-Gresa, M.P.; Blázquez, M.A.; Granell, A. Volatile Compounds in Citrus Essential Oils: A Comprehensive Review. Front. Plant Sci. 2019, 10, 12. [Google Scholar] [CrossRef]
- Bora, H.; Kamle, M.; Mahato, D.K.; Tiwari, P.; Kumar, P. Citrus Essential Oils (CEOs) and Their Applications in Food: An Overview. Plants 2020, 9, 357. [Google Scholar] [CrossRef] [PubMed]
- Boussaada, O.; Chemli, R. Seasonal Variation of Essential Oil Composition of Citrus aurantium L. Var. Amara. J. Essent. Oil Bear. Plants 2007, 10, 109–120. [Google Scholar] [CrossRef]
- Ferhat, M.A.; Meklati, B.Y.; Chemat, F. Comparison of Different Isolation Methods of Essential Oil from Citrus Fruits: Cold Pressing, Hydrodistillation and Microwave ‘Dry’ Distillation. Flavour. Fragr. J. 2007, 22, 494–504. [Google Scholar] [CrossRef]
- Moghaddam, M.; Mehdizadeh, L. Chapter 13—Chemistry of Essential Oils and Factors Influencing Their Constituents. In Handbook of Food Bioengineering; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 379–419. ISBN 978-0-12-811412-4. [Google Scholar]
- Razzaghi, S.E.; Arabhosseini, A.; Turk, M.; Soubrat, T.; Cendres, A.; Kianmehr, M.H.; Perino, S.; Chemat, F. Operational Efficiencies of Six Microwave Based Extraction Methods for Orange Peel Oil. J. Food Eng. 2019, 241, 26–32. [Google Scholar] [CrossRef]
- Oboh, G.; Ademosun, A.O.; Olumuyiwa, T.A.; Olasehinde, T.A.; Ademiluyi, A.O.; Adeyemo, A.C. Insecticidal Activity of Essential Oil from Orange Peels (Citrus sinensis) against Tribolium confusum, Callosobruchus maculatus and Sitophilus oryzae and Its Inhibitory Effects on Acetylcholinesterase and Na+/K+-ATPase Activiti. Phytoparasitica 2017, 45, 501–508. [Google Scholar] [CrossRef]
- Pugazhvendan, S.R.; Ross, P.R.; Elumalai, K. Insecticidal and Repellant Activities of Plants Oil against Stored Grain Pest, Tribolium Castaneum (Herbst) (Coleoptera: Tenebrionidae). Asian Pac. J. Trop Dis. 2012, 2, S412–S415. [Google Scholar] [CrossRef]
- Campolo, O.; Malacrinò, A.; Zappalà, L.; Laudani, F.; Chiera, E.; Serra, D.; Russo, M.; Palmeri, V. Fumigant Bioactivity of Five Citrus Essential Oils against Tribolium confusum. Phytoparasitica 2014, 42, 223–233. [Google Scholar] [CrossRef]
- Campolo, O.; Romeo, F.V.; Malacrinò, A.; Laudani, F.; Carpinteri, G.; Fabroni, S.; Rapisarda, P.; Palmeri, V. Effects of Inert Dusts Applied Alone and in Combination with Sweet Orange Essential Oil against Rhyzopertha dominica (Coleoptera: Bostrichidae) and Wheat Microbial Population. Ind. Crops Prod. 2014, 61, 361–369. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.M.; Mohamed, M.I.E.; Shawir, M.S.; Abou-Taleb, H.K. Chemical Composition, Insecticidal and Biochemical Effects of Essential Oils of Different Plant Species from Northern Egypt on the Rice Weevil, Sitophilus oryzae L. J. Pest Sci. 2016, 89, 219–229. [Google Scholar] [CrossRef]
