
Distribution of Genetic Diversity in Beta patula Aiton Populations from Madeira Archipelago, Portugal

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Sampling Procedure
2.2. Genetic Analysis
2.3. Data Treatment and Statistical Methods
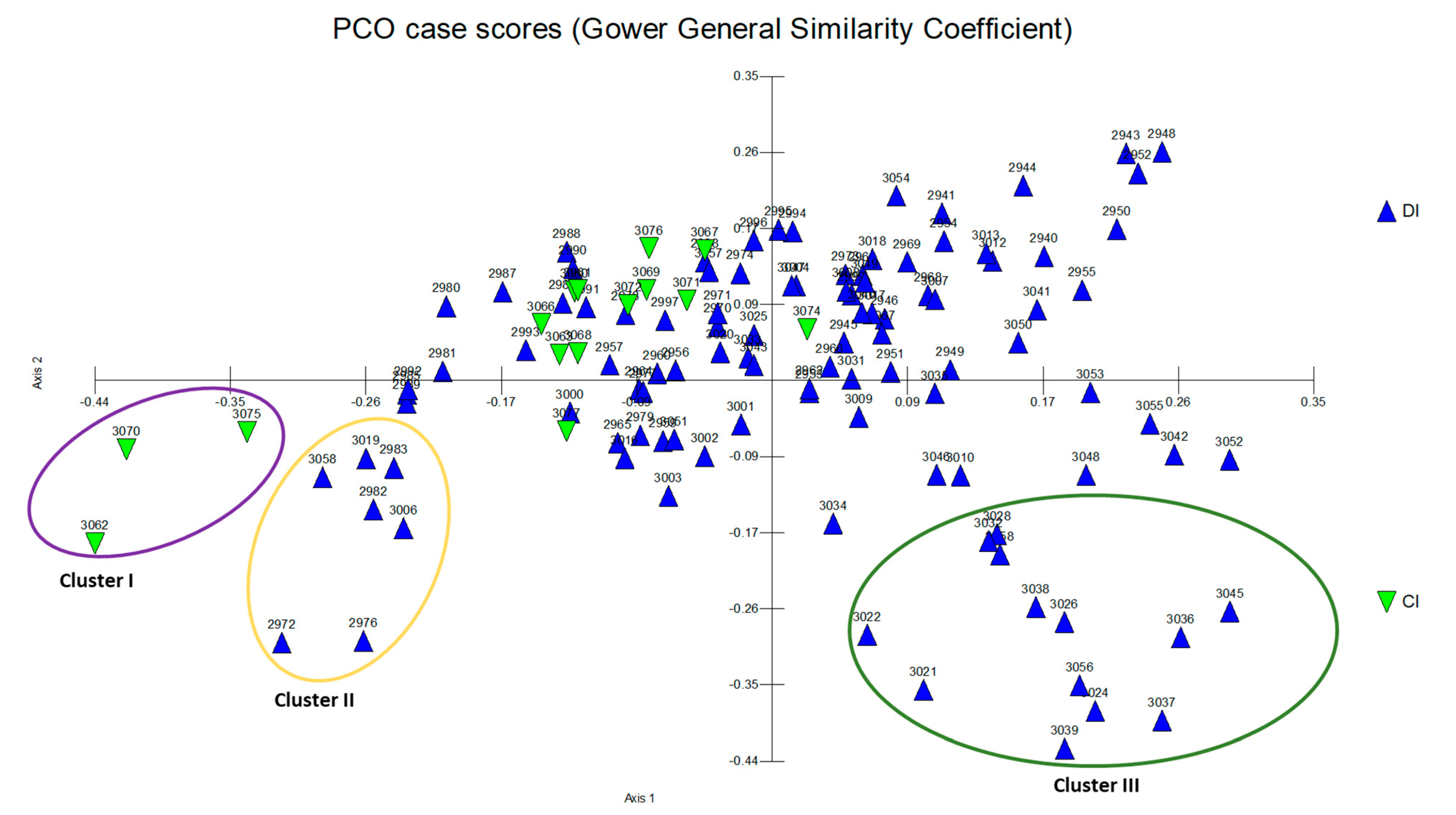
3. Results
4. Discussion
5. Final Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abreu, C.; Aguiar, A.M.; Carvalho, P.; Jardim, R.; Melo, I.; Oliveira, P.; Sérgio, C.; Serrano, A.R.; Vieira, P. Listagem dos Fungos, Flora e Fauna Terrestres dos Arquipélagos da Madeira e Selvagens: A list of the Terrestrial Fungi, Flora and Fauna of Madeira and Selvagens Archipelagos; Borges, P.A., Ed.; Direcçao Regional do Ambiente, Governo Regional da Madeira: Funchal, Portugal, 2008. [Google Scholar]
- Vincent, H.; Amri, A.; Castañeda-Álvarez, N.P.; Dempewolf, H.; Dulloo, E.; Guarino, L.; Hole, D.; Mba, C.; Toledo, A.; Maxted, N. Modeling of crop wild relative species identifies areas globally for in situ conservation. Commun. Biol. 2019, 2, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frese, L. Sugar beets and related wild species—From collecting to utilisation. In Schriften zu Genetischen Ressourcen; Knüpffer, H., Ochsmann, J., Eds.; Rudolf Mansfeld and Plant Genetic Resources; Martin Roesberg: Witterschlick, Germany, 2003; Volume 22, pp. 170–181. [Google Scholar]
- Frese, L. Conservation and Access to Sugarbeet Germplasm. Sugar Tech 2010, 12, 207–219. [Google Scholar] [CrossRef]
- Harlan, J.R.; de Wet, J.M. Toward a Rational Classification of Cultivated Plants. Taxon 1971, 20, 509–517. [Google Scholar] [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; European Commission, Directorate-General for Environment, Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Food and Agriculture Organization of the United Nations—FAO. International Treaty on Plant Genetic Resources for Food and Agriculture; Annex, I., Ed.; FAO: Rome, Italy, 2009. [Google Scholar]
- Goldman, I.L.; Navazio, J.P. History and Breeding of Table Beet in the United States. In Plant Breeding Reviews; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2002; pp. 357–388. [Google Scholar]
- Dohm, J.C.; Minoche, A.E.; Holtgräwe, D.; Capella-Gutiérrez, S.; Zakrzewski, F.; Tafer, H.; Rupp, O.; Sörensen, T.R.; Stracke, R.; Reinhardt, R.; et al. The genome of the recently domesticated crop plant sugar beet (Beta vulgaris). Nature 2014, 505, 546–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nóbrega, H.; Freitas, G.; Zavattieri, M.A.; Ragonezi, C.; Frese, L.; de Carvalho, M.A.A.P. Monitoring system and in situ conservation of endemic and threatened Beta patula Aiton populations in Madeira Region. Genet. Resour. Crop Evol. 2021, 68, 939–956. [Google Scholar] [CrossRef]
