
Corn Silk Extract: A Potential Modulator for Producing Functional Low Cholesterol Chicken Eggs

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Declaration
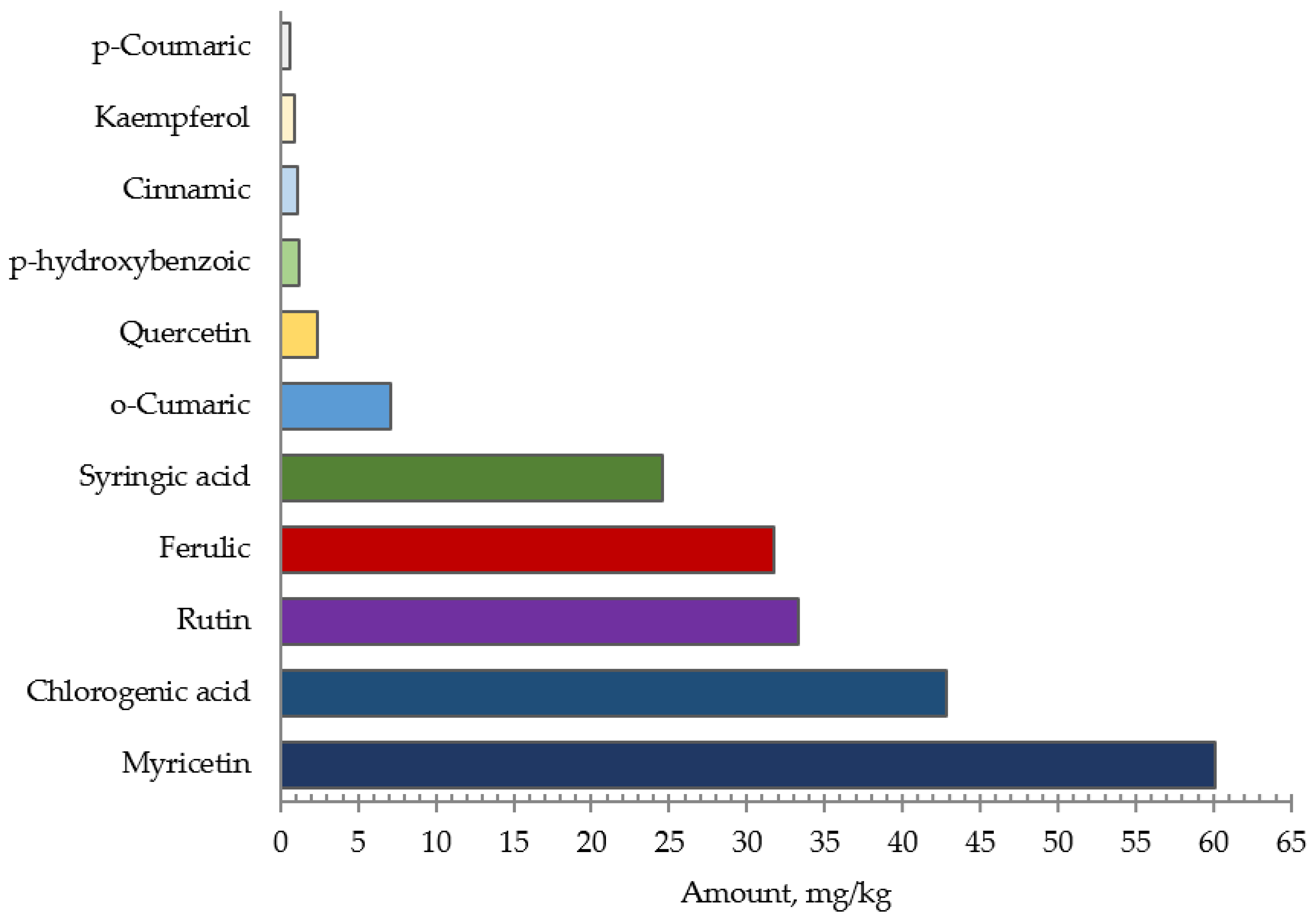
2.2. Corn Silk Extract Phenolic Acid Profile and Total Antioxidant Capacity Test
2.3. Birds’ Management and the Experimental Design
2.4. Egg Production Performance
2.5. Egg Quality Evaluation
 (AH-1.7W^0.37 + 7.57)]
(AH-1.7W^0.37 + 7.57)]
2.6. Collection and Preparation of Blood and Liver Samples
2.7. Blood and Liver Lipid Profile and Egg Yolk Cholesterol Content
2.8. Plasma Stress Markers and Biochemical Parameters
2.9. Statistical Analysis
3. Results
3.1. Egg Production Performance
3.2. Egg Quality Parameters
3.3. Blood Cholesterol Profile and Egg Cholesterol Content
3.4. Stress Markers and Liver Function
3.5. Liver and Kidney Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nowosad, K.; Sujka, M.; Pankiewicz, U.; Kowalski, R. The application of PEF technology in food processing and human nutrition. J. Food Sci. Technol. 2021, 58, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Balthazar, C.F.; Guimarães, J.F.; Coutinho, N.M.; Pimentel, T.C.; Ranadheera, C.S.; Santillo, A.; Albenzio, M.; Cruz, A.G.; Sant’Ana, A.S. The future of functional food: Emerging technologies application on prebiotics, probiotics and postbiotics. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2560–2586. [Google Scholar] [CrossRef] [PubMed]
- Papanikolaou, Y.; Fulgoni, V.L. 3rd. Eggs are cost-efficient in delivering several shortfall nutrients in the American diet: A cost-analysis in children and adults. Nutrients 2020, 12, 2406. [Google Scholar] [CrossRef] [PubMed]
- Spence, J.D.; Jenkins, D.J.; Davignon, J. Dietary cholesterol and egg yolks: Not for patients at risk of vascular disease. Can. J. Cardiol. 2010, 26, e336–e339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krittanawong, C.; Narasimhan, B.; Wang, Z.; Virk, H.U.H.; Farrell, A.M.; Zhang, H.; Tang, W.H.W. Association between egg consumption and risk of cardiovascular outcomes: A systematic review and meta-analysis. Am. J. Med. 2021, 134, 76–83.e72. [Google Scholar] [CrossRef] [PubMed]
- Zhong, V.W.; Van Horn, L.; Cornelis, M.C.; Wilkins, J.T.; Ning, H.; Carnethon, M.R.; Greenland, P.; Mentz, R.J.; Tucker, K.L.; Zhao, L.; et al. Associations of dietary cholesterol or egg consumption with incident cardiovascular disease and mortality. JAMA 2019, 321, 1081–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Wu, F.; Mao, L.; Zhu, F.; Zhang, Y.; Chen, X.; Jiao, J.; Zhang, Y. Egg and cholesterol consumption and mortality from cardiovascular and different causes in the United States: A population-based cohort study. PLoS Med. 2021, 18, e1003508. [Google Scholar] [CrossRef]