- Oyedeji, A.O.; Okunowo, W.O.; Osuntoki, A.A.; Olabode, T.B.; Ayo-Folorunso, F. Insecticidal and Biochemical Activity of Essential Oil from Citrus Sinensis Peel and Constituents on Callosobrunchus maculatus and Sitophilus zeamais. Pestic. Biochem. Physiol. 2020, 168, 104643. [Google Scholar] [CrossRef]
- Raja, N.; Albert, S.; Ignacimuthu, S.; Dorn, S. Effect of Plant Volatile Oils in Protecting Stored Cowpea Vigna unguiculata (L.) Walpers against Callosobruchus maculatus (F.) (Coleoptera: Bruchidae) Infestation. J. Stored Prod. Res. 2001, 37, 127–132. [Google Scholar] [CrossRef]
- Nattudurai, G.; Baskar, K.; Paulraj, M.G.; Islam, V.I.H.; Ignacimuthu, S.; Duraipandiyan, V. Toxic Effect of Atalantia monophylla Essential Oil on Callosobruchus maculatus and Sitophilus oryzae. Environ. Sci. Pollut. Res. 2017, 24, 1619–1629. [Google Scholar] [CrossRef]
- Sahaf, B.Z.; Moharramipour, S. Fumigant Toxicity of Carum Copticum and Vitex Pseudo-Negundo Essential Oils against Eggs, Larvae and Adults of Callosobruchus maculatus. J. Pest Sci. 2008, 81, 213–220. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Rani, P.U.; Kavallieratos, N.G. The Use of Plant Extracts for Stored Product Protection BT—Advances in Plant Biopesticides; Singh, D., Ed.; Springer: New Delhi, India, 2014; pp. 131–147. ISBN 978-81-322-2006-0. [Google Scholar]
- Shukla, R.; Singh, P.; Prakash, B.; Dubey, N.K. Assessment of Essential Oil of Acorus calamus L. and Its Major Constituent β-Asarone in Post Harvest Management of Callosobruchus chinensis L. J. Essent. Oil Bear. Plants 2016, 19, 542–552. [Google Scholar] [CrossRef]
- Viteri Jumbo, L.O.; Haddi, K.; Faroni, L.R.D.; Heleno, F.F.; Pinto, F.G.; Oliveira, E.E. Toxicity to, Oviposition and Population Growth Impairments of Callosobruchus maculatus Exposed to Clove and Cinnamon Essential Oils. PLoS ONE 2018, 13, e0207618. [Google Scholar] [CrossRef]
- Matos, L.F.; da Cruz Lima, E.; de Andrade Dutra, K.; Navarro, D.M.D.A.F.; Alves JL, R.; Silva, G.N. Chemical Composition and Insecticidal Effect of Essential Oils from Illicium verum and Eugenia caryophyllus on Callosobruchus maculatus in Cowpea. Ind. Crops Prod. 2020, 145, 112088. [Google Scholar] [CrossRef]
- Papachristos, D.P.; Stamopoulos, D.C. Repellent, Toxic and Reproduction Inhibitory Effects of Essential Oil Vapours on Acanthoscelides obtectus (Say) (Coleoptera: Bruchidae). J. Stored Prod. Res. 2002, 38, 117–128. [Google Scholar] [CrossRef]
- Fatiha, R.A.; Kada, R.; Khelil, M.A.; Pujade-Villar, J. Biological Control against the Cowpea Weevil (Callosobruchus chinensis L., Coleoptera: Bruchidae) Using Essential Oils of Some Medicinal Plants. J. Plant Prot. Res. 2014, 54, 211–217. [Google Scholar] [CrossRef]
- Keane, S.; Ryan, M.F. Purification, Characterisation, and Inhibition by Monoterpenes of Acetylcholinesterase from the Waxmoth, Galleria mellonella (L.). Insect. Biochem. Mol. Biol. 1999, 29, 1097–1104. [Google Scholar] [CrossRef]