- Pinheiro de Carvalho, M.A.; Nóbrega, H.; Frese, L.; Freitas, G.; Abreu, U.; Costa, G.; Fontinha, S. Distribution and abundance of Beta patula Aiton and other crop wild relatives of cultivated beets on Madeira. J. Cultiv. Plants 2010, 62, 357–366. [Google Scholar]
- Pinheiro De Carvalho, M.A.A.; Nóbrega, H.; Freitas, G.; Fontinha, S.; Frese, L. Towards the establishment of a genetic reserve for Beta patula aiton. In Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces; Maxted, N., Dulloo, M.E., Ford-Lloyd, B.V., Frese, L., Iriondo, J., de Carvalho, M.A.P., Eds.; CABI Publishing: Wallingford, UK, 2012; pp. 36–44. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M.; Matthews, T.J.; Borregaard, M.K.; Triantis, K.A. Island biogeography: Taking the long view of natures labor atories. Science 2017, 357, eaam8326. [Google Scholar] [CrossRef] [Green Version]
- Fois, M.; Podda, L.; Médail, F.; Bacchetta, G. Endemic and alien vascular plant diversity in the small Mediterranean islands of Sardinia: Drivers and implications for their conservation. Biol. Conser. 2020, 244, 108519. [Google Scholar] [CrossRef]
- Traveset, A.; Navarro, L. Plant reproductive ecology and evolution in the Mediterranean islands: State of the art. Plant Biol. 2018, 20, 63–77. [Google Scholar] [CrossRef] [Green Version]
- Médail, F. The specific vulnerability of plant biodiversity and vegetation on Mediterranean islands in the face of global change. Reg. Environ. Chang. 2017, 17, 1775–1790. [Google Scholar] [CrossRef] [Green Version]
- Sciandrello, S.; Cambria, S.; del Galdo, G.G.; Guarino, R.; Minissale, P.; Pasta, S.; Tavilla, G.; Cristaudo, A. Floristic and vegetation changes on a small mediterranean island over the last century. Plants 2021, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- Maxted, N.; Avagyan, A.; Frese, L.; Iriondo, J.; Brehm, J.M.; Singer, A.; Kell, S. Preserving Diversity: A Concept for In Situ Conservation of Crop Wild Relatives in Europe. In Situ and On-farm Conservation Network; European Cooperative Programme for Plant Genetic Resources: Rome, Italy, 2013. [Google Scholar]
- Greaver, T.L.; Sternberg, L.S.L. Decreased precipitation exacerbates the effects of sea level on coastal dune ecosystems in open ocean islands. Glob. Chang. Biol. 2010, 16, 1860–1869. [Google Scholar] [CrossRef]
