
Deletion of the OsLA1 Gene Leads to Multi-Tillering and Lazy Phenotypes in Rice
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Planting and Investigation of Agronomic Traits
2.2. Construction and Genetic Analysis of the F2 Segregated Population
2.3. Mixed-Pool Sequencing and Single-Nucleotide Polymorphism Analysis
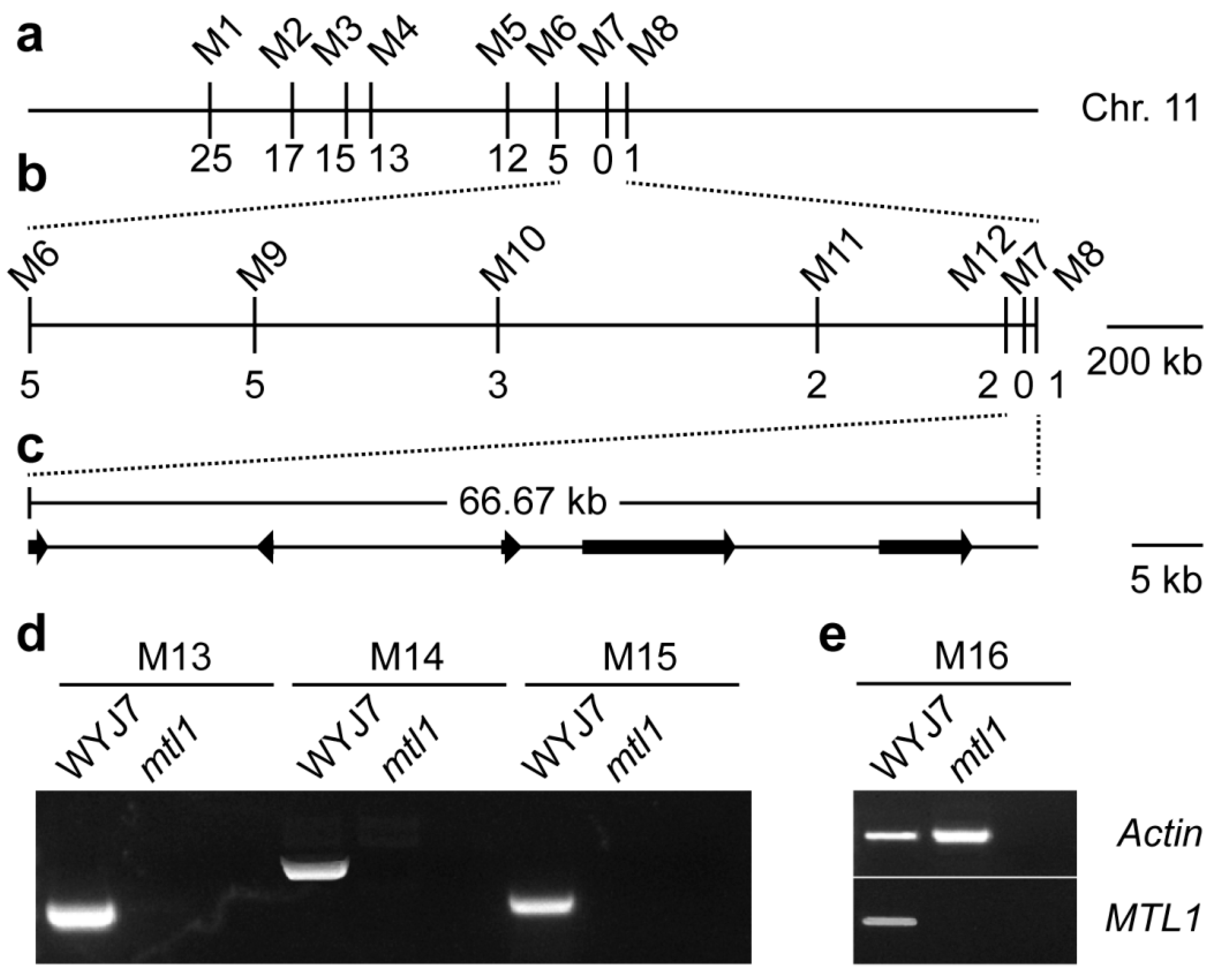
2.4. Fine Mapping of the MTL1 Gene
2.5. Analysis of the Candidate Genes
3. Results
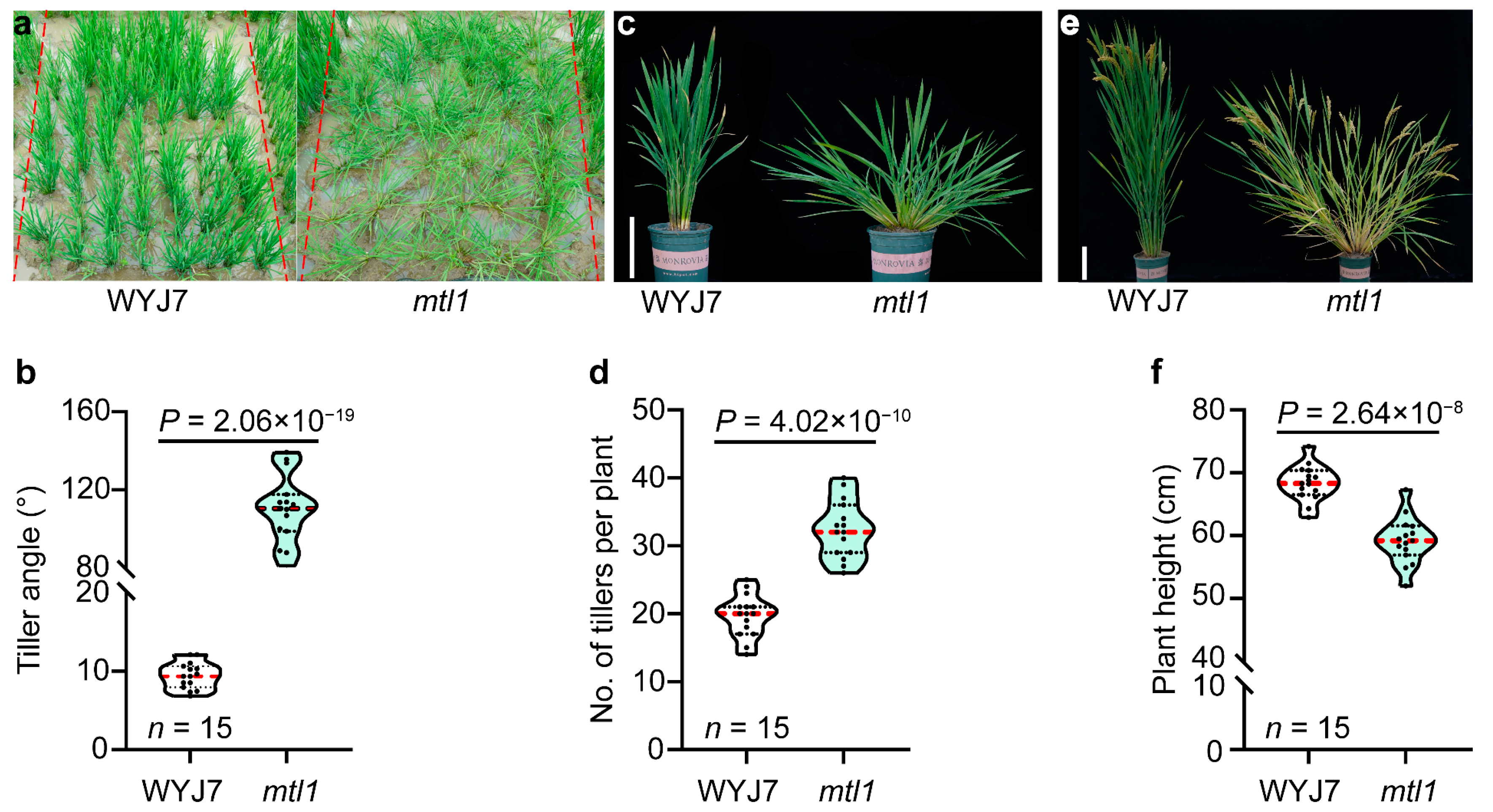
3.1. Phenotypic Analysis of the mtl1 Mutant
3.1.1. Plant Architecture Analysis of mtl1
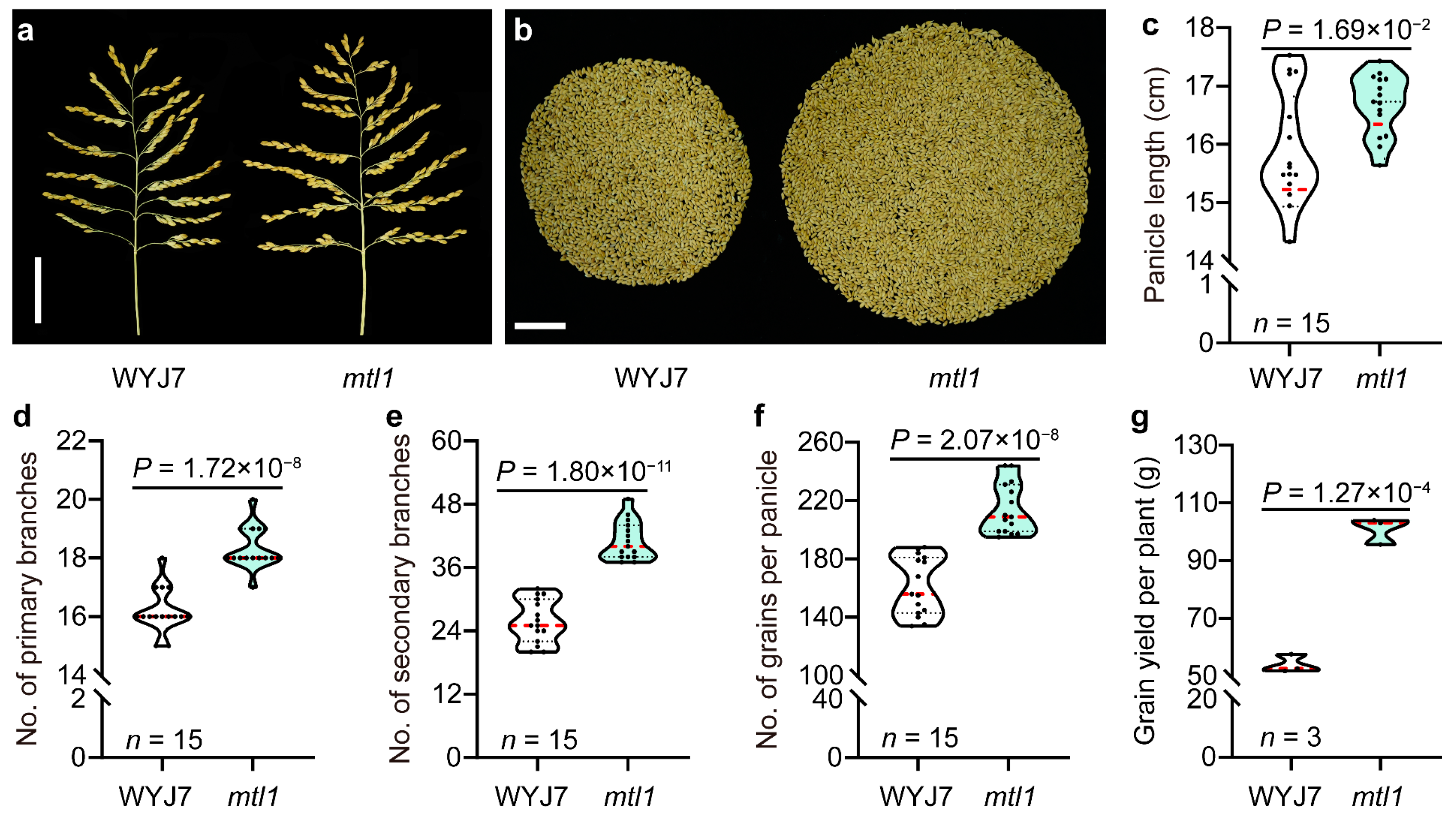
3.1.2. Panicle Type Analysis of mtl1
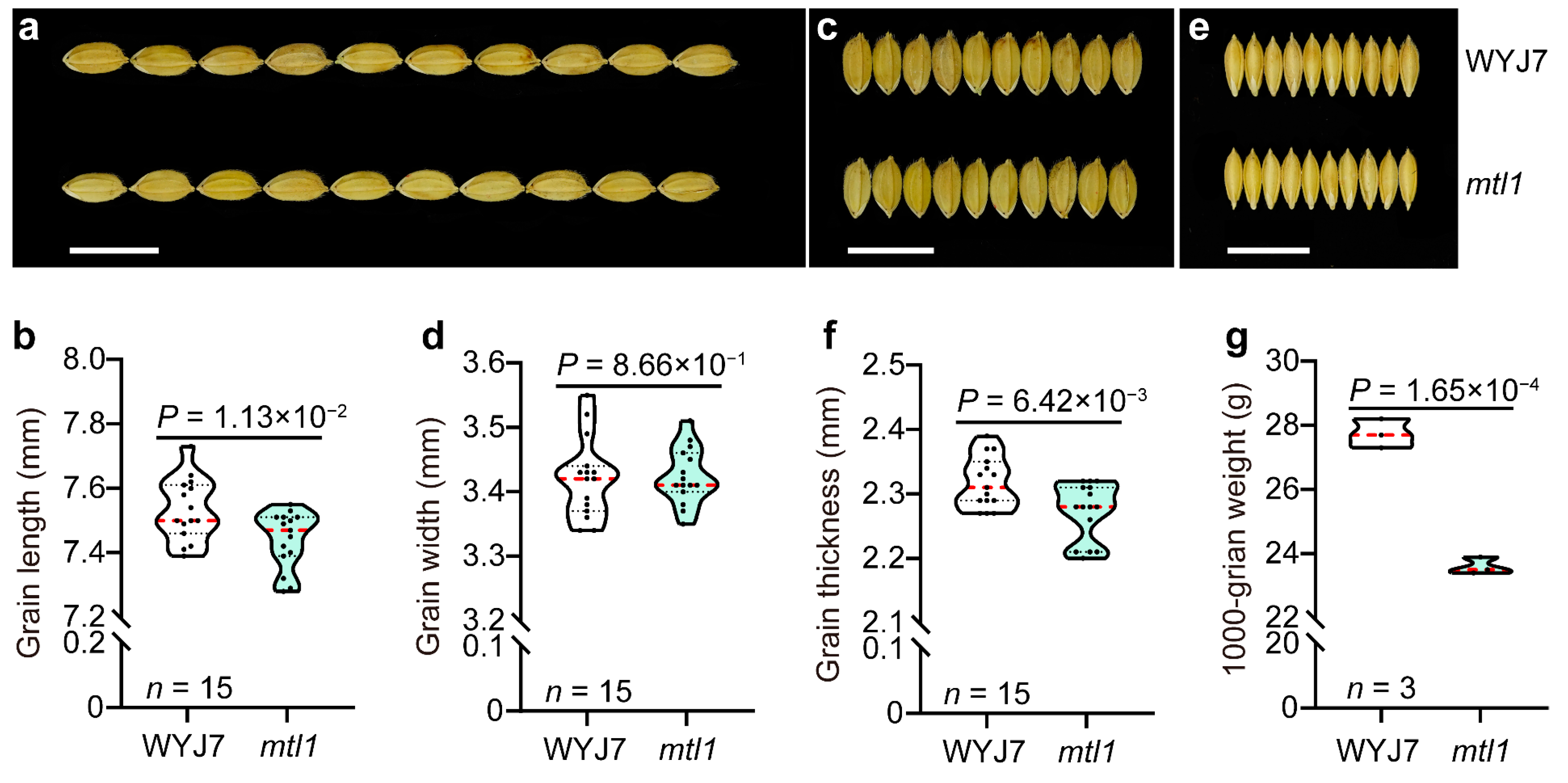
3.1.3. Grain Type Analysis of mtl1
3.2. Genetic Analysis of the mtl1 Mutant Phenotypes in Rice
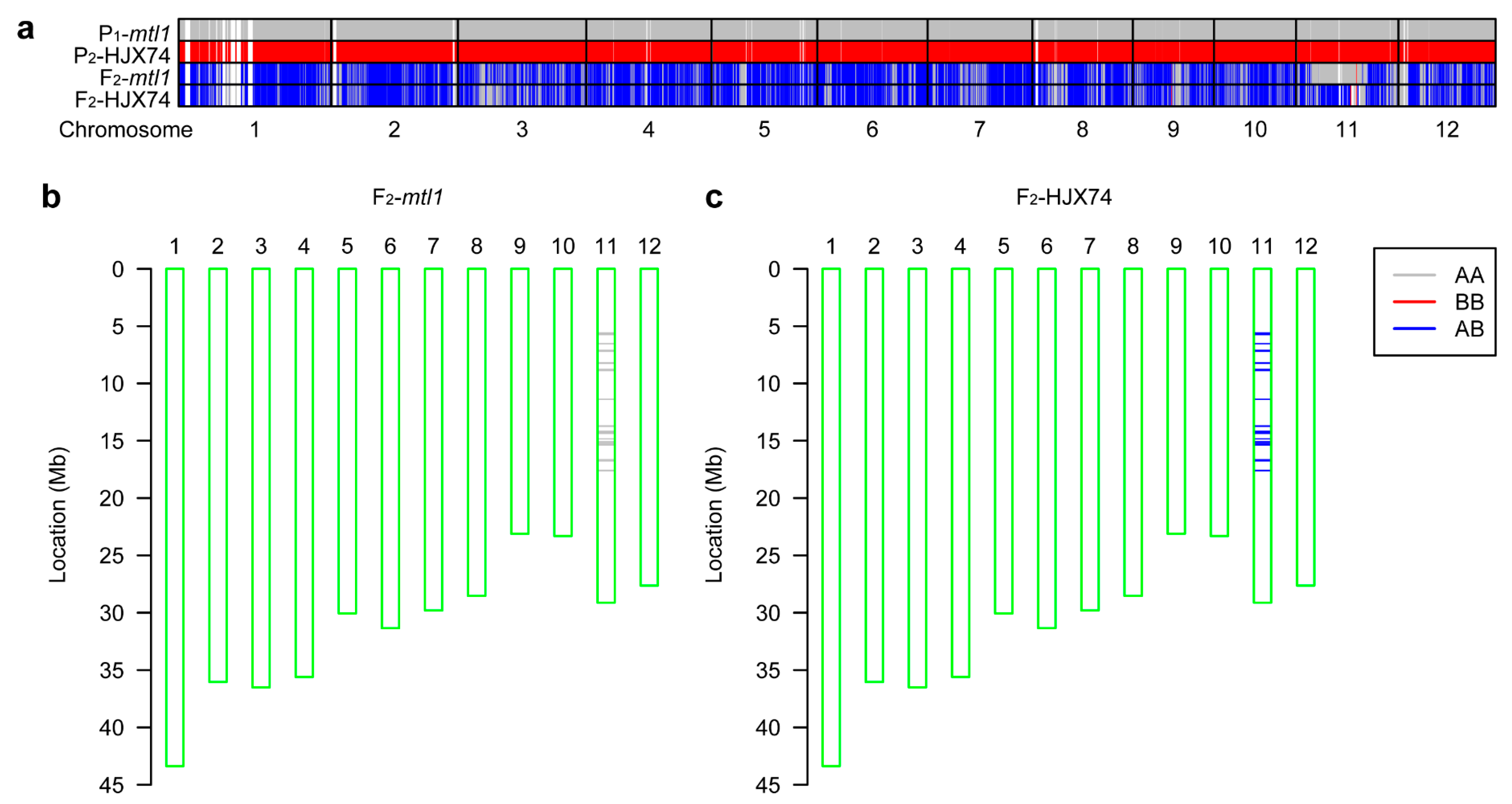
3.3. Preliminary Mapping of the MTL1 Gene
3.4. Fine Mapping of the MTL1 Gene
3.5. Analysis of the Candidate Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jiao, Y.Q.; Wang, Y.H.; Xue, D.W.; Wang, J.; Yan, M.X.; Liu, G.F.; Dong, G.J.; Zeng, D.L.; Lu, Z.F.; Zhu, X.D.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar] [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef]
- Qian, Q.; Guo, L.B.; Smith, S.M.; Li, J.Y. Breeding high-yield superior quality hybrid super rice by rational design. Natl. Sci. Rev. 2016, 3, 283–294. [Google Scholar] [CrossRef]