- López, M.D.; Pascual-Villalobos, M.J. Mode of Inhibition of Acetylcholinesterase by Monoterpenoids and Implications for Pest Control. Ind. Crops Prod. 2010, 31, 284–288. [Google Scholar] [CrossRef]
- Kim, S.-W.; Kang, J.; Park, I.-K. Fumigant Toxicity of Apiaceae Essential Oils and Their Constituents against Sitophilus oryzae and Their Acetylcholinesterase Inhibitory Activity. J. Asia Pac. Entomol. 2013, 16, 443–448. [Google Scholar] [CrossRef]
- Abdelgaleil, S.A.M.; Mohamed, M.I.E.; Badawy, M.E.I.; El-Arami, S.A.A. Fumigant and Contact Toxicities of Monoterpenes to Sitophilus oryzae (L.) and Tribolium castaneum (Herbst) and Their Inhibitory Effects on Acetylcholinesterase Activity. J. Chem. Ecol. 2009, 35, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Priestley, C.M.; Williamson, E.M.; Wafford, K.A.; Sattelle, D.B. Thymol, a Constituent of Thyme Essential Oil, Is a Positive Allosteric Modulator of Human GABAA Receptors and a Homo-Oligomeric GABA Receptor from Drosophila melanogaster. Br. J. Pharmacol. 2003, 140, 1363–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloomquist, J.R.; Boina, D.R.; Chow, E.; Carlier, P.R.; Reina, M.; Gonzalez-Coloma, A. Mode of Action of the Plant-Derived Silphinenes on Insect and Mammalian GABAA Receptor/Chloride Channel Complex. Pestic. Biochem. Physiol. 2008, 91, 17–23. [Google Scholar] [CrossRef]
- Rattan, R.S. Mechanism of Action of Insecticidal Secondary Metabolites of Plant Origin. Crop Prot. 2010, 29, 913–920. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of Monoterpenoid Insecticides on [3H]-TBOB Binding in House Fly GABA Receptor and 36Cl—Uptake in American Cockroach Ventral Nerve Cord. Pestic. Biochem Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Quantitative Structure–Activity Relationships of Monoterpenoid Binding Activities to the Housefly GABA Receptor. Pest Manag. Sci. 2012, 68, 1122–1129. [Google Scholar] [CrossRef] [Green Version]
- Li, A.S.; Iijima, A.; Huang, J.; Li, Q.X.; Chen, Y. Putative Mode of Action of the Monoterpenoids Linalool, Methyl Eugenol, Estragole, and Citronellal on Ligand-Gated Ion Channels. Engineering 2020, 6, 541–545. [Google Scholar] [CrossRef]
- Enan, E.E. Molecular and Pharmacological Analysis of an Octopamine Receptor from American Cockroach and Fruit Fly in Response to Plant Essential Oils. Arch Insect. Biochem. Physiol. 2005, 59, 161–171. [Google Scholar] [CrossRef]
- Enan, E.E. Molecular Response of Drosophila melanogaster Tyramine Receptor Cascade to Plant Essential Oils. Insect Biochem. Mol. Biol. 2005, 35, 309–321. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of Octopaminergic Receptors by Essential Oil Constituents Isolated from Aromatic Plants: Possible Mode of Action against Insect Pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef]