- Drouin-Chartier, J.-P.; Chen, S.; Li, Y.; Schwab, A.L.; Stampfer, M.J.; Sacks, F.M.; Rosner, B.; Willett, W.C.; Hu, F.B.; Bhupathiraju, S.N. Egg consumption and risk of cardiovascular disease: Three large prospective US cohort studies, systematic review, and updated meta-analysis. BMJ 2020, 368, m513. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Lv, J.; Guo, Y.; Bian, Z.; Si, J.; Yang, L.; Chen, Y.; Zhou, Y.; Zhang, H.; Liu, J.; et al. Associations of egg consumption with cardiovascular disease in a cohort study of 0.5 million Chinese adults. Heart 2018, 104, 1756. [Google Scholar] [CrossRef] [Green Version]
- Key, T.J.; Appleby, P.N.; Bradbury, K.E.; Sweeting, M.; Wood, A.; Johansson, I.; Kühn, T.; Steur, M.; Weiderpass, E.; Wennberg, M.; et al. Consumption of meat, fish, dairy products, and eggs and risk of ischemic heart disease. Circulation 2019, 139, 2835–2845. [Google Scholar] [CrossRef]
- Carson, J.A.S.; Lichtenstein, A.H.; Anderson, C.A.M.; Appel, L.J.; Kris-Etherton, P.M.; Meyer, K.A.; Petersen, K.; Polonsky, T.; Van Horn, L. Dietary cholesterol and cardiovascular risk: A Science advisory from the american heart association. Circulation 2020, 141, e39–e53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, S. The role of chicken eggs in human nutrition. J. Food Nutr. Metab. 2021, 3, 2–9. [Google Scholar] [CrossRef]
- Nimalaratne, C.; Wu, J. Hen egg as an antioxidant food commodity: A review. Nutrients 2015, 7, 8274–8293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Jia, R.; Gong, H.; Celi, P.; Zhuo, Y.; Ding, X.; Bai, S.; Zeng, Q.; Yin, H.; Xu, S.; et al. The effect of oxidative stress on the chicken ovary: Involvement of microbiota and melatonin interventions. Antioxidants 2021, 10, 1422. [Google Scholar] [CrossRef]
- Li, G.-M.; Liu, L.-P.; Yin, B.; Liu, Y.-Y.; Dong, W.-W.; Gong, S.; Zhang, J.; Tan, J.-H. Heat stress decreases egg production of laying hens by inducing apoptosis of follicular cells via activating the FasL/Fas and TNF-α systems. Poult. Sci. 2020, 99, 6084–6093. [Google Scholar] [CrossRef]
- Abbas, A.O.; Alaqil, A.A.; El-Beltagi, H.S.; Abd El-Atty, H.K.; Kamel, N.N. Modulating laying hens productivity and immune performance in response to oxidative stress induced by E. coli challenge using dietary propolis supplementation. Antioxidants 2020, 9, 893. [Google Scholar] [CrossRef]
- Hasanudin, K.; Hashim, P.; Mustafa, S. Corn silk (Stigma maydis) in healthcare: A phytochemical and pharmacological review. Molecules 2012, 17, 9697–9715. [Google Scholar] [CrossRef] [Green Version]
- Tian, S.; Sun, Y.; Chen, Z. Extraction of flavonoids from corn silk and biological activities in vitro. J. Food Qual. 2021, 2021, 7390425. [Google Scholar] [CrossRef]
- Nawaz, H.; Muzaffar, S.; Aslam, M.; Ahmad, S. Phytochemical Composition: Antioxidant Potential and Biological Activities of Corn in Corn—Production and Human Health in Changing Climate; Amanullah, A., Fahad, S., Eds.; IntechOpen: London, UK, 2018; pp. 49–67. [Google Scholar]