- Frese, L. Combining static and dynamic management of PGR: A case study of Beta genetic resources. In Managing Plant Genetic Diversity; CABI Publishing: Wallingford, UK, 2002; pp. 133–147. [Google Scholar]
- Maxted, N.; Ford-Lloyd, B.V.; Jury, S.; Kell, S.; Scholten, M. Towards a definition of a crop wild relative. Biodivers Conserv. 2006, 15, 2673–2685. [Google Scholar] [CrossRef]
- Kell, S.P.; Maxted, N.; Frese, L.; Iriondo, J.M. In situ Conservation of Crop Wild Relatives: A Strategy for Identifying Priority Genetic Reserve Sites. In Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces; CAB International: Wallingford, UK, 2012; Volume 1, pp. 7–19. [Google Scholar]
- Young, A.G.; Brown, A.H.D. Comparative population genetic structure of the rare woodland shrub Daviesia suaveolens and its common congener D. mimosoides. Conserv. Biol. 1996, 10, 1220–1228. [Google Scholar] [CrossRef]
- Ribeiro, I.C.; Pinheiro, C.; Ribeiro, C.M.; Veloso, M.M.; Simões-Costa, M.C.; Evaristo, I.; Paulo, O.S.; Ricardo, C.P. Genetic diversity and physiological performance of portuguese wild beet (Beta vulgaris spp. maritima) from three contrasting habitats. Front. Plant Sci. 2016, 7, 1293. [Google Scholar] [CrossRef] [Green Version]
- Frese, L.; Nachtigall, M.; Enders, M.; Carvalho, M.A.D. Beta patula Aiton: Genetic diversity analysis. In Agrobiodiversity Conservation: Securing the Diversity of Crop Wild Relatives and Landraces; CABI Publishing: Wallingford, UK, 2012; pp. 45–51. [Google Scholar]
- Silva, L.B.; Sardos, J.; de Sequeira, M.M.; Silva, L.; Crawford, D.; Moura, M. Understanding intra and inter-archipelago population genetic patterns within a recently evolved insular endemic lineage. Plant Syst. Evol. 2016, 302, 367–384. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Elam, D.R. Population genetic consequences of small population size: Implications for plant conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]
- Nóbrega, H.; Freitas, G.; Pinheiro de Carvalho, M.Â. Beta patula Population Assessment from 2014 to 2022 in Madeira Archipelago, Portugal, Version 1.13; Sampling Event Dataset; University of Madeira: Funchal, Portugal, 2022. [Google Scholar] [CrossRef]
- Rodríguez del Río, Á.; Minoche, A.E.; Zwickl, N.F.; Friedrich, A.; Liedtke, S.; Schmidt, T.; Himmelbauer, H.; Dohm, J.C. Genomes of the wild beets Beta patula and Beta vulgaris ssp. maritima. Plant J. 2019, 99, 1242–1253. [Google Scholar] [CrossRef] [Green Version]