- Wang, B.; Smith, S.M.; Li, J.Y. Genetic Regulation of Shoot Architecture. Annu. Rev. Plant Biol. 2018, 69, 437–468. [Google Scholar] [CrossRef]
- Schumacher, K.; Schmitt, T.; Rossberg, M.; Schmitz, G.; Theres, K. The Lateral suppressor (Ls) gene of tomato encodes a new member of the VHIID protein family. Proc. Natl. Acad. Sci. USA 1999, 96, 290–295. [Google Scholar] [CrossRef]
- Greb, T.; Clarenz, O.; Schafer, E.; Muller, D.; Herrero, R.; Schmitz, G.; Theres, K. Molecular analysis of the LATERAL SUPPRESSOR gene in Arabidopsis reveals a conserved control mechanism for axillary meristem formation. Genes Dev. 2003, 17, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Qian, Q.; Fu, Z.M.; Wang, Y.H.; Xiong, G.S.; Zeng, D.L.; Wang, X.Q.; Liu, X.F.; Teng, S.; Hiroshi, F.; et al. Control of tillering in rice. Nature 2003, 422, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Koumoto, T.; Shimada, H.; Kusano, H.; She, K.C.; Iwamoto, M.; Tanako, M. Rice monoculm mutation moc2, which inhibits outgrowth of the second tillers, is ascribed to lack of a fructose-1,6-bisphosphatase. Plant Biotechnol. J. 2013, 30, 47–56. [Google Scholar] [CrossRef]