- Shi, S.; Yu, B.; Li, W.; Shan, J.; Ma, T. Corn silk decoction for blood lipid in patients with angina pectoris: A systematic review and meta-analysis. Phytother. Res. 2019, 33, 2862–2869. [Google Scholar] [CrossRef]
- Solihah, M.A.; Rosli, W.D.W.; Nurhanan, A.R. Phytochemicals screening and total phenolic content of Malaysian Zea mays hair extracts. Int. Food Res. J. 2012, 19, 1533–1538. [Google Scholar]
- Guo, J.; Liu, T.; Han, L.; Liu, Y. The effects of corn silk on glycaemic metabolism. Nutr. Metab. 2009, 6, 47. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Da-yun, S.; Jing-shu, Z.; Hong-li, Z. Microwave-assisted extraction and antihyperlipidemic effect of total flavonoids from corn silk. Afr. J. Biotechnol. 2011, 10, 14583–14586. [Google Scholar] [CrossRef] [Green Version]
- Haslina; Wahjuningsih, S.B. Effect of corn silk powder extracts using in vivo to lipid profile and liver fat. IOP Conf. Ser. Earth Environ. Sci. 2020, 443, 012010. [Google Scholar] [CrossRef]
- Saheed, S.; Oladipipo, A.E.; Abdulazeez, A.A.; Olarewaju, S.A.; Ismaila, N.O.; Emmanuel, I.A.; Fatimah, Q.D.; Aisha, A.Y. Toxicological evaluations of Stigma maydis (corn silk) aqueous extract on hematological and lipid parameters in Wistar rats. Toxicol. Rep. 2015, 2, 638–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirrella, A.A.; Abdo, S.E.; El-Naggar, K.; Soliman, M.M.; Aboelenin, S.M.; Dawood, M.A.O.; Saleh, A.A. Use of corn silk meal in broiler diet: Effect on growth performance, blood biochemistry, immunological responses, and growth-related gene expression. Animals 2021, 11, 1170. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.Q.; Dong, Y.Y.; Qin, X.; Yuan, J.M.; Han, M.M.; Zhang, K.K.; Shi, S.R.; Song, X.Y.; Zhang, J.Z.; Li, J.H. Dietary supplementation of methionine mitigates oxidative stress in broilers under high stocking density. Poult. Sci. 2021, 100, 101231. [Google Scholar] [CrossRef] [PubMed]
- El-Mergawi, R.; Al-Humaid, A.; El-Rayes, D. Phenolic profiles and antioxidant activity in seeds of ten date cultivars from Saudi Arabia. J. Food Agric. Environ. 2016, 14, 38–43. [Google Scholar]
- Haugh, R. The Haugh unit for measuring egg quality. U.S. Egg Poult. Mag. 1937, 43, 552–555. [Google Scholar]
- Nekouei, O.; Yau, D.; MacKinnon, B.; Magouras, I.; Conan, A.; Elsohaby, I.; Paudel, S.; Pfeiffer, D.U. Quality assessment of day-old chickens on the broiler farms of Hong Kong. Animals 2022, 12, 1520. [Google Scholar] [CrossRef]
- Alzarah, M.I.; Alaqil, A.A.; Abbas, A.O.; Nassar, F.S.; Mehaisen, G.M.K.; Gouda, G.F.; Abd El-Atty, H.K.; Moustafa, E.S. Inclusion of Citrullus colocynthis seed extract into diets Induced a hypolipidemic effect and improved layer performance. Agriculture 2021, 11, 808. [Google Scholar] [CrossRef]
