
Identification and Verification of qGS11, a QTL Controlling Grain Size and Heading Date in Rice

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Field Trials and Phenotypic Evaluation
2.3. DNA Marker Analysis
2.4. Data Analysis
3. Results
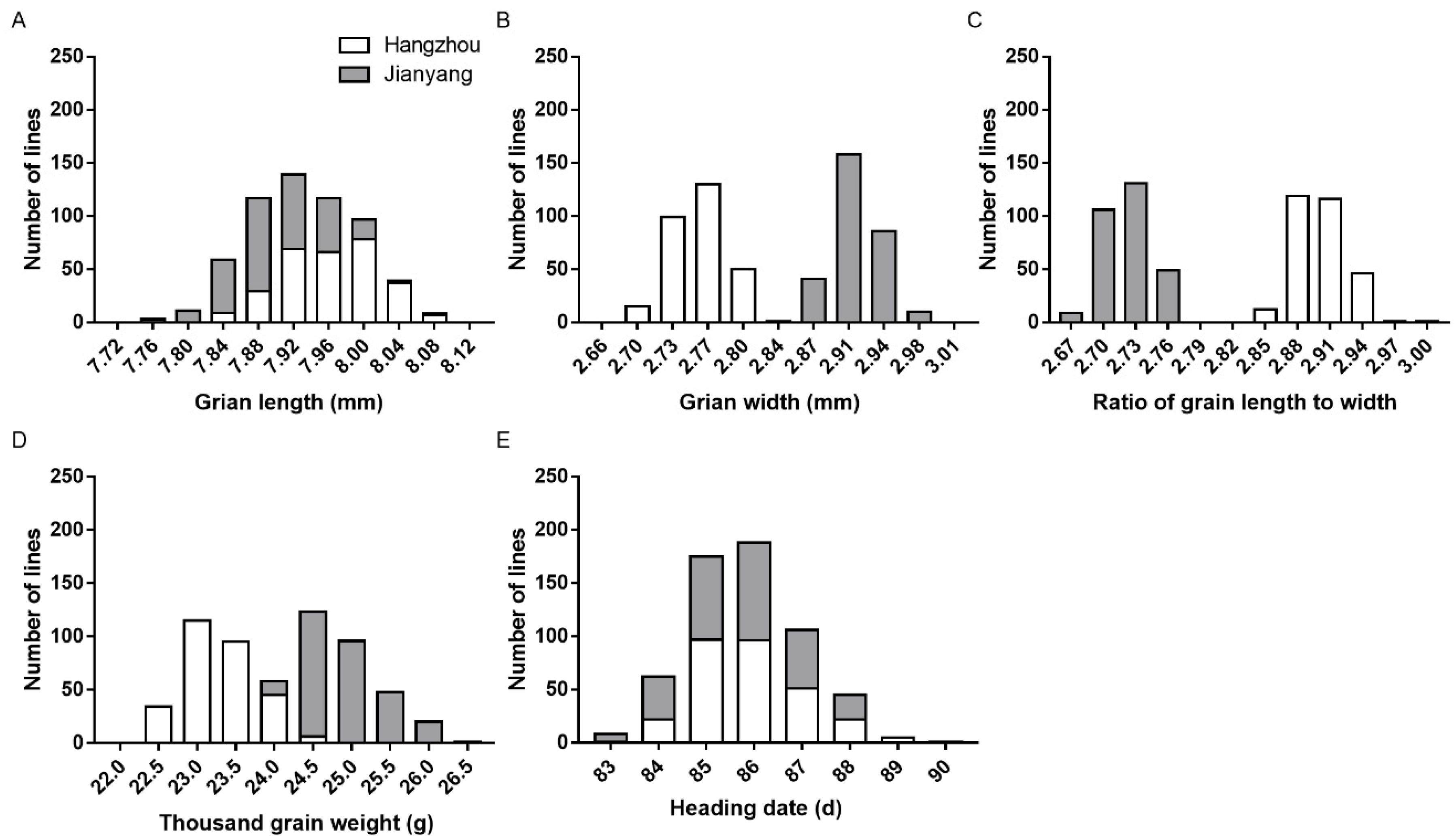
3.1. Phenotypic Performance of the ZL1 and ZL2 Populations
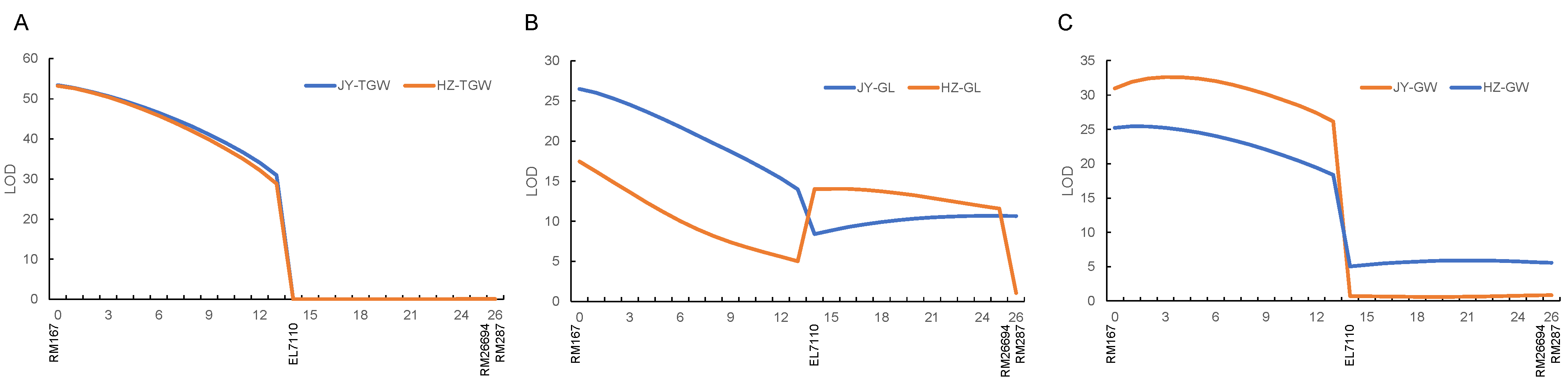
3.2. QTLs Detected in the ZL1 Population
3.3. QTLs Detected in the ZL2 Population
3.4. Validation of qGS11 Using NILs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zuo, Z.-W.; Zhang, Z.-H.; Huang, D.-R.; Fan, Y.-Y.; Yu, S.-B.; Zhuang, J.-Y.; Zhu, Y.-J. Control of thousand-grain weight by OsMADS56 in rice. Int. J. Mol. Sci. 2022, 23, 125. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lu, L.; Bai, X.; Zhou, X.; Wu, B.; Yuan, M.; Yang, L.; Xing, Y. A minor QTL, SG3, encoding an R2R3-MYB protein, negatively controls grain length in rice. Theor. Appl. Genet. 2020, 133, 2387–2399. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.N.; Wang, L.-L.; Zhu, Y.-J.; Fan, Y.-Y.; Zhuang, J.-Y.; Zhang, Z.-H. Identification through fine mapping and verification using CRISPR/Cas9-targeted mutagenesis for a minor QTL controlling grain weight in rice. Theor. Appl. Genet. 2021, 134, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Silvio, S.; Tuberosa, R. To clone or not to clone plant QTLs: Present and future challenges. Trends Plant Sci. 2005, 10, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Endo-Higashi, N.; Izawa, T. Flowering time genes Heading date 1 and Early heading date 1 together control panicle development in rice. Plant Cell Physiol. 2011, 52, 1083–1094. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhang, Z.-h.; Zhuang, J.-y. Quantitative Trait loci for heading date and their relationship with genetic control of yield traits in rice (Oryza sativa). Rice Sci. 2013, 20, 1–12. (In Chinese) [Google Scholar] [CrossRef]