- Lu, Z.F.; Shao, G.N.; Xiong, J.S.; Jiao, Y.Q.; Wang, J.; Liu, G.F.; Meng, X.B.; Liang, Y.; Xiong, G.S.; Wang, Y.H.; et al. MONOCULM 3, an ortholog of WUSCHEL in rice, is required for tiller bud formation. J. Genet. Genomics 2015, 42, 71–78. [Google Scholar] [CrossRef]
- Tanaka, W.; Ohmori, Y.; Ushijima, T.; Matsusaka, H.; Matsushita, T.; Kumamaru, T.; Kawano, S.; Hirano, H.Y. Axillary Meristem Formation in Rice Requires the WUSCHEL Ortholog TILLERS ABSENT1. Plant Cell 2015, 27, 1173–1184. [Google Scholar] [CrossRef]
- Xia, T.Y.; Chen, H.Q.; Dong, S.J.; Ma, Z.Y.; Ren, H.B.; Zhu, X.D.; Fang, X.H.; Chen, F. OsWUS promotes tiller bud growth by establishing weak apical dominance in rice. Plant J. 2020, 104, 1635–1647. [Google Scholar] [CrossRef]
- Ishikawa, S.; Maekawa, M.; Arite, T.; Onishi, K.; Takamure, I.; Kyozuka, J. Suppression of tiller bud activity in tillering dwarf mutants of rice. Plant Cell Physiol. 2005, 46, 79–86. [Google Scholar] [CrossRef]
- Lin, H.; Wang, R.X.; Qian, Q.; Yan, M.X.; Meng, X.B.; Fu, Z.M.; Yan, C.Y.; Jiang, B.; Su, Z.; Li, J.Y.; et al. DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell 2009, 21, 1512–1525. [Google Scholar] [CrossRef]