- Wascher, F.L.; Stralis-Pavese, N.; McGrath, J.M.; Schulz, B.; Himmelbauer, H.; Dohm, J.C. Genomic distances reveal relationships of wild and cultivated beets. Nat. Commun. 2022, 13, 2021. [Google Scholar] [CrossRef]
- Desplanque, B.; Boudry, P.; Broomberg, K.; Saumitou-Laprade, P.; Cuguen, J.; Van Dijk, H. Genetic diversity and gene flow between wild, cultivated and weedy forms of Beta vulgaris L. (Chenopodiaceae), assessed by RFLP and microsatellite markers. Theor. Appl. Genet. 1999, 98, 1194–1201. [Google Scholar] [CrossRef]
- Simko, I.; Eujayl, I.; van Hintum, T.J.L. Empirical evaluation of DArT, SNP, and SSR marker-systems for genotyping, clustering, and assigning sugar beet hybrid varieties into populations. Plant Sci. 2012, 184, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Laurent, V.; Devaux, P.; Thiel, T.; Viard, F.; Mielordt, S.; Touzet, P.; Quillet, M.C. Comparative effectiveness of sugar beet microsatellite markers isolated from genomic libraries and GenBank ESTs to map the sugar beet genome. Theor. Appl. Genet. 2007, 115, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.; Romeiras, M.M.; Castro, M.; Duarte, M.C.; Loureiro, J. Hidden diversity in wild Beta taxa from Portugal: Insights from genome size and ploidy level estimations using flow cytometry. Plant Sci. 2013, 207, 72–78. [Google Scholar] [CrossRef]
- Chao, S.; Somers, D.; Wheat and Barley DNA Extraction in 96-Well Plates. MAS Wheat. Available online: https://www.yumpu.com/en/document/view/11880064/wheat-and-barley-dna-extraction-in-96-well-plates-maswheat (accessed on 1 June 2017).
- Abbasi, Z.; Majidi, M.M.; Arzani, A.; Rajabi, A.; Mashayekhi, P.; Bocianowski, J. Association of SSR markers and morpho-physiological traits associated with salinity tolerance in sugar beet (Beta vulgaris L.). Euphytica 2015, 205, 785–797. [Google Scholar] [CrossRef]
- McGrath, J.M.; Trebbi, D.; Fenwick, A.; Panella, L.; Schulz, B.; Laurent, V.; Barnes, S.; Murray, S.C. An open-source first-generation molecular genetic map from a sugarbeet × table beet cross and its extension to physical mapping. Crop. Sci. 2007, 47, 27–44. [Google Scholar] [CrossRef]
- Gower, J.C. Some distance properties of latent root and vector methods used in multivariate analysis. Biometrika 1966, 53, 325–338. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar]
- ESRI. Inc Esri Support ArcMap 10.6. 2018. Available online: https://support.esri.com/en/products/desktop/arcgis-desktop/arcmap/10-6-1 (accessed on 1 June 2021).