- Xue, W.; Xing, Y.; Weng, X.; Zhao, Y.; Tang, W.; Wang, L.; Zhou, H.; Yu, S.; Xu, C.; Li, X.; et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat. Genet. 2008, 40, 761–767. [Google Scholar] [CrossRef]
- Yan, W.; Liu, H.; Zhou, X.; Li, Q.; Zhang, J.; Lu, L.; Liu, T.; Liu, H.; Zhang, C.; Zhang, Z.; et al. Natural variation in Ghd7.1 plays an important role in grain yield and adaptation in rice. Cell Res. 2013, 23, 969–971. [Google Scholar] [CrossRef]
- Wei, X.; Xu, J.; Guo, H.; Jiang, L.; Chen, S.; Yu, C.; Zhou, Z.; Hu, P.; Zhai, H.; Wan, J. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010, 153, 1747–1758. [Google Scholar] [CrossRef]
- Zhang, Z.-H.; Wang, K.; Guo, L.; Zhu, Y.-J.; Fan, Y.-Y.; Cheng, S.-H.; Zhuang, J.-Y. Pleiotropism of the photoperiod-insensitive allele of Hd1 on heading date, plant height and yield traits in rice. PLoS ONE 2012, 7, e52538. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.-H.; Cao, L.-Y.; Chen, J.-Y.; Zhang, Y.-X.; Zhuang, J.-Y.; Cheng, S.-H. Effects of Hd2 in the presence of the photoperiod-insensitive functional allele of Hd1 in rice. Biol. Open 2016, 5, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-J.; Fan, Y.-Y.; Wang, K.; Huang, D.-R.; Liu, W.-Z.; Ying, J.-Z.; Zhuang, J.-Y. Rice flowering locus T1 plays an important role in heading date influencing yield traits in rice. Sci. Rep. 2017, 7, 4918. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, Y.-J.; Zhu, A.-D.; Fan, Y.-Y.; Huang, T.-X.; Zhang, J.-F.; Xie, H.-A.; Zhuang, J.-Y. Identification and verification of quantitative trait loci affecting milling yield of rice. Agronomy 2020, 10, 75. [Google Scholar] [CrossRef]
- Zhu, A.-D. Mapping Minor-Effect QTL for Grain Weight and Shape and Validation of Five QTL in Rice. Master’s Degree, Chinese Academy of Agricultural Sciences, Hangzhou, China, 2019.