- Ito, S.; Kitahata, N.; Umehara, M.; Hanada, A.; Kato, A.; Ueno, K.; Mashiguchi, K.; Kyozuka, J.; Yoneyama, K.; Yamaguchi, S.; et al. A new lead chemical for strigolactone biosynthesis inhibitors. Plant Cell Physiol. 2010, 51, 1143–1150. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, X.; Xiong, G.S.; Liu, H.H.; Chen, F.L.; Wang, L.; Meng, X.B.; Liu, G.F.; Yu, H.; Yuan, Y.D.; et al. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 2013, 504, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Kagiyama, M.; Hirano, Y.; Mori, T.; Kim, S.Y.; Kyozuka, J.; Seto, Y.; Yamaguchi, S.; Hakoshima, T. Structures of D14 and D14L in the strigolactone and karrikin signaling pathways. Genes Cells 2013, 18, 147–160. [Google Scholar] [CrossRef]
- Jin, J.; Huang, W.; Gao, J.P.; Yang, J.; Shi, M.; Zhu, M.Z.; Luo, D.; Lin, H.X. Genetic control of rice plant architecture under domestication. Nat. Genet. 2008, 40, 1365–1369. [Google Scholar] [CrossRef]
- Tan, L.B.; Li, X.R.; Liu, F.X.; Sun, X.Y.; Li, C.G.; Zhu, Z.F.; Fu, Y.C.; Cai, H.W.; Wang, X.K.; Xie, D.X.; et al. Control of a key transition from prostrate to erect growth in rice domestication. Nat. Genet. 2008, 40, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.S.; Lin, Z.W.; Li, H.X.; Li, X.J.; Li, J.Y.; Wang, Y.H.; Zhang, X.; Zhu, Z.F.; Zhai, W.X.; Wang, X.K.; et al. TAC1, a major quantitative trait locus controlling tiller angle in rice. Plant J. 2007, 52, 891–898. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.J.; Zhao, H.; Xie, W.B.; Han, Z.M.; Li, G.W.; Yao, W.; Bai, X.F.; Hu, Y.; Guo, Z.L.; Lu, K.; et al. A Novel Tiller Angle Gene, TAC3, together with TAC1 and D2 Largely Determine the Natural Variation of Tiller Angle in Rice Cultivars. PLoS Genet. 2016, 12, e1006412. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sun, H.Y.; Jiang, J.H.; Sun, X.Y.; Tan, L.B.; Sun, C.Q. TAC4 controls tiller angle by regulating the endogenous auxin content and distribution in rice. Plant Biotechnol. J. 2021, 19, 64–73. [Google Scholar] [CrossRef]
- Li, P.J.; Wang, Y.H.; Qian, Q.; Fu, Z.M.; Wang, M.; Zeng, D.L.; Li, B.H.; Wang, X.J.; Li, J.Y. LAZY1 controls rice shoot gravitropism through regulating polar auxin transport. Cell Res. 2007, 17, 402–410. [Google Scholar] [CrossRef]
- Yoshihara, T.; Iino, M. Identification of the gravitropism-related rice gene LAZY1 and elucidation of LAZY1-dependent and -independent gravity signaling pathways. Plant Cell Physiol. 2007, 48, 678–688. [Google Scholar] [CrossRef] [PubMed]
- Sang, D.J.; Chen, D.Q.; Liu, G.F.; Liang, Y.; Huang, L.Z.; Meng, X.B.; Chu, J.F.; Sun, X.H.; Dong, G.J.; Yuan, Y.D.; et al. Strigolactones regulate rice tiller angle by attenuating shoot gravitropism through inhibiting auxin biosynthesis. Proc. Natl. Acad. Sci. USA 2014, 111, 11199–11204. [Google Scholar] [CrossRef]