- Price, D.N.; Berry, M.S. Comparison of Effects of Octopamine and Insecticidal Essential Oils on Activity in the Nerve Cord, Foregut, and Dorsal Unpaired Median Neurons of Cockroaches. J. Insect. Physiol. 2006, 52, 309–319. [Google Scholar] [CrossRef] [PubMed]
- De Souza, L.; Cardoso, M.D.; Konig, I.F.; Ferreira, V.R.; Caetano, A.R.; Campolina, G.A.; Haddi, K. Toxicity, Histopathological Alterations and Acetylcholinesterase Inhibition of Illicium verum Essential Oil in Drosophila Suzukis. Agriculture 2022, 12, 1667. [Google Scholar] [CrossRef]
- Ciriminna, R.; Lomeli-Rodriguez, M.; Demma Carà, P.; Lopez-Sanchez, J.A.; Pagliaro, M. Limonene: A Versatile Chemical of the Bioeconomy. Chem. Commun. 2014, 50, 15288–15296. [Google Scholar] [CrossRef] [PubMed]
- Malacrinò, A.; Campolo, O.; Laudani, F.; Palmeri, V. Fumigant and Repellent Activity of Limonene Enantiomers Against Tribolium confusum Du Val. Neotrop. Entomol. 2016, 45, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Huang, S.; Li, C.; Chen, S.; Peng, Z. Biological Activity of the Essential Oil from the Leaves of Piper Sarmentosum Roxb. (Piperaceae) and Its Chemical Constituents on Brontispa longissima (Gestro) (Coleoptera: Hispidae). Pestic. Biochem. Physiol. 2010, 96, 132–139. [Google Scholar] [CrossRef]
- Rossi, Y.E.; Palacios, S.M. Fumigant Toxicity of Citrus Sinensis Essential Oil on Musca domestica L. Adults in the Absence and Presence of a P450 Inhibitor. Acta Trop 2013, 127, 33–37. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Jalali Sendi, J. A Review on Recent Research Results on Bio-Effects of Plant Essential Oils against Major Coleopteran Insect Pests. Toxin Rev. 2015, 34, 76–91. [Google Scholar] [CrossRef]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological Effects of Essential Oils—A Review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Koul, O.; Walia, S.; Dhaliwal, G.S.; Nagar, P. Essential Oils as Green Pesticides Potential and Constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N° | Compounds * | RI ** | RT *** | Formula | Content (%) |
|---|---|---|---|---|---|
| 1 | Octan-1-ol | 925 | 5.89 | C8H18O | 0.63 |
| 2 | α-Pinene | 930 | 8.19 | C10H16 | 1.01 |
| 3 | Camphene | 944 | 8.65 | C10H16 | 0.03 |
| 4 | Sabinene | 971 | 9.62 | C10H16 | 0.05 |
| 5 | Limonene | 1006 | 11.51 | C10H16 | 86.18 |
| 7 | α-Terpinene | 1021 | 15.61 | C10H16 | 0.06 |
| 8 | 7-Propylidene-bicyclo [4.1.0] heptane | 1025 | 16.41 | C10H16 | 0.08 |
| 6 | Trans-Limonene-1.2 oxide | 1056 | 22.27 | C10H16O | 0.06 |
| 9 | Terpinen-4-ol | 1083 | 24.87 | C10H18O | 0.08 |
| 10 | 3-Carene | 1099 | 27.63 | C10H16 | 0.08 |
| 11 | Ocimene | 1138 | 35.58 | C10H16 | 0.07 |
| 12 | Cyclobutane. 1.2-bis (1-methyle thenyl)-. trans- | 1162 | 41.78 | C10H16 | 0.03 |
| 13 | Thymol | 1174 | 42.18 | C10H14O | 0.03 |
| 14 | Terpinen-1-ol | 1189 | 42.45 | C10H18O | 3.81 |
| 15 | Linalool | 1196 | 42.59 | C10H18O | 1.73 |
| 16 | Undecanal | 1236 | 43.34 | C11H22O | 4.57 |
| 17 | Geranyl acetate | 1255 | 43.52 | C12H2002 | 0.07 |