- Zhang, H.-W.; Fan, Y.-Y.; Zhu, Y.-J.; Chen, J.-Y.; Yu, S.-B.; Zhuang, J.-Y. Dissection of the qTGW1.1 region into two tightly-linked minor QTLs having stable effects for grain weight in rice. BMC Genet. 2016, 17, 98. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Huang, N.; Bennett, J.; Khush, G.S. PCR-Based Marker-Assisted Selection in Rice Breeding; IRRI Discussion Paper Series No.12; International Rice Research Institute: Los Baños, Philippines, 1995. [Google Scholar]
- Chen, X.; Temnykh, S.; Xu, Y.; Cho, Y.G.; McCouch, S.R. Development of a microsatellite framework map providing genome-wide coverage in rice (Oryza sativa L.). Theor. Appl. Genet. 1997, 95, 553–567. [Google Scholar] [CrossRef]
- Meng, L.; Li, H.; Zhang, L.; Wang, J. QTL Icimapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 2015, 3, 269–283. [Google Scholar] [CrossRef]
- McCouch, S.R.; CGSNL. Committee on Gene Symbolization, Nomenclature and Linkage, Rice Genetic Cooperative. In Gene Nomenclature System for Rice; Springer: Berlin, Germany, 2008; Volume 1, pp. 72–84. [Google Scholar]
- Dai, W.-M.; Zhang, K.-Q.; Wu, J.-R.; Wang, L.; Duan, B.-W.; Zheng, K.-L.; Cai, R.; Zhuang, J.-Y. Validating a segment on the short arm of chromosome 6 responsible for genetic variation in the hull silicon content and yield traits of rice. Euphytica 2008, 160, 317–324. [Google Scholar] [CrossRef]
- Mauricio, R. Mapping quantitative trait loci in plants: Uses and caveats for evolutionary biology. Nat. Rev. Genet. 2001, 2, 370–381. [Google Scholar] [CrossRef]
- Gu, H.; Zhang, K.; Gull, S.; Chen, C.; Ran, J.; Zou, B.; Wang, P.; Liang, G. Fine mapping of qTGW7b, a minor effect QTL for grain weight in rice. Int. J. Mol. Sci. 2022, 23, 8926. [Google Scholar] [CrossRef]
- Bao, J. Genes and QTLs for rice grain quality improvement. In Rice-Germplasm, Genetics and Improvement; Yan, W., Bao, J., Eds.; Intech: Rijeka, Croatia, 2014; pp. 239–278. [Google Scholar]
- Mei, D.-Y.; Zhu, Y.-J.; Yu, Y.-H.; Fan, Y.-Y.; Huang, D.-R.; Zhuang, J.-Y. Quantitative trait loci for grain chalkiness and endosperm transparency detected in three recombinant inbred line populations of indica rice. J. Integr. Agric. 2013, 12, 1–11. [Google Scholar] [CrossRef]
- Nakagawa, M.; Shimamoto, K.; Kyozuka, J. Overexpression of RCN1 and RCN2, rice TERMINAL FLOWER1/CENTRORADIALIS homologs, confers delay of phase transition and altered panicle morphology in rice. Plant J. 2002, 29, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhu, S.S.; Liu, T.Z.; Wang, C.M.; Cheng, Z.J.; Zhang, X.; Chen, L.P.; Sheng, P.K.; Cai, M.H.; Li, C.N.; et al. DELAYED HEADING DATE1 interacts with OsHAP5C/D, delays flowering time and enhances yield in rice. Plant Biotecnol. J. 2019, 17, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Han, S.-H.; Yoo, S.-C.; Lee, B.-D.; An, G.; Peak, N.-C. Rice FLAVIN-BIDING, KELCH REPEAT, F-BOX1 (OsFKF1) promotes flowering independent of photoperiod. Plant Cell Environ. 2015, 38, 2527–2540. [Google Scholar] [CrossRef]