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE Software Package Version 1.31 for Population Genetic Analysis; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Analysis of Gene Diversity in Subdivided Populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [Green Version]
- Ascarini, F.; Nóbrega, H.G.M.; Leite, I.S.; Freitas, G.; Ragonezi, C.; Amely Zavattieri, M.; Pinheiro de Carvalho, M.A.A. Assessing the diversity of sea beet (Beta vulgaris L. ssp. maritima) populations. J. Agric. Sci. Technol. 2021, 23, 685–698. [Google Scholar]
- Wright, S. Evolution and Genetics of Populations. In Volume 4: Variability within and among Natural Populations; University of Chicago Press: Chicago, IL, USA, 1984. [Google Scholar]
- Biancardi, E.; Panella, L.W.; Lewellen, R.T. Morphology, Physiology, and Ecology. In Beta Maritima: The Origin of Beets; Schmidt, T., Ed.; Springer: New York, NY, USA, 2012; pp. 109–117. [Google Scholar]
- Cureton, A.N.; Newbury, H.J.; Raybould, A.F.; Ford-Lloyd, B.V. Genetic structure and gene flow in wild beet populations: The potential influence of habitat on transgene spread and risk assessment. J. Appl. Ecol. 2006, 43, 1203–1212. [Google Scholar] [CrossRef]
- Lienert, J. Habitat fragmentation effects on fitness of plant populations—A review. J. Nat. Conserv. 2004, 12, 53–72. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.L.; Skolnick, M.; Davis, R. Construction of a genetic linkage map in man using restriction fragment lenght polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Frese, L.; Nachtigall, M.; Iriondo, J.M.; Rubio Teso, M.L.; Duarte, M.C.; Pinheiro de Carvalho, M.Â.A. Genetic diversity and differentiation in Patellifolia (Amaranthaceae) in the Macaronesian archipelagos and the Iberian Peninsula and implications for genetic conservation programmes. Genet. Resour. Crop Evol. 2019, 66, 225–241. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, S.E.J.D.; Briscoe, D.A.; Ballou, J.D. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Fikre, T.; Tesfaye, K.; Assefa, K. Genetic Diversity of Ethiopian Tef [(Eragrostis tef (Zucc.) Trotter] Released and Selected Farmers’ Varieties along with Two Wild Relatives as Revealed by Microsatellite Markers. J. Crop Sci. Biotechnol. 2018, 21, 367–374. [Google Scholar] [CrossRef]
- Mekbib, Y.; Huang, S.X.; Ngarega, B.K.; Li, Z.Z.; Shi, T.; Ou, K.F.; Liang, Y.T.; Chen, J.M.; Yang, X.Y. The level of genetic diversity and differentiation of tropical lotus, Nelumbo nucifera Gaertn. (Nelumbonaceae) from Australia, India, and Thailand. Bot. Stud. 2020, 61, 15. [Google Scholar] [CrossRef]
- He, Q.; Li, X.W.; Liang, G.L.; Ji, K.; Guo, Q.G.; Yuan, W.M.; Zhou, G.Z.; Chen, K.S.; van de Weg, W.E.; Gao, Z.S. Genetic Diversity and Identity of Chinese Loquat Cultivars/Accessions (Eriobotrya japonica) Using Apple SSR Markers. Plant Mol. Biol. Rep. 2011, 29, 197–208. [Google Scholar] [CrossRef]