- Wang, B.; Xiao, T.; Ruan, J.; Liu, W. Beneficial effects of corn silk on metabolic syndrome. Curr. Pharm. Des. 2017, 23, 5097–5103. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.J.; Zhao, J.L. Corn silk (Zea mays L.), a source of natural antioxidants with α-amylase, α-glucosidase, advanced glycation and diabetic nephropathy inhibitory activities. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 110, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Al-Harthi, M.A. The Effect of natural and synthetic antioxidants on performance, egg quality and blood constituents of laying hens grown under high ambient temperature. Ital. J. Anim. Sci. 2014, 13, 3239. [Google Scholar] [CrossRef]
- Abdel Magied, H.A.; Selim, N.A.; Habib, H.H.; El- Komy, H.M.A.; Mostafa, M.A.S.A. Strengthening the antioxidant status of laying hens through summer season by using different categories of antioxidant sources. J. Anim. Poult. Prod. 2020, 11, 539–547. [Google Scholar] [CrossRef]
- Obianwuna, U.E.; Oleforuh-Okoleh, V.U.; Wang, J.; Zhang, H.J.; Qi, G.H.; Qiu, K.; Wu, S.G. Potential implications of natural antioxidants of plant origin on oxidative stability of chicken albumen during storage: A review. Antioxidants 2022, 11, 630. [Google Scholar] [CrossRef]
- Bai, D.; Liu, K.; He, X.; Tan, H.; Liu, Y.; Li, Y.; Zhang, Y.; Zhen, W.; Zhang, C.; Ma, Y. Effect of dietary chlorogenic acid on growth performance, antioxidant function, and immune response of broiler breeders under immune stress and stocking density stress. Vet. Sci. 2022, 9, 582. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, H.; Zhang, J.; Zhou, B.; Zhuang, S.; He, X.; Wang, T.; Wang, C. Effects of different levels of rutin on growth performance, immunity, intestinal barrier and antioxidant capacity of broilers. Ital. J. Anim. Sci. 2022, 21, 1390–1401. [Google Scholar] [CrossRef]
- Millán, J.; Pintó, X.; Muñoz, A.; Zúñiga, M.; Rubiés-Prat, J.; Pallardo, L.F.; Masana, L.; Mangas, A.; Hernández-Mijares, A.; González-Santos, P.; et al. Lipoprotein ratios: Physiological significance and clinical usefulness in cardiovascular prevention. Vasc. Health Risk Manag. 2009, 5, 757–765. [Google Scholar]
- Cha, J.H.; Kim, S.R.; Kang, H.J.; Kim, M.H.; Ha, A.W.; Kim, W.K. Corn silk extract improves cholesterol metabolism in C57BL/6J mouse fed high-fat diets. Nutr. Res. Pract. 2016, 10, 501–506. [Google Scholar] [CrossRef] [Green Version]