- Zhang, N.; Yu, H.; Yu, H.; Cai, Y.Y.; Huang, L.Z.; Xu, C.; Xiong, G.S.; Meng, X.B.; Wang, J.Y.; Chen, H.F.; et al. A Core Regulatory Pathway Controlling Rice Tiller Angle Mediated by the LAZY1-Dependent Asymmetric Distribution of Auxin. Plant Cell 2018, 30, 1461–1475. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Sang, D.J.; Huang, L.Z.; Song, Y.Q.; Du, M.C.; Cao, J.J.; Wang, W.G. PROG1 acts upstream of LAZY1 to regulate rice tiller angle as a repressor. Crop J. 2023, 11, 386–393. [Google Scholar] [CrossRef]
- Xu, X.; Liu, B.M.; Zhang, L.L.; Wu, Y.J. Mutagenic effects of heavy ion irradiation on rice seeds. Nucl. Instrum. Methods Phys. Res. 2012, 290, 19–25. [Google Scholar] [CrossRef]
- Edwards, K.; Johnstone, C.; Thompson, C. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucleic Acids Res. 1991, 19, 1349. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, J.Y. Molecular basis of plant architecture. Annu. Rev. Plant Bio. 2008, 59, 253–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Li, J.Y. Rice, rising. Nat. Genet. 2008, 40, 1273–1275. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.G.; Gao, H.B.; Liang, Y.; Li, J.Y.; Wang, Y.H. Molecular basis underlying rice tiller angle: Current progress and future perspectives. Mol. Plant. 