- Baraket, G.; Chatti, K.; Saddoud, O.; Ben Abdelkarim, A.; Mars, M.; Trifi, M.; Hannachi, A.S. Comparative Assessment of SSR and AFLP Markers for Evaluation of Genetic Diversity and Conservation of Fig, Ficus carica L., Genetic Resources in Tunisia. Plant Mol. Biol. Rep. 2011, 29, 171–184. [Google Scholar] [CrossRef]
- Sharma, S.S.; Negi, M.S.; Sinha, P.; Kumar, K.; Tripathi, S.B. Assessment of Genetic Diversity of Biodiesel Species Pongamia pinnata Accessions using AFLP and Three Endonuclease-AFLP. Plant Mol. Biol. Rep. 2011, 29, 12–18. [Google Scholar] [CrossRef]
- Kong, Q.; Li, X.; Xiang, C.; Wang, H.; Song, J.; Zhi, H. Genetic Diversity of Radish (Raphanus sativus L.) Germplasm Resources Revealed by AFLP and RAPD Markers. Plant Mol. Biol. Rep. 2011, 29, 217–223. [Google Scholar] [CrossRef]
- Matthews, T.J.; Rigal, F.; Kougioumoutzis, K.; Trigas, P.; Triantis, K.A. Unravelling the small-island effect through phylogenetic community ecology. J. Biogeogr. 2020, 47, 2341–2352. [Google Scholar] [CrossRef]
- Ottewell, K.M.; Bickerton, D.C.; Byrne, M.; Lowe, A.J. Bridging the gap: A genetic assessment framework for population-level threatened plant conservation prioritization and decision-making. Divers. Distrib. 2016, 22, 174–188. [Google Scholar] [CrossRef]
- Jornal Oficial da Região Autónoma da Madeira. JORAM. Resolução nº 1226/2015 Aprova a alteração dos Limites dos Sítios Classificados da Rede Natura 2000 PTMAD0001—Laurissilva da Madeira; PTMAD0003—Ponta de São Lourenço; PTMAD0006—Moledos—Madalena do Mar e PTMAD0007. Available online: https://ifcn.madeira.gov.pt/images/Doc_Artigos/Legislacao/AreasProtegidas/pnm/altrn2000maddez2015.pdf (accessed on 1 June 2021).
- European Comission Natura 2000—The Macaronesian Region. Available online: https://ec.europa.eu/environment/nature/natura2000/biogeog_regions/macaronesian/index_en.htm (accessed on 1 June 2021).
- Maxted, N.; Labokas, J.; Palme, A. Crop wild relative conservation strategies. Planning and implementing national and regional conservation strategies. In Proceedings of the a Joint Nordic/ECPGR Workshop, Vilnius, Lithuania, 19–22 September 2016; pp. 19–22. [Google Scholar]
- Cruz, M.J.; Santos, F.D.; Aguiar, R.; Oliveira, R.P.; Correia, A.; Tavares, T.; Pereira, J.S. Impactos das alterações climáticas nos ecossistemas terrestres da ilha da madeira. In Proceedings of the I Congresso Nacional Sobre Alterações Climáticas, CLIMA 2008, Aveiro, Portugal, 29–30 September 2008. [Google Scholar]
- Maggioni, L.; Maxted, N.; Engels, J. The European Cooperative Programme For Plant Genetic Resources (ECPGR) And Planta Europa: An Opportunity For Synergies. Plant Europa. News 2008, winter, 10. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Type | Designations | Traits | References |
|---|---|---|---|
| SSR | 2KWS (SSR2) | Leaf and root K+, Leaf Ca2+, Root Na+, WSC, ECS; SC + WSC, ANC; saline and non-saline conditions. | Abbasi et al. [36] |
| SSR | BQ584037 | Phosphatidylglycerol-phosphate synthase. | McGrath et al. [37] |
| SSR | BQ588629 | BSD domain-containing protein (Pfam PF03909), root K+, WSY, WSC, SY, under saline conditions. | Abbasi et al. [36] |