- Saito, H.; Yuan, Q.; Okumoto, Y.; Doi, K.; Yoshimura, A.; Inoue, H.; Teraishi, M.; Tsukiyama, T.; Tanisaka, T. Multiple alleles at Early flowering 1 locus making variation in the basic vegetative growth period in rice (Oryza sativa L.). Theor. Appl. Genet. 2009, 119, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Saito, H.; Okumoto, Y.; Inoue, H.; Nishida, H.; Tsukiyama, T.; Teraishi, M.; Tanisaka, T. Identification of a novel gene ef7 conferring an extremely long basic vegetative growth phase in rice. Theor. Appl. Genet. 2009, 119, 675–684. [Google Scholar] [CrossRef]
- Gao, H.; Jin, M.; Zheng, X.; Chen, J.; Yuan, D.; Xin, Y.; Wang, M.; Huang, D.; Zhang, Z.; Zhou, K.; et al. Days to heading 7, a major quantitative locus determining photoperiod sensitivity and regional adaptation in rice. Proc. Natl. Acad. Sci. USA 2014, 111, 16337–16342. [Google Scholar] [CrossRef]
- Wei, H.-H.; Li, C.; Xing, Z.-P.; Wang, W.-T.; Dai, Q.-G.; Zhou, G.-S.; Wang, L.; Xu, K.; Huo, Z.-Y.; Guo, B.-W.; et al. Suitable growing zone and yield potential for late-maturity type of Yongyou japonica/indica hybrid rice in the lower reaches of Yangtze River. J. Integr. Agric. 2015, 1, 13. [Google Scholar] [CrossRef]
- Wei, X.; Liu, L.; Xu, J.; Jiang, L.; Zhang, W.; Wang, J.; Zhai, H.; Wan, J. Breeding strategies for optimum heading date using genotypic information in rice. Mol. Breed. 2010, 25, 287–298. [Google Scholar] [CrossRef]
- Fujino, K.; Yamanouchi, U.; Yano, M. Roles of the Hd5 gene controlling heading date for adaptation to the northern limits of rice cultivation. Theor. Appl. Genet. 2013, 126, 611–618. [Google Scholar] [CrossRef]
- Naranjo, L.; Talón, M.; Domingo, C. Diversity of floral regulatory genes of japonica rice cultivated at northern latitudes. BMC Genom. 2014, 15, 101. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Population | Trait | Location | Mean | SD | CV | Range | Skewness | Kurtosis |
|---|---|---|---|---|---|---|---|---|
| ZL1 | GL | Hangzhou | 7.962 | 0.054 | 0.68 | 7.830−8.111 | −0.08 | −0.51 |
| Jianyang | 7.902 | 0.053 | 0.67 | 7.753−8.073 | 0.13 | −0.10 | ||
| GW | Hangzhou | 2.757 | 0.026 | 0.94 | 2.692−2.819 | 0.09 | −0.39 | |
| Jianyang | 2.913 | 0.022 | 0.76 | 2.870−2.980 | 0.31 | −0.30 | ||
| RLW | Hangzhou | 2.900 | 0.023 | 0.79 | 2.846−2.997 | 0.36 | 0.36 | |
| Jianyang | 2.722 | 0.021 | 0.77 | 2.670−2.768 | 0.04 | −0.59 | ||
| TGW | Hangzhou | 23.27 | 0.47 | 2.02 | 22.29−24.45 | 0.26 | −0.61 | |
| Jianyang | 24.89 | 0.47 | 1.89 | 23.93−26.47 | 0.50 | −0.24 | ||
| HD | Hangzhou | 85.8 | 1.1 | 1.28 | 83.1−89.5 | 0.54 | 0.22 | |
| Jianyang | 85.7 | 1.2 | 1.40 | 83.0−88.2 | 0.09 | −0.57 | ||
| ZL2 | GL | Hangzhou | 8.043 | 0.046 | 0.57 | 7.906–8.167 | 0.01 | −0.33 |