2022, 15, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Lv, S.W.; Wu, W.G.; Fu, Y.C.; Liu, F.X.; Wang, B.B.; Li, W.G.; Gu, P.; Cai, H.W.; Sun, C.Q.; et al. The domestication of plant architecture in African rice. Plant J. 2018, 94, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Z.; Zhao, S.S.; Li, X.R.; Zhang, B.S.; Jiang, L.Y.; Tang, Y.Y.; Zhao, J.; Ma, X.; Cai, H.W.; Sun, C.Q.; et al. Deletions linked to PROG1 gene participate in plant architecture domestication in Asian and African rice. Nat. Commun. 2018, 9, 4157. [Google Scholar] [CrossRef]
- Jiang, J.H.; Tan, L.B.; Zhu, Z.F.; Fu, Y.C.; Liu, F.X.; Cai, H.W.; Sun, C.Q. Molecular evolution of the TAC1 gene from rice (Oryza sativa L.). J. Genet. Genomics 2012, 39, 551–560. [Google Scholar] [CrossRef]
- Li, Z.; Liang, Y.; Yuan, Y.D.; Wang, L.; Meng, X.B.; Xiong, G.S.; Zhou, J.; Cai, Y.Y.; Han, N.P.; Hua, L.K.; et al. OsBRXL4 Regulates Shoot Gravitropism and Rice Tiller Angle through Affecting LAZY1 Nuclear Localization. Mol. Plant 2019, 12, 1143–1156. [Google Scholar] [CrossRef]
- Dong, Z.B.; Jiang, C.; Chen, X.Y.; Zhang, T.; Ding, L.; Song, W.B.; Luo, H.B.; Lai, J.S.; Chen, H.B.; Liu, R.Y.; et al. Maize LAZY1 mediates shoot gravitropism and inflorescence development through regulating auxin transport, auxin signaling, and light response. Plant Physiol. 2013, 163, 1306–1322. [Google Scholar] [CrossRef]