- Hargis, P.S. Modifying egg yolk cholesterol in the domestic fowl—A review. World’s Poult. Sci. J. 1988, 44, 17–29. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, W.; Du, Y.; Liu, X.; Geng, Z. Genetic parameters for yolk cholesterol and transcriptional evidence indicate a role of lipoprotein lipase in the cholesterol metabolism of the Chinese wenchang chicken. Front. Genet. 2019, 10, 902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A dietary molecule with diverse biological activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Chang, C.; Zhang, L.; Liu, Y.; Huang, X.; Chen, Z. Chlorogenic acid improves late diabetes through adiponectin receptor signaling pathways in db/db mice. PLoS ONE 2015, 10, e0120842. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The pharmacological potential of rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant defence systems and oxidative stress in poultry biology: An update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F. Antioxidant systems in poultry biology: Superoxide dismutase. J. Anim. Res. Nutr. 2016, 1, 1–17. [Google Scholar] [CrossRef]
- Ognik, K.; Cholewińska, E.; Sembratowicz, I.; Grela, E.; Czech, A. The potential of using plant antioxidants to stimulate antioxidant mechanisms in poultry. World’s Poult. Sci. J. 2016, 72, 291–298. [Google Scholar] [CrossRef]
- Czerska, M.; Mikołajewska, K.; Zieliński, M.; Gromadzińska, J.; Wąsowicz, W. Today’s oxidative stress markers. Med. Pracy 2015, 66, 393–405. [Google Scholar] [CrossRef]
- Ho, E.; Karimi Galougahi, K.; Liu, C.C.; Bhindi, R.; Figtree, G.A. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [Green Version]
- El Golli-Bennour, E.; Bacha, H. Hsp70 expression as biomarkers of oxidative stress: Mycotoxins’ exploration. Toxicology 2011, 287, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Abass, A.O.; Kamel, N.N.; Khalifa, W.H.; Gouda, G.F.; El-Manylawi, M.A.F.; Mehaisen, G.M.K.; Mashaly, M.M. Propolis supplementation attenuates the negative effects of oxidative stress induced by paraquat injection on productive performance and immune function in turkey poults. Poult. Sci. 2017, 96, 4419–4429. [Google Scholar] [CrossRef] [PubMed]
- Akbarian, A.; Michiels, J.; Degroote, J.; Majdeddin, M.; Golian, A.; De Smet, S. Association between heat stress and oxidative stress in poultry; mitochondrial dysfunction and dietary interventions with phytochemicals. J. Anim. Sci. Biotechnol. 2016, 7, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.; Inbaraj, B.S.; Kaur, S.; Rasane, P.; Nanda, V. Phytochemical analysis and characterization of corn silk (Zea mays, G5417). Agronomy 2022, 12, 777. [Google Scholar] [CrossRef]
- Rajeshwari, H.; Sivapriya, T. Analysis of nutrients, phytochemicals, antioxidant and antimicrobial activity of corn silk extract (Zea mays L. Stigma). Int. J. Health Allied Sci. 2021, 10, 275–279. [Google Scholar] [CrossRef]