| 18 | Neryl acetate | 1265 | 43.59 | C12H2002 | 0.08 |
| 19 | δ-Elemene | 1288 | 43.72 | C15H24 | 0.08 |
| 20 | CaryophylleneE | 1335 | 44.57 | C15H24 | 0.07 |
| 21 | Geracrene-A or β-Bisabolene | 1347 | 44.68 | C15H24 | 0.09 |
| 22 | Quercetin-7,3′,4′-trimethyl ether | 1368 | 44.81 | C18H16O7 | 0.06 |
| Monoterpenes hydrocarbon | 87.59 | ||||
| Oxygenated monoterpenes | 5.71 | ||||
| Sesquiterpenes hydrocarbon | 0.24 | ||||
| Other | 5.41 | ||||
| Total | 98.95 | ||||
| Yield of EO (%) | 1.25 ± 0.1 | ||||
| Gender | Model | R² | F | df | p |
|---|---|---|---|---|---|
| Females | y = −6.24 + 8.88T + 0.13C +0.14TC | 0.92 | 89.08 | 3, 22 | <0.0001 |
| T | 0.66 | 31.93 | 1, 22 | <0.0001 | |
| C | 0.54 | 4.53 | 1, 22 | 0.0448 | |
| T × C | 0.62 | 29.26 | 1, 22 | <0.0001 | |
| Males | y = 10.62 + 2.98T + −0.05C + 0.47TC | 0.95 | 94.95 | 3, 14 | <0.0001 |
| T | 0.69 | 171.87 | 1, 14 | <0.0001 | |
| C | 0.46 | 113.40 | 1, 14 | <0.0001 | |
| T × C | 0.59 | 90.30 | 1, 14 | <0.0001 |
| Concentration (µL/L Air) | Eggs/10 Pairs | Adult Numbers/10 Pairs |
|---|---|---|
| 0 | 507.83 ± 12.66 a* | 426.17 ± 5.62 a |
| 25 | 224.67 ± 4.61 b | 109.33 ± 4.57 b |
| 50 | 150.83 ± 3.36 c | 30.50 ± 1.38 c |
| 100 | 94.83 ± 3.24 d | 9.17 ± 1.14 d |
| 200 | 53.50 ± 2.29 e | 0.00 ± 0.00 e |
| F | 782.23 | 2879.43 |
| df | 4, 25 | 4, 25 |
| p | <0.0001 | <0.0001 |
| Concentration * (µL/L Air) | T (Days) | Sex Ratio | R0 (Female/Female) | r (Female/Day) | λ E (Female/Day) | DT (Days) |
|---|---|---|---|---|---|---|
| 0 | 28.17 ± 0.82 a** | 0.97 ± 0.03 a | 21.70 ± 0.58 a | 0.110 ± 0.004 a | 1.12 ± 0.004 a | 6.36 ± 0.25 a |
| 25 | 27.01 ± 0.78 a | 0.97 ± 0.03 a | 5.55 ± 0.24 b | 0.064 ± 0.003 b | 1.07 ± 0.004 b | 11.02 ± 0.57 b |
| 50 | 28.39 ± 0.77 a | 0.87 ± 0.04 a | 1.63 ± 0.08 c | 0.017 ± 0.002 c | 1.02 ± 0.002 c | 42.52 ± 4.17 c |
| 100 | 28.54 ± 1.10 a | 1.42 ± 0.08 b | 0.38 ± 0.05 d | −0.04 ± 0.01 d | 0.96 ± 0.01 d | −22.70 ± 4.86 d*** |
| F | 0.41 | 26.66 | 1904.51 | 272.91 | 245.00 | 68.94 |
| df | 3, 20 | 3, 20 | 3, 20 | 3, 20 | 3, 20 | 3, 20 |
| p | 0.747 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Kasimi, R.; Douiri, F.; Haddi, K.; Boughdad, A. Bioactivity of Essential Oil from Citrus aurantium Peel against the Pulse Beetle Callosbruchus maculatus F. on Chickpea. Agriculture 2023, 13, 232. https://doi.org/10.3390/agriculture13020232
El Kasimi R, Douiri F, Haddi K, Boughdad A. Bioactivity of Essential Oil from Citrus aurantium Peel against the Pulse Beetle Callosbruchus maculatus F. on Chickpea. Agriculture. 2023; 13(2):232. https://doi.org/10.3390/agriculture13020232
Chicago/Turabian StyleEl Kasimi, Rachid, Fatima Douiri, Khalid Haddi, and Ahmed Boughdad. 2023. "Bioactivity of Essential Oil from Citrus aurantium Peel against the Pulse Beetle Callosbruchus maculatus F. on Chickpea" Agriculture 13, no. 2: 232. https://doi.org/10.3390/agriculture13020232