| GW | Hangzhou | 2.712 | 0.026 | 0.96 | 2.627–2.789 | −0.12 | 0.10 | |
| RLW | Hangzhou | 2.978 | 0.022 | 0.74 | 2.912–3.062 | 0.46 | 0.76 | |
| TGW | Hangzhou | 23.07 | 0.38 | 1.65 | 21.83–24.15 | −0.25 | 0.19 | |
| HD | Hangzhou | 84.9 | 1.1 | 1.29 | 80.8–89.6 | 0.21 | 0.89 |
| Traits | GL | GW | RLW | TGW | ||||
|---|---|---|---|---|---|---|---|---|
| Hangzhou | Jianyang | Hangzhou | Jianyang | Hangzhou | Jianyang | Hangzhou | Jianyang | |
| GW | 0.576 *** | 0.434 *** | ||||||
| RLW | 0.175 *** | 0.470 *** | −0.703 *** | −0.590 *** | ||||
| TGW | 0.766 *** | 0.616 *** | 0.896 *** | 0.817 *** | −0.406 *** | −0.244 *** | ||
| HD | −0.194 *** | 0.108 | −0.504 *** | −0.058 | 0.442 *** | 0.155 ** | −0.468 *** | −0.180 ** |
| QTL | Interval | Hangzhou | Jianyang | ||||||
|---|---|---|---|---|---|---|---|---|---|
| LOD | A | D | R2 (%) | LOD | A | D | R2 (%) | ||
| qGL11.1 | RM167—EL7110 | 17.4 | 0.029 | 0.013 | 18.3 | 26.5 | 0.039 | 0.006 | 16.5 |
| qGL11.2 | EL7110—RM26694 | 14.1 | −0.028 | −0.008 | 13.7 | 10.7 | −0.025 | −0.009 | 6.1 |
| qGW11 | RM167—EL7110 | 25.4 | 0.016 | −0.001 | 45.5 | 32.6 | 0.017 | 0.0004 | 40.2 |
| qRLW11 | EL7110—RM26694 | 24.1 | −0.016 | −0.007 | 32.3 | 13.2 | −0.011 | −0.005 | 20.3 |
| qTGW11 | RM167—EL7110 | 53.2 | 0.41 | 0.02 | 56.6 | 53.4 | 0.41 | 0.01 | 56.7 |
| qHD11 | EL7110—RM26694 | 14.2 | −0.6 | −0.2 | 20.4 | 9.8 | −0.6 | −0.2 | 14.5 |
| QTL | Interval | LOD | A | D | R2 (%) |
|---|---|---|---|---|---|
| qGW11 | RM167–EL7110 | 3.7 | 0.006 | −0.009 | 5.6 |
| qRLW11 | EL7110–RM26694 | 4.1 | −0.006 | 0.007 | 6.9 |
| qTGW11 | RM167–EL7110 | 5.6 | 0.14 | −0.02 | 9.1 |
| qHD11 | RM167–EL7110 | 7.8 | −0.5 | 0.2 | 11.3 |
| Trait | Phenotypic (Mean ± SD) | p | A | R2 (%) | |
|---|---|---|---|---|---|
| NILTQ | NILIR | ||||
| GL | 7.658 ± 0.046 | 7.696 ± 0.051 | <0.0001 | 0.019 | 9.5 |
| GW | 3.006 ± 0.014 | 3.049 ± 0.019 | <0.0001 | 0.022 | 56.3 |
| RLW | 2.548 ± 0.020 | 2.524 ± 0.024 | <0.0001 | −0.012 | 20.3 |
| TGW | 23.96 ± 0.24 | 24.9 ± 0.34 | <0.0001 | 0.47 | 59.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, C.-L.; Wang, S.-L.; Fan, Y.-Y.; Huang, T.-X.; Zhuang, J.-Y.; Zhu, Y.-J.; Zhang, H. Identification and Verification of qGS11, a QTL Controlling Grain Size and Heading Date in Rice. Agriculture 2022, 12, 1384. https://doi.org/10.3390/agriculture12091384
Zheng C-L, Wang S-L, Fan Y-Y, Huang T-X, Zhuang J-Y, Zhu Y-J, Zhang H. Identification and Verification of qGS11, a QTL Controlling Grain Size and Heading Date in Rice. Agriculture. 2022; 12(9):1384. https://doi.org/10.3390/agriculture12091384
Chicago/Turabian StyleZheng, Chang-Lin, Shi-Lin Wang, Ye-Yang Fan, Ting-Xu Huang, Jie-Yun Zhuang, Yu-Jun Zhu, and Hui Zhang. 2022. "Identification and Verification of qGS11, a QTL Controlling Grain Size and Heading Date in Rice" Agriculture 12, no. 9: 1384. https://doi.org/10.3390/agriculture12091384