- Rahman, N.A.; Wan Rosli, W.I. Nutritional compositions and antioxidative capacity of the silk obtained from immature and mature corn. J. King Saud Univ. Sci. 2014, 26, 119–127. [Google Scholar] [CrossRef] [Green Version]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Suresh Kumar, C. Syringic acid (SA)—A review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Subramani, P.; Sampathkumar, N.; Ravindiran, G.; Rajalingam, D.; Kumar, B. Evaluation of nephroprotective and antioxidant potential of Tragia involucrata. Drug Invent. Today 2009, 1, 55–60. [Google Scholar]
- Pandya, D.; Nagrajappa, A.K.; Ravi, K.S. Assessment and correlation of urea and creatinine levels in saliva and serum of patients with chronic kidney disease, diabetes and hypertension- A research study. J. Clin. Diagn. Res. JCDR 2016, 10, ZC58–ZC62. [Google Scholar] [CrossRef]
- Wans, E.M.; Ahmed, M.M.; Mousa, A.A.; Tahoun, E.A.; Orabi, S.H. Ameliorative effects of corn silk extract on acetaminophen-induced renal toxicity in rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 1762–1774. [Google Scholar] [CrossRef]
- Chen, J.Y.; Sun, X.Y.; Ouyang, J.M. Modulation of calcium oxalate crystal growth and protection from oxidatively damaged renal epithelial cells of corn silk polysaccharides with different molecular weights. Oxidative Med. Cell. Longev. 2020, 2020, 6982948. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Ingredients | Content |
|---|---|
| Yellow corn, g/kg | 565.5 |
| Soybean meal (44%), g/kg | 276.0 |
| Wheat bran, g/kg | 10.0 |
| Soybean oil, g/kg | 30.0 |
| Bone meal, g/kg | 30.0 |
| Limestone, g/kg | 80.0 |
| Salt, g/kg | 4.0 |
| Premix 1, g/kg | 3.0 |
| DL-methionine, g/kg | 1.5 |
| Calculated chemical analysis | |
| Metabolizable energy, kcal/kg | 301.15 |
| Crude protein, g/kg | 174.7 |
| Calcium, g/kg | 40.2 |
| Available phosphorus, g/kg | 5.2 |
| Lysine, g/kg | 9.5 |
| Methionine, g/kg | 4.2 |
| Linoleic acid, g/kg | 28.8 |
| Chemical analysis (%) | |
| Dry matter | 89.00 |
| Crude protein | 16.75 |
| Crude fat | 6.60 |
| Crude fiber | 4.70 |
| Ash | 12.90 |
| Calcium | 4.22 |
| Phosphorus | 0.42 |
| Parameters | Control | CSE-100 | CSE-200 | CSE-400 |
|---|---|---|---|---|
| EN, 7d | 6.38 b ± 0.06 | 6.52 b ± 0.07 | 6.82 a ± 0.05 | 6.66 a ± 0.02 |
| EW, g/egg | 59.13 d ± 0.15 | 60.40 c ± 0.15 | 61.67 b ± 0.15 | 62.73 a ± 0.11 |
| Egg mass/week | 378.12 d ± 1.98 | 390.34 c ± 1.79 | 404.97 b ± 1.83 | 417.68 a ± 1.64 |
| FI, g/day | 113.42 b ± 0.18 | 112.52 c ± 0.19 | 111.60 d ± 0.15 | 115.13 a ± 0.17 |
| FCR | 2.10 a ± 0.01 | 2.02 b ± 0.01 | 1.93 c ± 0.01 | 1.93 c ± 0.01 |
| Parameters | Control | CSE-100 | CSE-200 | CSE-400 |
|---|---|---|---|---|
| Albumin index | 9.45 c ± 0.06 | 9.73 b ± 0.04 | 9.84 b ± 0.03 | 10.01 a ± 0.04 |
| Yolk index | 40.38 c ± 0.05 | 41.10 b ± 0.05 | 42.27 a ± 0.35 | 42.31 a ± 0.06 |
| Yolk color | 7.92 d ± 0.05 | 8.49 c ± 0.04 | 9.09 ± 0.03b | 9.67 a ± 0.06 |
| Shell thickness, mm | 0.31 d ± 0.001 | 0.35 c ± 0.001 | 0.37 b ± 0.002 | 0.40 a ± 0.002 |
| Shell strength, kg/cm2 | 3.76 d ± 0.01 | 3.95 c ± 0.01 | 4.29 b ± 0.03 | 4.49 a ± 0.03 |