| SSR | FDSB1027 | Tissue Na+ content, SY, WSY; saline responses. | Abbasi et al. [36] |
| EST-SSR | FDSB1250 | Hydrolase family protein (Pfam PF00657); GDSL esterase/lipase; WSC under non-saline conditions. | Abbasi et al. [36] |
| SSR | SB04 | Leaf Na+, Leaf K+, WSC under saline conditions. | - |
| SSR | SB13 | WSC, SC, root ANC under saline conditions. | Abbasi et al. [36] |
| SSR | SB15 | Sugar yield-related traits: SY, WSY, RY, WSC, ECS; saline responses. | Abbasi et al. [36] |
| Molecular Marker | Linkage Group | Alleles | PIC |
|---|---|---|---|
| 2KWS (SSR2) | 2 | 11 | 0.870 |
| BQ584037 | 2 | 10 | 0.858 |
| BQ588629 | 1 | 9 | 0.662 |
| FDSB1027 | 3 | 8 | 0.670 |
| FDSB1250 | 7 | 5 | 0.406 |
| SB04 | 5 | 8 | 0.739 |
| SB13 | 3 | 6 | 0.605 |
| SB15 | 5 | 17 | 0.734 |
| Desembarcadouro Islet (DI) | Chão Islet (CI) | |||||||
|---|---|---|---|---|---|---|---|---|
| Marker | H′ | Ho | Nei | FIS | H′ | Ho | Nei | FIS |
| 2KWS | 2.0803 | 0.1048 | 0.8601 | 0.8782 | 1.6566 | 0.1333 | 0.7911 | 0.8315 |
| BQ584037 | 2.0976 | 0.7238 | 0.8591 | 0.1575 | 1.7458 | 0.6667 | 0.8044 | 0.1713 |
| BQ588629 | 1.2872 | 0.1524 | 0.6079 | 0.7493 | 0.4851 | 0.1333 | 0.2400 | 0.4444 |
| FDSB1027 | 1.3882 | 0.0190 | 0.6716 | 0.9716 | 1.0776 | 0.0000 | 0.5778 | 1.0000 |
| FDSB1250 | 0.7821 | 0.0381 | 0.4067 | 0.9063 | 0.4851 | 0.0000 | 0.2400 | 1.0000 |
| SB04 | 1.5791 | 0.0857 | 0.7264 | 0.8820 | 1.2382 | 0.1333 | 0.6867 | 0.8058 |
| SB13 | 1.0695 | 0.0857 | 0.5723 | 0.8502 | 1.4154 | 0.1333 | 0.7289 | 0.8171 |
| SB15 | 1.7787 | 0.1524 | 0.7227 | 0.7891 | 1.1711 | 0.1333 | 0.6489 | 0.7945 |
| Mean | 1.5078 | 0.1702 | 0.6784 | 1.1593 | 0.1667 | 0.5897 | ||
| St. Dev. | 0.4684 | 0.2287 | 0.1512 | 0.4749 | 0.2108 | 0.2279 | ||
| Source of Variation | df | SS | MS | Variance Estimated | Percentage of Variance (%) | F-Statistics | p |
|---|---|---|---|---|---|---|---|
| Between populations | 1 | 19.493 | 19.493 | 0.281 | 9 | FST = 0.094 | <0.001 |
| Among individuals | 118 | 559.086 | 4.738 | 2.029 | 68 | FIS = 0.749 | <0.001 |
| Within individuals | 120 | 81.500 | 0.679 | 0.679 | 23 | FIT = 0.773 | <0.001 |
| Total | 239 | 660.079 | - | 2.990 | 100 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ragonezi, C.; Nóbrega, H.; Leite, M.I.; Freitas, J.G.R.d.; Macedo, F.L.; Carvalho, M.Â.A.P.d. Distribution of Genetic Diversity in Beta patula Aiton Populations from Madeira Archipelago, Portugal. Agriculture 2023, 13, 27. https://doi.org/10.3390/agriculture13010027
Ragonezi C, Nóbrega H, Leite MI, Freitas JGRd, Macedo FL, Carvalho MÂAPd. Distribution of Genetic Diversity in Beta patula Aiton Populations from Madeira Archipelago, Portugal. Agriculture. 2023; 13(1):27. https://doi.org/10.3390/agriculture13010027
Chicago/Turabian StyleRagonezi, Carla, Humberto Nóbrega, Maria Inês Leite, José G. R. de Freitas, Fabrício Lopes Macedo, and Miguel Â. A. Pinheiro de Carvalho. 2023. "Distribution of Genetic Diversity in Beta patula Aiton Populations from Madeira Archipelago, Portugal" Agriculture 13, no. 1: 27. https://doi.org/10.3390/agriculture13010027