- Chen, J.Y.; Yu, R.B.; Li, N.; Deng, Z.G.; Zhang, X.X.; Zhao, Y.R.; Qu, C.F.; Yuan, Y.F.; Pan, Z.X.; Zhou, Y.Y.; et al. Amyloplast sedimentation repolarizes LAZYs to achieve gravity sensing in plants. Cell 2023, 186, 4788–4802.e15. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrid Crosses. | F1 Phenotypes | F2 | χ23∶1 | ||
|---|---|---|---|---|---|
| HJX 74 Phenotypes | mtl1 Phenotypes | Total Number of Plants | |||
| mtl1 × HJX74 | HJX74 | 505 | 161 | 666 | 0.20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, Z.; Mei, T.; Feng, T.; Ai, H.; Ye, Y.; Duan, S.; Liu, B.; Huang, X. Deletion of the OsLA1 Gene Leads to Multi-Tillering and Lazy Phenotypes in Rice. Agriculture 2023, 13, 2166. https://doi.org/10.3390/agriculture13112166
Sun Z, Mei T, Feng T, Ai H, Ye Y, Duan S, Liu B, Huang X. Deletion of the OsLA1 Gene Leads to Multi-Tillering and Lazy Phenotypes in Rice. Agriculture. 2023; 13(11):2166. https://doi.org/10.3390/agriculture13112166
Chicago/Turabian StyleSun, Zhanglun, Tianrun Mei, Tingting Feng, Hao Ai, Yafeng Ye, Sumei Duan, Binmei Liu, and Xianzhong Huang. 2023. "Deletion of the OsLA1 Gene Leads to Multi-Tillering and Lazy Phenotypes in Rice" Agriculture 13, no. 11: 2166. https://doi.org/10.3390/agriculture13112166