| Haugh unit | 81.78 b ± 0.38 | 81.91 b ± 0.14 | 83.38 a ± 0.049 | 82.96 ab ± 0.35 |
| Parameters | Control | CSE-100 | CSE-200 | CSE-400 |
|---|---|---|---|---|
| Triglycerides, mg/dL | 216.8 a ± 6.22 | 199.5 ab ± 4.45 | 180.0 b ± 4.92 | 158.5 c ± 4.57 |
| Cholesterol, mg/dL | 165.7 a ± 6.13 | 166.2 a ± 5.31 | 148.2 ab ± 3.74 | 135.5 b ± 6.40 |
| HDL-CH, mg/dL | 42.40 c ± 1.07 | 45.53 bc ± 0.96 | 47.53 b ± 0.73 | 55.23 a ± 0.88 |
| LDL-CH, mg/dL | 114.5 a ± 1.23 | 101.5 b ± 3.80 | 94.3 b ± 2.94 | 82.9 c ± 1.74 |
| Plasma cholesterol ratio | 3.92 a ± 0.15 | 3.66 a ± 0.14 | 3.12 b ± 0.06 | 2.45 c ± 0.10 |
| Liver CH, mg/dL | 5.73 a ± 0.14 | 4.90 b ± 0.09 | 4.25 c ± 0.15 | 3.42 d ± 0.08 |
| Egg cholesterol content, mg/g | 12.82 a ± 0.18 | 11.68 b ± 0.14 | 11.20 b ± 0.15 | 10.43 c ± 0.08 |
| Parameters | Control | CSE-100 | CSE-200 | CSE-400 |
|---|---|---|---|---|
| MDA, μM/mL | 2.74 a ± 0.19 | 2.41 ab ± 0.15 | 1.96 bc ± 0.07 | 1.75 c ± 0.07 |
| TNF-α, pg/mL | 96.25 a ± 1.11 | 91.78 ab ± 1.15 | 87.57 bc ± 1.92 | 82.97 c ± 1.52 |
| HSP-70, ng/mL | 26.19 a ± 1.30 | 21.67 b ± 0.79 | 20.59 b ± 0.95 | 20.03 b ± 1.21 |
| Corticosterone, ng/mL | 6.31 a ± 0.20 | 6.10 a ± 0.30 | 5.38 ab ± 0.21 | 4.57 b ± 0.25 |
| SOD, U/mL | 4.73 c ± 0.13 | 5.04 bc ± 0.10 | 5.26 b ± 0.06 | 5.66 a ± 0.06 |
| T-AOC, U/mL | 6.47 d ± 0.17 | 7.17 c ± 0.16 | 8.13 b ± 0.10 | 8.90 a ± 0.10 |
| Parameters | Control | CSE-100 | CSE-200 | CSE-400 |
|---|---|---|---|---|
| Total protein, g/dL | 4.89 a ± 0.12 | 4.96 a ± 0.11 | 5.10 a ± 0.09 | 5.26 a ± 0.22 |
| Albumin, g/dL | 2.62 a ± 0.13 | 2.35 a ± 0.13 | 2.79 a ± 0.26 | 2.65 a ± 0.18 |
| Globulin, g/dL | 2.26 a ± 0.21 | 2.61 a ± 0.14 | 2.32 a ± 0.28 | 2.61 a ± 0.38 |
| ALT, U/mL | 13.39 a ± 0.42 | 12.15 b ± 0.17 | 11.51 b ± 0.20 | 10.12 c ± 0.21 |
| AST, U/mL | 30.78 a ± 1.28 | 27.03 ab ± 0.80 | 23.63 bc ± 0.96 | 21.07 c ± 0.70 |
| Urea, mg/dL | 5.88 a ± 0.05 | 5.42 b ± 0.10 | 5.21 b ± 0.04 | 4.78 c ± 0.08 |
| Creatinine, mg/dL | 0.30 a ± 0.01 | 0.28 ab ± 0.004 | 0.26 bc ± 0.006 | 0.24 c ± 0.004 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, A.O.; Alaqil, A.A.; Kamel, N.N.; Nassar, F.S. Corn Silk Extract: A Potential Modulator for Producing Functional Low Cholesterol Chicken Eggs. Agriculture 2023, 13, 65. https://doi.org/10.3390/agriculture13010065
Abbas AO, Alaqil AA, Kamel NN, Nassar FS. Corn Silk Extract: A Potential Modulator for Producing Functional Low Cholesterol Chicken Eggs. Agriculture. 2023; 13(1):65. https://doi.org/10.3390/agriculture13010065
Chicago/Turabian StyleAbbas, Ahmed O., Abdulaziz A. Alaqil, Nancy N. Kamel, and Farid S. Nassar. 2023. "Corn Silk Extract: A Potential Modulator for Producing Functional Low Cholesterol Chicken Eggs" Agriculture 13, no. 1: 65. https://doi.org/10.3390/agriculture13010065